
 , Michal Ďuračka 2, Marek Halenár 3, Juraj Pivko 4, Eduard Kolesár 3, Ľubica Chrastinová 4, Ľubomír Ondruška 4, Rastislav Jurčík 4, Adriana Kolesárová 2,3
, Michal Ďuračka 2, Marek Halenár 3, Juraj Pivko 4, Eduard Kolesár 3, Ľubica Chrastinová 4, Ľubomír Ondruška 4, Rastislav Jurčík 4, Adriana Kolesárová 2,31 Institute of Biotechnology, Slovak University of Agriculture in Nitra, 949 76 Nitra, Slovakia
2 AgroBioTech Research Centre, Slovak University of Agriculture in Nitra, 949 76 Nitra, Slovakia
3 Institute of Applied Biology, Slovak University of Agriculture in Nitra, 949 76 Nitra, Slovakia
4 Animal Production Research Centre Nitra, National Agricultural and Food Center, 951 41 Lužianky, Slovakia
Abstract
Background: Apricot kernels containing amygdalin (AMG) as the major cyanogenic glycoside are potentially useful as a complementary therapy for the management of several ailments including cancer. Nevertheless, little is known regarding the toxic and therapeutic doses of AMG, particularly in terms of male reproduction. Hence, this study evaluates selected qualitative characteristics of rabbit testicular tissue following in vivo administration of AMG or apricot kernels for 28 days. Methods: The rabbits were randomly divided into five groups (Control, P1, P2, P3, P4). The Control received no AMG/apricot kernels while the experimental groups P1 and P2 received a daily intramuscular injection of amygdalin at a dose of 0.6 and 3.0 mg/kg of body weight (b.w.) for 28 days, respectively. P3 and P4 received a daily dose of 60 and 300 mg/kg b.w. of crushed apricot kernels mixed with feed for 28 days, respectively. Changes to the testicular structure were quantified morphometrically, while tissue lysates were subjected to the evaluation of reactive oxygen species (ROS) production, total antioxidant capacity, activities of antioxidant enzymes, and glutathione concentration. The extent of damage to the proteins and lipids was quantified as well. Levels of selected cytokines were determined by the enzyme-linked immunosorbent assay while a luminometric approach was used to assess the activity of caspases. Results: Rabbits treated with 3.0 mg/kg b.w. AMG presented a significantly increased protein oxidation (p = 0.0118) accompanied by a depletion of superoxide dismutase (p = 0.0464), catalase (p = 0.0317), and glutathione peroxidase (p = 0.0002). Significantly increased levels of interleukin-1 beta (p = 0.0012), tumor necrosis factors alpha (p = 0.0159), caspase-3/7 (p = 0.0014), and caspase-9 (p = 0.0243) were also recorded in the experimental group P2 when compared to the Control. No effects were observed in the rabbits treated with apricot kernels at the oxidative, inflammatory, and histopathological levels. Conclusions: Apricot kernels did not induce toxicity in the testicular tissues of male rabbits, unlike pure AMG, which had a negative effect on male reproductive structures carried out through oxidative, inflammatory, and pro-apoptotic mechanisms.
Keywords
- amygdalin
- apricot kernels
- rabbits
- testes
- histology
- oxidative stress
- inflammation
- caspases
According to global statistics, cancer is the second major cause of death worldwide [1] representing a global public health concern with an increasing tendency, mostly due to population growth and a fast-aging population, particularly in developed countries [2]. The most common malignancies diagnosed in male patients comprise prostate, colorectal, lung, and bladder cancer; while breast, colorectal, uterine, and lung cancer are the most common in female patients [1]. Since cancer patients are met with numerous physical, emotional, and socio-economic challenges that have a major impact on their quality of life, major efforts have been invested in the development of faster and more efficient screening, diagnostic, and treatment strategies [3]. Despite remarkable progress in the understanding of cancer development that is accompanied by major advances in cancer therapy over the past decade, the fact that malignancies are generally aggressive, heterogenous, and dynamic pathologies [4] represents a persistent challenge for clinical oncology even in the new millennium [4].
The World Health Organization (WHO) defines cancer therapy as a treatment aimed to considerably prolong the life of oncology patients and to ensure a proper quality of life for cancer survivors [5]. While the leading cancer therapies comprise conventional options such as surgery, radiation therapy, chemotherapy, and proton therapy [2], complementary therapies have received more prominence and popularity as adjuvant remedies to improve the efficiency of conservative cancer treatment as well as a strategy to increase the quality of life of patients [6]. Although complementary remedies are by and large not included in standard patient care, particularly in developed countries as opposed to most African or Asian countries [7, 8, 9], their use is on the rise, most notably to enhance immunity, relieve pain, or ameliorate side-effects stemming either from the ailment itself or from conservative treatment [8]. Complementary therapies encompass a rather large and diverse group of approaches, that may be divided into three major categories, specifically mind–body therapies (such as meditation, hypnosis, acupuncture, relaxation techniques, yoga, tai chi, or massage therapy), traditional medicine (Chinese medicine, Ayurveda, or naturopathic medicine), and biologically based practices or natural products such as vitamins, botanicals, dietary supplements, herbs, spices, special foods, or diets [9]. Among these, apricot kernels, and their active component amygdalin, have received particular attention from the public as well as the scientific community.
Amygdalin (D-mandelonitrile-b-D-gentiobioside; AMG) is a cyanogenic glycoside
composed of two molecules of glucose: one molecule of benzaldehyde, which has
analgesic effects; and one molecule of hydrocyanic acid, which is regarded as an
antineoplastic agent [10]. The compound may be found in kernels of plants
belonging to the Rosaceae family such as almonds, apricots, peaches, or
apples [11]. AMG per se is non-toxic until hydrogen cyanide (HCN) is released
primarily through enzymatic hydrolysis catalyzed by
AMG has become a popular alternative treatment in patients suffering from diabetes mellitus, asthma, bronchitis, and a complementary cancer remedy [10]. The use of AMG or its synthetic derivate Laetrile in cancer therapy has been widely supported by in vitro studies on colon [12], cervical [14], bladder [15], or prostate cancer cells [16]. The rationale for using this molecule rests on the fact that as opposed to cancer cells, non-malignant cells present with higher levels of rhodanese—a liver mitochondrial enzyme that catalyzes the detoxification of HCN by sulphuration [17]. Nevertheless, the effects of AMG observed in in vivo studies are inconsistent; besides, peroral ingestion of AMG comes along with the risk of cyanide poisoning. Consequently, AMG has not yet been granted Food and Drug Administration (FDA) approval for its use as a therapeutic agent in the United States due to insufficient clinical verification of its therapeutic efficiency [18]. As such, particular attention must be paid to any effects AMG may exhibit on healthy individuals, since the current knowledge on the behavior of AMG on the general morphology and physiology is limited.
This paper is part of a larger investigation striving to determine the effects of AMG within a larger systemic context of health, physiology, and reproduction [19, 20, 21, 22, 23]. As opposed to most studies on AMG, taking advantage of rodents, we chose rabbits as our laboratory model. Whilst undoubtedly the use of mice or rats comes along reduced maintenance costs, small size, and ease of breeding as well as a wide array of available commercial reagents, and genetically modified models [24], the attractiveness of rabbits lies in their intermediate size between rodents and larger animal models, a longer life span, and higher success rate in the translation of findings to humans. From a practical point of view, the size of rabbits permits greater access to tissues and cells from one single animal, and a repeated sampling of blood. What is more, semen collection is more convenient, and as opposed to rodents, ejaculated spermatozoa may be readily obtained when needed [24, 25].
Possible beneficial or adverse effects of AMG on male reproduction have been
previously studied with regard to the endocrine balance as well as the quality of
ejaculated spermatozoa. Kolesár [26] reported no significant
changes to the levels of pituitary hormones following a 28-day administration of
pure AMG or apricot kernels. Similarly, the levels of 17
Pure AMG as well as chemicals necessary for the sample processing for histology and oxidative profile assessment were purchased from Sigma-Aldrich (St. Louis, MO, USA), unless stated otherwise. Crushed apricot kernels for the experimental design were obtained from Trasco (Žiar n. Hronom, Slovakia). Commercial kits used for the cytokine analysis were obtained from MyBioSource, Inc. (San Diego, CA, USA), while caspase activity was assessed using commercial kits purchased from Promega Corporation (Madison, WI, USA).
Meat line P91 Californian rabbit males (n = 20; 150 days old; 4.00
The animals were randomly divided into five groups. The Control group received
neither pure AMG nor apricot kernels. Rabbits from the experimental groups P1 and
P2 received a daily intramuscular (i.m.) injection of pure AMG (
Left testes were fixed in 10% neutral buffered formalin (Sigma-Aldrich, St.
Louis, MO, USA), dehydrated with ethanol (70% and 96% for 2 h each; 100% for 1
h) (Centralchem, Bratislava, Slovakia), and embedded with the Technovit 7100
cuttable embedding resin (Kulzer GmbH, Hanau, Germany). The samples were
processed with the AC-820 rotary microtome (American Optical Company, Vernon
Hills, IL, USA), stained with toluidine blue-basic fuchsin stain, and mounted
with Entellan onto microscopic slides. The slides were evaluated for
histopathological changes under the EC3 optical microscope (Leica Camera AG,
Wetzlar, Germany) [19]. Photomicrographs were then taken at a magnification of
10
Right testes were cut into 50 mg fragments that were homogenized using two
different protocols depending on the final analysis. For the evaluation of the
oxidative profile and cytokine levels, the fragments were homogenized with the
Radioimmunoprecipitation assay (RIPA) buffer/protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MO, USA) using the SFX 250 ultrasonic homogenizer
(Branson Ultrasonics, Brookfield, CT, USA) on ice at 28 kHz for 30 s. Following
centrifugation (11,828
Global ROS levels in each specimen were quantified with the luminol-based
chemiluminescent assay. Each sample was treated with 5 mmol/L luminol
(5-amino-2,3-dihydro-1,4-phthalazinedione) and subjected to chemiluminescent
assessment using the Glomax Multi
The total antioxidant capacity (TAC) representing a sum of all ROS-quenching
molecules was assessed with an improved chemiluminescent assay developed by
Muller et al. [30]. All samples were treated with a signal reagent
containing 282.2 mmol/L luminol, 41.8 mmol/L 4-iodophenol, 12 mol/L
H
Levels of protein carbonyls (PCs) representing oxidative products of proteins
were quantified with the help of the 2,4-dinitrophenylhydrazine (DNPH) method
introduced by Weber et al. [31]. Briefly, each sample normalized to 1 mg
protein/mL was pre-treated with trichloroacetic acid (TCA; 20% w/v; Sigma-Aldrich, St. Louis, MO, USA),
subsequently mixed with 1 mL DNPH (10 mM in 2 N hydrochloric acid (HCl)) and
incubated at 37 °C for 1 h. After that, 20% TCA was added to the
samples, which were then centrifuged (300
The degree of lipid peroxidation (LPO) to the testicular tissue, expressed
through malondialdehyde (MDA) production, was quantified with the thiobarbituric
acid reactive substance (TBARS) assay. Briefly, each sample was pretreated with
5% sodium dodecyl sulfate, and then mixed with 0.53% thiobarbituric acid
dissolved in 20% acetic acid (Centralchem, Bratislava, Slovakia). The samples
were subsequently boiled (100 °C, 1 h), then cooled down and centrifuged
at 1750
Activities of the antioxidant enzymes superoxide dismutase (SOD) and glutathione
peroxidase (GPx) were assessed with the RANSOD (#SD125/SD126) and RANSEL
(#RS504/RS505) commercial kits (Randox Laboratories, Crumlin, UK) and the Rx
Monza semi-automatic analyzer. The results are expressed as IU/g protein [28]. In
the meantime, catalase (CAT) activity was assessed according to the protocol
designed by Beers and Sizer [32] that follows H
Levels of tumor necrosis factor-alpha (TNF-
The activity of caspase-3/7 and -9 was evaluated using the
Caspase-Glo® 3/7 Assay (#G8090) and
Caspase-Glo® 9 Assay (G8210), respectively. Prior to the
analysis, the protein concentration in each lysate was adjusted to 1 mg/mL. An
equal volume of diluted (10 µg/mL) lysate was mixed with the appropriate
Caspase-Glo® reagent in 96-well, white-walled plates. The
plates were then incubated at room temperature for 1 h and subsequently, the
luminescent signal was captured using the Glomax Multi
General statistics and comparisons were performed using the GraphPad Prism
program (version 8.4.4 for Mac; GraphPad Software Incorporated, La Jolla, CA,
USA). The measured values were expressed as mean
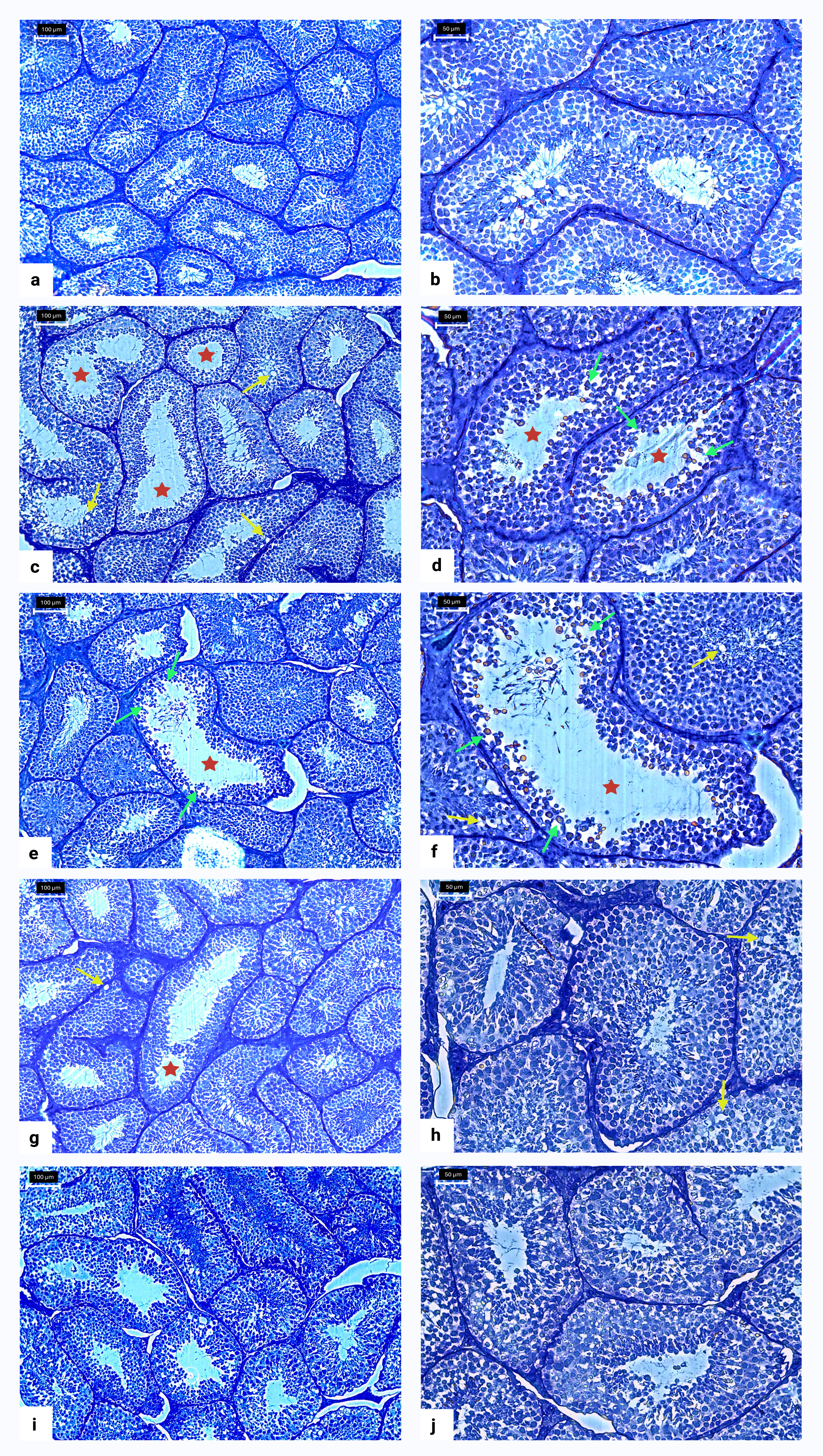
The results of the histopathological analysis of the Control and experimental samples are provided in Table 1 summarizing the observations made from all histological sections (n = 20) and slides (n = 60). Dystrophic changes observed in the seminiferous epithelium were most often manifested as hydropic-vacuolar dystrophy. Smaller or bigger vacuoles containing liquid were present in the seminiferous epithelium. Liquid was also found in the lumen and interstitium. The intensity of such changes was different, and slightly increased in the experimental groups when compared to the Control. Changes found in the Leydig cells of the experimental animals were comparable to the Control. Representative photomicrographs of the testicular tissue collected from the Control and experimental groups are depicted in Fig. 1.
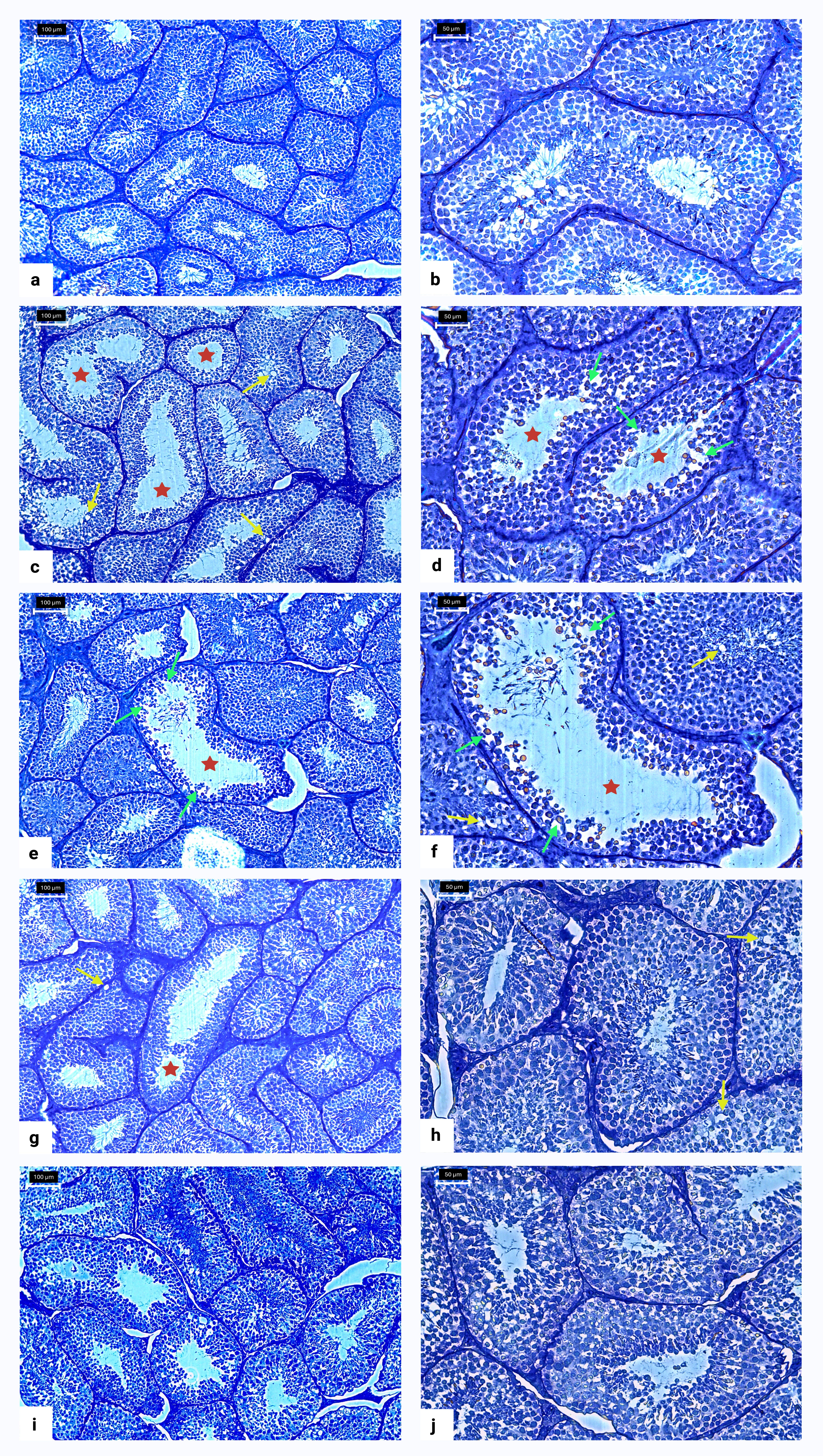 Fig. 1.
Fig. 1.Representative photomicrographs of testicular tissue, light microscopy. (a) Control animals; Scale bar = 100 μm. (b) Control animals; Scale bar = 50 μm. (c) Experimental animals administered with 0.6 mg/kg b.w. AMG i.m.; Scale bar = 100 μm. (d) Experimental animals administered with 0.6 mg/kg b.w. AMG i.m.; Scale bar = 50 μm. (e) Experimental animals administered with 3.0 mg/kg b.w. AMG i.m.; Scale bar = 100 μm. (f) Experimental animals administered with 3.0 mg/kg b.w. AMG i.m.; Scale bar = 50 μm. (g) Experimental animals administered with 60 mg/kg b.w. crushed apricot kernels orally; Scale bar = 100 μm. (h) Experimental animals administered with 60 mg/kg b.w. crushed apricot kernels orally; Scale bar = 50 μm. (i) Experimental animals administered with 300 mg/kg b.w. crushed apricot kernels orally; Scale bar = 100 μm. (j) Experimental animals administered with 300 mg/kg b.w. crushed apricot kernels orally; Scale bar = 50 μm. Seminiferous tubules of the Control group presented with unimpaired basal membranes, undisrupted spermatogenic line, and lumen filled with spermatozoa. Testicular tissue of the experimental groups intramuscularly treated with pure AMG contained smaller or bigger vacuoles filled with liquid (yellow arrow), and a disruption of the seminiferous epithelium was detected in some seminiferous tubules (green arrow). The lumen of several seminiferous tubules lacked spermatozoa (red star). Vacuolization and luminal loss of spermatozoa were to a lesser degree observed in the experimental group administered with 60 mg/kg b.w. apricot kernels. No visible changes in the testicular tissue were observed in the experimental animals supplemented with 300 mg/kg b.w. apricot kernels. AMG, amygdalin; i.m., intramuscular; b.w., body weight.
| Groups | Seminiferous epithelium | Interstitial Leydig cells | Interstitial edema | ||
| Dystrophia | Vacuoles | Dystrophia | Vacuoles | ||
| Control | + | + | + | - | + |
| P1 | ++ | ++ | + | - | - |
| P2 | + | + | + | - | - |
| P3 | ++ | ++ | + | + | + |
| P4 | + | ++ | + | + | + |
Intensity of changes: + low, ++ medium, - none detected.
The semi-quantitative analysis of the photomicrographs revealed that the relative volume of the germinal epithelium was not significantly different amongst the Control and experimental groups (Table 2). Nevertheless, a significantly higher relative volume of the interstitium (p = 0.0040) accompanied by a significantly lower luminar relative volume (p = 0.0042) was found in the P2 group in comparison to the Control.
| Germinal epithelium | Interstitium | Lumen | |
| Control | 66.01 |
10.53 |
23.46 |
| P1 | 63.29 |
14.98 |
21.73 |
| P2 | 62.92 |
21.13 |
15.95 |
| P3 | 70.07 |
9.48 |
20.45 |
| P4 | 65.13 |
12.91 |
21.96 |
Data are presented as mean
Morphometric measurements also revealed that when compared to the Control, a significantly decreased epithelial height was present in the P1 (p = 0.0104) as well as the P2 experimental group exposed to 3.0 mg/kg b.w. AMG i.m. (p = 0.0095; Table 3). Moreover, a significantly decreased luminar diameter was observed in the experimental groups P1 (p = 0.0305) and P2 (p = 0.0318) in comparison to the untreated Control.
| Tubular diameter (µm) | Epithelial height (µm) | Luminar diameter (µm) | |
| Control | 227.30 |
61.37 |
165.93 |
| P1 | 196.90 |
51.18 |
145.72 |
| P2 | 196.50 |
48.41 |
148.09 |
| P3 | 244.70 |
69.84 |
174.86 |
| P4 | 220.10 |
63.62 |
156.85 |
Data are presented as mean
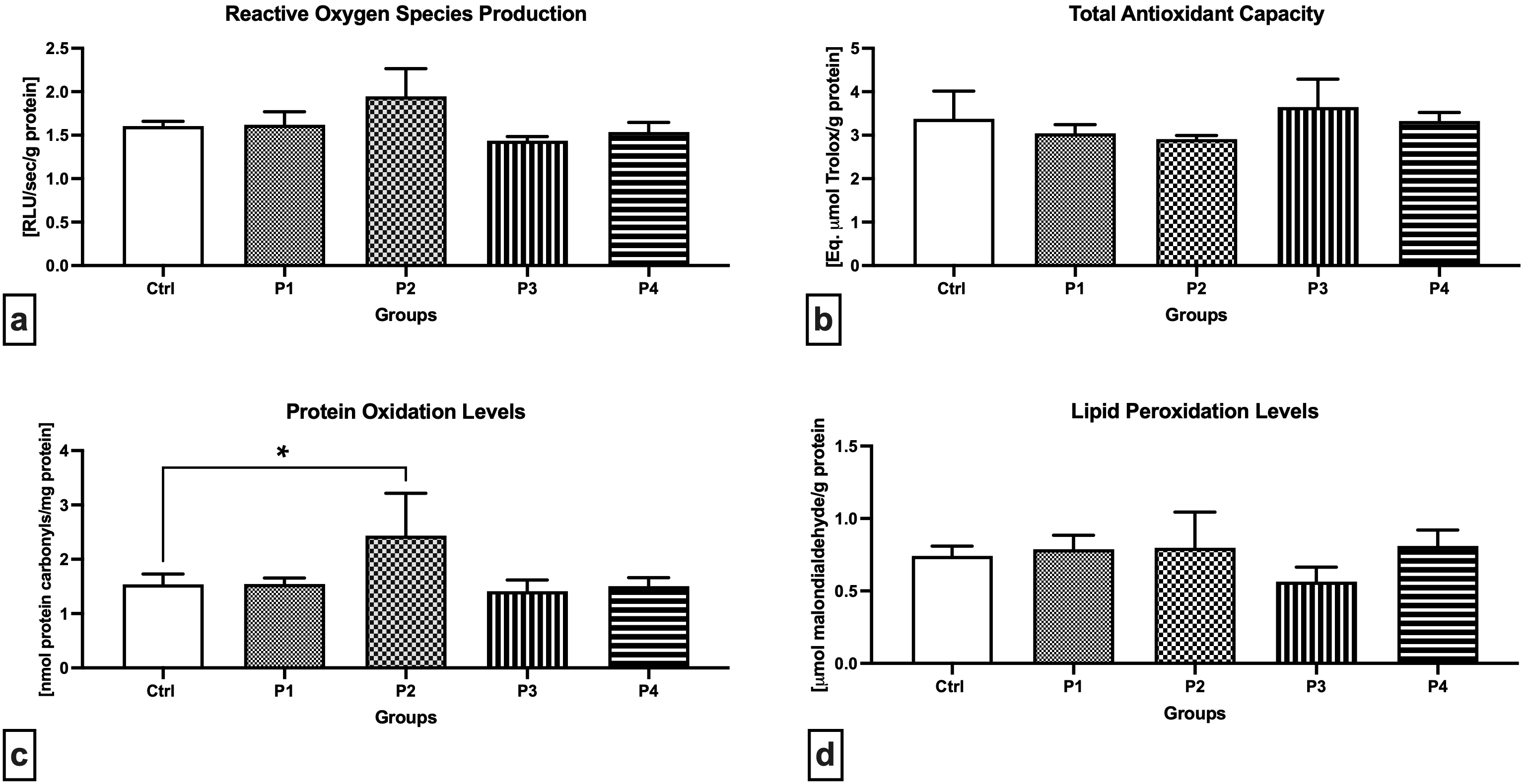
The chemiluminescent analysis revealed increased ROS levels in the P2 group when compared to the Control (Fig. 2a), although no significant differences were recorded. Inversely, a lower, although non-significant, ROS production was observed in the P3 experimental group in comparison to the Control. Changes in the oxidative milieu in the Control and experimental groups were corroborated by a non-significant decline in TAC in the P1 and P2 groups in comparison with the Control (Fig. 2b). In the meantime, experimental groups exposed to crushed apricot kernels (P3 and P4) presented a slight increase in TAC in comparison to the Control. The highest amount of protein carbonyls indicative of oxidative damage to the proteins was recorded in the P1 group administered with 3.0 mg/kg b.w. AMG i.m., which was significantly higher when compared to the Control (p = 0.0118; Fig. 2c). No significant fluctuations were observed in the remaining experimental groups in comparison with the Control. As opposed to protein oxidation, higher although non-significant levels of MDA as the primary marker of LPO were observed in the P2 experimental group (Fig. 2d). Moreover, oral administration of 60 mg/kg b.w. apricot kernels led to a non-significant decrease in the testicular MDA when compared to the Control.
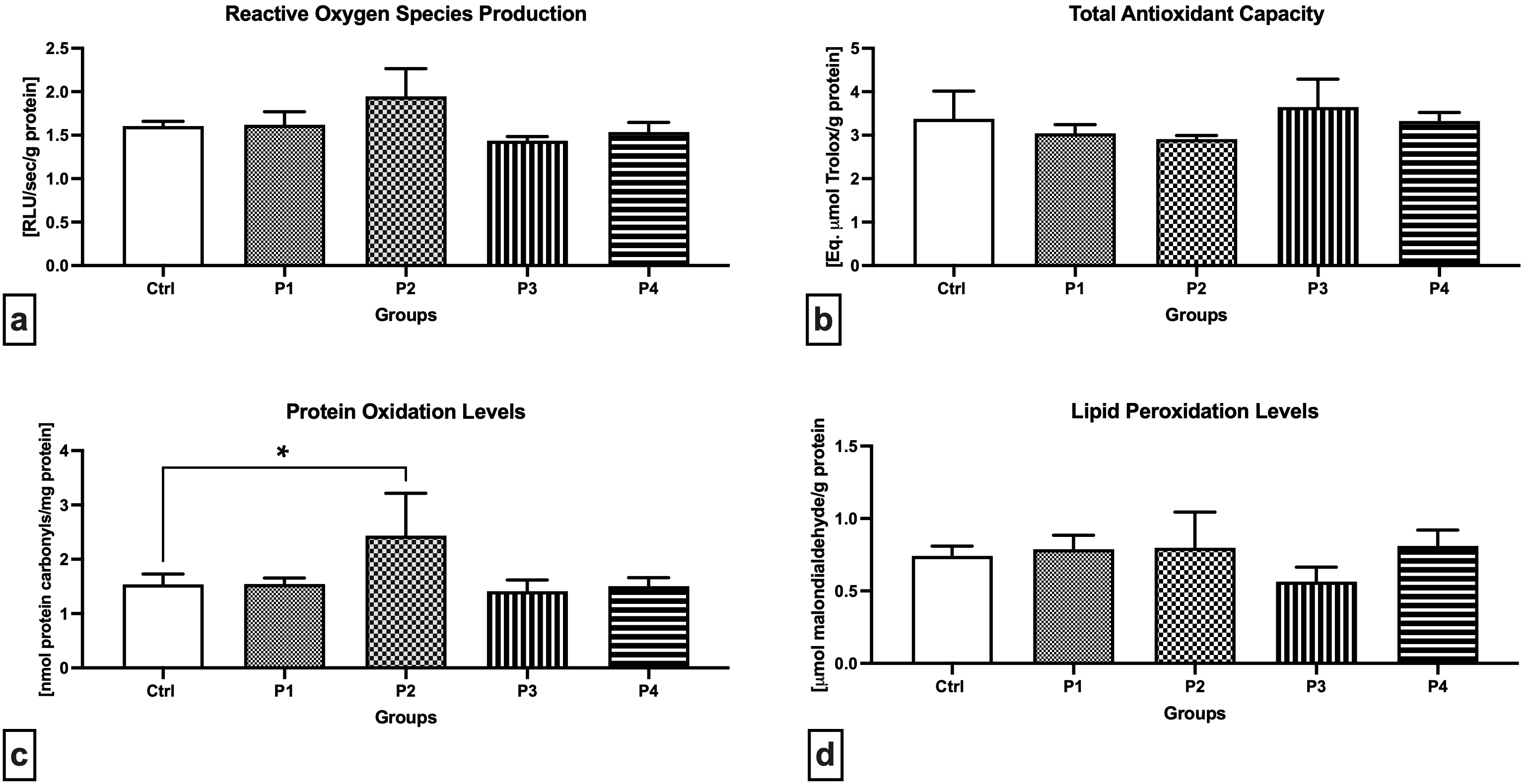 Fig. 2.
Fig. 2.Oxidative profile of the testicular tissue of the control and
experimental animals. (a) Reactive oxygen production. (b) Total antioxidant
capacity. (c) Oxidative damage to the proteins. (d) Oxidative damage to the
lipids. Data are presented as mean
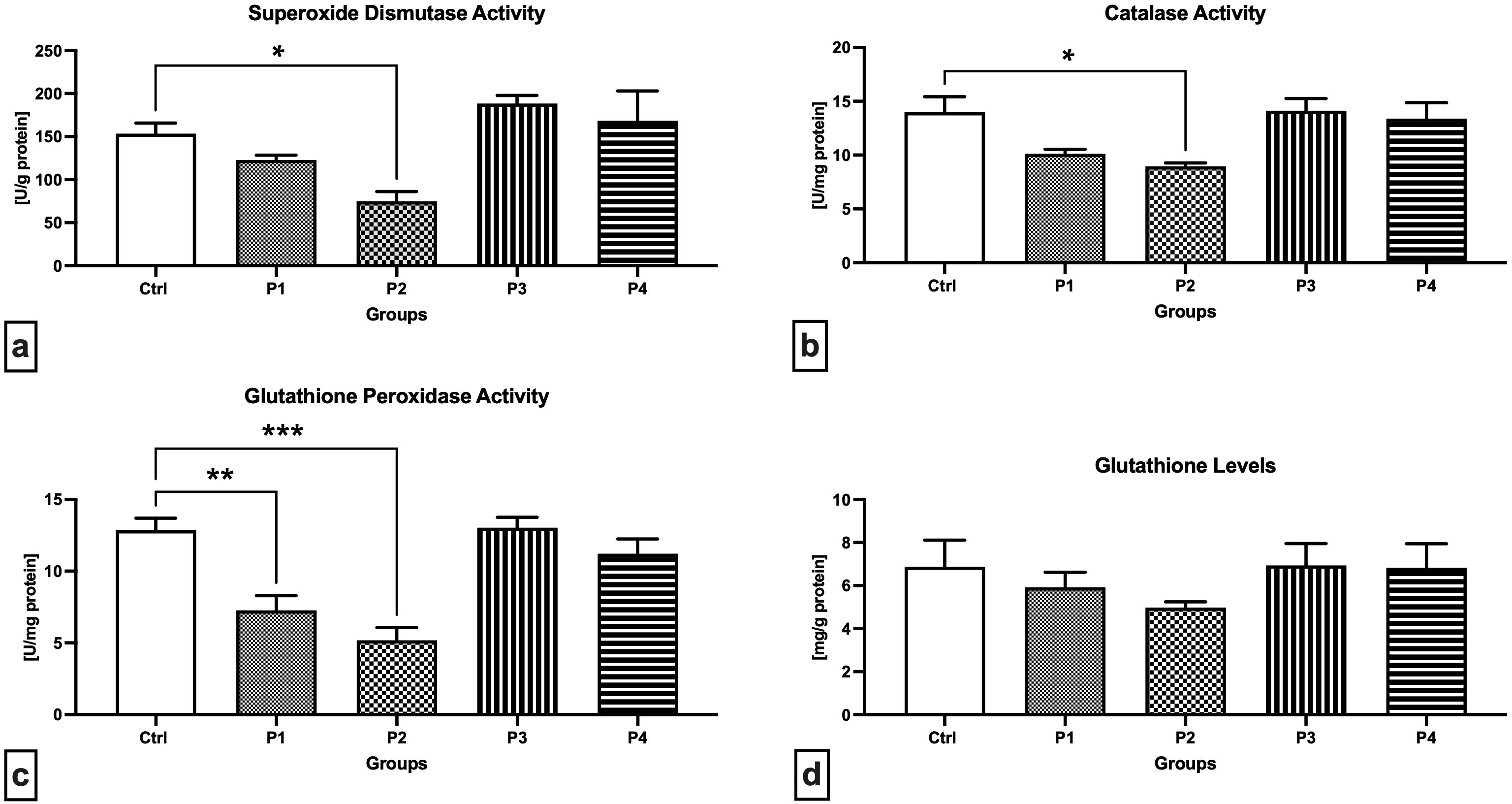
A closer look at the primary components of the antioxidant system of male reproductive structures revealed that SOD activity was significantly decreased in the experimental group P2 exposed to 3.0 mg/kg b.w. AMG i.m. when compared to the Control (p = 0.0464; Fig. 3a). On the other hand, a non-significant increase in SOD activity was recorded in the experimental animals receiving 60 or 300 mg/kg b.w. apricot kernels (P3 and P4, respectively). Similar changes were observed in the case of CAT, the activity of which was the lowest in the P2 experimental group, which was significantly different in comparison to the Control (p = 0.0317; Fig. 3b). No changes in the CAT activity were observed in the P3 and P4 experimental groups. More pronounced changes were recorded in the case of GPx activity with a significant decline observed in the P1 (p = 0.0043; Fig. 3c) and P2 group (p = 0.0002) when compared to the Control. While supplementation of 300 mg/kg b.w. apricot kernels (experimental group P4) led to a slight decrease in GPx activity, no changes were observed between the P3 experimental group and the Control. The levels of GSH as a primary testicular non-enzymatic antioxidant did not differ among the Control and the experimental groups (Fig. 3d). Nevertheless, decreased GSH concentrations were observed in the experimental groups P1 and P2 subjected to intramuscular injection of 0.6 or 3 mg/kg b.w. pure AMG, respectively, when compared to the Control.
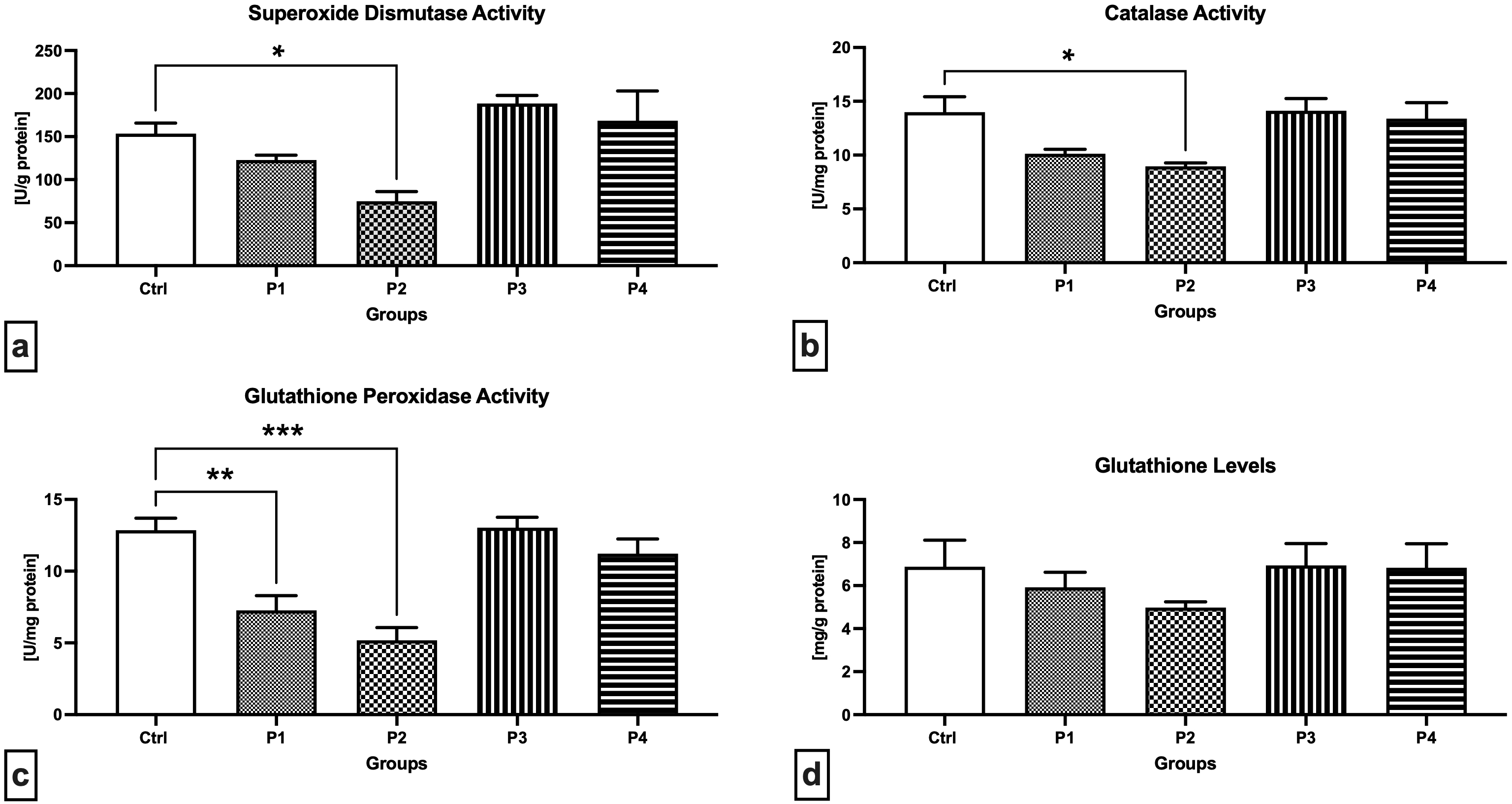 Fig. 3.
Fig. 3.Antioxidant characteristics of the testicular tissue of the
control and experimental animals. (a) Superoxide dismutase activity. (b)
Catalase activity. (c) Glutathione peroxidase activity. (d) Glutathione levels.
Data are presented as mean
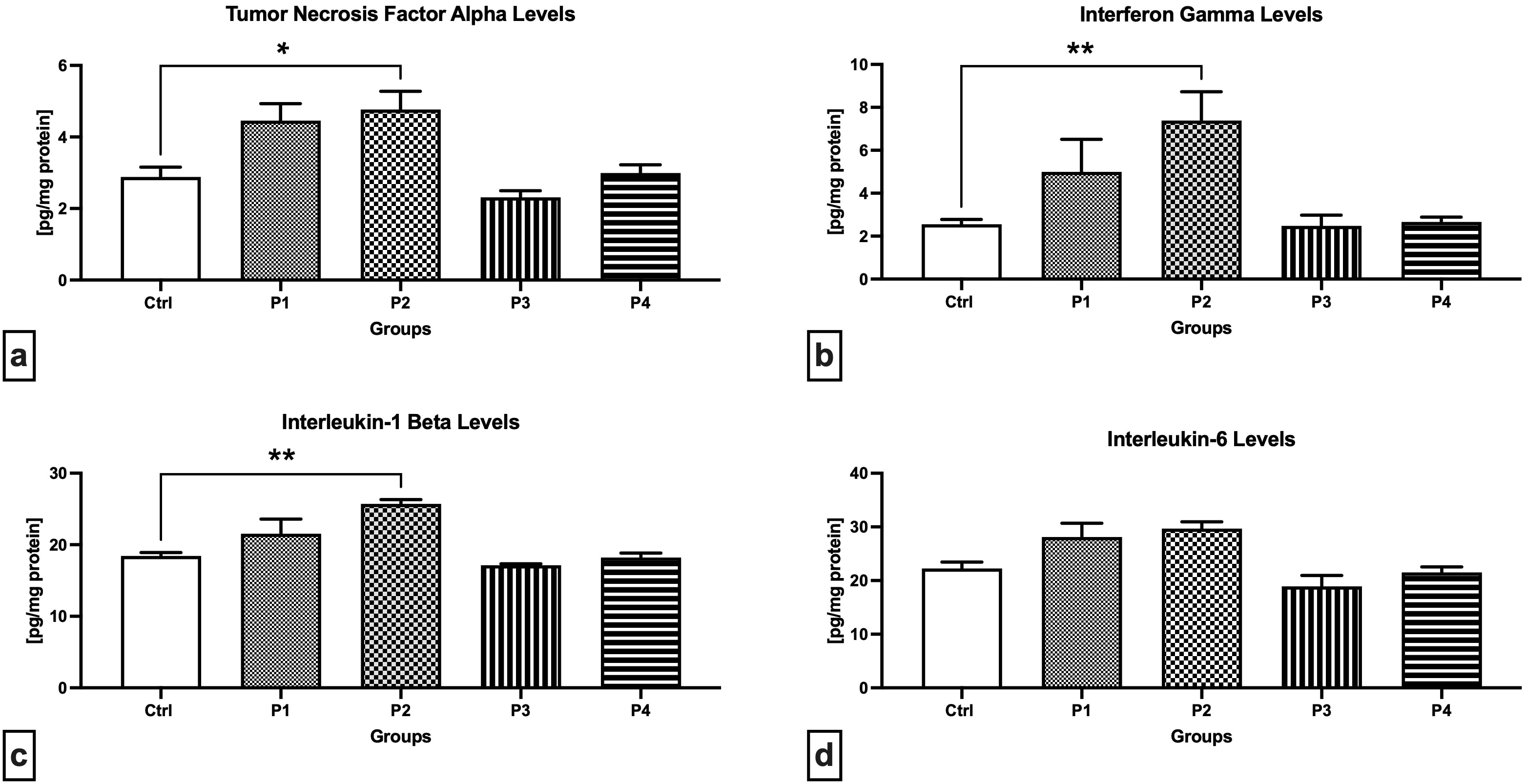
As revealed through the ELISA method, the P2 experimental group presented
significantly higher levels of TNF-
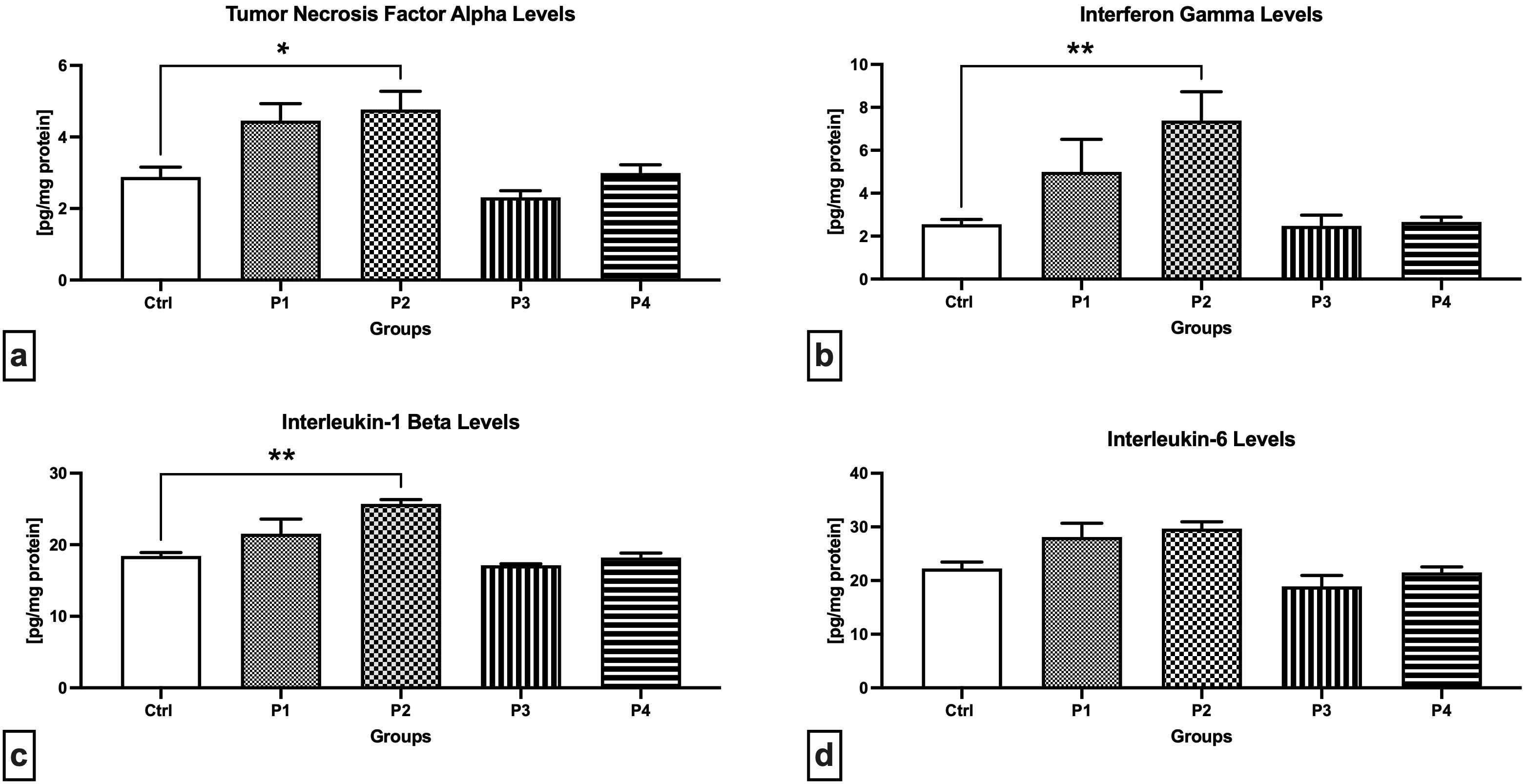 Fig. 4.
Fig. 4.Cytokine levels in the testicular tissue of the control and
experimental animals. (a) Tumor necrosis factor-alpha levels. (b)
Interferon-gamma levels. (c) Interleukin-1 levels. (d) Interleukin-6 levels. Data
are presented as mean
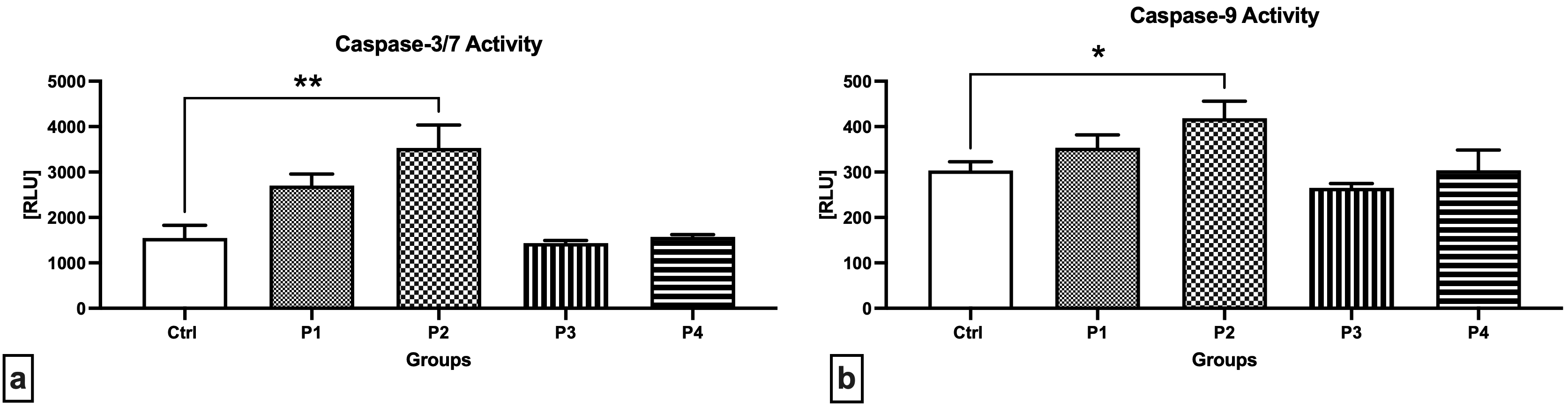
Notably higher activities of caspase-3/7 (Fig. 5a) as well as caspase-9 (Fig. 5b) were detected in the testicular tissue collected from the experimental groups exposed to 3.0 mg/kg b.w. AMG i.m., with significant changes when comparing the Control with the P2 group (p = 0.0014 in case of caspase-3/7; p = 0.0243 with respect to caspase-9). Slightly decreased enzymatic activities were recorded in the experimental groups supplemented with crushed apricot kernels (P3 and P4), although no significant changes were observed.
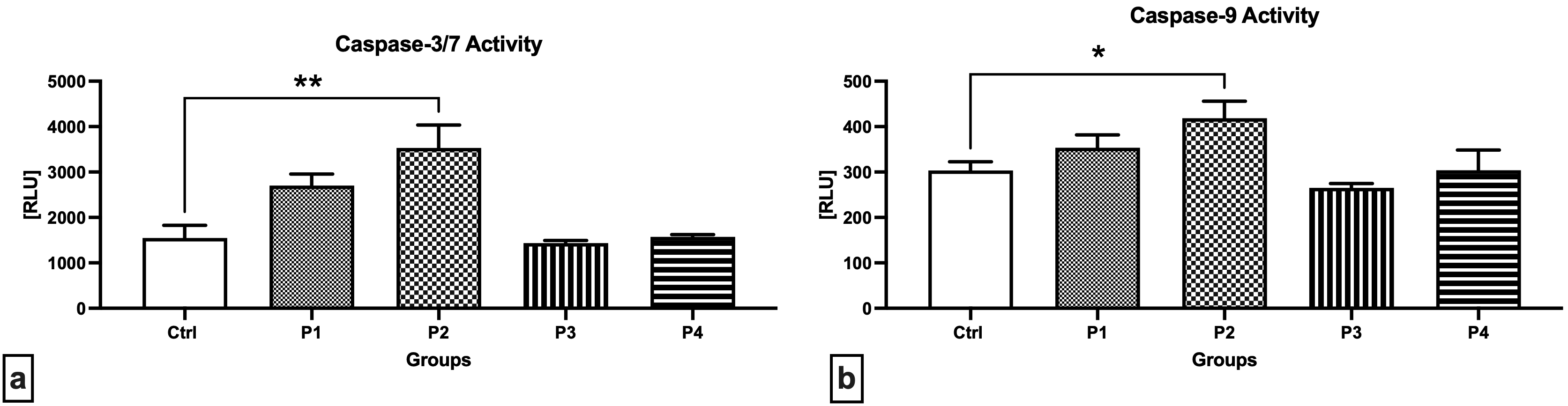 Fig. 5.
Fig. 5.Activity of caspases in the testicular tissue of the control and
experimental animals. (a) Caspase-3/7 activity. (b) Caspase-9 activity. Data are
presented as mean
Over the past years, several in vivo studies have emerged suggesting that AMG does not cause significant changes to the overall health status [22], bone [23], or renal structure [19] of the experimental animals. Nevertheless, it has been reported that intramuscular AMG administration may be associated with time-dependent changes to the balance of male reproductive hormones [26] and a decrease in sperm motion and kinematic characteristics [21]. In this sense, it must be remembered that tumorigenesis and spermatogenesis share some common characteristics, as both processes are characterized by rapidly dividing and differentiating cells, high energy requirements, and an exceptional metabolic profile. On the other hand, the structural and functional uniqueness renders the testes to respond in a very sensitive manner to any imbalance of the internal milieu or surrounding exterior. Since the male fertilization ability by and large depends on the structural integrity and functional activity of male gonads, this paper strived to elucidate the in vivo response of testicular tissue towards pure amygdalin administered intramuscularly, or to crushed apricot kernels supplemented orally.
Our data reveal that pure AMG, as opposed to apricot kernels, may affect the testicular dynamics, particularly when higher (“therapeutic”) doses are used. These effects manifested themselves primarily through a compromised activity of antioxidant enzymes representing the first line of defense against oxidative stress, accompanied by an increase of oxidized proteins, and inflammatory and apoptotic processes; however, these alterations were not translated into notable changes to the testicular architecture.
While the toxicity and efficacy of AMG in in vivo studies needs to be
further understood, it has been previously postulated that orally administered
AMG releases a high amount of cyanide [34], either through the activity of
Cyanide as a resulting product of AMG metabolism acts as a potent neurotoxin, which may cause alterations to vital organs including the heart, kidney, brain, or liver, although, Shivanoor and Muniswamy [39] observed no changes in the body or reproductive organ weights of rats perorally treated with (0.64 and 1.2 mg/kg) sodium cyanide for 90 days. As discussed earlier, the primary toxicity of HCN lies in its ability to bind to the cytochrome oxidase terminal in the mitochondrial respiratory chain causing an obstruction in the cells’ ability to use oxygen [40], ROS overgeneration that is accompanied by lipid peroxidation, and cell death [41, 42]. Furthermore, oxidative overload may develop if the effects of ROS are not properly counteracted with antioxidants [43].
While lipid peroxidation is considered the central mechanism of oxidative damage, only slight changes in the levels of MDA were observed in this study. Inversely, a significant increase in the levels of protein carbonyls was observed following intramuscular treatment with 3.0 mg/kg b.w. AMG. This observation agrees with Albogami et al. [37] who suggest that AMG decomposition is associated with the overproduction of benzaldehyde, which will then interact with hydrogen peroxide to form carboxylate and benzenediol [44]. Hence, we may hypothesize that protein carbonylation may be the primary oxidative mechanism of AMG through which ROS accumulation and excessive levels of benzaldehyde may trigger protein oxidation even before the lipids are compromised by peroxidation.
Intramuscular treatment particularly with 3.0 mg/kg b.w. AMG significantly
modulated the activity of all antioxidant enzymes and decreased the levels of
glutathione as a prominent testicular non-enzymatic antioxidant. Changes in the
patterns of antioxidant molecules indicate a shift in the oxidative milieu driven
by ROS overproduction and a depletion of the inherent antioxidant capacity, which
corroborates previous studies on AMG toxicity [22, 37, 43]. A compromised capacity
of SOD to dismutate superoxide to hydrogen peroxide (H
Previous reports have revealed a close association between testicular damage and increased antioxidant depletion accompanied by oxidative damage to the proteins and lipids [46]; nevertheless, our histology and morphometry data indicate only slight changes to the testicular architecture following treatment with AMG or apricot kernels. Inversely, Albogami et al. [37] have observed pathological alterations in the testicular tissue, such as fewer spermatogonia, cellular necrosis, epithelial disarray, and degeneration of Leydig and Sertoli cells. This discrepancy may be explained by the fact that such changes were observed only in rats administered with extremely high AMG doses (200 mg/kg b.w.), whereas animals exposed to lower AMG doses (50 and 100 mg/kg b.w.), presented with normal cell arrangement in the lumen of the seminiferous tubules with no histopathological alterations and active spermatogenesis. Hence, we may postulate that carefully selected low AMG doses may not have a detrimental impact on the structural integrity of male reproductive organs.
It is apparent, however, that induced cell death could be a paramount mechanism
of AMG toxicity even in the reproductive tissues. Our data collected from the
assessment of caspase activity agree with Erkkilä et al. [47] who
observed germ cell death with a concomitant decrease of adenosine monophosphate
(AMP), adenosine diphosphate (ADP), and adenosine triphosphate (ATP) in human
seminiferous tubules cultured in the presence of cyanide. The cessation of energy
production may stem from proton (H
Cell disintegration and subsequent death may be defined as a cluster of
molecular processes that may trigger the secretion of cytokines [50]. As
unraveled by earlier reports, cyanide toxicity is associated with the activation
of the transcription factor nuclear factor-kappa B (NF-
The results of this study revealed that neither of the selected doses of apricot kernels caused any structural alterations to the testicular tissues. Moreover, oral treatment with apricot kernels had slight antioxidant and anti-inflammatory effects, which agrees with the findings of Albogami et al. [37]. It seems that like other bioactive molecules, amygdalin presents a peculiar dichotomy—whilst high AMG doses may have toxic to lethal effects, carefully selected low doses present a biological activity that exhibits beneficial effects on the living system. Pure AMG or foodstuffs containing this molecule have been shown to act as ameliorating agents in studies of induced toxicity, particularly through chemotherapeutic agents [55, 56]. It seems that AMG or apricot kernels could become an interesting complementary option to support conventional therapy of a variety of ailments; this future prospect is, however, subject to larger cohort and/or case-control studies to clarify the existing discrepancies.
Despite notable changes observed in the Control and experimental groups, sample size is the most significant limitation of this study. A relatively low number of animals representing each pre-established group may have impacted the power and complexity of this study. At the same time, an undersized population of animals used in our experimental design may not be fully representative of the population, hence affecting the precision and validity of the collected data. As such, follow-up and/or complementary in vivo studies on a larger population of animal models (rodents or rabbits) may add more veracity to the data obtained in this report. Another limiting factor lies in the aim to only investigate subacute doses within a shorter timeframe. Hence, to generalize the results, the treatment period should have taken longer, whilst considering higher doses of AMG as well as apricot kernels. At the same time, knowledge of the specific signaling pathways that are affected or regulated by AMG on a genetic, epigenetic, or molecular level is more than necessary to conclusively determine the suitability of pure AMG or apricot kernels as relevant therapeutic agents.
The present study suggests that short-term intramuscular administration of amygdalin exerted negative effects on rabbit testicular tissue through oxidative, inflammatory, and pro-apoptotic mechanisms, which may lead to a decreased reproductive potential of the animals. On the other hand, oral supplementation of apricot kernels did not present with changes to the testicular milieu. The specific conditions leading to the toxicity of amygdalin and its food sources should be further studied to determine optimal doses, timeframes, and routes of amygdalin administration to ensure its safety and effectiveness as a potential complementary cancer treatment option in the future.
The datasets generated during and/or analyzed in this study are available from the corresponding author upon reasonable request.
AK and ET designed the study. ET, MĎ, MH, JP and EK performed the research. ĽC and ĽO oversaw the animal care and treatment regime. MH and RJ performed the animal sacrifice and sample collection. ET and MĎ analyzed the data. ET and AK wrote the manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work. All authors contributed to editorial changes in the manuscript, read and approved the final manuscript.
Institutional and national guidelines for the care and use of animals were followed appropriately, and all experimental procedures were approved by the State Veterinary and Food Institute of Slovak Republic, no. 3398/11–221/3 and Ethic Committee.
We wish to thank the CeRA Team of Excellence for their financial and promotional support.
This research was funded by the Slovak Agency for Research and Development, grant numbers APVV-18-0312 and APVV-21-0095; and by the Scientific Grant Agency of the Ministry of Education, Science, Research and Sport of the Slovak Republic and the Slovak Academy of Sciences, grant number VEGA 1/0266/20.
Given her role as Guest Editor, Eva Tvrdá had no involvement in the peer-review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Amedeo Amedei. The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.