







1 Key Laboratory of Plant Resources Conservation and Utilization, College of Biological Resources and Environmental Sciences, Jishou University, 416000 Jishou, Hunan, China
2 College of Biology and Environmental Sciences, Jishou University, 416000 Jishou, Hunan, China
3 National and Local United Engineering Laboratory of Integrative Utilization of Eucommia ulmoides, 416000 Jishou, Hunan, China
Abstract
Background: Eucommia ulmoides Oliver is a unique high-quality
natural rubber tree species and rare medicinal tree species in China. The rapid
characterization of E. ulmoides gene function has been severely hampered
by the limitations of genetic transformation methods and breeding cycles. The
polyethylene glycol (PEG)-mediated protoplast transformation system is a
multifunctional and rapid tool for the analysis of functional genes in
vivo, but it has not been established in E. ulmoides. Methods:
In this study, a large number of highly active protoplasts were isolated from the
stems of E. ulmoides seedlings by enzymatic digestion, and green
fluorescent protein expression was facilitated using a PEG-mediated method.
Results: Optimal enzymatic digestion occurred when the enzyme was
digested for 10 h in an enzymatic solution containing 2.5% Cellulase R-10 (w/v),
0.6% Macerozyme R-10 (w/v), 2.5% pectinase (w/v), 0.5% hemicellulase (w/v),
and 0.6 mol/L mannitol. The active protoplast yield under this condition was 1.13
Keywords
- E. ulmoides
- stem
- protoplast isolation
- transient transformation
Eucommia ulmoides Oliver is in the family Eucommiaceae and is a relict
plant of the tertiary period [1]. It is distributed in the subtropical and
temperate areas of China (24°50
Historically, due to technological limitations, researchers have primarily concentrated on pharmacology [7, 8] and resource collection [9, 10] when studying and utilizing E. ulmoides, but have not searched for functional genes [11], which has severely limited the research of molecular breeding and related biological characteristics of E. ulmoides. With the advancement of omics technology and the availability of E. ulmoides genome and transcriptome resources, there is a growing interest in identifying E. ulmoides functional genes.
However, genetic transformation technology still suffers from limitations of the cost of time, labor, and economy. For instance, a particle bombardment method and Agrobacterium-mediated method can establish a stable genetic transformation system in E. ulmoides, but they are limited by expensive equipment, low transformation efficiency, difficult genetic transformation, challenging in vitro regeneration, the long time required, and low functional expression efficiency of genes. These factors hinder the high-throughput analyses of functional genes.
Transient expression of protoplasts is a general and convenient technique for functional gene analysis. Protoplasts can be obtained by mechanical isolation or enzymolysis. Protoplasts can be used for high-throughput analysis of subcellular localization, gene function, promoter activity, protein interactions, hybridization, and gene expression. Currently, protoplast transformation systems have been established in a variety of commercial crops, including rice [12], sugarcane [13], perennial ryegrass [14], freesia [15], orchid [16], and soybean [17], of which some results have been achieved in the protoplast isolation of a few woody economic plants such as cassava [18], Magnolia [19], camellia oleifera [20], and poplar [21], indicating that the establishment of an efficient protoplast isolation method is feasible in the study of gene function of woody plants.
To the best of our knowledge, establishment of a protoplast transient expression system of E. ulmoides has not been reported. As the model plant of natural rubber and woody medicinal plants, it is necessary to establish a transient expression system, which is of great significance for the study of functional genes of E. ulmoides.
The aim of this study was to establish an efficient and simple protoplast isolation and transient transformation system for E. ulmoides. Efficient protoplasts were obtained by optimizing the factors that affect the yield and activity of protoplasts including the concentration of mannitol, enzymolysis time, and combination of enzymes. Furthermore, using green fluorescent protein (GFP) as the reporter gene, we verified the feasibility of the polyethylene glycol (PEG)-mediated protoplast transient expression system of E. ulmoides.
The results showed that the system is efficient and convenient, and can be used as a reference for future gene function studies of medicinal plants and hard rubber plants.
The stems of E. ulmoides seedlings were selected for protoplast isolation in this study. Whole E. ulmoides seeds were soaked in 400 mg/L gibberellic acid 3 solution overnight. Then the seeds were transferred to a petri dish covered with wet absorbent paper, and grown in a dark incubator at 25 °C for 3 days. The budding E. ulmoides seeds were transferred to loose soil for 1 week of seedling cultivation. The E. ulmoides seedlings grew in a growth cabinet (RTOP-310Y; Zhejiang TOP Cloud-Agri Technology Co., Ltd., Zhejiang, China) at a temperature of 26/22 °C (day/night) and a 16/8 h light-dark cycle.
The Escherichia coli DH5
One-week-old E. ulmoides seedlings collected were rinsed in pure water
until the surface was clean, and only the stems of seedlings were used as
material for protoplast isolation. Approximately 0.3 g stems were weighed and cut
into 0.2–0.4 mm stem segments using a sharp razor. Then all stem segments were
immediately transferred to a 3 mL enzyme solution (10 mmol/L MES
[Macklin, Shanghai, China] KOH, pH 5.7, 0.22 mmol/L KH
The concentration of protoplasts was calculated with a hemocytometer (XB.K.25;
QiuJing, Shanghai, China). Protoplast viability was measured by staining with
Evans blue. Specifically, protoplasts and 0.25% Evans blue staining solution
(Phygene, Fuzhou, China) were mixed in a 1:1 ratio and then incubated for 5 min
at room temperature in the dark. Living protoplasts (unstained) and dead
protoplasts (blue) were visualized with an optical microscope. Protoplast
viability (%) = number of unstained protoplasts in view/total number of
protoplasts in view
The harvested protoplasts were resuspended in MMG solution (0.6 mol/L mannitol,
15 mmol/L MgCl
To establish a transient transformation system for high-throughput functional gene analysis in E. ulmoides, the selection of appropriate materials to obtain high-yielding and high-viability protoplasts is a crucial factor. Since woody plants contain high lignin, we selected stems of 1-week-old birth E. ulmoides seedlings as materials to assess the effect of protoplast isolation.
To obtain high quality protoplasts in good condition, different concentrations
of mannitol were added to the enzyme solution containing 2.5% Cellulase R-10
(w/v), 0.6% Macerozyme R-10 (w/v), 2.5% pectinase (w/v), and 1.5%
hemicellulase (w/v) to explore the effect of osmotic pressure on the quantity and
quality of protoplasts. After 4 h of enzymatic digestion, the total number of
protoplasts, number of viable protoplasts, and the survival rate were counted
(Fig. 1). Significantly, total protoplast yield (19.80
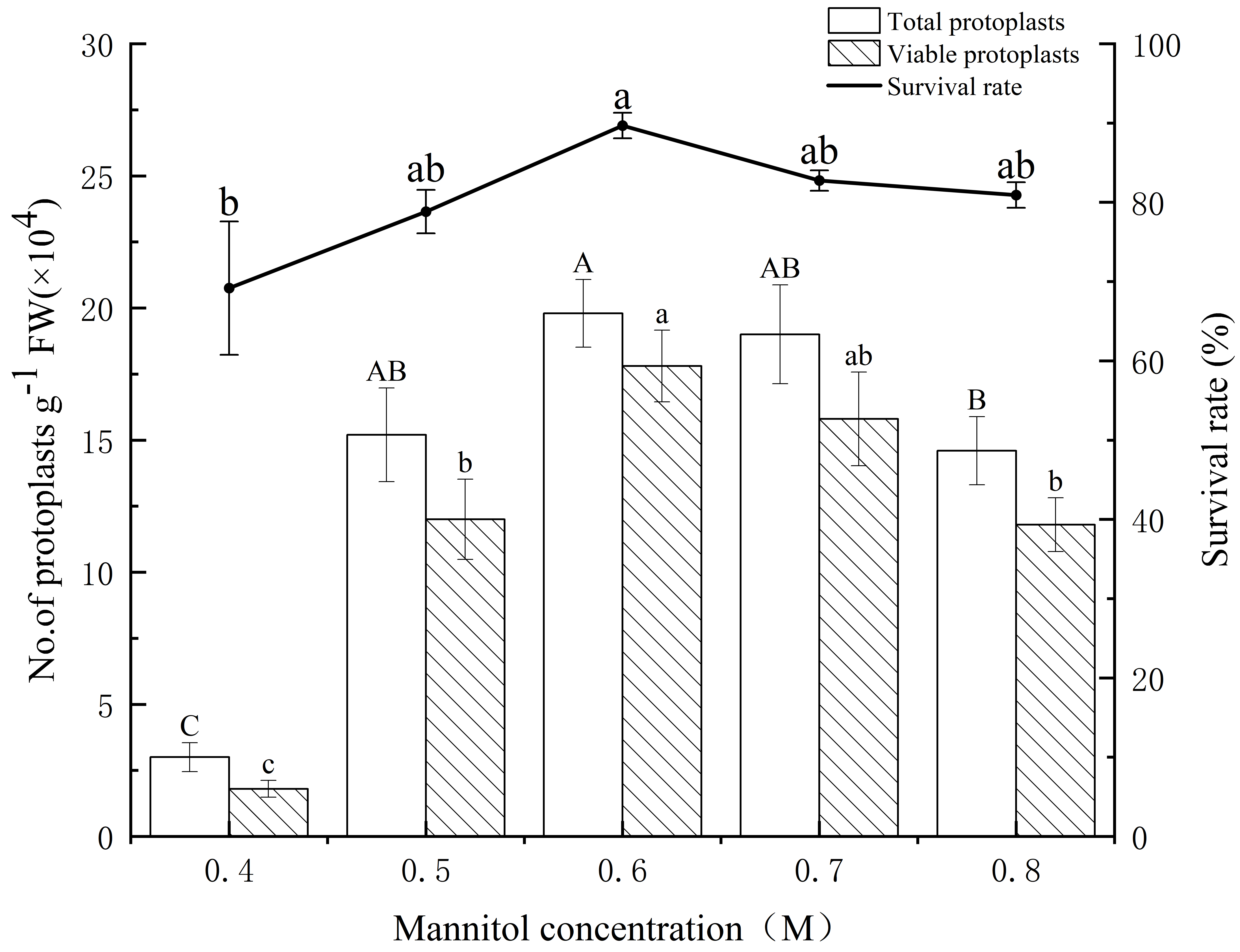 Fig. 1.
Fig. 1.Yield and survival rate of isolated protoplasts of E.
ulmoides at different mannitol concentrations. The white and patterned bars
indicate total yield of protoplasts and yield of viable protoplasts,
respectively. The black solid line indicates the survival rate. Values indicate
the mean
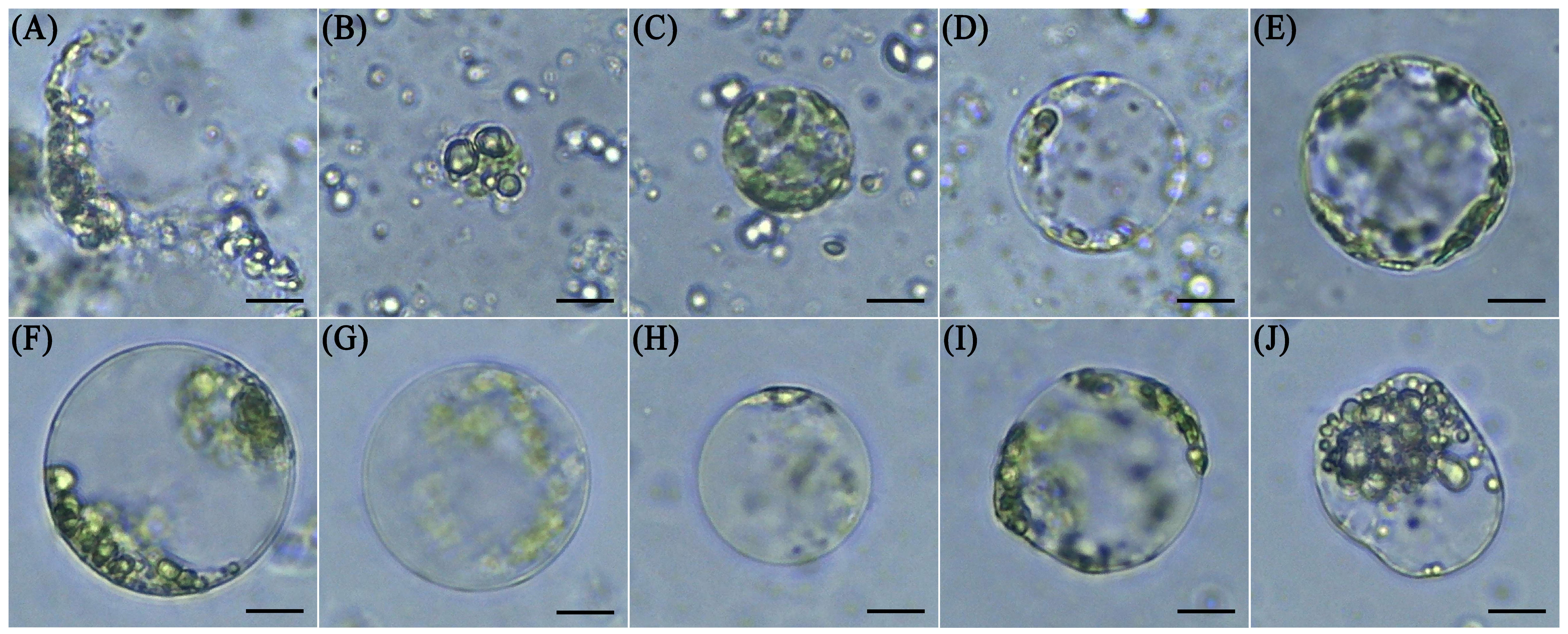 Fig. 2.
Fig. 2.Characterization of protoplasts in mannitol concentrations of 0.4 mol/L to 0.8 mol/L. Protoplast states in 0.4 mol/L mannitol (A,B). Protoplast states in 0.5 mol/L mannitol (C,D). Protoplast states in 0.6 mol/L mannitol (E,F). Protoplast states in 0.7 mol/L mannitol (G,H). Protoplast states in 0.8 mol/L mannitol (I,J). Scale bar, 15 µm.
Next, we analyzed the effect of enzymatic digestion time (1–14 h) on the
isolation efficiency of protoplasts from the stems of E. ulmoides
seedlings. The experimental data (Fig. 3) showed that the total yield of
protoplasts began to significantly increase in the 4th hour of enzymatic
digestion and peaked at 10 h (92.80
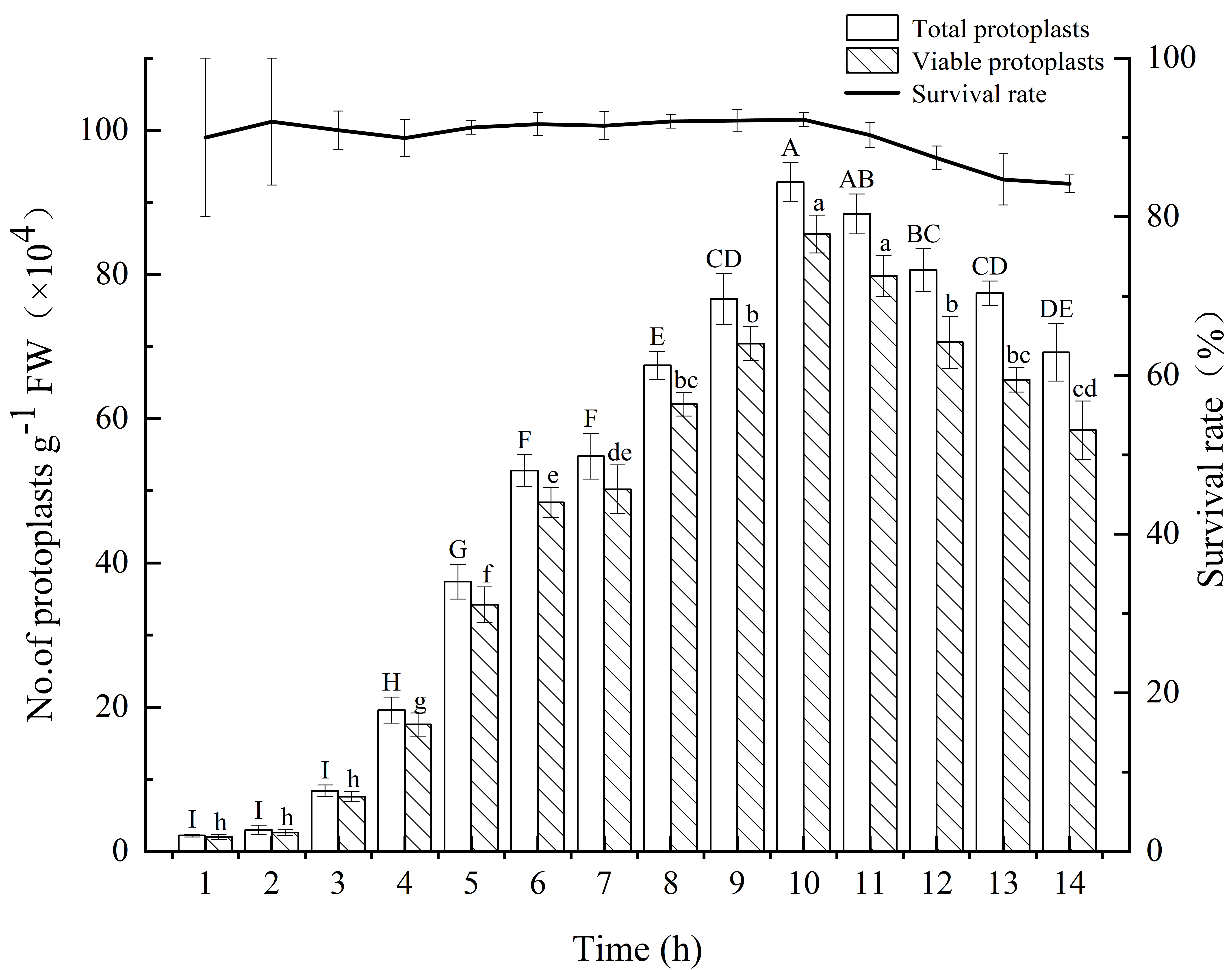 Fig. 3.
Fig. 3.Yield and survival rate of isolated protoplasts from E.
ulmoides for different enzymatic digestion times. The white and patterned bars
indicate total yield of protoplasts and yield of viable protoplasts,
respectively. The black solid line indicates the survival rate. Values indicate
the mean
For the isolation of protoplasts, it is often not the most appropriate approach
to obtain the optimal level of each factor separately by simply superimposing
one-way analysis of variance. Therefore, we selected four enzymes, each of which
had three concentration levels (Table 1), and performed a total of nine
treatments with an orthogonal experimental design, as shown in Table 2. It can be
seen that the highest yield of protoplasts (12.00
| Combination | Cellulase R-10 (%) | Pectinase (%) | Macerozyme R-10 (%) | Hemicellulase (%) |
| 1 | 1.5 | 1.5 | 0.3 | 0.5 |
| 2 | 2.5 | 2.5 | 0.6 | 1.5 |
| 3 | 3.5 | 3.5 | 0.9 | 2.5 |
| Treatment No. | Cellulase R-10 (%) | Pectinase (%) | Macerozyme R-10 (%) | Hemicellulase (%) | Protoplast yield (×10 |
Active protoplast yield (×10 |
Protoplast viability (%) |
| 1 | 1 | 1 | 1 | 1 | 4.00 | 3.33 | 85.00% |
| 2 | 1 | 2 | 3 | 3 | 7.00 | 6.67 | 95.83% |
| 3 | 1 | 3 | 2 | 2 | 8.33 | 7.67 | 92.13% |
| 4 | 2 | 1 | 3 | 2 | 9.00 | 8.67 | 96.30% |
| 5 | 2 | 2 | 2 | 1 | 12.00 | 11.33 | 94.84% |
| 6 | 2 | 3 | 1 | 3 | 5.33 | 4.67 | 87.78% |
| 7 | 3 | 1 | 2 | 3 | 5.33 | 4.67 | 87.78% |
| 8 | 3 | 2 | 1 | 2 | 4.00 | 3.67 | 93.33% |
| 9 | 3 | 3 | 3 | 1 | 5.67 | 5.00 | 87.78% |
| Protoplast yield | K1 | 19.33 | 18.33 | 13.33 | 21.67 | ||
| K2 | 26.33 | 23.00 | 25.67 | 21.33 | |||
| K3 | 15.00 | 19.33 | 21.67 | 17.67 | |||
| Range | 11.33 | 4.67 | 12.33 | 4.00 | |||
| Rank | Macerozyme R-10 |
||||||
| Active protoplast yield | K1 | 17.67 | 16.67 | 11.67 | 19.67 | ||
| K2 | 24.67 | 21.67 | 23.67 | 20.00 | |||
| K3 | 13.33 | 17.33 | 20.33 | 16.00 | |||
| Range | 11.33 | 5.00 | 12.00 | 4.00 | |||
| Rank | Macerozyme R-10 |
||||||
| Protoplast viability | K1 | 2.88 | 2.75 | 2.60 | 2.68 | ||
| K2 | 2.76 | 2.72 | 2.83 | 2.82 | |||
| K3 | 2.60 | 2.72 | 2.76 | 2.71 | |||
| Range | 0.28 | 0.03 | 0.23 | 0.14 | |||
| Rank | Cellulase R-10 |
||||||
| Category | Protoplasts yield | Active protoplasts yield | Protoplast viability |
| Protoplast yield | 1 | ||
| Active protoplast yield | 0.998 | 1 | |
| Protoplast viability | 0.711 | 0.752 | 1 |
Overall, combined with the above optimized parameters, a large amount of protoplasts could be efficiently isolated from the stems of E. ulmoides seedlings. That is, the concentrations of Cellulase R-10, Macerozyme R-10, pectinase, hemicellulase, and mannitol in the enzymatic digestion solution were 2.5%, 0.6%, 2.5%, 0.5%, and 0.6 mol/L, respectively. The enzymatic digestion time was 10 h. The vast majority of protoplasts harvested by centrifugal precipitation were round (Fig. 4A), and Evans blue staining showed a high viability of protoplasts (Fig. 4B).
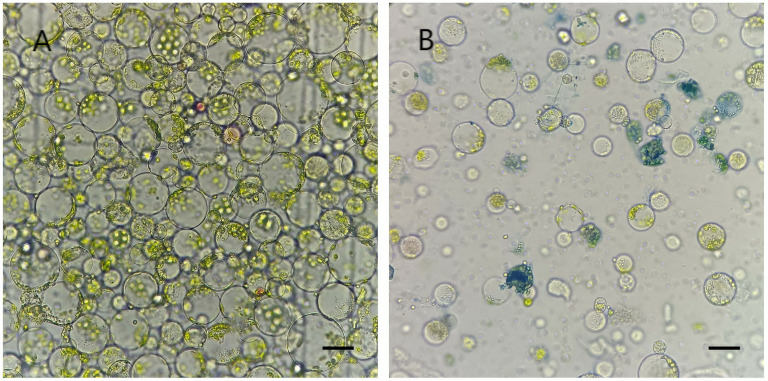 Fig. 4.
Fig. 4.Isolated protoplasts from the stems of E. ulmoides seedlings. (A) Morphological features of protoplasts observed under a light microscope. (B) Evans blue-stained inactive protoplasts under a light microscope. Scale bar, 20 µm.
The feasibility of PEG-mediated transfection of the pCAMBIA1303-mGFP5 plasmid into E. ulmoides was initially explored. The plasmid encoded mGFP under the control of CaMV35S promoter. The pCAMBIA1303-mGFP5 was transfected at 25 °C in the dark in 35% PEG4000. GFP was detected in the cytoplasm with an obvious distribution (Fig. 5). Meanwhile, the effects of different transfection times on transfection efficiency were explored, and the results showed that the transformation efficiency was as high as 35.01% after 15 min of transfection (Fig. 6).
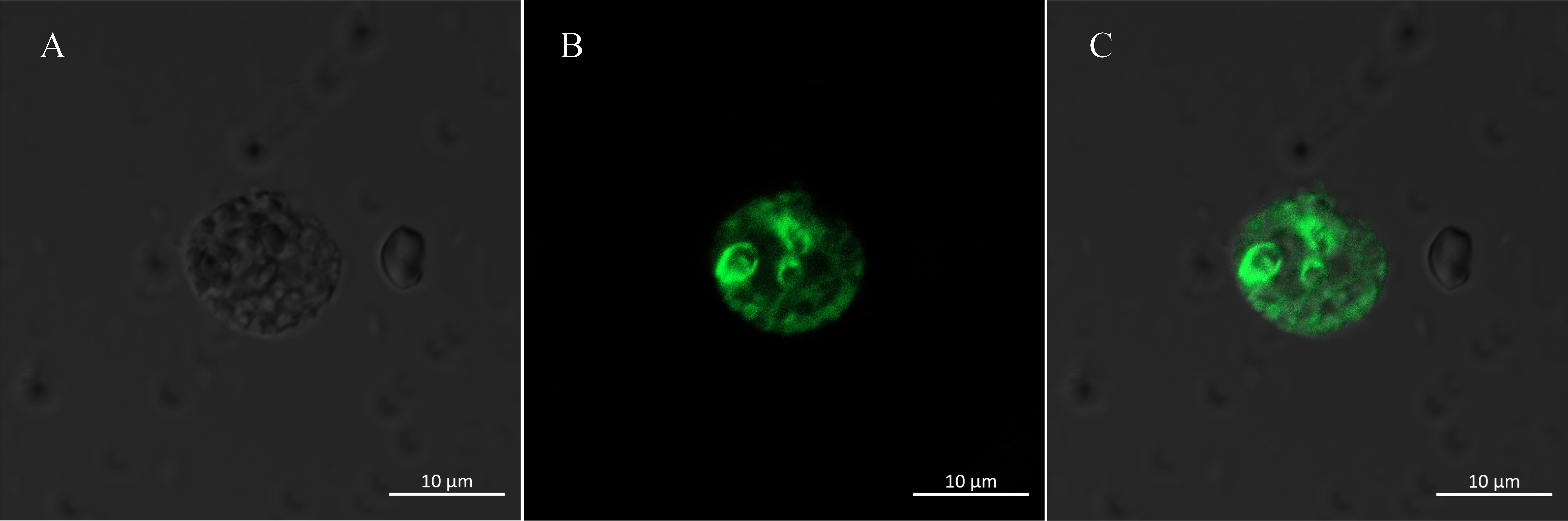 Fig. 5.
Fig. 5.pCAMBIA1303-mGFP5 distribution in the protoplasts of E. ulmoides. (A) mGFP fluorescence image. (B) Bright field image. (C) Merged field image. Scale bar, 10 µm.
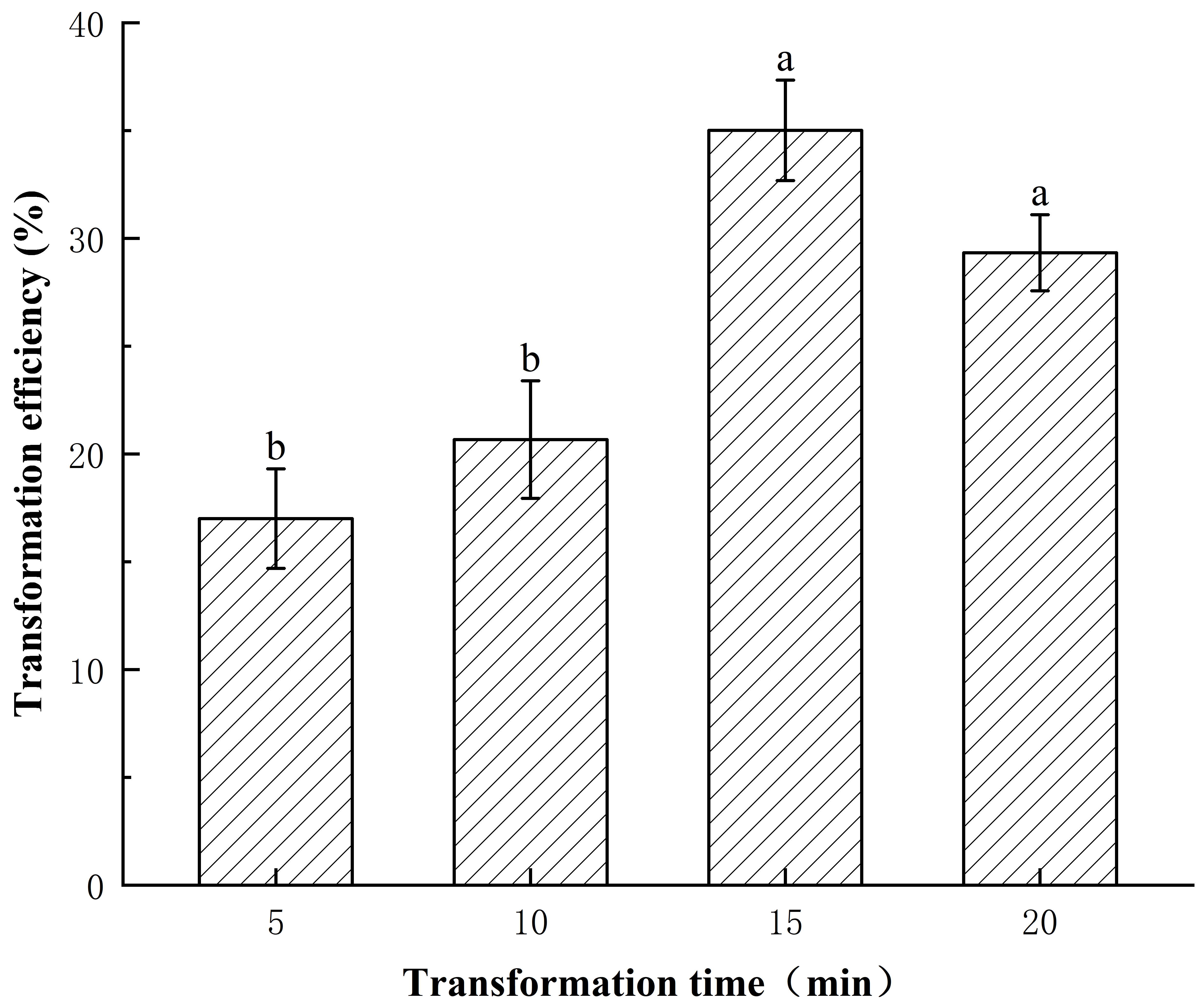 Fig. 6.
Fig. 6.Transfection efficiency of protoplasts of E. ulmoides
at different transfection times. Transfection efficiency was calculated after 15
h of cultivation. Values indicate the mean
Plant protoplasts can be isolated from plants’ specific tissues or organs and can efficiently provide multiple independent single cell systems to solve specific problems because they retain their cellular properties [16]. It was once thought that the source tissues suitable for protoplast isolation were derived from non-seedling plants [22]. However, several studies in recent years have indicated that protoplasts can be isolated from young plant organs such as hypocotyls, leaves, roots, and root hairs [23], which provides a reference idea for the isolation of protoplasts from plants of different species.
The establishment of a stable protoplast isolation system is a prerequisite for generating a large number of protoplasts with high viability. The isolation of protoplasts is typically influenced by several factors including material selection, enzyme type and concentration, osmotic pressure, and enzymatic digestion time. Although efficient protoplast isolation systems have been reported in several herbaceous plants in recent years, isolation of sufficient amounts of live protoplasts in woody plants is still a challenge. This is especially true for Eucommia, which has great potential for exploitation because the whole plant, except the xylem, is rich in Eucommia-rubber (hard rubber), which is a major challenge for the isolation of E. ulmoides protoplasts.
Experiments have been reported on the selection of materials from plants from the outdoors that can affect the efficiency of protoplast isolation due to their thick cuticle and thick cell walls [24], but the preparation and supply of test tube seedlings often require skillful aseptic manipulation, economic cost, and time cost. With the aim of convenience, economy, and efficiency, we used stems from 1-week-old live seedlings of E. ulmoides cultured at a simulated room temperature, which were not lignified and contained less Eucommia rubber to conduct the study.
Free protoplasts are fragile when they have no cell wall to maintain their shape, and this fragility is reflected in the inability of protoplasts to remain isotonic with the external environment, resulting in swell, burst, or shrinkage of protoplasts. The addition of an appropriate concentration of osmotic stabilizer to the enzymatic solution can maintain the equilibrium of osmotic pressure in and out of the protoplasts. Most current protoplast isolation systems add mannitol and sorbitol to maintain the osmotic pressure of free protoplasts. The appropriate concentration of the osmotic stabilizer depends on the properties of the material itself. The results of several studies showed that appropriate addition of mannitol in the range of 0.4 mol/L to –0.8 mol/L can maintain the shape of protoplasts for a long time, depending on the material. Our results indicated that both hypoosmolarity and hyperosmolarity were detrimental to isolation of protoplasts from the stems of E. ulmoides seedlings. When the mannitol concentration was 0.4 mol/L to –0.5 mol/L, most of the protoplasts swelled and ruptured. When the mannitol concentration was greater than 0.6 mol/L, some of the protoplasts shrunk, which was not conducive to the progress of transient transformation. Therefore, the addition of 0.6 mol/L mannitol was determined to be the optimum concentration for protoplast isolation from the stems of E. ulmoides seedlings.
Enzymatic digestion time is one of the important factors in obtaining protoplast yield and viability, which varies according to different tissues and organs. When protoplasts were isolated from the leaf mesophyll of Catalpa bungei, the enzymatic digestion time was too short, resulting in inadequate enzymatic digestion of protoplasts, and the yield could not meet the need of instantaneous transformation, whereas excessive enzymatic digestion time could lead to a decrease of protoplast activity [23]. The experimental results suggested that the protoplast total yield and live protoplast yield showed a stepwise increase within 10 h, and peaked at 10 h. Therefore, enzymatic digestion time of 10 h is appropriate. Unexpectedly, we found that even though there was a decreasing trend in protoplast total yield and live protoplast yield after 10 h, the decrease in viability was not significant. Based on the comparison of previous methods with this study, it was speculated that the survival rate of protoplasts might be mainly influenced by the following factors: (i) the oscillation rate during enzymatic digestion; (ii) the state of the experimental material; and (iii) the composition of inorganic salts in the enzymatic solution.
The type and concentration of enzymes directly affect the preparation of
protoplasts. Cellulases, pectinases, macerozymes, and hemicellulases are commonly
used for the degradation of cell walls, and the chemical composition of cell
walls varies depending on the species and material sources, so the appropriate
enzyme combinations and enzyme concentrations for efficient preparation of
protoplasts is not uniform. Protoplast yields of 6.5
Establishing a stable genetic transformation system is an important constraint for genetic improvement of woody plants. Protoplast-based transient expression is a rapid, economical, and efficient alternative system compared to the traditional stable expression of transgenes, and is important for the analysis of plant gene function. This system has proven to be a powerful experimental tool in molecular biology for a wide range of applications such as subcellular localization [27], promoter activity analysis [28], protein–protein interactions [29], protein–DNA interactions [30], protein-based biochemical assays [31] and quantitative real-time polymerase chain reaction (QRT-PCR) for gene expression analysis.
In this study, a PEG-mediated transient protoplast transformation method based on the protoplast isolation system was established for stems of E. ulmoides seedlings, using mGFP as a reporter gene, and the feasibility of the system was verified. At the same time, the effect of transformation time on transformation efficiency was initially investigated by taking the transformation time as an independent variable. Notably, even though 15 min in 35% PEG solution was beneficial for transformation efficiency (up to 35.01%), similar to the results of other studies, there are still optimization factors in our transformation system and the efficiency may be further improved.
In this study, we utilized the stems of E. ulmoides seedlings as the
material and systematically investigated 9 enzyme digestion solutions, 5 mannitol
concentrations, and 14 durations, leading to the development of an optimized
protocol for efficient isolation of E. ulmoides protoplasts. The
optimized enzyme solution, composed of 2.5% Cellulase R-10 (w/v), 0.6%
Macerozyme R-10 (w/v), 2.5% pectinase (w/v), 0.5% hemicellulase (w/v), and 0.6
mol/L mannitol, enabled the generation of 1.13
All data points generated or analyzed during this study are included in this article and there are no further underlying data necessary to reproduce the results. The raw data are available from the corresponding author on reasonable request.
Conceptualization—BH, MD and QZ; Data acquision—BH, RL, WS and JP; Formal analysis—YW, LM and YD; Methodology—BH, CW and QZ; Writing original draft—BH; Corresponding—QZ. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Eucommia ulmoides (Jishou University, Hunan, China) plants were used in this study. The plant materials were provided by Prof. Qiang Zhou (Jishou University, Hunan, China).
We are grateful to the Institute of Botany, the Chinese Academy of Sciences for supporting this work.
This research was funded by National Natural Science Foundation of China under Grant [32160388].
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.