

1 Head and Neck Surgery, Yunnan Cancer Hospital, 650118 Kunming, Yunnan, China
2 Medical School, Kunming University of Science and Technology, 650500 Kunming, Yunnan, China
†These authors contributed equally.
Abstract
At present, many problems remain to be solved in studying the pathogenesis of thyroid cancer. Ferroptosis is a programmed cell death mode discovered in recent years, and many studies have found that ferroptosis plays a significant role in the prognosis and progression of thyroid cancer. The researchers showed that ferroptosis-related genes are essential in diagnosing thyroid cancer. Therefore, this paper summarizes some pathological and clinical characteristics of thyroid cancer and makes a series of combs on the relationship between ferroptosis and the basis and function of thyroid cancer, thus providing specific ideas for the diagnosis and treatment of thyroid cancer.
Keywords
- ferroptosis
- thyroid cancer
- prognosis
- progression
- ferroptosis-related genes
Thyroid carcinoma includes four main types: papillary carcinoma, follicular carcinoma, medullary carcinoma, and undifferentiated carcinoma. Thyroid cancer is one of the most common malignant tumors, accounting for about 1% of all solid malignant tumors [1]. The incidence rate is mainly female, which has jumped to the top three in recent years. The mortality rate is relatively low while the incidence rate is high, showing that most thyroid cancers are inert [2, 3, 4]. Due to the high incidence of thyroid cancer, the incidence of cancer in adolescents and young people (AYA) has increased significantly in the last ten years [5]. Due to the high incidence of differentiated thyroid cancer, there is an urgent need for more effective early diagnosis tools and treatment strategies for this disease. Recent research shows that circular RNA (circRNA) plays a vital role in the occurrence and development of cancer [6]. CircRNAs can play their role through various mechanisms, the most common of which is that it affects the expression of target genes through sponge microRNAs (miRNAs). At the same time, other functions also include regulating its activity and transcription of host genes through interaction with protein [7, 8, 9]. The imbalance of circRNA is related to the occurrence and development of many diseases, especially cancer [10]. In this review, we briefly summarized the relationship between thyroid cancer and ferroptosis-related genes. Then, we discussed the potential role of ferroptosis-related genes as diagnostic and prognostic markers. At the same time, we also summarize the role of ferroptosis-related genes in promoting or inhibiting the development of thyroid cancer. Finally, we put forward some future research directions in this field.
Thyroid cancer is a malignant tumor of the endocrine system, mainly from thyroid follicular cells and a few from parafollicular cells [11]. The thyroid gland secretes various hormones, including thyroid hormone, which has a robust regulatory function on the body’s metabolism. Thyroid cancer can be divided into differentiated thyroid cancer and undifferentiated thyroid cancer. Its pathological types include papillary cancer, follicular cancer, medullary cancer, and undifferentiated cancer, among which differentiated papillary cancer is the most common, accounting for more than 80% of all thyroid cancers [12]. Followed by medullary carcinoma developed from thyroid follicular cells, accounting for about 10% of all thyroid cancers [13]. Undifferentiated carcinoma (UC) is a type of thyroid cancer with a poor prognosis. It develops rapidly, with cervical lymph node metastasis in about 50% of early stage, and distant metastasis often occurs through the blood supply, with high mortality [14].
According to the staging of thyroid cancer by the American Association for Cancer Research (AARC), the staging indicators include age, and tumor infiltration degree. Among them, the age of 55 is an essential factor affecting the staging, and the progress of thyroid cancer is divided into four stages (I to IV). As cancer progresses from stage I to stage IV, the survival rate of patients will decrease. The main feature of stage I and stage II is that the tumor cells are confined in the thyroid gland, but the maximum diameter of the tumor is different. The main feature of stage III is that tumor cells spread outside the thyroid gland and invade. Stage IV is defined as distant metastasis (for example, lung, bone, and other organs, and tumors can also occur around the carotid artery).
In recent years, with the continuous improvement of ultrasonic imaging technology for thyroid cancer, the sensitivity of various indicators in laboratory diagnosis, such as Thyroglobulin Antibody (TgAb), Thyroid Stimulating Hormone Receptor Antibody (TRAb), and the development of Convolutional Neural Networks (CNNs) and Dimensionality Reduction Techniques (DRTs) technology and Classification Regularized Uniform Manifold Approximation and Projection (CReUMAP) embedding have high accuracy in judging thyroid nodules, which leads to an increase in the screening rate of thyroid cancer and an apparent upward trend in the incidence and diagnosis rate [15, 16]. There are many treatments for thyroid cancer (such as surgery, radionuclide therapy, TSH suppression therapy, and external radiation therapy), among which surgery is the primary treatment, and most patients have a good prognosis after surgery [17]. Although most thyroid cancer is an inert tumor, it is also prone to cervical lymph node metastasis. About 30%–80% of patients with papillary thyroid cancer have cervical lymph node metastasis. Lymph nodes in the central region (region VI) are the most prone to metastasis, which can reach about 33% of all patients [18, 19, 20]. Patients with advanced thyroid cancer can try to use neoadjuvant therapy and other measures.
Unlike other types of cell death, such as apoptosis, pyroptosis, and autophagy, ferroptosis is an iron-dependent cell death characteristic of lipid peroxidation [21]. Ferroptosis is involved in the progression of many types of cancers, such as non-small cell lung cancer (NSCLC), colorectal cancer (CRC), gastric cancer (GC), and thyroid cancer (TC), and also affects the immunotherapeutic effects [22, 23, 24, 25, 26]. Erastin and orafenib cause ferroptosis due to glutathione depletion in the antioxidant defense system. Ferroptosis-related Genes (FRGs) are co-regulated in thyroid cancer, including APOE, BCL3, ALOX5AP, lncRNAl, and variant 4 (ETV4). In addition, there is a strong relationship between ferroptosis and the regulation of FRGs expression, which will be discussed individually in the following developments. Ferroptosis is roughly divided into several parts in regulating thyroid cancer, including activation of iron-containing enzymes by excess iron, lipid peroxidation in polyunsaturated fatty acids, and defense disorder of an antioxidant system [27, 28]. Therefore, we reviewed the progress on biological functions and clinical implications of ferroptosis in thyroid cancer.
Ferroptosis plays a vital role in the prognosis of thyroid cancer, and the FRGs found in the current study are closely related to the prognosis of thyroid cancer. Based on the expression data of FRGs in the thyroid cancer dataset from The Cancer Genome Atlas (TCGA), two subtypes, cluster 1 and cluster 2, were identified. The proportion of FRGs that positively associated with ferroptosis in cluster 2 was higher than that in cluster 1. On the contrary, the proportion of FRGs that negatively correlated with ferroptosis in cluster 1 was higher than that in cluster 2. Nras and Braf were identified as the predominant mutations in cluster 2 and cluster 1, respectively (Tables 1,2, Ref. [29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46]) [29]. Based on the thyroid cancer dataset from TCGA, a 10-FRG signature including TXNRD1, TFRC, TF, SRXN1, PGD, ISCU, DRD4, DPP4, CDKN2A, and ANGPTL7 was identified, and this 10-FRG signature was an independent prognostic factor for both overall survival (OS) and disease-free survival (DFS). After analyzing the thyroid cancer dataset from TCGA, high expression of AKR1C3 and low expression levels of BID, FBXW7, GPX4, and MAP3K5 were found to be positively correlated with unfavorable outcomes in thyroid cancer patients, respectively. Moreover, these genes were also the independent prognosis factors (Tables 1,2) [30]. Except for TFRC, nine other genes of the 10-FRG signature were similar to TCGA data and differentially expressed in an independent cohort of thyroid cancer (n = 75). Quite significantly, sorafenib, a ferroptosis inducer and clinical drug for thyroid cancer, reduced the cell viability of thyroid cancer cells and changed most of the 10-FRG signature except PGD (Tables 1,2) [31]. Huang et al. [32] conducted a prognosis-predictive model including HSPA5, ferroptosis AURKA, and TSC22D3 based on FRGs, and this model was an independent prognosis prediction factor of progression-free survival (Tables 1,2). Eight FRGs were identified as an independent prognostic risk model in thyroid cancer, and the eight FRGs were closely associated with immune-related biological processes [47].
| CircRNA (CircBase ID) | Detection Methods, Cohort Size/Subtype | Sample Source | Prognosis | Diagnosis | Refs |
| FGRS | qRT-PCR tomor tissues (n = 502), normal tissues (n = 56) | Tissues | – | – | [29] |
| FGRS (AKR1C3) | qRT-PCR tomor tissues (n = 75), normal thyroid tissues | Tissues | + | – | [30] |
| FGRs | qRT-PCR tomor tissues (n = 75), normal tissues (n = 75) | Tissues | + | – | [31] |
| FRGs | qRT-PCR PTC samples (n = 501), normal samples | – | PFS | – | [32] |
| ALOX5AP-rs4076128 | SNP qRT-PCR papillary thyroid carcinoma (n = 520), normal thyroid tissues (n = 520) | Tissues | – | – | [33] |
| BCL3-s34698726 | |||||
| BCL3-rs8100239 | |||||
| circ-0067934 | qRT-PCR tomor tissues, normal tissues | Tissues | – | – | [34] |
| Neferine | western blot, qRT-PCR | Tissues, Serum | – | + | [35] |
| FTO | qRT-PCR PTC tissues, normal tissues | Tissues | – | – | [36] |
| ALKBH5 | qRT-PCR tomor tissues,normal thyroid tissues (mRNA protein levels) | Tissues | – | – | [37] |
| Circ KIF4A | qRT-PCR tomor tissues (n = 52,070), normal thyroid tissues | Tissues | + | – | [38] |
| lncRNA AC034213.1 | qRT-PCR tomor tissues (n = 502), normal tissues | Tissues | + | – | [39] |
| PR-DE-IRFeGs | qRT-PCR THCA tissues, adjacent normal thyroid tissues | Tissues | OS+ | – | [40] |
| CERS6-AS1, LASP1 | qRT-PCR tomor tissues, normal tissues | Tissues | + | – | [41] |
| miR-497-5p | |||||
| FRGs | qRT-PCR tomor tissues (n = 75), adjacent normal tissues (78.8%) | Tissues | + | – | [42] |
| ETV4 | qRT-PCR PTC tissues (n = 30), adjacent normal tissues (n = 30) | Tissues | – | – | [43] |
| FRGs | RT-qPCR tomor tissues, normal tissues (n = 45) | Tissues | + | – | [44] |
| FGRs | qRT-PCR tomor tissues (n = 510), normal thyroid tissues (n = 58) | Tissues | + | – | [45] |
| Vitamin C | western blot, qRT-PCR | Tissues | – | + | [46] |
+ represents that they are related to prognosis or diagnosis; – represents that they are not; FGRs, ferroptosis-related genes; PR-DE-IRFeGs, prognosis-related differentially expressed immune-related ferroptosis genes; ALKBH5, alkB homolog 5; CERS6-AS1, ceramide synthase 6 antisense RNA 1; LASP1, lim and SH3 protein1; FTO, fat mass and obesity-associated protein; OS, overall survival; FRGs, Ferroptosis-related Genes; Circ KIF4A, Circular Transcripts of KIF4A; ETV4, E26 Transformation Specific (ETS) Variant 4; SNP, Single Nucleotide Polymorphism; qRT-PCR, Quantitative reverse transcription polymerase chain reaction; PTC, Papillary thyroid cancer; THCA, Thyroid Carcinoma; RT-qPCR, Real-time Quantitive PCR; PFS, Progression-free Survival.
| CircRNA (CircBase ID) | Functions | Molecular Mechanisms | Refs |
| FGRs | in vitro: ↑ tumor growth; in vivo: ↑ prolifera | hub gengs/Braf and Nras mutations | [29] |
| FGRS (AKR1C3) | in vivo: ↑ prolifera/invasion/migration abilities | AKR1C3 | [30] |
| FGRs | in vitro: ↑ tumor growth | – | [31] |
| FRGs | in vivo: ↑ immune cell infiltration (TMB/TME/HLA) | – | [32] |
| ALOX5AP-rs4076128 | in vivo: ↑ susceptibility/genetic polymorphisms | APOE/BCL-3/ALOX5AP | [33] |
| BCL3-s34698726 | |||
| BCL3-rs8100239 | |||
| circ-0067934 | in vivo: ↓ prolifera | miR-545-3p/SLC7A11 | [34] |
| Neferine | in vivo: ↑ prolifera/invasion | Nrf2/HO-1/NQO1 | [35] |
| FTO | in vivo: ↓ tumor progrssion | m6A-TIAM1-Nrf2/HO-1 | [36] |
| ALKBH5 | in vivo: ↓ tumor progrssion | m6A-TIAM1-Nrf2/HO-1 | [37] |
| Circ KIF4A | in vitro: ↓ tumor growth | Circ KIF4A-miR-1231-GPX4 | [38] |
| lncRNA AC034213.1 | over survive/tomur purity/immune checkpoint blockade | CTLA4/PD-L1 | [39] |
| PR-DE-IRFeGs | in vitro: ↑ tumor progression | BRAF/NRAS/HRAS mutation | [40] |
| CERS6-AS1, LASP1 | in vivo: ↓ tumor cell viablity | miR-497-5P/LASP1 | [41] |
| miR-497-5p | |||
| FRGs | in vitro: ↑ prognostic survival | – | [42] |
| ETV4 | in vitro: ↑ cell proliferation, cell cycle progression; in vivo: ↑ tumor growth | SLC7A11 | [43] |
| FRGs | in vitro: ↑ prognosis; in vivo: ↑ immune status | – | [44] |
| FGRs | – | – | [45] |
| Vitamin C | in vitro: ↑ ATC cells, GPX4 inactivation, ROS accumulation | GPX4 | [46] |
ROS, reactive oxygen species; TMB, Tumor Mutation Burden; TME, Tumor Microenvironment; HLA, Human Leukocyte Antigen; ATC, Anaplastic Thyroid Cancer; GPX4, Glutathione Peroxidase 4; AKR1C3, Aldo‐keto Reductase Family 1 Member C3; APOE/BCL-3/ALOX5AP, Apolipoprotein E / BCL3 Transcription Coactivator/ Arachidonate 5-Lipoxygenase Activating Protein; SLC7A11, The Solute Carrier Family 7 Member 11; Nrf2/HO-1/NQO1, NF-E2-related Factor2/Heme Oxygenase 1/NAD(P)H Quinone Dehydrogenase 1; TIAM1, TIAM Rac1 Associated GEF 1; CTLA4/PD-L1, Cytotoxic T-lymphocyte Associated Protein 4/Programmed Death 1; BRAF/NRAS/HRAS, B-Raf Proto-oncogene, Serine/Threonine Kinase/NRAS Proto-oncogene, GTPase/HRas Proto-oncogene, GTPase.
In recent years, discoveries have been made in the progression of ferroptosis in thyroid cancer, mainly focusing on the differential expression of intermediate hormone levels and receptor activity between thyroid cancer tissues and normal tissues. After analyzing the thyroid cancer dataset from TCGA, 176 of 259 were differentially expressed in thyroid tumor tissues compared with adjacent normal tissues, and these differentially expressed genes were enriched in thyroid hormone, lipid metabolism, and activation of receptors in Gene Ontology (GO) analysis and enriched in thyroid hormone synthesis and Peroxisome Proliferator-activated Receptors (PPAR) signaling in Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis [31]. After performing the GO and KEGG pathway analyses in the thyroid cancer dataset from TCGA, immune cell infiltration, HLA, TME, TMB, and ssGSEA were identified to be markedly correlated with ferroptosis [32]. Different genes also have different associations with thyroid cancer risk. Polymorphisms of FRGs, including APOE, BCL3, and ALOX5AP, were investigated, and the results showed that BCL3-s34698726 and rs8100239 were significantly correlated with an increased risk of thyroid cancer. At the same time, ALOX5AP-rs4076128 was associated with a decreased risk of thyroid cancer (Tables 1,2) [33].
In comparison between thyroid cancer tissue and normal tissue, upregulated or
down-regulated target genes in vivo and in vitro inhibited iron death in thyroid
cancer cells to varying degrees. Compared with normal thyroid follicular
epithelial Nthy-ori 3-1 cells, cicr_0067934 was overexpressed in thyroid cancer
FTC133 and TPC-1 cells. Both in vitro and in vivo experiments showed that
silencing circ_0067934 induced apoptosis and inhibited proliferation of thyroid
cancer cells, and also increased levels of ferroptosis markers, including
Fe
IRT6 was overexpressed in thyroid cancer, and its upregulation sensitized thyroid cancer cells to ferroptosis through NCOA4-mediated autophagic degradation of ferritin. On the contrary, the silence of SIRT6 induced ferroptosis resistance. Significantly, sulfasalazine, as a clinical ferroptosis agonist, suppressed the tumor growth of thyroid cancer cells in vivo [26]. Haem oxygenase-1 (HO-1) participates in the regulation of oxidative stress and tumorigenesis. In follicular thyroid cancer tissues, HO-1 was overexpressed, whereas its upregulation activated the ferroptosis signaling pathway. A recent study reported that curcumin treatment caused ferroptosis and impeded tumor growth of thyroid cancer cells by enhancing HO-1 [48]. GPX4 was overexpressed in thyroid tumor tissues than in normal ones, and its high expression was correlated with pathologic stage III–IV, stage T3–T4, and poor prognosis, and functional study elucidated its promotive role in cell proliferation and suppressive role to ferroptosis [49]. Blocking and genetic knockdown of GPX4 significantly induced ferroptosis, impeded migration of thyroid cells, and functionally inhibited phosphorylation of NPM1 [50].
Both in 2D and 3D cultures, blocking of GPX4 reduced GSH, increased ROS, and
eventually caused ferroptosis of thyroid cancer cells and attenuated cell
migration [51]. Based on the 5-gene signature (LINC00900, LINC02454, AC012038.2,
DPP4-DT, and AC055720.2), thyroid cancer patients were divided into two groups
including low-risk and high-risk groups, and these 5-gene signatures was
identified as an independent prognostic factor in thyroid cancer. Further study
confirmed overexpression of LINC00900 and LINC02454. Furthermore, immune-related
pathways against tumors were enriched in low-risk thyroid cancer patients [52].
AlkB homolog 5 (ALKBH5) was reduced in thyroid cancer tissues, and overexpression
of ALKBH5 suppressed cell proliferation, and potentiated cellular Fe
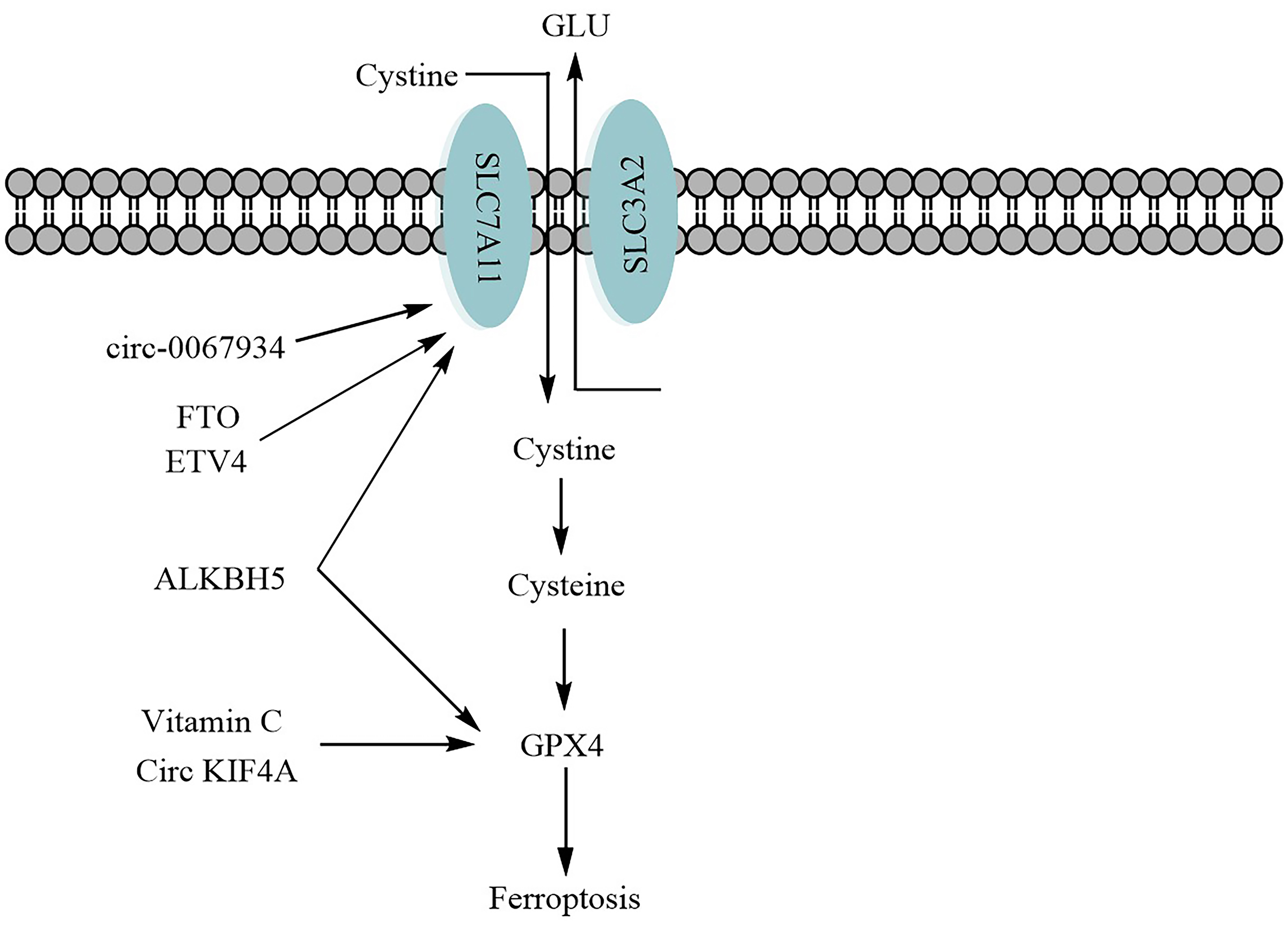
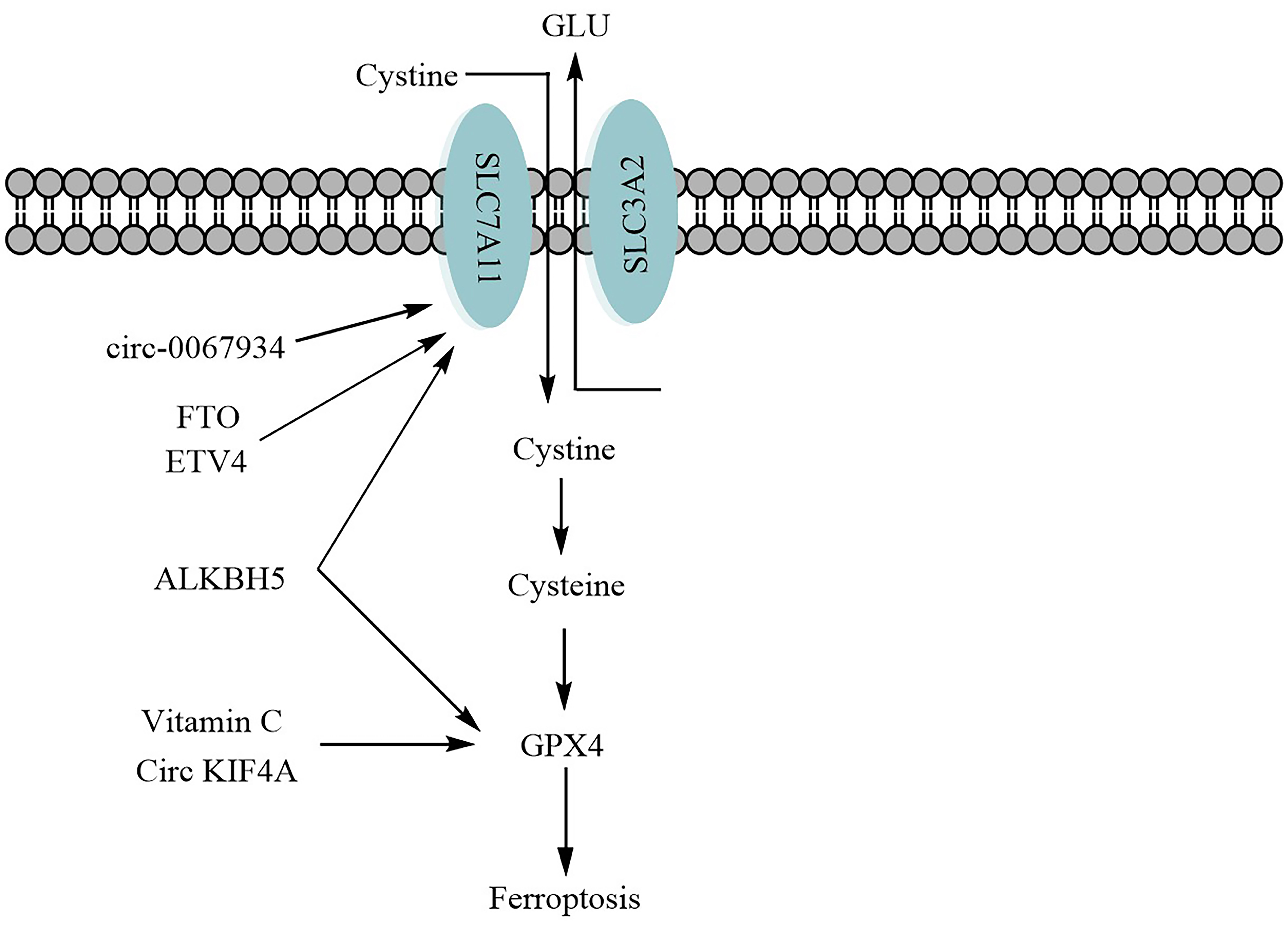 Fig. 1.
Fig. 1.Possible mechansim of targeting ferroptosis to enhance anti-thyroid cancer. FTO, Fat mass and obesity-associated protein; ETV4, variant 4; Circ KIF4A, Circular transcripts of KIF4A; SLC7A11, the solute carrier family 7 member 11; SLC3A2, the solute carrier family 3 member 2; GLU, glutathione.
Ferroptosis-associated lncRNA prognostic model (Ferr-LPM) was identified in thyroid cancer, and the patients with higher scores showed poor prognosis. Quite significantly, the Ferr-LPM score was negatively correlated with tumor purity and positively linked with immune checkpoint blockade response (Tables 1,2) [39]. Prognosis-related differentially expressed immune-related ferroptosis genes (PR-DE-IRFeGs) expression level was remarkably correlated with BRAF/NRAS/HRAS mutation and suggested that immunity and ferroptosis probably participate in the progression of thyroid cancer through BRAF/NRAS/HRAS mutation and anticancer immunity (Tables 1,2) [40].
Both ceramide synthase 6 antisense RNA 1 (CERS6-AS1) and lim and SH3 protein1 (LASP1) were highly expressed, and miR-497-5p was lowly expressed in papillary thyroid tumor tissues and cells and also identified as potential prognostic factors. Functional experiments indicated that the knockdown of CERS6-AS1 impaired cell viability and induced ferroptosis of cancer cells via sponging miR-497-5p and positively regulating downstream LASP1. Notably, the downregulation of CERS6-AS1 attenuated the growth of thyroid cancer cells (Tables 1,2) [41]. Distinguishing between thyroid cancer and thyroid follicular adenoma is an excellent challenge in the clinic. Two hundred four proteins were differentially expressed between cancer and adenoma after analyzing the proteosomes of both adenoma and cancer samples. Interestingly, ferroptosis-associated genes were dysregulated in cancer compared to adenoma, indicative of ferroptosis involving tumorigenesis of thyroid tissues [54].
About 78.8% of ferroptosis-related genes were changed expressed in thyroid tumor tissues compared with normal tissues. Twelve of these genes showed predictive prognosis value, and eight of twelve genes, including GPX4, DPP4, ISCU, GSS, PGD, MIOX, TFRC, and TF, were constructed as a prognostic model. The high-risk group showed a shortened survival time (Tables 1,2) [42].
ETV4 as a transcription factor was overexpressed in papillary thyroid cancer tissues, and the functional study revealed that the knockdown of ETV4 restrained cell cycle progression and cell proliferation. Mechanically, ETV4 bound to the promoter of SLC7A11 and upregulated its expression. In vivo experiments showed that the silence of ETV4 impeded tumor growth of thyroid cancer cells (Tables 1,2) [43]. Forty-five ferroptosis-related genes were differentially expressed in thyroid cancer tissues compared with normal tissues, and 6 of these genes were correlated with the prognosis of thyroid cancer patients. A 5-gene signature was eventually identified to predict prognosis in thyroid cancer. Notably, the high-risk and low-risk groups based on the 5-gene signature had distinctive immune status (Tables 1,2) [44]. The 8-gene ferroptosis model was identified to predict the outcome of thyroid cancer patients, and this signature was an independent prognostic marker in thyroid cancer (Tables 1,2) [45]. Vitamin C is involved in the pathogenesis of thyroid cancer through the MAPK/ERK and PI3K/AKT pathways [55]. Vitamin C, an anti-tumor agent, was found to activate ferroptosis with inactivation of GPX4 and suppress cell proliferation. Mechanically, vitamin C promoted ferritin degradation via ferritinophagy and subsequentially increased cellular free iron and ROS levels, accumulating ROS and lipid peroxidation (Tables 1,2) [46].
Expression of nine genes, including GPX4, ANXA1, CLU, TGFBR3, TNFRSF12A, PSEN1, LEF1, TIMP3, and BNIP3L was correlated with overall survival of thyroid cancer patients, and GPX4 was upregulated in thyroid cancer tissues (THCA) after analyzing GEPIA database [56]. High expression of SLC7A11 was associated with shortened overall survival time in thyroid cancer patients, and its expression was also correlated with improved mortality (OR 3.87, 95% CI 1.18–17.31) [57]. High expression of both SLC7A11 and cric_0008274 was correlated with unfavorable outcomes in thyroid cancer patients, and silence of cric_0008274 remarkably reduced cell migration and adhesion and improved cell apoptosis in vitro and tumor growth in vivo through affecting miR-154-3p/SLC7A11 signaling pathway [58].
Ferroptosis is a modality of regulatory cell death. In recent years, with the deepening of various studies, it has been found that ferroptosis has an increasingly evident relationship with the pathogenesis of various malignant tumors, and the signaling pathways involved in ferroptosis play an essential role in the occurrence and development of malignant tumors. Thyroid cancer is one of the common malignant tumors. Although most of its malignant degree is not high, its incidence is increasing yearly, and it is getting younger and younger. Since the pathogenesis of thyroid cancer needs to be further improved, this review summarizes the involvement of ferroptosis in the development of thyroid cancer cells. With the deepening of the research, the mechanism of ferroptosis-related genes is discussed. At present, although there is no specific drug or technology related to ferroptosis applied in clinical work, I believe that in future clinical work, the research on ferroptosis mechanism will lead to the development of oncogene targeting drugs, which will undoubtedly be a major breakthrough for patients with advanced thyroid cancer. However, the detailed mechanism of thyroid cancer cells’ resistance to ferroptosis has not been fully revealed, especially in the aspect of tumor immunity, which requires further research.
RH and XL designed the research study. ZS performed the research. JY and YR provided help and advice on analyzing the data. RH and ZS wrote the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work to take public responsibility for appropriate portions of the content and agreed to be accountable for all aspects of the work in ensuring that questions related to its accuracy or integrity.
Not applicable.
Not applicable.
This research received no external funding.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.