








1 Department of Ophthalmology, Peking Union Medical College Hospital, Chinese Academy of Medical Sciences & Peking Union Medical College, 100730 Beijing, China
2 Key Laboratory of Ocular Fundus Diseases, Chinese Academy of Medical Sciences, 100730 Beijing, China
Abstract
Background: NOD-, LRR-, and pyrin domain-containing protein 3
(NLRP3) is a common inflammatory factor that induces inflammation by increasing
the expression of related cytokines. Although the NLRP3 inflammasome has been
implicated in many ophthalmic diseases, its role in myopia is largely unknown.
The aim of this study was to explore the relationship between myopia progression
and the NLRP3 pathway. Methods: A form-deprivation myopia (FDM)
mouse model was used. Different degrees of myopic shift were achieved via
monocular form deprivation with 0-, 2-, and 4-week covering, and by 4-week
covering followed by 1-week uncovering (the blank, FDM2, FDM4, and FDM5 groups,
respectively) in both wild-type and NLRP3 (-/-) C57BL/6J mice. Axial length and
refractive power were measured to assess the specific degree of myopic shift. The
protein levels of NLRP3 and of related cytokines in the sclera were evaluated by
Western blotting and immunohistochemistry. Collagen I and matrix
metalloproteinase-2 (MMP-2), which affect extracellular matrix (ECM) remodeling
of the sclera, were also examined to clarify the possible underlying mechanism.
Results: In wild-type mice, the FDM4 group had the most
significant myopic shift. Both the increase in refractive power and the
elongation in axial length were significantly different between the experimental
and control eyes in the FDM2 group. The protein levels of NLRP3, caspase-1,
IL-1
Keywords
- NLRP-3
- myopia
- inflammation
Myopia is a highly prevalent eye disease worldwide [1]. An epidemiological survey found that in urbanized East Asia and North America, more than half the children who complete their high school education are myopic [2]. A recent meta-analysis has suggested that close to half of the world’s population may be myopic by 2050, with as many as 10% being highly myopic [3]. Low to moderate degrees of myopia make daily life inconvenient due to blurred distance vision. This is usually corrected with spectacles or contact lenses, or by refractive surgery. In some cases, high degree myopia is also correctable using these optical approaches, although there is a significant increase in the incidence of pathological myopia, myopic maculopathy, chorioretinal degeneration, retinal degeneration and detachment [4]. However, permanent vision loss can occur in these situations. The prevention and treatment of myopia, and particularly high degree myopia, has therefore become an important international public health priority [1].
In the vast majority of myopic cases (
Previous studies have shown that chronic inflammation is involved in myopia
progression [13, 14]. The population-based cohort study showed that patients with
autoimmune diseases such as type 1 diabetes mellitus, systemic lupus
erytheomatosus, uveitis, and allergic conjunctivitis have a higher risk of myopia
compared to patients without autoimmune disease. In animal models of myopia,
inflammatory markers such as c-Fos, nuclear factor kappa B (NF
Healthy 3-week-old male wild-type C57BL/6J mice (Beijing Vital River Laboratory Animal Technology Co., Ltd., Beijing, China) and NLRP3 (-/-) C57BL/6J mice (ViewSolid Biotech Co., Beijing, China) of the same age and with body weights of 12–14 g were used in this study. Mouse genotypes were confirmed by DNA sequencing technology. Both types of mice were randomly divided into four groups, comprising three FDM groups (12 mice in each group) and a blank group that was comprised of 3 subgroups (6 mice in each subgroup). The FDM mouse model involved covering the right eye with latex balloons for 2 weeks (FDM2 group), 4 weeks (FDM4 group), or 4 weeks followed by 1-week uncovered (FDM5 group). The balloons were also used to make collars that prevented the mice from using their paws to remove the balloon covering their eye. The mice in the blank group did not receive any treatment and were examined when they were 5, 7 and 8 weeks old, thus corresponding to the ages of mice in the FDM2, FDM4 and FDM5 groups, respectively. All mice were exposed to daily periods of 12 hours of natural light and 12 hours of dark. Water, vegetables, and vitamins were provided ad libitum. Animal treatment adhered to the Association for Research in Vision and Ophthalmology’s Statement on the Use of Animals in Ophthalmic and Vision Research. The animal experiments were conducted under pathogen-free conditions in line with the institutional animal care protocols approved by the Peking Union Medical College Hospital (XHDW-2022-52).
Mice were anesthetized with an intraperitoneal injection of 1% pentobarbital sodium and the pupils were then rapidly dilated with compound tropicamide. The refraction of both eyes was measured in a dark room using a banded optometric apparatus (Suzhou Sixty Six Vision Technology Co., Ltd., Suzhou, China; Model YZ24) in the direction of the visual axis. After the mice were sacrificed, their eyes were removed with a forceps and stripped of as much peripheral connective tissue as possible so that the axis of the eye was oriented in the same direction as the micrometer (Dongguan Sanqiang Hardware Machinery Co., Model 211-101). Readings were taken just as the spiral micrometer touched the corneal surface, which determined the axial length (AL). Three measurements were made, and the average value was rounded to the nearest 0.001 mm. All measurements were performed and recorded by experienced personnel.
The removed eye samples were fixed in 4% formaldehyde solution at 4 ℃ and then
cut into sections, blocked, and incubated with primary antibodies for observation
under a fluorescent microscope. For the negative control, normal serum was used
instead of the primary antibody. The evaluation of fluorescence was carried out
in double-blind fashion. The primary antibodies were specific for NLRP3 (1:200
dilution; #15101, cell signaling technology, USA), IL-1
The sclera was snap-frozen in liquid nitrogen, ground into a powder and then
placed in lysis solution for 60 minutes. The scleral lysates were prepared in SDS
lysis buffer containing a mixture of protease and phosphatase inhibitors (Pierce,
Rockford, IL). The proteins were quantified using a Bradford protein assay and 40
SPSS 22.0 (IBM Corp., Chicago, IL, USA) statistical software was used for
statistical analysis. Measurement data in this study were tested for normality
using the Shapiro-Wilk test and expressed as the mean
To determine whether the myopic shift in wild-type and NLRP3 -/- mice was
comparable in the subsequent study, we first compared the refractive status of
the blank group in both wild-type and NLRP3 -/- mice. No significant differences
were found in axial length (AL) or refraction power between the two mouse types
at the same age (Fig. 1B,C). Immunohistochemical staining showed that NLRP3 was
expressed at much lower levels in NLRP3 -/- mice compared to wild-type mice.
Other cytokines related to the NLRP3 pathway and to the scleral ECM, including
IL-1
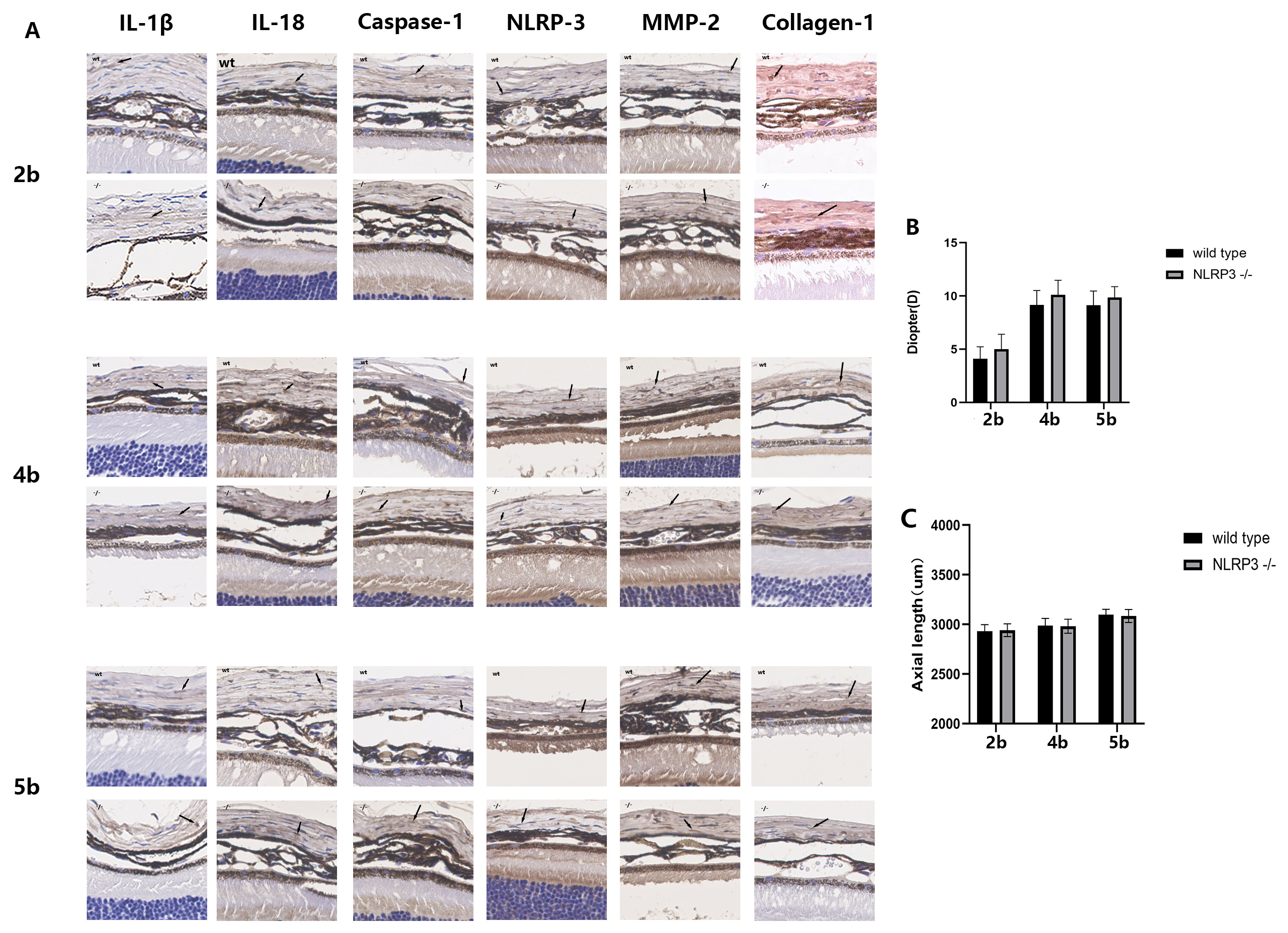 Fig. 1.
Fig. 1.Refractive status and related cytokine expression in the blank
group of wild-type mice and NLRP3-/- mice. 2b, 4b, 5b represent the eyes from
the blank group at the same age as the FDM2, FDM4 and FDM5 groups, respectively.
(A) Immunohistochemical images of sclera. Each image corresponds to an actual
size of 100
| 2c | 2b | 2e | 4c | 4b | 4e | 5c | 5b | 5e | |
| +/- | +/- | +/- | +/- | +/- | +/- | +/- | +/- | +/- | |
| IL-1 |
1.2 |
0.9 |
5.1 |
2.1 |
1.8 |
9.5 |
2.9 |
2.6 |
5.4 |
| IL-18 | 3.3 |
2.7 |
7.3 |
6.2 |
6.1 |
8.8 |
4.0 |
4.7 |
8.0 |
| Caspase-1 | 1.5 |
1.8 |
4.3 |
1.8 |
2.0 |
8.5 |
2.6 |
2.3 |
1.6 |
| NLRP-3 | 2.4 |
2.2 |
5.7 |
3.0 |
3.5 |
7.4 |
3.1 |
3.4 |
2.2 |
| MMP-2 | 1.1 |
1.6 |
7.6 |
4.8 |
5.1 |
9.4 |
3.0 |
3.0 |
3.5 |
| Collagen-1 | 4.7 |
5.0 |
3.5 |
3.3 |
4.0 |
2.1 |
4.8 |
4.4 |
2.8 |
| The values shown in the table are the average IRS from 6 eyes in the same group.
Data are expressed as the mean | |||||||||
We next investigated whether changes to the NLRP3 inflammasome in the sclera were associated with myopia development. To do this, the myopic shift, AL elongation, and expression levels of NLRP3 and caspase-1 in the experimental eyes were compared with the control eyes (i.e., the uncovered contralateral eye) in the FDM2 and FDM4 groups of wild-type mice.
In the FDM4 group, significant myopic shift (Fig. 2C; 5.30
 Fig. 2.
Fig. 2.The expression of inflammatory cytokines related to the NLRP3
pathway and of scleral tissue remodeling proteins increased over time in
experimental eyes, and decreased after removal of the eye mask. Form deprivation
promoted myopic shift in wild-type C57/BL6J mice. 2c represents the control eyes
in the FDM2 group, 2e represents the experimental eyes in the FDM2 group, 2b
represents the eyes in the blank group of mice with the same age as FDM2, and so
on for FDM4 and FDM5. (A) Immunohistochemical images of sclera. Scleral tissues
were obtained from wild-type mice. Each image corresponds to an actual size of
100
Western blotting and immunohistochemistry revealed that the expression levels of NLRP3 and caspase-1 protein were both correlated with the duration of form deprivation. In both the FDM2 and FDM4 groups, the NLRP3 and caspase-1 levels in the sclera of experimental eyes were higher than in the control eyes (Fig. 3A–C, and Fig. 2A). NLRP3 and caspase-1 expression in the experimental eyes of the FDM4 group were higher than in the control eyes of the same group (Fig. 3B,C, p = 0.006), and also higher than in the experimental eyes of the FDM2 group (Fig. 3B,C, p = 0.032). This was consistent with the results observed for myopic shift and AL elongation. Similar results were also obtained using the immunohistochemical semi-quantitative method for comparison (Table 1). These findings suggest that myopic progression and activation of the NLRP3 inflammasome pathway occur in the FDM mouse model.
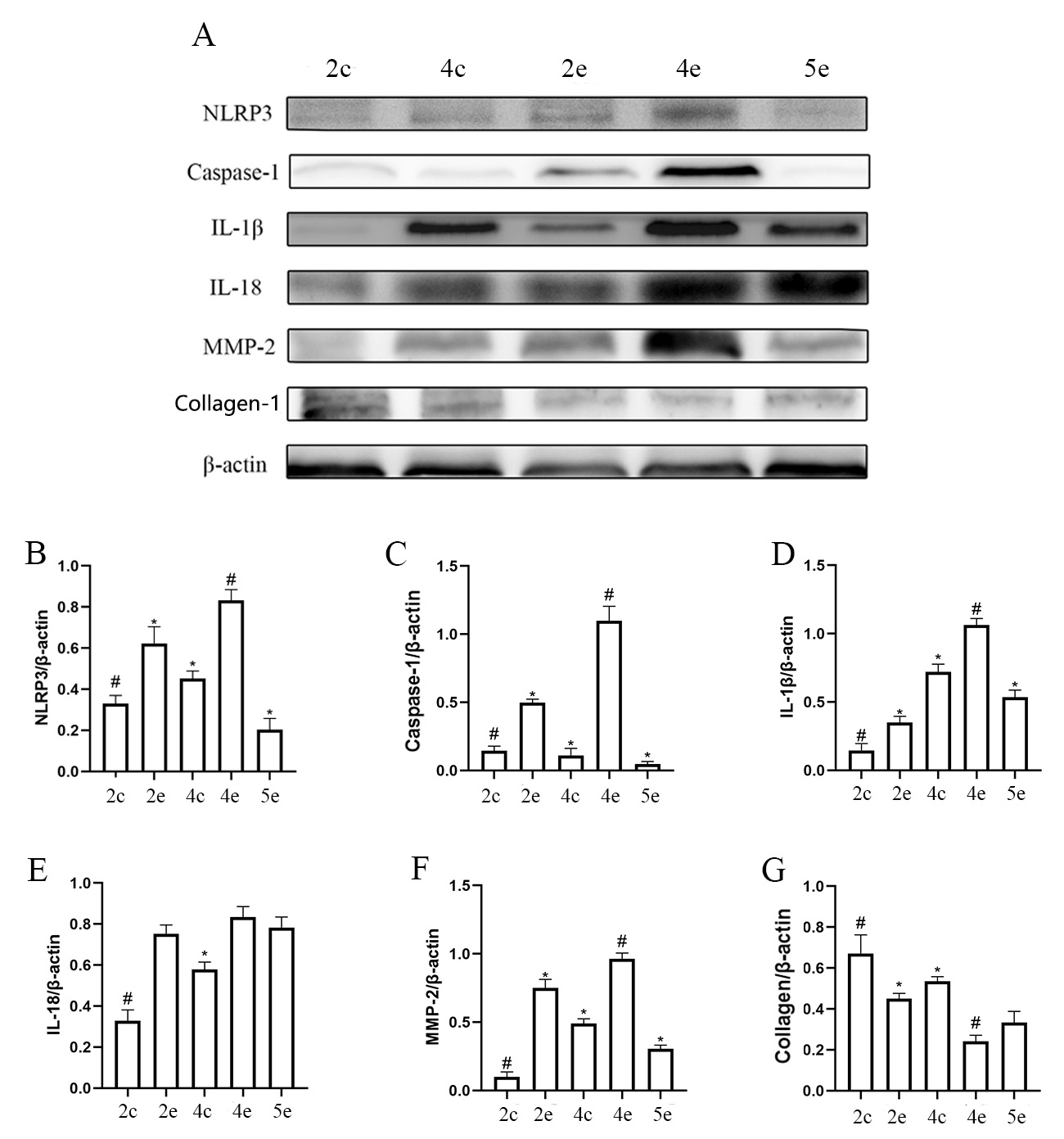 Fig. 3.
Fig. 3.Form deprivation promoted scleral NLRP3 inflammasome activation
in wild-type C57/BL6J mice. 2c represents the control eyes in the FDM2 group, 2e
represents the experimental eyes in the FDM2 group, and so on for the FDM4 and
FDM5 groups. (A) Representative Western blot results for NLRP3, caspase-1,
IL-1
The relationship between NLRP3 expression and myopia was further investigated by
comparing the AL, myopic shift, and NLRP3 expression in the experimental eyes of
wild-type mice in the FDM5 group with those from the FDM4 group. One week after
removal of the covering, the expression levels of NLRP3 and caspase-1 were
significantly lower, as observed by Western blotting (Fig. 3B,C) and
immunohistochemistry (Fig. 2A). The myopic shift in the experimental eyes was
also significantly reduced (Fig. 2B; 9.90
To clarify the effect of NLRP3 on the development of myopia, we examined FDM using NLRP3 -/- mice and measured the refraction power, AL and expression levels of NLRP3 and caspase-1 in the different experimental groups. The results were compared with those of wild-type mice from the same age group.
Refractive changes in the experimental eyes from the FDM4 group of NLRP3 -/-
mice were again significantly different to those of control eyes (Fig. 4B; 7.33
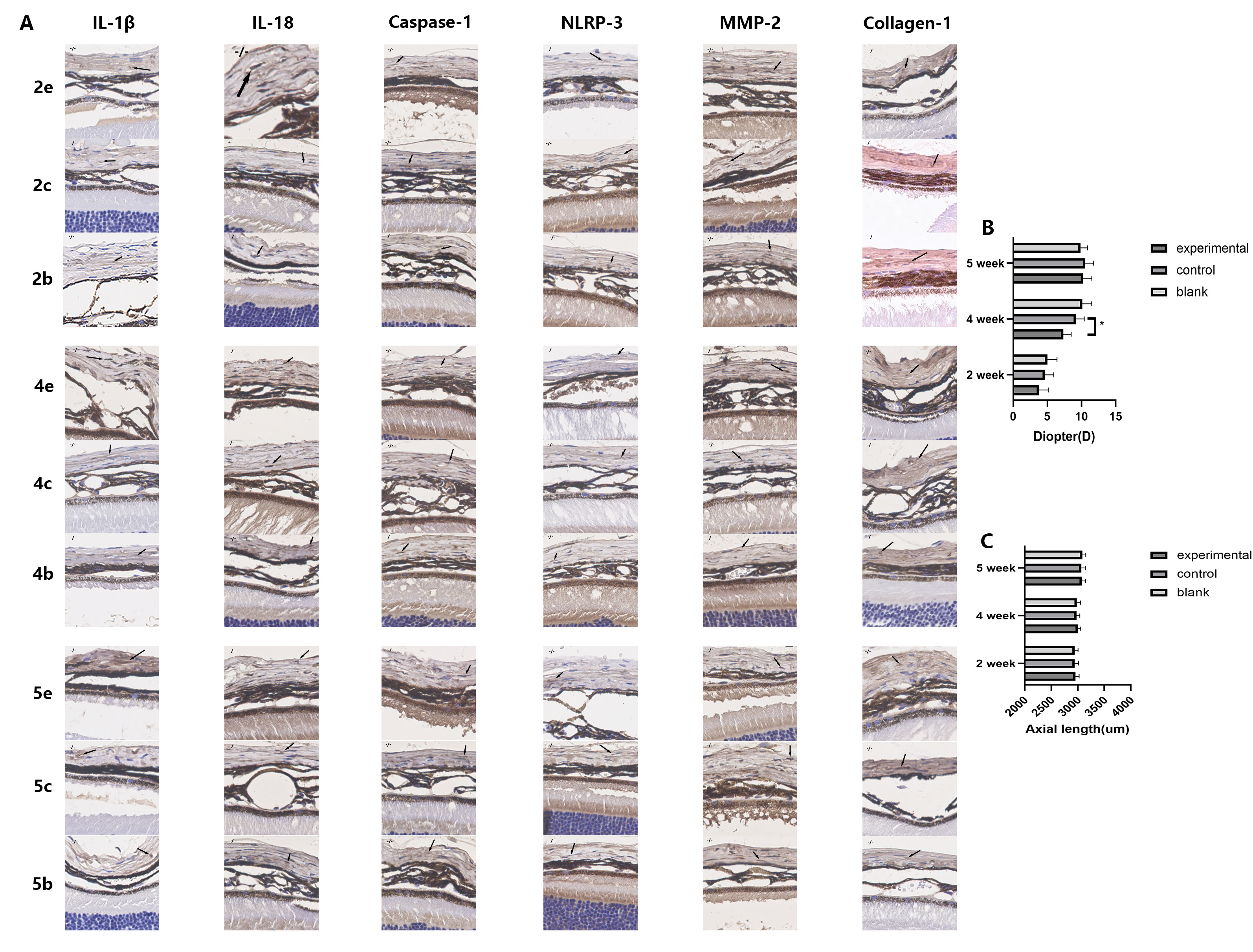 Fig. 4.
Fig. 4.The expression of inflammatory cytokines related to the NLRP3
pathway and of scleral tissue remodeling proteins increased over time in NLRP3
-/- mice, but this did not reach statistical significance. 2c represents the
control eyes in the FDM2 group, 2e represents the experimental eyes in the FDM2
group, 2b represents the eyes in the blank group with the same age as the FDM2
group, and so on for the FDM4 and FDM5 groups. (A) Immunohistochemical images of
scleral tissues obtained from NLRP3 -/- mice. Each image corresponds to an actual
size of 100
Furthermore, NLRP3 expression in NLRP3 -/- mice was not statistically different between the various groups (Fig. 5D and Fig. 4A) and relatively low (Fig. 6B Table 1). The expression level for Caspase-1 in the experimental eyes of the FDM4 group was higher than in the other groups of NLRP3 -/- mice (Fig. 5F), but significantly lower than that observed in wild-type mice (Fig. 6A). The myopic shift in the FDM mouse model was suppressed when NLRP3 expression was lower.
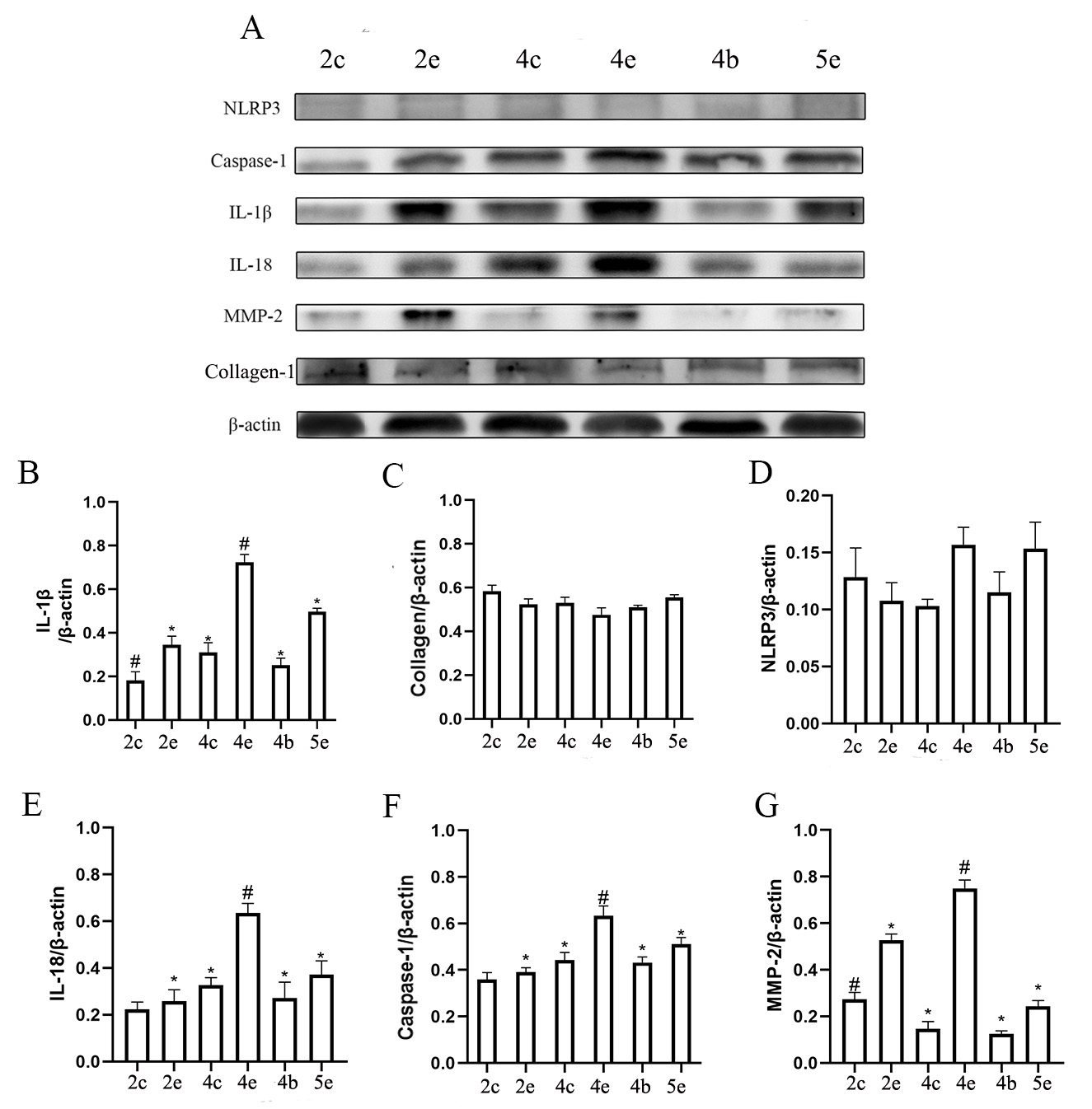 Fig. 5.
Fig. 5.NLRP3 knockout reduced scleral NLRP3 inflammasome activation in
the FDM mouse model. 2c represents the control eyes in the FDM2 group, 2e
represents the experimental eyes in the FDM2 group, and so on for the FDM4 and
FDM5 groups. (A) Representative Western blot for expression of the NLRP3,
caspase-1, IL-1
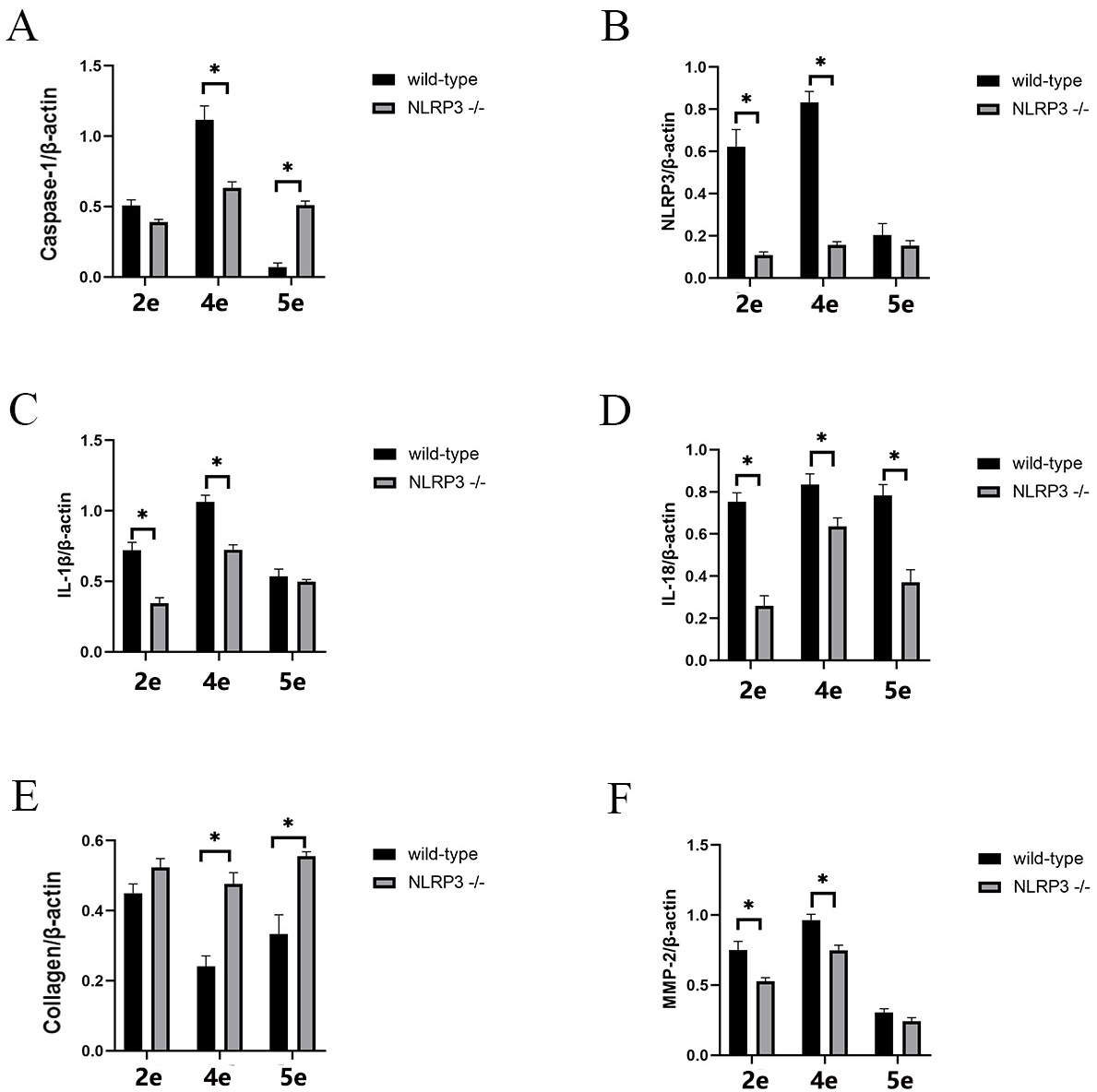 Fig. 6.
Fig. 6.Deletion of NLRP3 reduces the development of FDM and the
expression of inflammatory cytokines related to NLRP3. Comparison of caspase-1
(A), NLRP3 (B), IL-1
To explain the above findings, we measured the expression of IL-18 and
IL-1
The expression of collagen I was completely different to IL-1
In wild-type mice, IL-18 showed high expression in all experimental eyes, with no statistical difference between groups. In contrast, the experimental eyes in the FDM4 group of NLRP3 -/- mice showed a statistically different level of IL-18 expression to all of the other groups (Fig. 5E), although they also showed reduced levels compared to wild-type mice (Fig. 6D). It is possible that the FDM model induces IL-18 activation through pathways other than NLRP3/caspase-1, therefore explaining why masking still increased IL-18 expression in NLRP3 -/- mice.
In the FDM4 group of NLRP3 -/- mice, no statistical differences were observed between the expression of each factor in the eyes of the blank group compared to the eyes of the controls. This demonstrates that the FDM model did not affect the expression level of each factor in the control eyes of NLRP3 -/- mice (Fig. 5B–G).
Axial elongation is the most important form of change in the progression of myopia [20], and was recapitulated in the FDM mouse model used in the current study. In the FDM4 group, the refraction shift towards myopia and axial elongation were both significantly different in the experimental eyes compared to the control eyes. Moreover, the difference diminished after the mask had been removed for one week. These results confirm a successful establishment of the myopic model and validate the findings of previous studies [21]. There were no significant differences in either refraction or AL between eyes in the blank group and control eyes in the FDM4 group, suggesting that masking one eye did not affect development of the unmasked control eye. Together with the observation of no significant differences in myopic shift and AL elongation between the control eyes in the FDM4 group and the eyes in the blank group, we believe that monocular form deprivation in wild-type mice has no significant effect on the untreated eye in FDM models.
In this study, immunohistochemistry and Western blotting showed that protein expression levels for NLRP3, Caspase-1, IL-18, and MMP-2 were highly consistent with myopic shift and AL elongation in wild-type mice. These results strongly suggest the NLRP3 pathway is associated with the progression of myopia. Previous studies have shown that NLRP3 is involved in various eye diseases such as keratitis, diabetic retinopathy, dry eye disease and age-related macular degeneration [22, 23, 24]. The current study has for the first time revealed an association between the NLRP3 pathway and myopia.
The role of inflammation in myopia progression has been described previously
[25, 26]. Lin and colleagues reported that expression levels for inflammatory
cytokines including c-Fos, nuclear factor
Previous studies have demonstrated that MMP-2 can induce scleral ECM remodeling,
thereby playing a key role in myopia development. IL-18 and IL-1
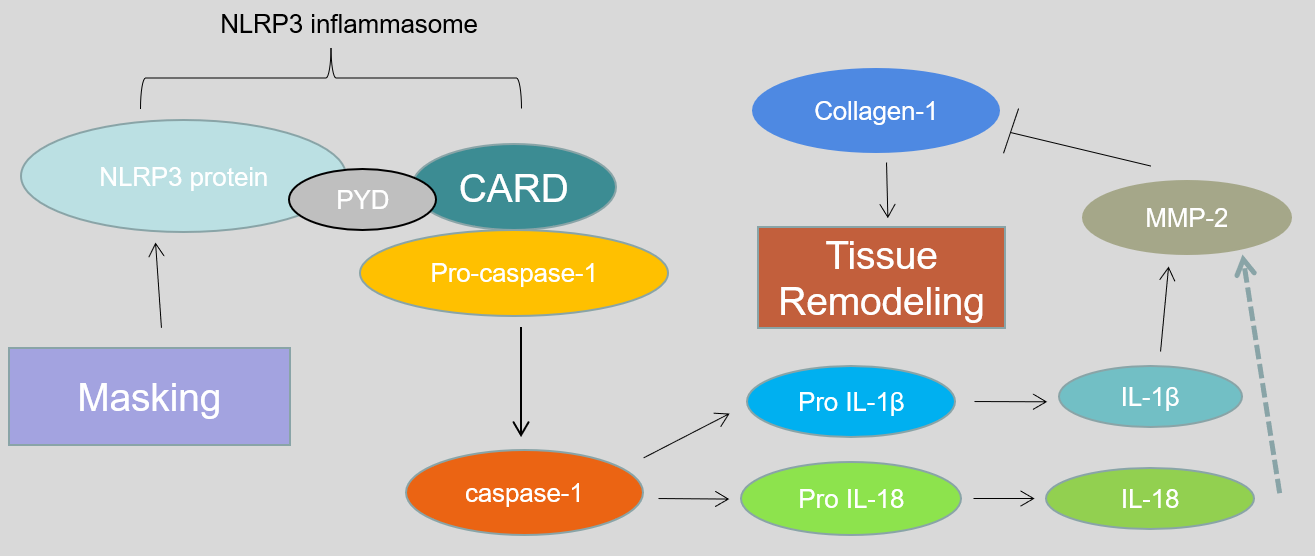 Fig. 7.
Fig. 7.Possible mechanism explaining how the up-regulation of NLRP3 induces a myopic shift.
NLRP3 -/- mice experienced a significant myopic shift in the form-deprivation eyes compared with control eyes, although to a much lesser extent than in wild-type mice. Similiar trends were observed for axial elongation. A possible explanation could be that NLRP3 was only partially suppressed in the knockout mice used in this study, and the residual expression of NLRP3 could still play a role in the development of myopia. Another explanation is that myopia is a multifactorial disease, and hence the progression of myopia is only partly determined by NLRP3 activation. The FDM model could also undergo myopia progression through other pathways.
The situation with IL-18 was somewhat complicated by the fact that its generally high expression in the experimental eyes of wild-type mice prevented us from determining whether it was also involved in the aforementioned mechanism of myopia progression. However, the expression level of IL-18 in NLRP3 -/- mice was consistent with a myopic shift. This suggests there are other pathways in the FDM model besides NLRP3 that can increase the IL-18 expression level and thus be associated with myopia progression.
Despite a plethora of research on myopia, the molecular/cellular mechanisms that underlie its pathology are still not well understood, thereby slowing the search for more effective pharmacological controls. So far, the anticholinergic blocking agent atropine has been the most effective and is used by clinicians in off-label ways for myopia control. However, there are concerns regarding potential long-term ocular or systemic side-effects of atropine usage. NLRP3 is a common inflammatory factor involved in the development and progression of many eye diseases. Inhibitors of NLRP3 have proven effective in the treatment of these diseases [30, 31, 32]. The present study demonstrated the involvement of NLRP3 in the progression of myopia in the FDM model. Moreover, the suppression of NLRP3 expression reduced the extent of myopic shift, suggesting that NLRP3 inhibitors could potentially be used to prevent or slow the development of myopia.
A limitation of this study was that only protein expression levels were assessed, with no analysis at the transcriptional level using methods such as RT-PCR. In addition, Western blotting and immunohistochemistry are semi-quantitative descriptions and somewhat inadequate for precise numerical evaluation.
In summary, this is the first report to our knowledge that NLRP3 could be
involved in myopic shift in the FDM mouse model. The underlying mechanism might
be that NLRP3 promotes the activation of IL-1
All data generated or analyzed during this study are included in this published article.
ZC designed the research study. ZC and KX performed the research. QL provided help and advice on the research design and paper writing. ZC analyzed the data. ZC wrote the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript.
Animal treatment adhered to the Association for Research in Vision and Ophthalmology’s Statement for the Use of Animals in Ophthalmic and Vision Research, and the animal experiments were conducted under pathogen-free conditions in line with the institutional animal care protocols approved by the Peking Union Medical College Hospital (XHDW-2022-52).
The authors of this article would like to thank Yongfeng Lv and Lu Sun for their guidance on optometry techniques.
This research was funded by National Natural Science Foundation of China (Number 81870685) and the Natural Science Foundation of Beijing Municipality (Number 7172173).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.