
 , Ginnae Ahn 1,6,*
, Ginnae Ahn 1,6,*1 Department of Food Technology and Nutrition, Chonnam National University, 59626 Yeosu, Republic of Korea
2 Choung Ryong Fisheries Co., Ltd., 63612 Jeju, Republic of Korea
3 Jeju International Marine Science Center for Research & Education, Korea Institute of Ocean Science & Technology (KIOST), 63349 Jeju, Republic of Korea
4 Food Safety and Processing Research Division, National Institute of Fisheries Science, 46083 Busan, Republic of Korea
5 Department of Marine Bio and Medical Sciences, Hanseo University, 32158 Seosan-si, Republic of Korea
6 Department of Marine Bio-Food Sciences, Chonnam National University, 59626 Yeosu, Republic of Korea
Abstract
Backgrounds: Melanogenesis, regulated by genetic, hormonal, and
environmental factors, occurs in melanocytes in the basal layer of the epidermis.
Dysregulation of this process can lead to various skin disorders, such as
hyperpigmentation and hypopigmentation. Therefore, the present study investigated
the effect of ultrasonic-assisted ethanol extract (SHUE) from Sargassum
horneri (S. horneri), brown seaweed against melanogenesis in
Keywords
- Sargassum horneri
- melanogenesis
- tyrosinase inhibition
- B16F10 murine melanocytes
Human skin, hair, and eye coloration are intricately governed by a biological process called melanogenesis. This process involves synthesizing and distributing melanin, a polymerized pigment originating from the amino acid tyrosine. Beyond its role in coloration, melanin is pivotal in safeguarding the skin against harmful ultraviolet (UV) radiation and contributing to overall homeostasis [1]. In addition to being a fundamental interest, understanding the intricate mechanisms of melanogenesis has important implications in various fields, including dermatology, genetics, and cosmetic research.
Melanogenesis begins with the activation of specialized pigment-producing cells known as melanocytes, located in the basal layer of the epidermis, hair follicles, and uveal tract of the eye [2]. Melanocytes derive from melanoblasts originating from embryonic neural crest cells [3] and, in the skin, one melanocyte is surrounded by approximately 36 keratinocytes [4]. Melanogenesis is regulated by a complex interplay of genetic, cellular, and environmental factors. Upon stimulation, melanocytes synthesize and deposit melanin within specialized organelles called melanosomes. These melanosomes are subsequently transported and transferred to adjacent keratinocytes, ultimately enhancing the pigmentation of tissues [4]. The production of eumelanin, which gives brown to black color, and pheomelanin, which imparts yellow to red hues, occurs through additional enzymatic processes. This outcome depends on factors such as the enzyme ratios, substrate availability, and the pH conditions inside melanosomes [4].
Several regulatory mechanisms control melanogenesis, ensuring its fine-tuned
regulation in response to physiological and environmental cues. The most notable
regulators include
Understanding the molecular intricacies of melanogenesis has been the subject of intense scientific investigation. Researchers have identified numerous factors involved in melanocyte development, differentiation, and pigment production. Elucidating the signaling pathways, transcription factors, and regulatory elements that govern melanogenesis has provided valuable insights into the complex interplay between genetic determinants and environmental influences [8]. Melanogenesis goes beyond determining pigmentation; it has clinical significance linked to skin disorders. Conditions like melasma and post-inflammatory hyperpigmentation, for example, stem from excessive melanin production and uneven distribution, leading to hyperpigmentation issues [9].
As per the reports, laboratory scale-70% ethanol extracts of Sargassum
spp. found around the Korean Sea indicated tyrosinase inhibitory effects [10].
Another interesting study conducted by S.H. CHA and the team found that
Sargassum silquastrum reduced cellular melanin synthesis and tyrosinase
activity along with the inhibitory effects on the pigmentation of zebrafish
in vivo model [11]. Moreover, M.S. Azam et al. [12] reported
that ethanolic extract from Sargassum serratifolium indicated hypo
pigmenting properties on B16F10 murine melanocytes. In addition,
Sargassum cristaefolium and Sargassum thunbergii have exhibited
the inhibitory potential of melanin production in B16F10 murine melanocytes [13, 14]. By considering the potential of Sargassum spp. on skin whitening,
Sargassum horneri 70% ethanol extract (SHE70), S. horneri 50%
ethanol extract (SHE50), and S. horneri ultrasonic-assisted ethanol
extract (SHUE) were screened for their melanin inhibitory potential in
Dimethyl sulfoxide (DMSO),
3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT), bovine
serum albumin (BSA), ethidium bromide, gallic acid,
S. horneri was collected along the coast of Jeju Island and its species were identified by the Biodiversity Research Institute (Jeju, South Korea). S. horneri was washed with running water and dried at room temperature. The 200 g of dried sample soaked in distilled water and 30 kg of 50% ethanol in water (v/v) were reacted in the ultrasonic apparatus (MD-1200PG, Mirae ultrasonic, Gyeonggi-do, KR) at 4 °C and ultrasonic frequency of 28 kHz. After 5 h, the supernatant was filtered using Whatman No. 6 filter paper (Whatman Maidstone, Buckinghamshire, UK) following centrifugation at 4800 rpm for 10 min. The resulting ultrasonic-assisted S. horneri ethanol extract (SHUE) was concentrated by using rotary evaporation.
For the preparation of ethanol extracts, 150 g of the dried S. horneri powder was separately reacted with 3 L of 50% or 70% ethanol for 12 h at room temperature. Following that, the extracts were centrifuged, and the supernatants were filtered by using a vacuum filtration apparatus. Each supernatant was rotary evaporated and used as SHE50 and SHE70 respectively in the study.
Following the procedures outlined in one of our earlier research, the total
polyphenolic content, total protein content, and carbohydrate content of SHE70,
SHE50, and SHUE were determined [15]. To assess the phenolic composition, a
gradient concentration of gallic acid was used as the reference standard. The
total protein content was determined using the Lowry method with BSA as the
reference standard, and the carbohydrate content was assessed using the
phenol–sulfuric method with d-glucose as the reference standard. The
High-performance liquid chromatographic (HPLC) analysis was implemented to
identify fucosterol content in the sample. Waters Alliance e2695 Separations
Module (Milford, MA, USA) equipped with Waters 2489 UV/Vis Detector (USA) and YMC
Pack-Pro C18 column (4.6
The B16F10 murine melanocytes were purchased from the Korean Cell Line Bank
(KCLB, Seoul, Republic of Korea). The cells were validated by Short Tandem Repeat (STR) profiling and
tested negative for mycoplasma and were cultured in DMEM supplemented with 10%
heat-inactivated FBS along with a 1% penicillin/streptomycin antibiotic mixture.
A controlled humidified environment with 5% CO
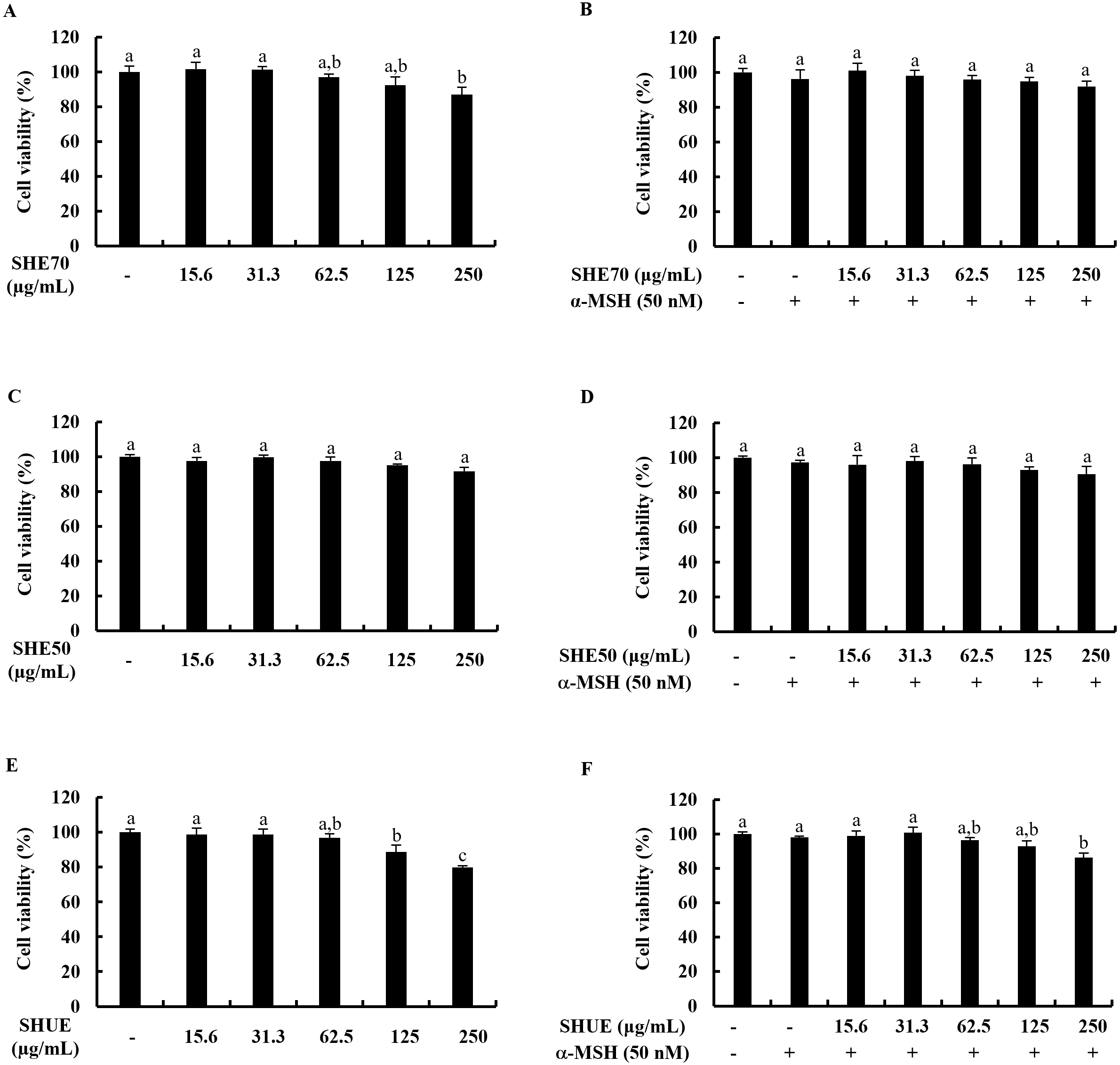
To examine the cytotoxicity of SHE70, SHE50, and SHUE as well as their
cytoprotective effects on B16F10 murine melanocytes against
The B16F10 murine melanocytes were seeded in 10 cm cell culture dishes at a
density of 3
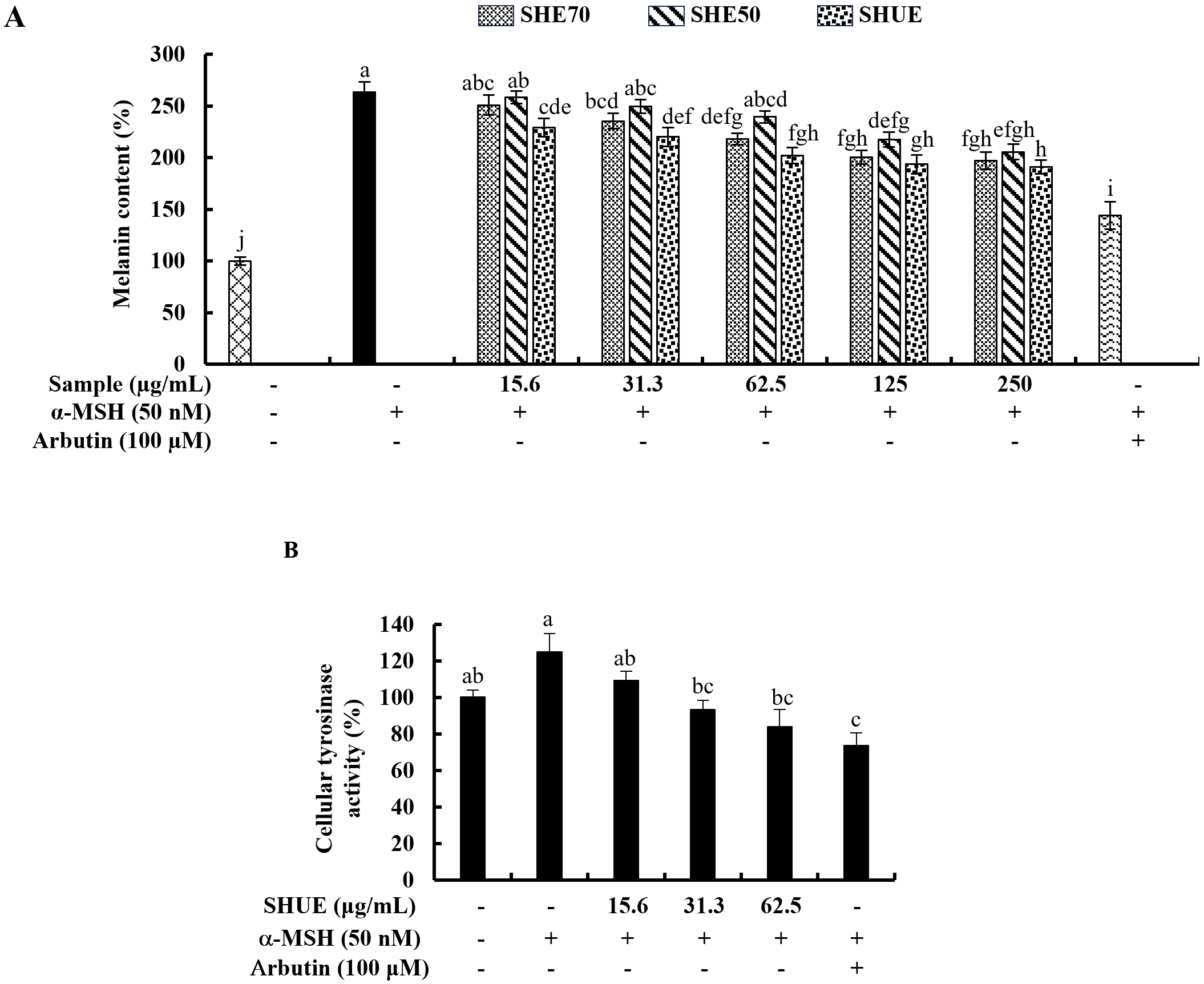
Cellular tyrosinase activity was assessed by using cells cultured in 10 cm cell
culture dishes. The analysis method was obtained from one of the previous studies
[17]. In brief,
Cells were seeded (1
All statistical analyses of the study were performed using the SPSS software
(IBM, SPSS Inc., Version 24.0, Chicago, IL, USA). One-way analysis of variance
(ANOVA) followed by Duncan’s multiple range tests was used to evaluate the
significant variations among data sets, and data were presented as the mean
As per the analysis, yield, total carbohydrate content, crude protein content,
and total phenolic compound content of each sample were presented in Table 1. The
extract yield of three independent replicates indicated the highest average yield
on a dry basis was 28.70
| Sample | Yield (%) |
Carbohydrate (%) |
Crude protein (%) |
Total phenolic compounds (%) |
| SHE70 | 9.98 |
4.08 |
1.95 |
2.38 |
| SHE50 | 7.28 |
3.35 |
1.19 |
1.37 |
| SHUE | 28.70 |
8.74 |
14.31 |
6.25 |
B16F10 murine melanocytes were treated with 15.6, 31.3, 62.5, 125, and 250
µg/mL concentration of each sample either alone or together with
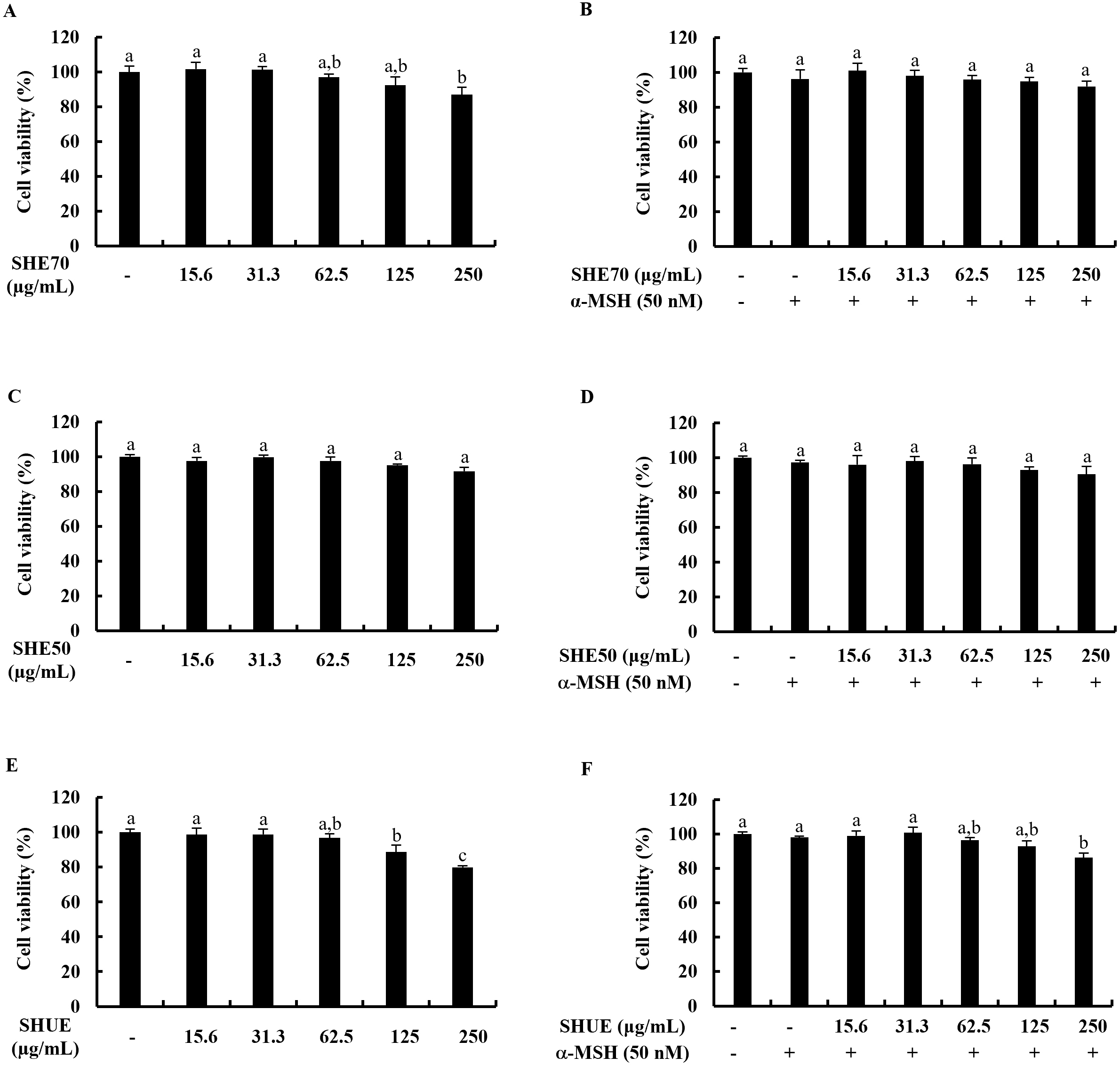 Fig. 1.
Fig. 1.Effect of extracts on B16F10 murine melanocytes. Cytotoxicity
analysis of (A) SHE70, (C) SHE50, (E) SHUE, and analysis of the effect on cell
viability of (B) SHE70, (D) SHE50, and (F) SHUE. Values were expressed as mean
The potential of each sample in inhibiting intracellular melanin synthesis was
analyzed and the results illustrated in Fig. 2A indicated significant
intracellular melanin inhibitory activity at 31.3 µg/mL of SHUE.
Further, as per the results indicated in Fig. 2A, SHUE effectively downregulated
the cellular melanin content in a dose-dependent manner in contrast to the
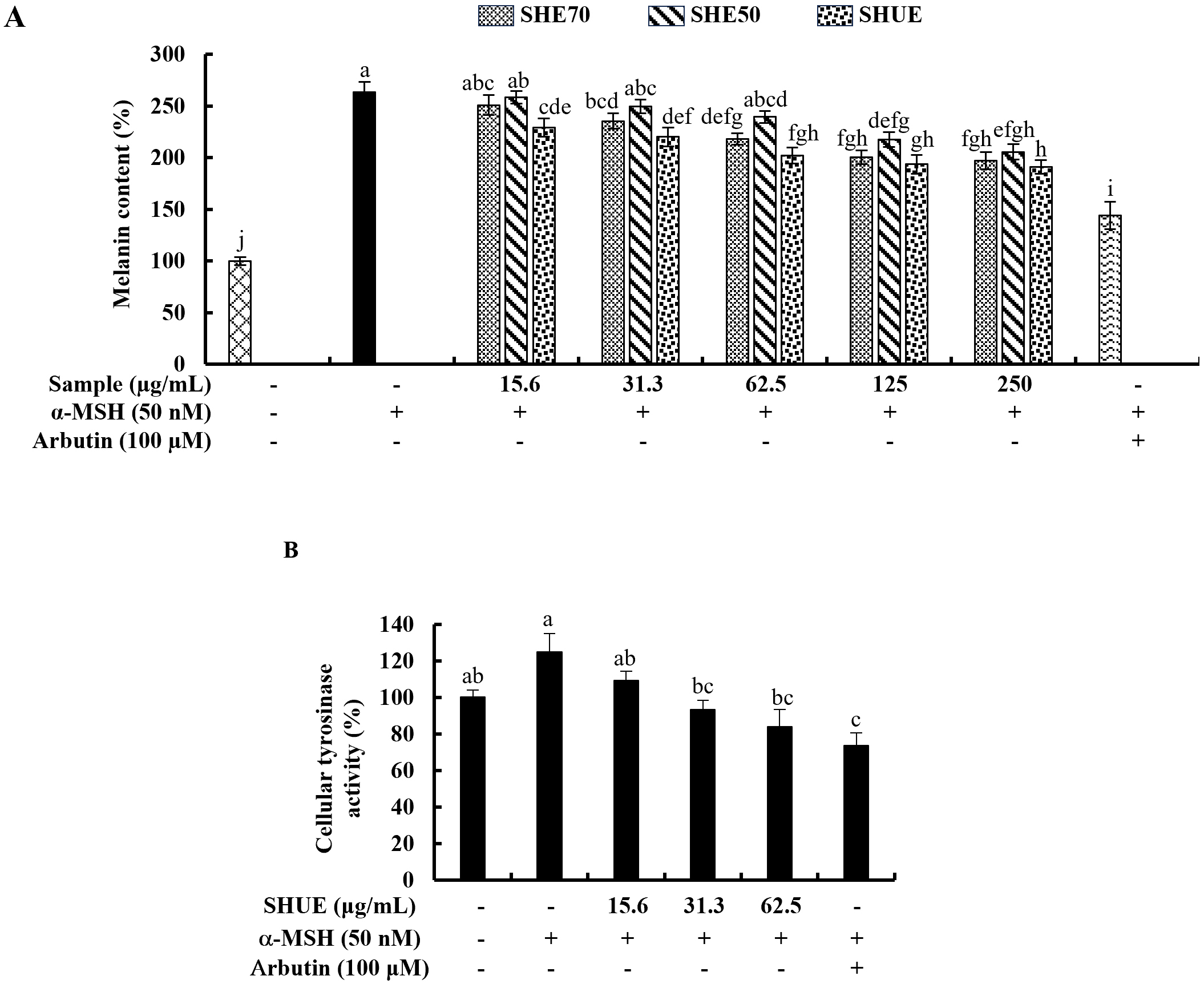 Fig. 2.
Fig. 2.Effect of SHUE on melanin synthesis and tyrosinase activity.
(A) Effect of SHE70, SHE50, and SHUE on melanin synthesis in
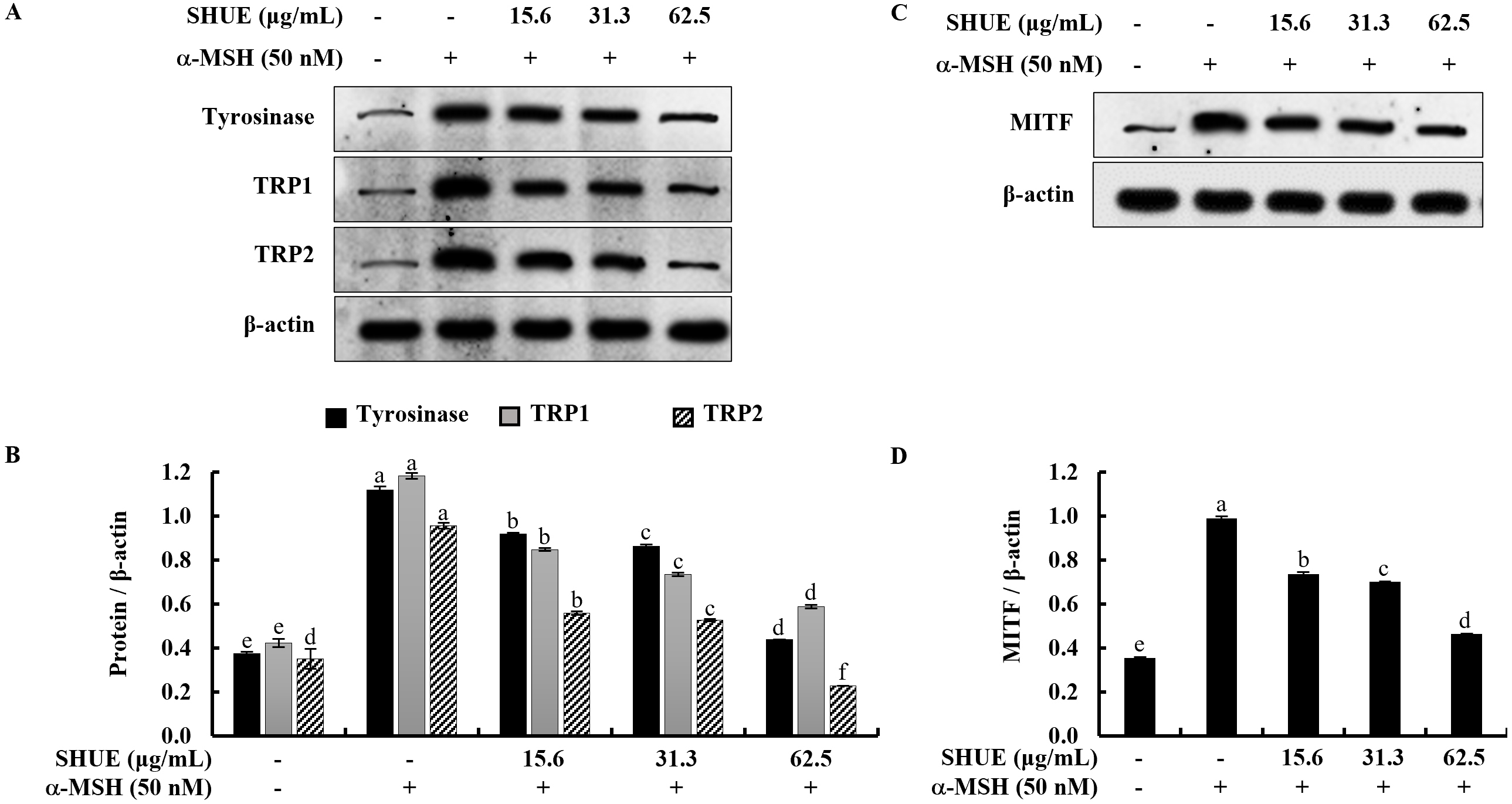
Given the significant reduction in melanin content observed with SHUE, the study
was extended to investigate its potential impact on the expression of melanogenic
enzymes, including tyrosinase and tyrosinase-related proteins (TRPs). Western
blot analysis results showed that SHUE downregulated the expression of
tyrosinase, TRP1, and TRP2 in a dose-dependent manner (Fig. 3A).
Microphthalmia-associated transcription factor (MITF) has been reported to act as
a master transcription factor for the expression of tyrosinase, TRP1, and TRP2 in
the process of melanin biosynthesis [21]. In Fig. 3C, the analytical results
indicated that SHUE similarly downregulated MITF expression. In addition,
relative folds indicated in Fig. 3B,D confirmed the significant downregulation of
MITF, tyrosinase, and tyrosinase-related proteins (TRPs) (Original western blot images can be found in Supplementary Fig. 2). Taken together, these
data showed the inhibition of melanogenesis by SHUE occurred in
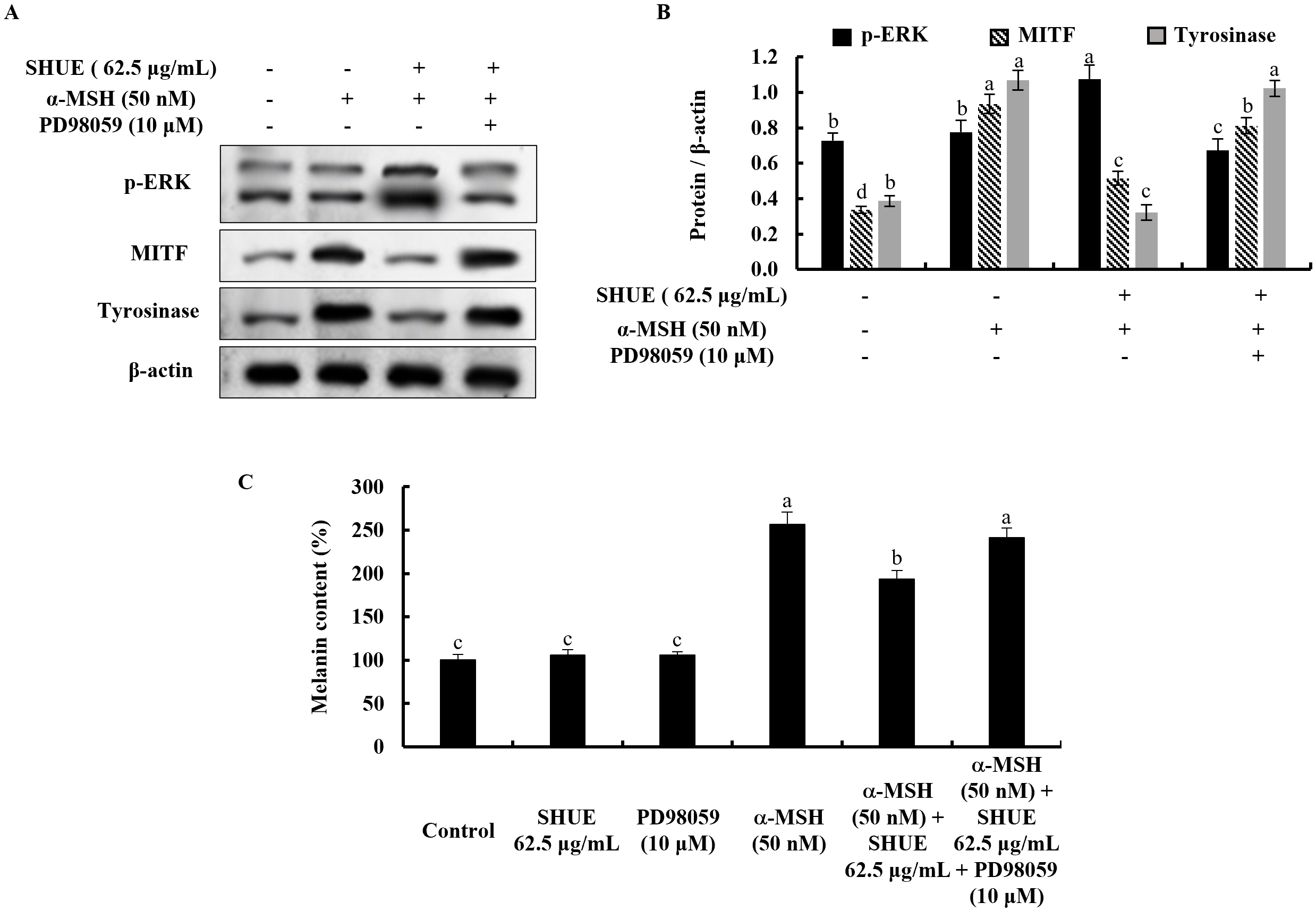
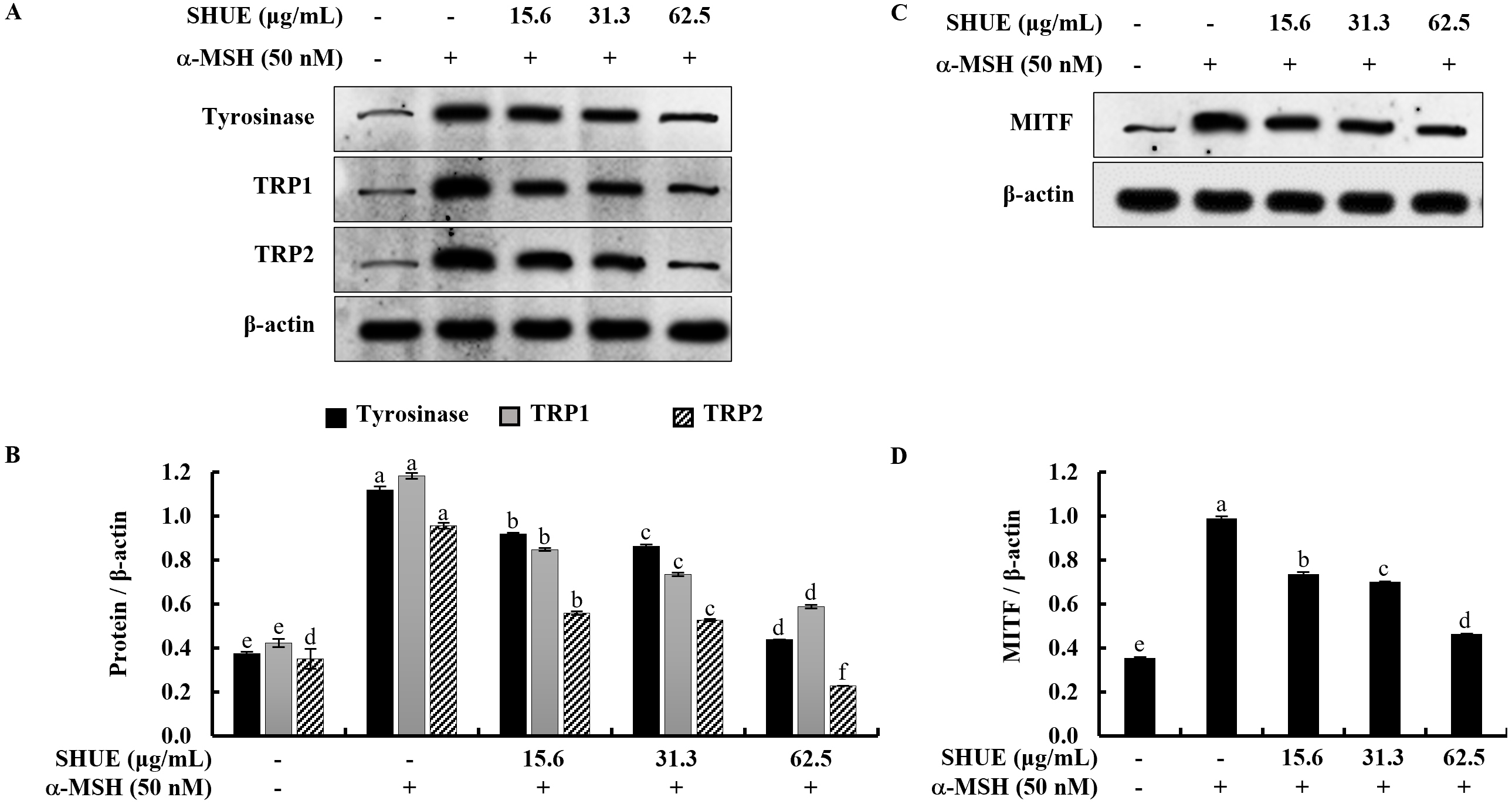 Fig. 3.
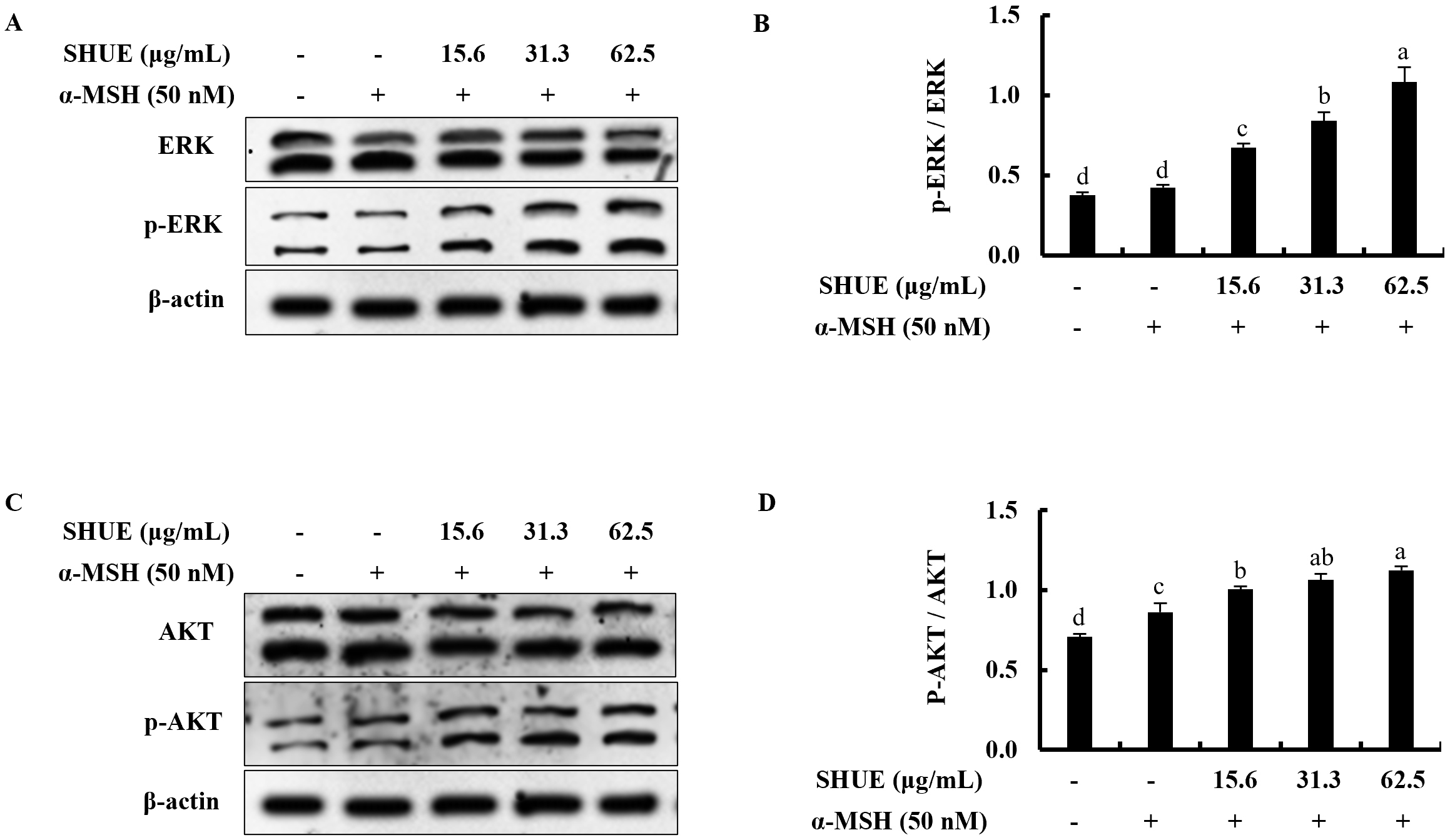
Fig. 3.Effect of SHUE on the expression of extracellular signal-regulated kinase (ERK)-associated
melanogenesis-related protein expression. (A,C) Western blot analysis and, (B,D)
relative folds. Values were expressed as mean
The effect of ERK on the melanogenesis process was assessed via Western
blotting. Under
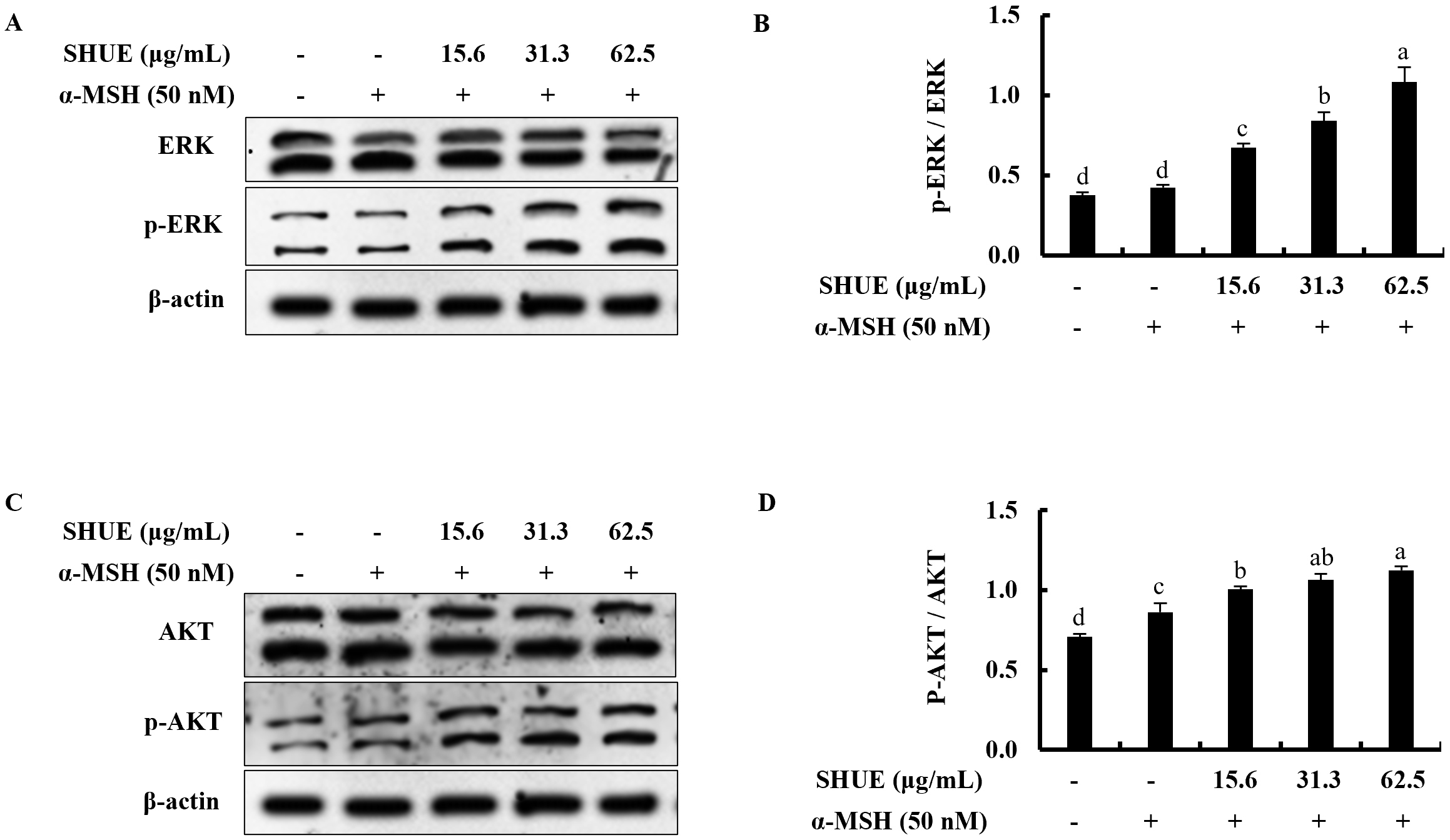 Fig. 4.
Fig. 4.Effect of SHUE on the expression of ERK and AKT signaling
proteins in
With the addition of an ERK inhibitor (PD98059), the ERK phosphorylation is
decreased in the SHUE-treated cells (Fig. 5A,B). Simultaneously, MITF and
tyrosinase indicated expression level changes toward melanogenesis with the
treatment of PD98059. This process occurs through the ERK-involved signaling
pathway in
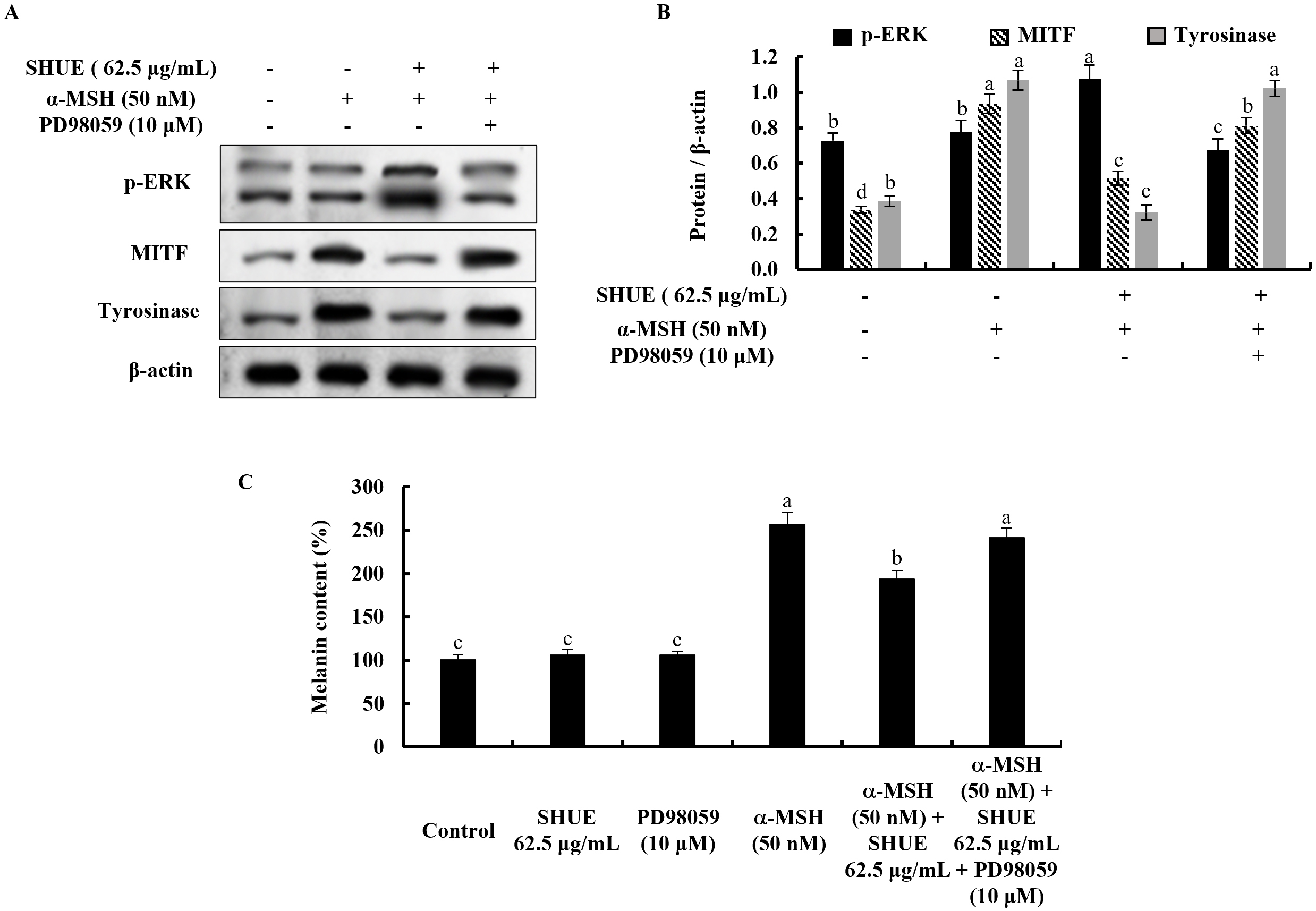 Fig. 5.
Fig. 5.Effect of the PD98059 in
Skin whitening products have gained popularity among consumers worldwide, with an estimated 15% of the global population engaging in their use. As reported in 2022, approximately $13 billion was expended on skincare and cosmetic products alone in the Asia-Pacific region [22]. Unfortunately, the global circulation of skin-lightening cosmetics containing illicit and hazardous ingredients is readily available in various countries and sold through both conventional and online ways [23]. With health concerns, the quest for naturally available multidisciplinary drugs for skin whitening is of paramount significance in dermatological research [24]. Such compounds hold the potential to address hyperpigmentation and other skin-related issues while minimizing adverse effects [24]. Natural ingredients offer a promising avenue to develop safe and effective cosmeceutical agents, as they often encompass a range of bioactive compounds with complementary mechanisms of action [25]. By revealing the power of natural sources, we can tap into a wealth of biodiversity and traditional knowledge, accelerating the discovery of novel compounds for achieving even skin tone. This pursuit aligns with the growing demand for safer alternatives in the cosmetics and dermatology industries, emphasizing the need for a holistic approach to skin whitening that prioritizes both efficacy and safety.
In the present study, we prepared the three different extracts (SHE50, SHE70, and SHUE) based on two extraction approaches, room temperature S. horneri ethanol extraction and ultrasonic assisted ethanol extraction. The ultrasonic-assisted ethanol extraction method and room temperature ethanol extraction method which were used in this study are both implemented in extracting compounds from botanical materials, yet they diverge in efficiency and the techniques used. The ultrasonic-assisted ethanol extraction method utilizes high-frequency sound waves to enhance the extraction process. This results in increased penetration of the solvent into the plant matrix [26]. It facilitates the extraction of target compounds more rapidly and efficiently compared to room temperature extraction, which relies solely on the natural diffusion of the solvent. Moreover, ultrasonic-assisted extraction typically requires shorter extraction times and lower temperatures, thus preserving thermally sensitive compounds better [27]. On the other hand, room-temperature ethanol extraction is simpler and more accessible, requiring no specialized equipment. Results obtained from the compositional analysis as well as HLPC analysis for the target compound (Fucosterol) indicated that SHUE consists of a higher yield and a better intracellular melanin reduction potential in contrast to the other two extracts in the preliminary investigation.
When discussing melanogenesis, the key enzyme namely tyrosinase which is a
copper-containing glycoprotein catalyzes the hydroxylation of L-tyrosine to
L-3,4-dihydroxyphenylalanine (L-DOPA). Subsequently, L-DOPA is converted to
dopaquinone [28]. Recent studies have shed light on the role of
microenvironmental factors in melanogenesis. For instance, UV radiation is a
potent inducer of melanin production, acting through the activation of various
signaling cascades, including the mitogen-activated protein kinase (MAPK) pathway
[4, 29]. UV exposure triggers DNA damage and oxidative stress, leading to the
activation of melanocyte-stimulating factors and the upregulation of melanogenic
enzymes. These adaptive responses serve as a protective mechanism against
UV-induced DNA damage by increasing the production and distribution of melanin
[30]. In addition to UV radiation, other environmental factors, such as hormonal
changes, inflammation, and chemical exposure, can influence melanogenesis.
Hormones, particularly estrogens, have been shown to modulate melanocyte function
and melanin synthesis [31]. Inflammatory mediators, such as prostaglandins and
cytokines, can disrupt melanogenesis by altering the expression and activity of
melanogenic enzymes [9]. A neuropeptide hormone,
Intracellular melanin content correlates directly with the activity of
tyrosinase [17]. To begin assessing the impact of SHUE on
Many similar studies about natural extracts or compounds have shown that they
directly inhibit the catalytic activity of tyrosinase by regulating the molecular
structure [36, 37, 38]. However, the cell lysate contained tyrosinase as well as
TRP-1, TRP-2, and various other signaling molecules. Following the reveal of the
potential of SHUE to suppress tyrosinase activity in this study, we resolved to
investigate how SHUE regulates the expression of proteins involved in melanin
synthesis. Moreover, the mechanism behind its suppression of melanin production
was investigated. TRP-2, one of the enzymes involved in melanin synthesis,
catalyzes the reaction from DOPA chrome to 5,6-dihydroxyindole-2-carboxylic acid
(DHICA) [39]. Therefore, like tyrosinase, a decrease in the protein expression
level of TRP-2 is considered a cause of melanin production suppression. MITF is
the transcription factor of tyrosinase, TRP-1, and TRP-2; and it is known as the
most important transcription factor in melanin production. Protein expression
analyzed by western blotting revealed that the expression levels of tyrosinase,
TRP-1, and TRP-2 were significantly decreased by the SHUE treatment in stimulated
B16F10 murine melanocytes. Furthermore, the expression of MITF was also decreased
by the SHUE treatment in stimulated B16F10 murine melanocytes. Findings of the
study suggest that SHUE suppresses tyrosinase activity, based on the
downregulation of tyrosinase and MITF expression levels, resulting in the
suppression of melanin synthesis. Recent studies indicate that the ERK kinase
cascade and the AKT pathway play roles in regulating melanin synthesis. As
reported, they trigger MITF phosphorylation, which breaks it down and ultimately
reduces melanin production when ERK and AKT are activated [40]. Moreover, the
melanocyte differentiation program activates the MITF expression. Therefore, this
cascade is consistent with the idea that MITF is a master regulator of
melanogenesis. In addition, MITF is a nuclear mediator of Wnt signals during
melanocyte differentiation by inhibiting Glycogen synthase kinase 3
In summary, our study indicates that SHUE effectively inhibits melanin synthesis
and tyrosinase activity by targeting the ERK pathway, leading to the suppression
of MITF, tyrosinase, TRP-1, and TRP-2 in
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
All authors contributed to the conception and design of the research study. KGISK, AMKJ, CIK, and YSA performed the research. SJH, JYO, EAK, SHC and GA provided help and advice on the experiments. KGISK, AMKJ, SJH, and EAK performed data analysis, curation, and visualization. SHC confirmed the reproduction of the results. CIK, YSA, JYO, and GA provided funding and resources. CIK, YSA, and GA were responsible for overall project supervision. KGISK wrote the draft manuscript, and AMKJ, CIK, YSA, SJH, JYO and EAK contributed to its review and editing. SHC and GA conceived the manuscript and contributed to its review and editing. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects.
S. horneri was collected along the West coast of Jeju island. And sample was identified by the Biodiversity Research Institute in Jeju, South Korea (voucher specimen number is SH2017J005).
Not applicable.
This work was supported by the Technology Development Program (S3266086) funded by the Ministry of SMEs and Startups (MSS, Korea).
Given Dr. Seon-Heui Cha’s role as an editorial board member, Dr. Seon-Heui Cha was not involved in the peer review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Marcello Iriti. Mr. Chang-Ik Ko and Yong-Seok Ahn work researchers at Choung Ryong Fisheries Co., Ltd. as researchers and were involved ultrasonic extract preparation and provided it for the study. All responsibility for this article was delegated to Dr. Seon-Heui Cha and Dr. Ginnae Ahn. Other authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.