
 , Wen Du 1, Shuai Shang 1,*
, Wen Du 1, Shuai Shang 1,*1 College of Biological and Environmental Engineering, Binzhou University, 256600 Binzhou, Shandong, China
Academic Editors: Josef Jampílek and Vesna Jaćević
Abstract
Introduction: Non-alcoholic fatty liver disease (NAFLD) has become one of the most common liver diseases globally and its negative impact has grad- ually attracted attention. Method: In order to explore whether camphor seed oil has a certain link effect on NAFLD, we identified its remodeling of intestinal flora and liver function index through the use of camphor seed kernel oil as food treatment (CCSKO). Results: Our results showed that camphor seed oil significantly improved the liver index, TG content and histopathology of the NAFLD mice compared with other groups. Meanwhile, we found significant differences in the intestinal microbiota of mice in different treatment groups. Discussion: The CCSKO treatment might significantly increase the abundance of Bacteroidetes, which were involved in many important metabolic activities in the human colon, including carbohydrate fermentation, utilization of nitrogenous substances, and biotransformation of bile acids and other steroids. Conclusions: These findings indicate a strong contribution in shaping the gut microbiota by treatment groups, shedding light on the impact of camphor tree seed kernel oil on gut microbiota.
Keywords
- non-alcoholic fatty liver disease
- camphor seed kernel oil
- intestinal microbiota
- mice
Non-alcoholic fatty liver disease (NAFLD) is a special kind of liver disease, including the liver disease encompassing steatosis (fatty change), non-alcoholic steatohepatitis (NASH) and cirrhosis in the absence of alcohol abuse. With the development of economy and the improvement of living standards, the dietary structure and lifestyle have also changed greatly. The prevalence of NAFLD is reported to be increasing rapidly in parallel with the dramatic rise in obesity and diabetes [1, 2], and is increasingly recognized as the leading cause for liver dysfunction and cirrhosis in the non-alcoholic. Nowadays, viral hepatitis negative population has become a social problem with increasing morbidity and mortality worldwide. As the metabolic risk factors, the incidence of NAFLD has been increasing in recent years, and even gradually affecting younger people. NAFLD has become one of the most common liver diseases globally and has gradually attracted attention [3]. Beyond diet and exercise, no effective therapeutic approaches exist to eliminate NAFLD, or prevent its progression to NASH and end-stage liver disease [4, 5]. The prevailing hypothesis suggests that disease progression might be triggered when an acute inflammatory insult, oxidative stress, adipose tissue expandability, changes in climate, agro-alimentary logistics, and political direction on hepatic steatosis [4, 6].
As a major nutrient, oil is the main source and storage form of energy for the body. Except for providing energy for the body, the oil has many other important physiological functions, including the survival and development of human beings. Currently, the cooking oil is still soybean oil, rapeseed oil, corn oil, peanut oil, lard, etc. However, excessive dietary fat intake leads to the increasing incidence of metabolic diseases such as obesity, cardiovascular and cerebrovascular diseases, diabetes, etc. If nutrition is taken for a long time storage of fat metabolism of the liver and muscle tissue, leading to nutritional imbalance. In that case, buffer storage cells can force excess nutrients a lot in the process of physiological metabolism, and bring the related metabolic tissue at a high concentration of the metabolites of the environment for a long time, eventually leading to cell functional disorder and abnormal lipid metabolism [7]. Therefore, the three low and one high dietary principles of low oil, low sugar, low salt and high fiber are advocated. Theoretically, reducing the fat intake may prevent disease, but low-fat or fat-free foods may affect the taste.
The medium carbon chain fat (MCT) as a short carbon chain is good flavor and has healthy functions, attracting more and more attention in the fields of oil and nutrition. In the liver, the MCT is quickly and efficiently decomposed to produce energy, and its digestion and absorption rate is 4 times than that of the ordinary long chain fatty acids. And its metabolism speed is 10 times than that of the ordinary long chain fatty acids. The MCT is easy to digest and metabolize, which can limit the intake of calories and prevent the increase of fat [8, 9]. And the MCT can rapidly cross the mitochondrial inner membrane into the mitochondria of hepatocytes for beta oxidation, which is rapid in oxidation and not easy to accumulate in adipose tissue and liver tissue [10]. Caprylic acid MCT can provide energy efficiently without causing fat deposition, which can reduce the pressure on the liver. Thus, the MCT is difficult to form store, cause the body fat deposition and cause obesity [8]. The MCT composed of capric acid and octanoic acid which has metabolic pathway nutritional and clinical therapeutic value, which is used as a nutritional drug to treat fat metabolism disorders [11]. Adding the MCT to diet can increase calories and reduce fat deposits.
Camphor tree (Cinnamomum camphora (L.) Presl) as a specialty of China’s precious economic forest species is rich in natural camphor, aromatic oil and other medicinal substances [12]. Previous study found that these plants were rich in volatile oil and contained aromatic compounds of diterpenoid tannins and glycoside alkaloids, lignin organic acid polysaccharides and pigments [13]. Though, the camphor tree has high economic value, there is very little research on the camphor tree seeds kernel oil and the lipid-lowering effect of reducing weight. Each year, a large number of mature camphor tree seeds scatter everywhere, not only polluting the environment to a rough estimate, but also causing the waste resource. The intestinal flora plays an important role in metabolites and has a biological barrier function involved in the regulation of immune response. In addition, these intestinal bacteria perform structural and tissue functions. The composition of the intestinal flora can reflect the health status of the host. More and more studies focus on how the intestinal flora is shaped during the occurrence of disease, and expect to find potential signals of pathology from the intestinal flora. Therefore, this study was expected to complete the following experiments. First, we hoped to find out the relevant indicator flora by comparing the bacterial flora changes before and after the occurrence of non-alcoholic fatty liver; Second, we hoped to identify how the use of camphor tree seed kernel oil as food treatment could remodel the intestinal flora.
Four-week-old C57BL/6J male mice (Reg. No. SYXK (Lu) 20160008) were purchased
from the Experimental Animal Center of Binzhou University. The mice were housed
five individuals per cage under standardized environmental conditions
(temperature 22
At the end of the experiment, the mice in each group fasted and could not stop
drinking water for 12 h. All mice were anesthetized by
inhalation of 2.5% isoflurane, killed by intraperitoneal injection of 300
The weight of the liver in each sample was about 0.5 g. The intestinal contents of the mice were collected and stored at –20 °C for gene sequencing. The body weight of each mouse was recorded and anesthetized with ether. After disinfection with 75% alcohol, the abdominal cavity was quickly opened, the liver was removed, and the surrounding tissues were removed. Each liver was then made into homogenate. The enzyme standard instrument was used to test the triglyceride (TG) and triglyceride (TC).
The total DNA was extracted in duplicate from the mice gut content.
Approximately 250 mg of gut contents and 10 L of water samples were used for each
DNA extraction. The E.Z.N.A. ®Stool DNA Kit (D4015, Omega, Inc.,
USA) was used to extract DNA extractions from different samples according to
manufacturer’s instructions in the present study. The 50
Raw sequencing reads were obtained from the Illumina. BaseSpace website and analyzed with the Quantitative Insights into Microbial Ecology (Qiime v1.7.0) pipeline.
All experiments were performed in triplicates. All data are presented as the
means
For bioinformatic analyses, the mothur v1.39.5 [14] was used to to demultiplex and quality the raw sequencing reads. Then, the suspect sequences were removed based on existing databases. At least 97% sequence similarity, the operational taxonomic units (OTUs) were determined with Uparse v7.0.1001. The GraPhlAn was used to graph the OTUs identified in each microbiome [15]. The Qiime v1.7.0 was used to calculate the beta diversity and alpha of the gut microbiomes of the mice. In order to further measure alpha diversity, the mothur v1.39.5 [14] was used to calculate Good’s coverage scores based on 10,000 iterations. The R v3.2.2 was used to calculate the Shannon, chao1, Simpson alpha diversity indices and abundance-based coverage estimator evenness metrics. Then, the beta diversity of the OTUs was evaluated. Based on the rarefied data and square-root transformed the read abundance data, the composition of identified community members in the mice was calculated with Bray-Curtis dissimilarity matrix [16]. The principal component analysis (PCA) was performed in R v3.2.2 to calculate the overall similarity in bacterial community structure across the micAvailability of supporting data.
The GenBank/EMBL/DDBJ accession numbers reported in this paper are as follows: the associated BioProject and Bio-Sample numbers are PRJNA765784 and SAMN21584113-21584137, respectively.
During the experiment, the mental state of mice in each group was stable, drinking and feeding were normal, and there was no abnormal performance or death. After successfully constructing the NAFLD model, mice showed slow movement, reduced activity, and weight loss. Compared with the blank group, fecal information showed a significant sense of greasiness.
After poisoning, the weight of the damaged organs will change, and the organ coefficient will change accordingly. According to the viscera index in Table 1, the blank group and camphor seed kernel oil were compared. The results showed that the experimental data of the two groups were similar, the ratio of organs and body weight was relatively constant, and the organs in the camphor seed kernel oil group were less damaged. The liver index of the natural recovery group was increased with congestive viscera.
| Group | liver index |
| Control group (NC) | 4.93% |
| Natural recovery group (NR) | 5.08% |
| Camphor seed kernel oil group (CCSKO) | 4.88% |
| Note: The data are presented as means | |
The liver is an important site for the synthesis and metabolism of endogenous lipids and lipoproteins, which is closely related to lipid metabolism. The fat that enters the liver through lipid metabolism is continuously synthesized into TG and enters the blood circulation in the form of lipoprotein, forming a dynamic balance. Once the synthesis and operation balance in the liver are unbalanced, TG accumulation in the liver will lead to the formation of non-alcoholic fatty liver. As shown in Table 2, TG concentration in the liver of mice in the camphor seed kernel oil group was not significantly different from that in the normal control group. In contrast, TG concentration in the liver of mice in the natural recovery group was significantly higher than that in the control and the camphor seed kernel oil groups.
| Group | Triglyceride concentration (mmol/L) |
| Control | 2.034 |
| NR | 2.188 |
| CCSKO | 2.110 |
| Note: The data are presented as the means | |
As shown in Table 3, TC concentration in the liver of mice in the camphor seed kernel oil group was not significantly different from that in the normal control group. In contrast, TC concentration in the liver of mice in the natural recovery group was significantly higher than that in the normal control group and the camphor seed kernel oil group.
| Group | Cholesterol degrees (mmol/L) |
| Control | 0.618 |
| NR | 0.883 |
| CCSKO | 0.750 |
| Note: The data are presented as the means | |
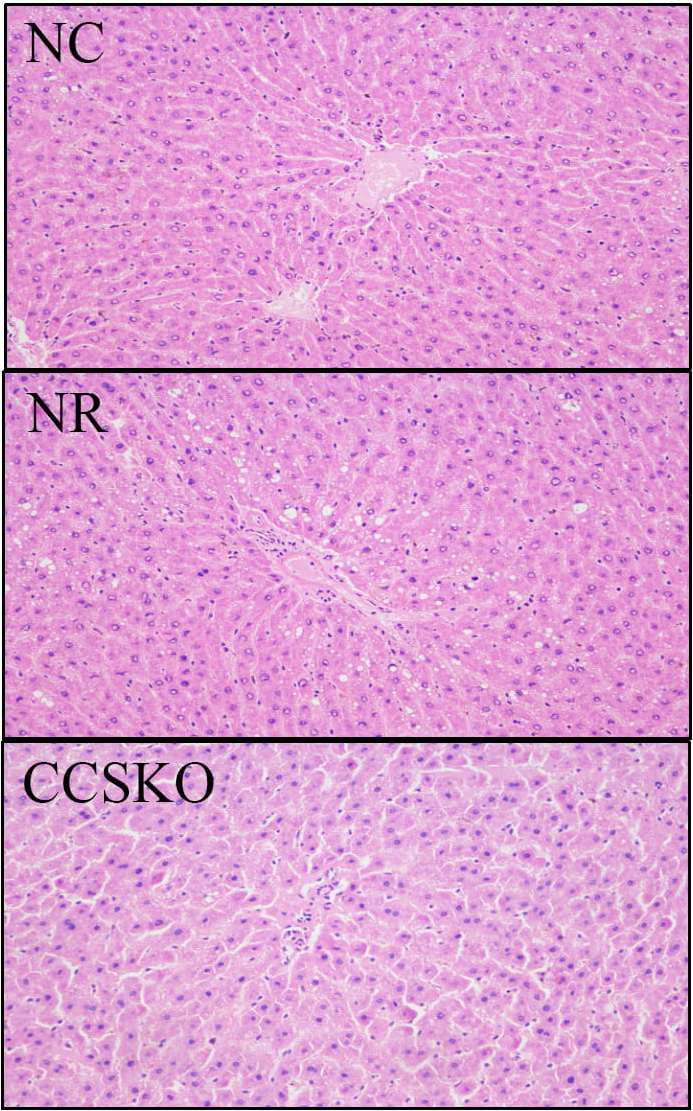
In the NR group, lipid-droplet vacuoles of different sizes, round and tension were found in the cytoplasm of liver cells, and the liver tissues were damaged to different degrees (Fig. 1). The cellular structure of liver tissue in CCSKO group was similar to that in normal control group, and the structure was intact without degeneration and necrosis.
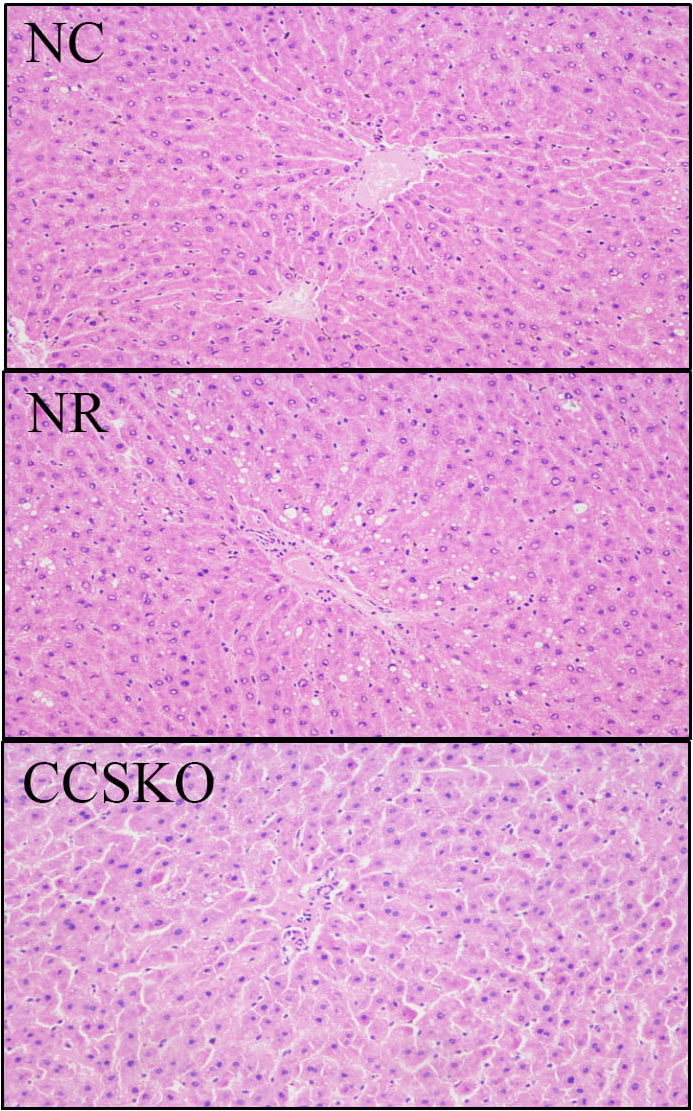 Fig. 1.
Fig. 1.HE staining of the mouse liver.
A total of 1867,251 high-quality sequences for subsequent analysis were obtained from 25 samples in five subgroups using high-throughput sequencing technology, filtration and quality control. After clustering with 97% similarity, each sample contained an average of 1687 OTUs, among which the sample with the most OUT was NC_4 (2223) and the sample with the least OUT was Model_3 (1166).
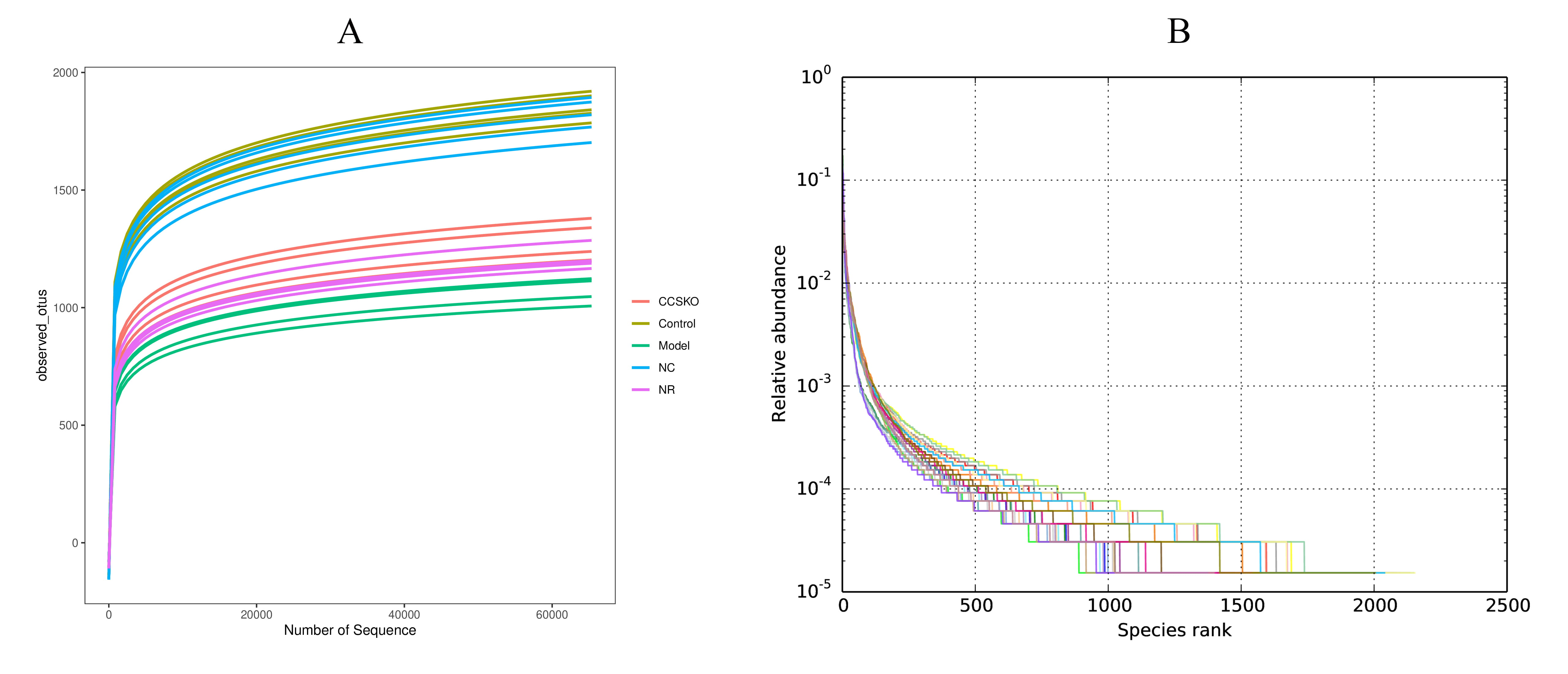
The dilution curve was constructed by random sampling of the sequencing sequence to extract the number of sequences and the number they represented (Fig. 2). It can be seen from Fig. 1A that when the sequence number reaches 55,000, the curve tends to be flat, indicating that our sequencing data volume is asymptotically reasonable. Fig. 1B shows the species richness and evenness of the samples in both horizontal and vertical directions.
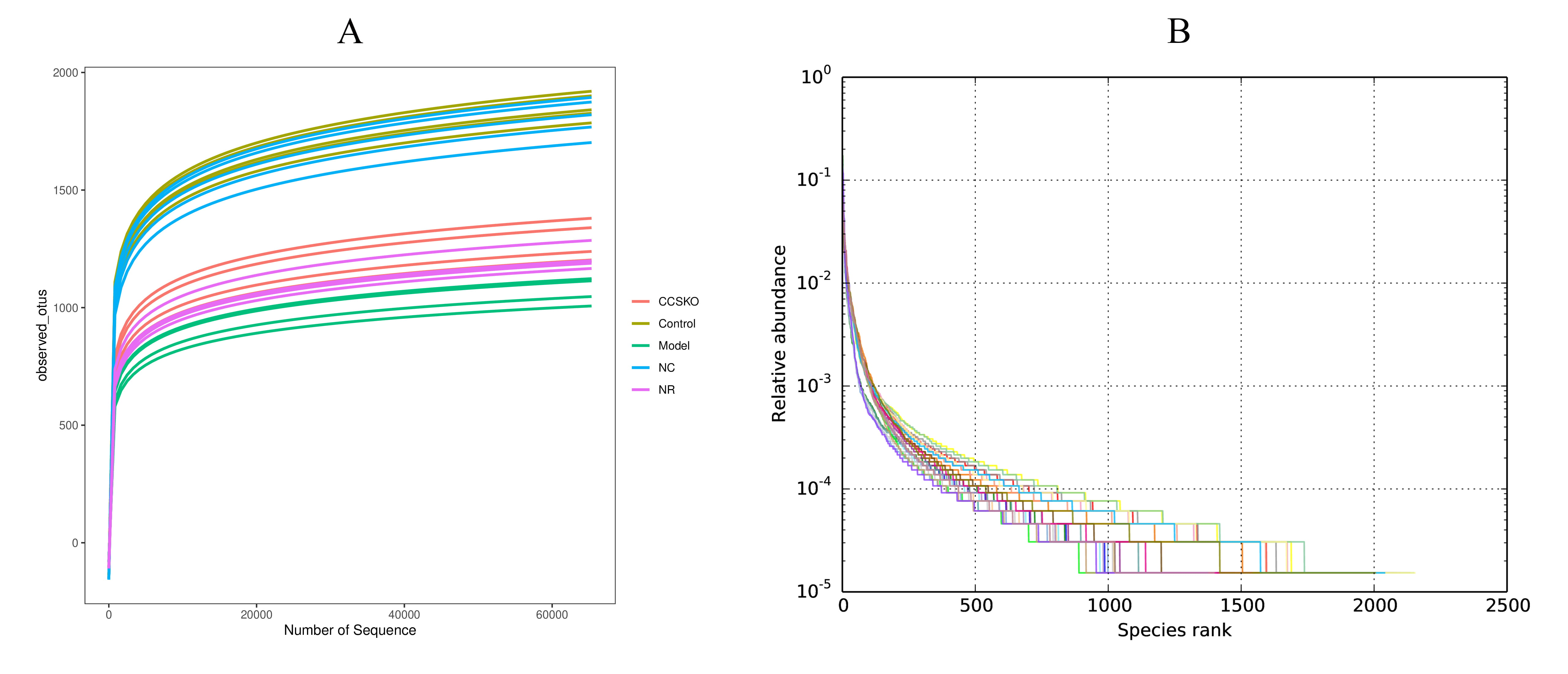 Fig. 2.
Fig. 2.(A) Dilution curve (B) Rank Abundance curve. The former indirectly reflects the rationality of the size of our sequencing data and the species Abundance in the sample. On the rank abundance curve, the wider the curve span, the higher the relative abundance of the species on the horizontal direction, and the smoothness of the curve indicates the homogeneity of the species on the vertical direction.
Diversity can reflect the richness and evenness of the microbial community, including Chao1 index, Ace index, Shannon index and Simpson index. Among them, Chao1 index and Ace index reflect the number of species in the community, without considering the abundance of each species in the community. Shannon index and Simpson index reflect community diversity, which is affected by species richness and species evenness in the sample community. The diversity index is shown in Table 1.
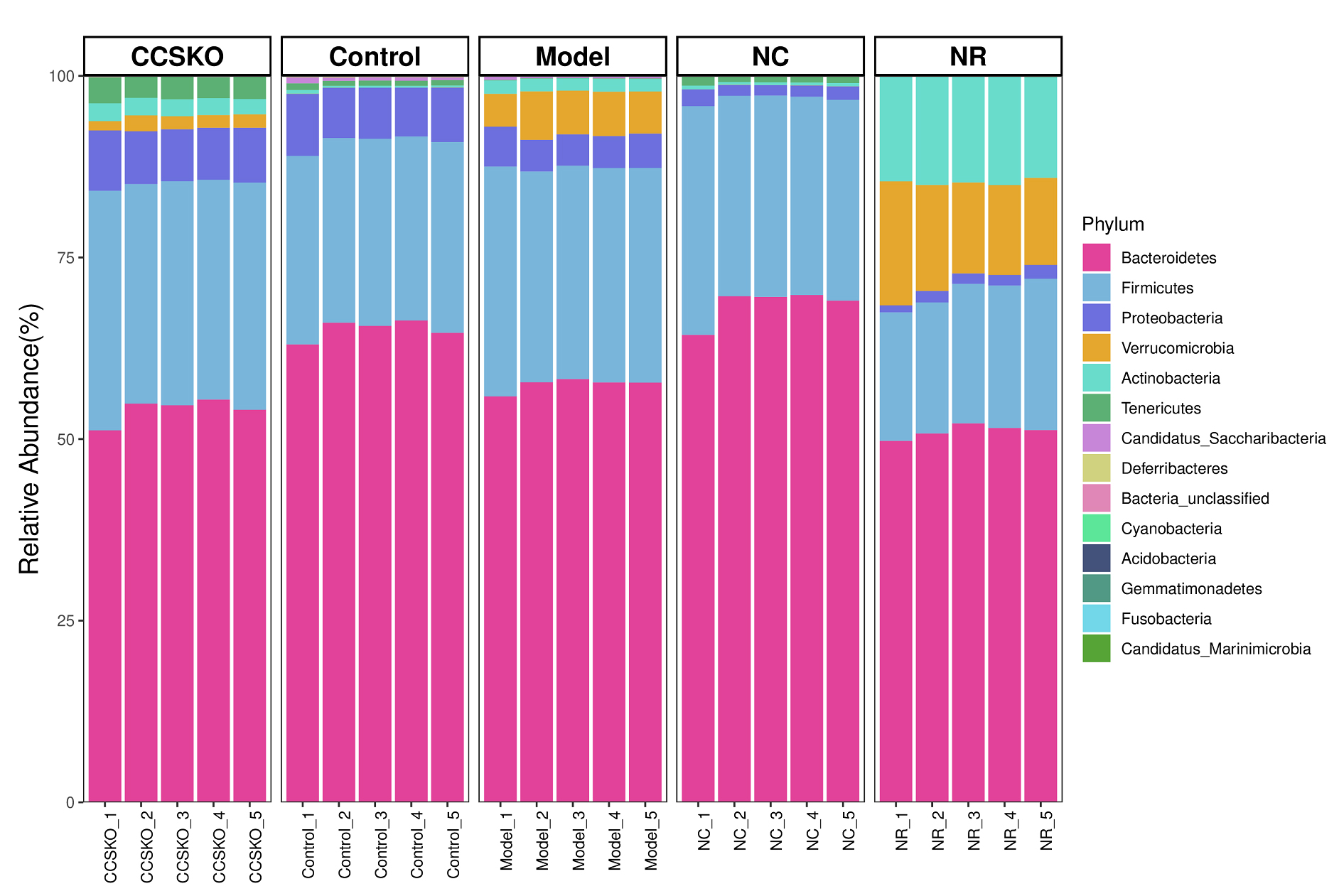
A total of 14 phyla, 27 classes, 53 orders, 92 families, 190 genera and 263 species were identified from 25 samples from 5 subgroups. At the gate level, Bacteroidetes is the most abundant in each group, Firmicutes second. The sum of Bacteroidetes is more than 70% in each group (Fig. 3), and the NC group has the highest abundance, which is 96.85%. Proteobacteria ranked the fifth in abundance in NR group (1.46%), and the third in the other four groups (CCSKO, 7.49%, Control, 7.34%, Model, 4.66%, and NC, 1.72%). It is worth noting that the abundance of Verrucomicrobia and Actinobacteria in the NR group was much higher than that of the other four groups, with the abundance of 13.71% and 14.52%, respectively. Another thing to note is that Acidobacteria only does not exist in the Control group, but is distributed in the other four groups, and its abundance is all 0.01%.
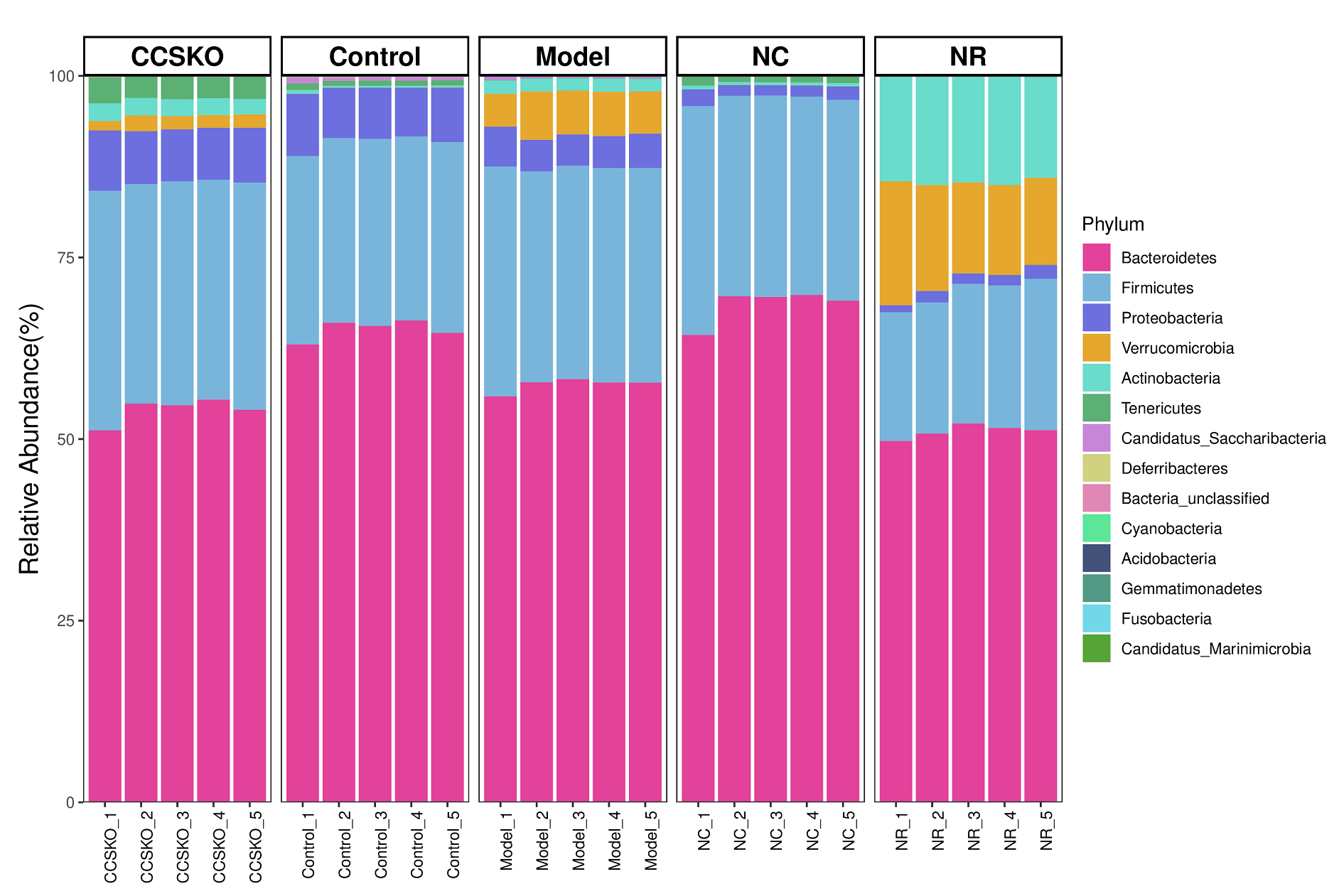 Fig. 3.
Fig. 3.Species distribution among groups based on the Bray-Curtis Distance at the gate level.
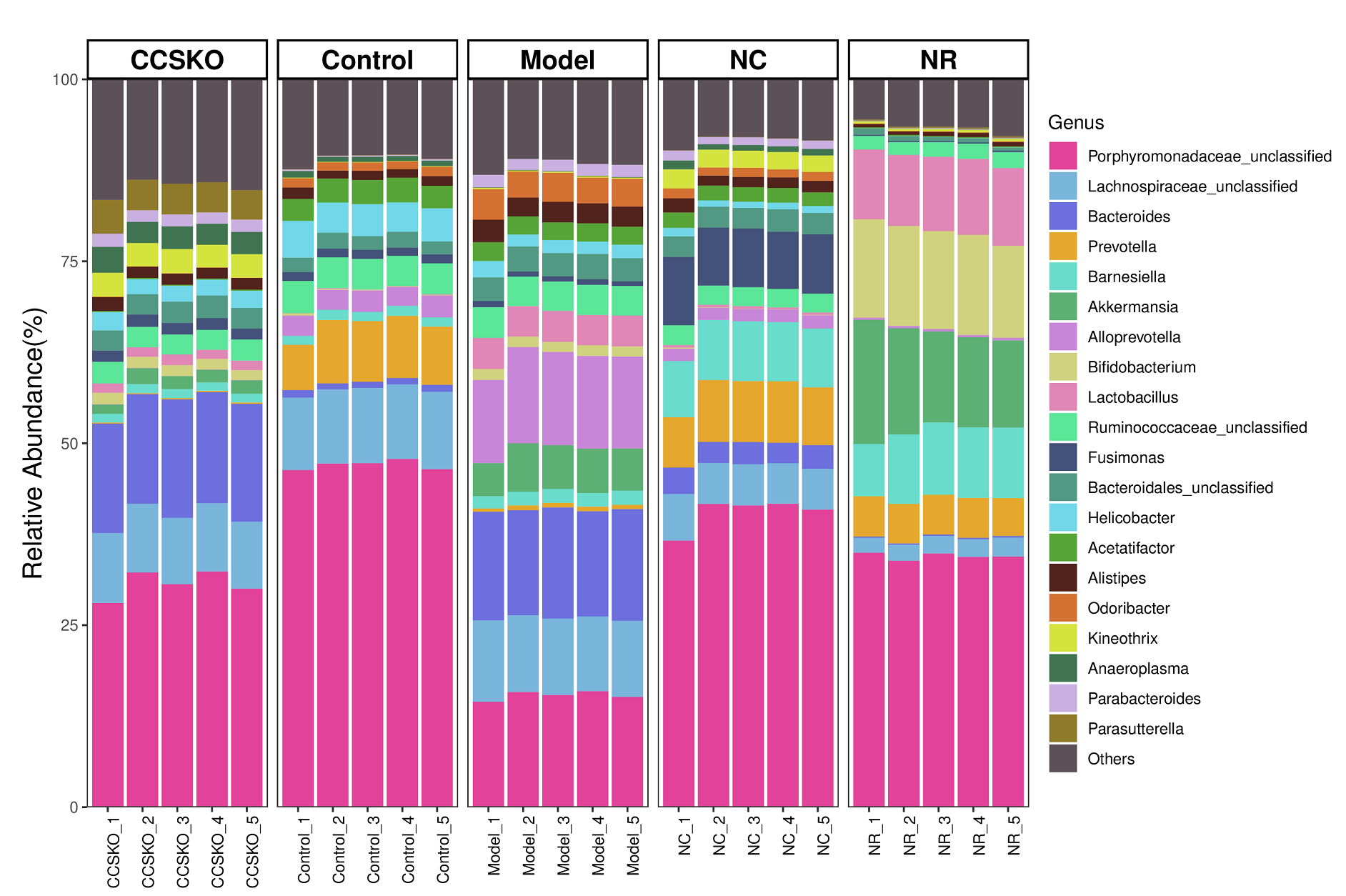
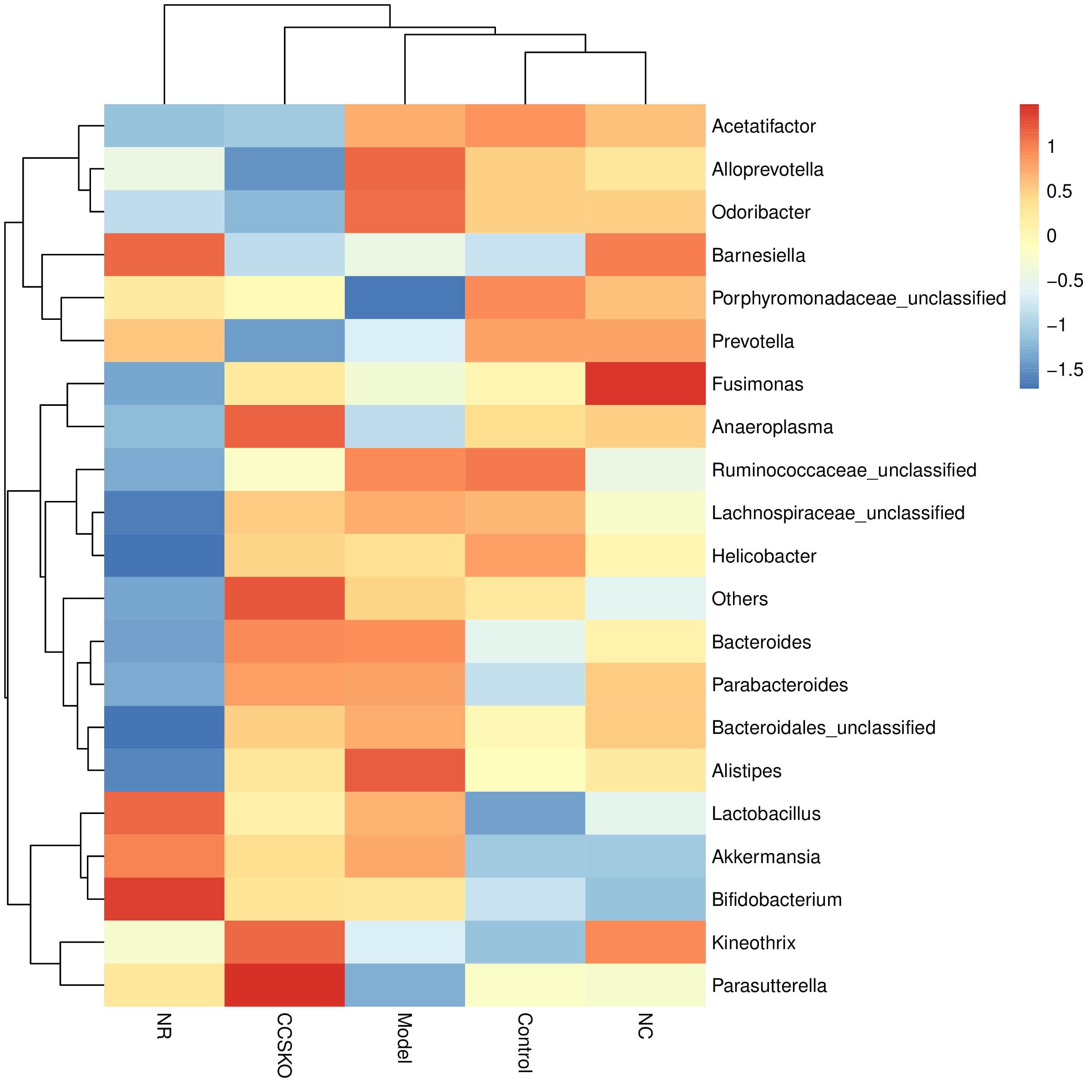
At the genus level, the highest abundance was Porphyromonadaceae_unclassified (Fig. 4). There was a great difference between the five groups, with the highest abundance being 47.01% in the Control group and the lowest being 15.37% in the Model group. The Bacteroides in CCSKO group and Model group were significantly higher than the other two groups. Helicobacter and Acetatifactor were significantly higher in the Control group than in the other three. In the NR group, Bifidobacterium and Lactobacillus were also significantly higher than the other four groups. It is worth noting that each microflora in the CCSKO group is more diverse, and Parasutterella distribution is significantly higher than that of the other four subgroups.
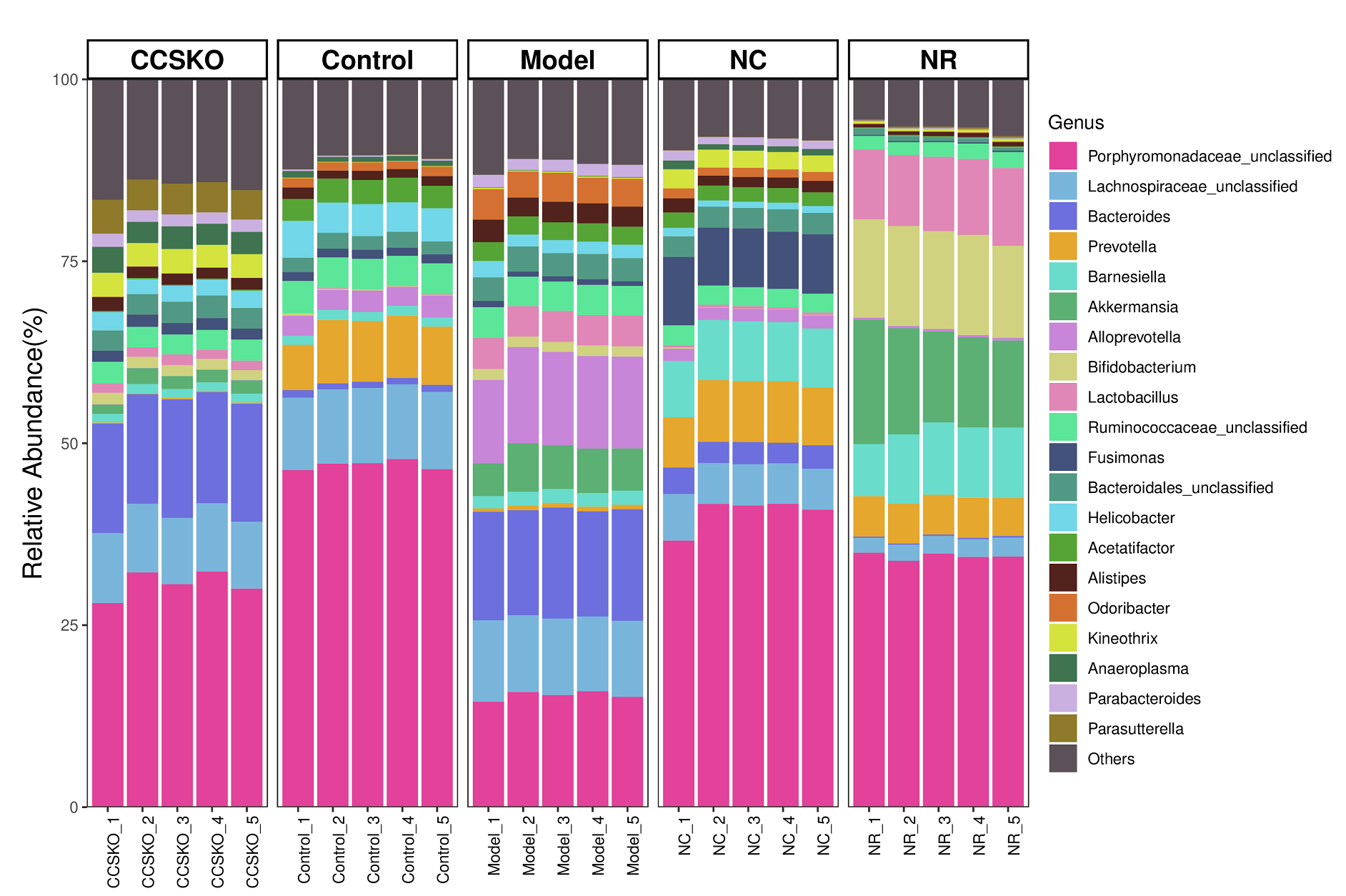 Fig. 4.
Fig. 4.Species distribution among groups based on bray-Curtis Distance at genus level.
According to the relative abundance table of samples, the community composition data of the 20 communities with the highest relative abundance at each level of classification were clustered according to the abundance distribution of taxa or the similarity degree between samples (Fig. 5). The taxa and samples were sequenced respectively according to the clustering results, and presented by the heat map. It can be seen from the figure that the relative abundance of the most dominant genera of the NR group is low, while that of the CCSKO group is high. From the clustering results, the tree graph based on Bray-Curtis Distance is similar to the clustering heat graph.
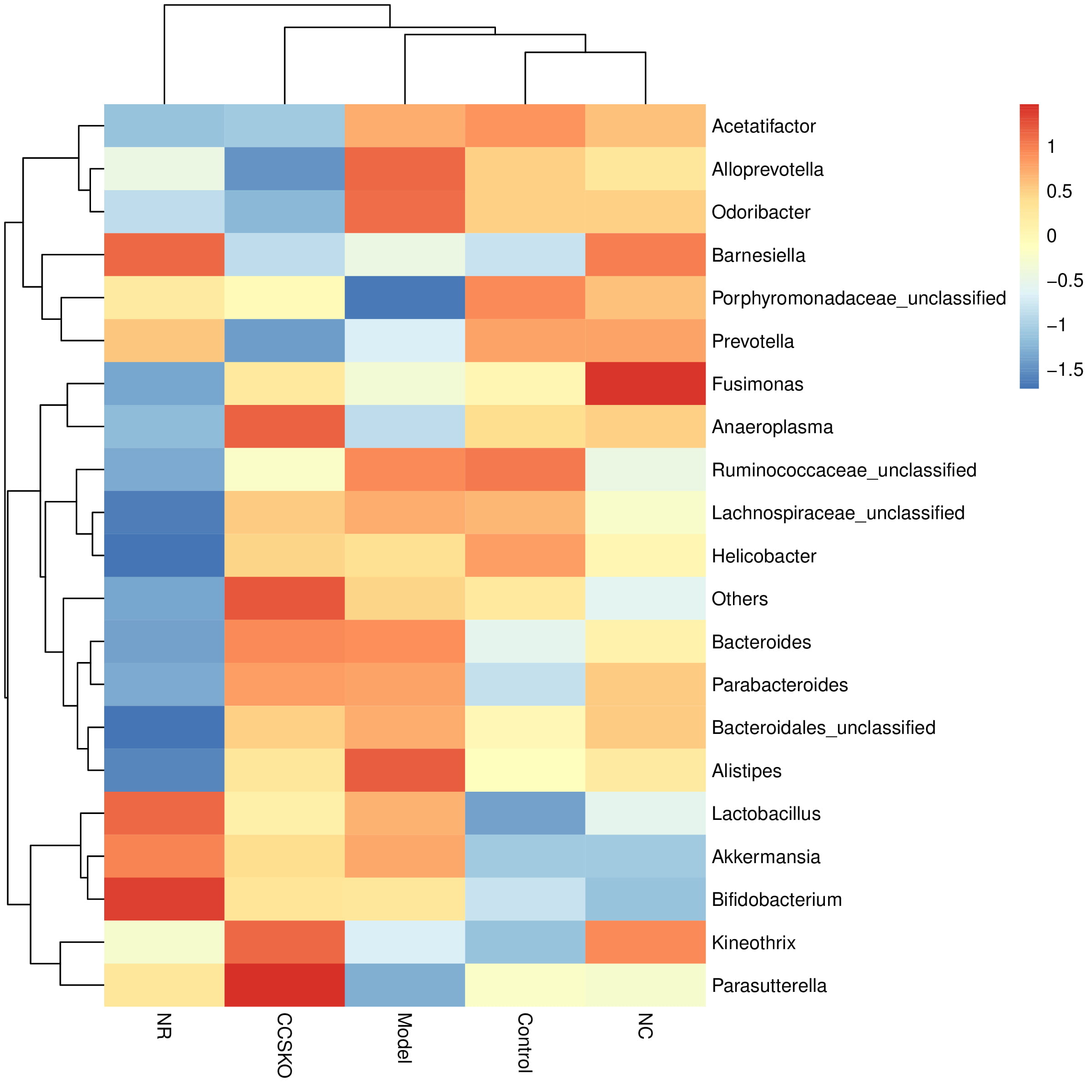 Fig. 5.
Fig. 5.Clustering heat map of relative abundance of species at genus level.
We used similarity analysis to check the rationality of grouping. Anosim Analysis (Analysis of similarities) is based on the sample distance matrix obtained by the Unweighted Unifrac algorithm, and the differences within and between the sample groups are determined by sorting the sample distance grade, and the statistical significance of the differences between the original sample groups is evaluated by the replacement test. The R values of ANOSIM were all 1, indicating that the greater the difference between groups, and the smaller the difference between groups, the better the grouping effect.
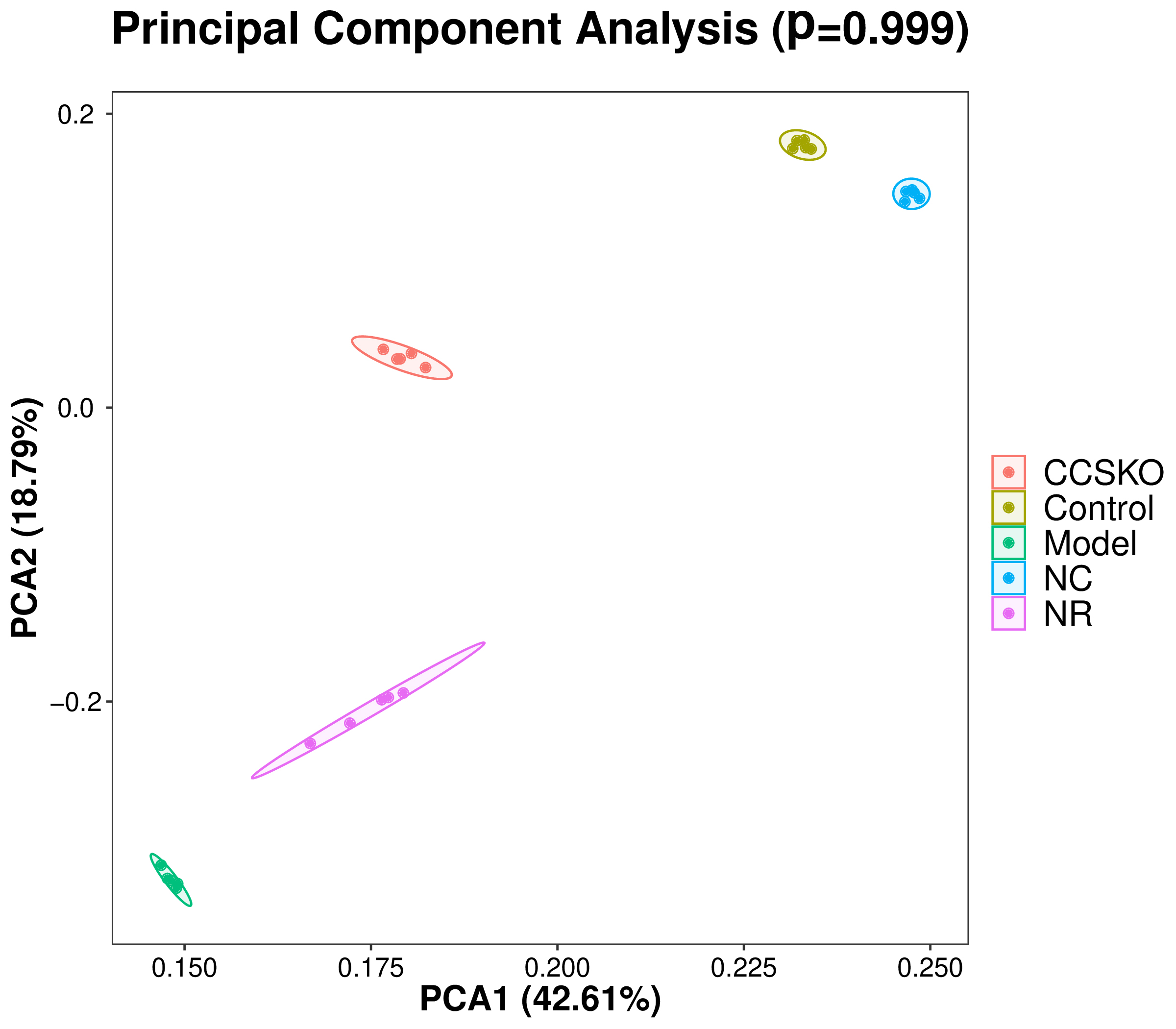
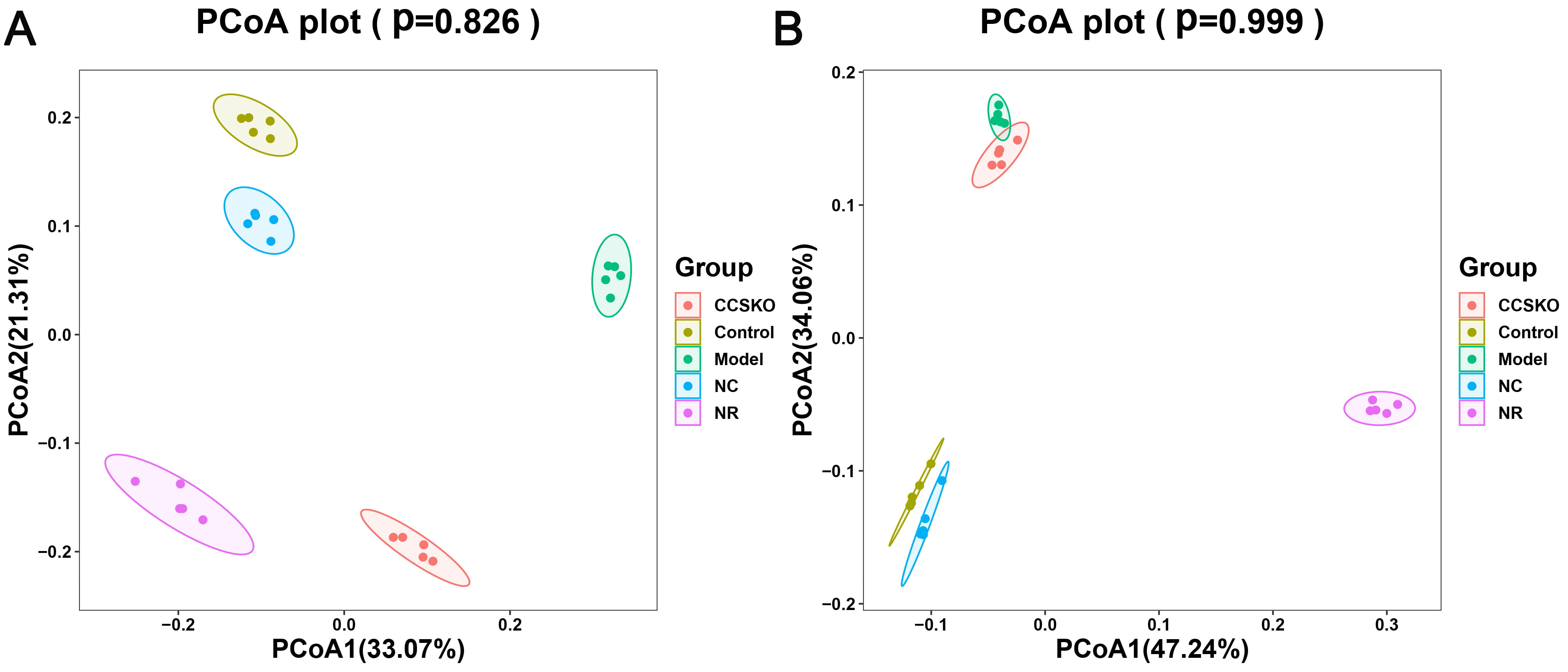
It can be seen from the PCA diagram that the biological duplication between samples is good and the composition of the same sample is similar (Fig. 6). The effects of camphor seed kernel oil (CCSKO) on intestinal flora of hepatic lipidosis mice were determined by analyzing the 16S rRNA gene sequence of intestinal microbial samples from each group. The principal coordinate analysis (PCoA) based on UniFrac distance showed that the intestinal microbial community of each experimental group had obvious clustering. CCSKO was more closely clustered than NR group, indicating that the addition of camphor seed kernel oil (CCSKO) significantly changed the bacterial flora structure (Fig. 7).
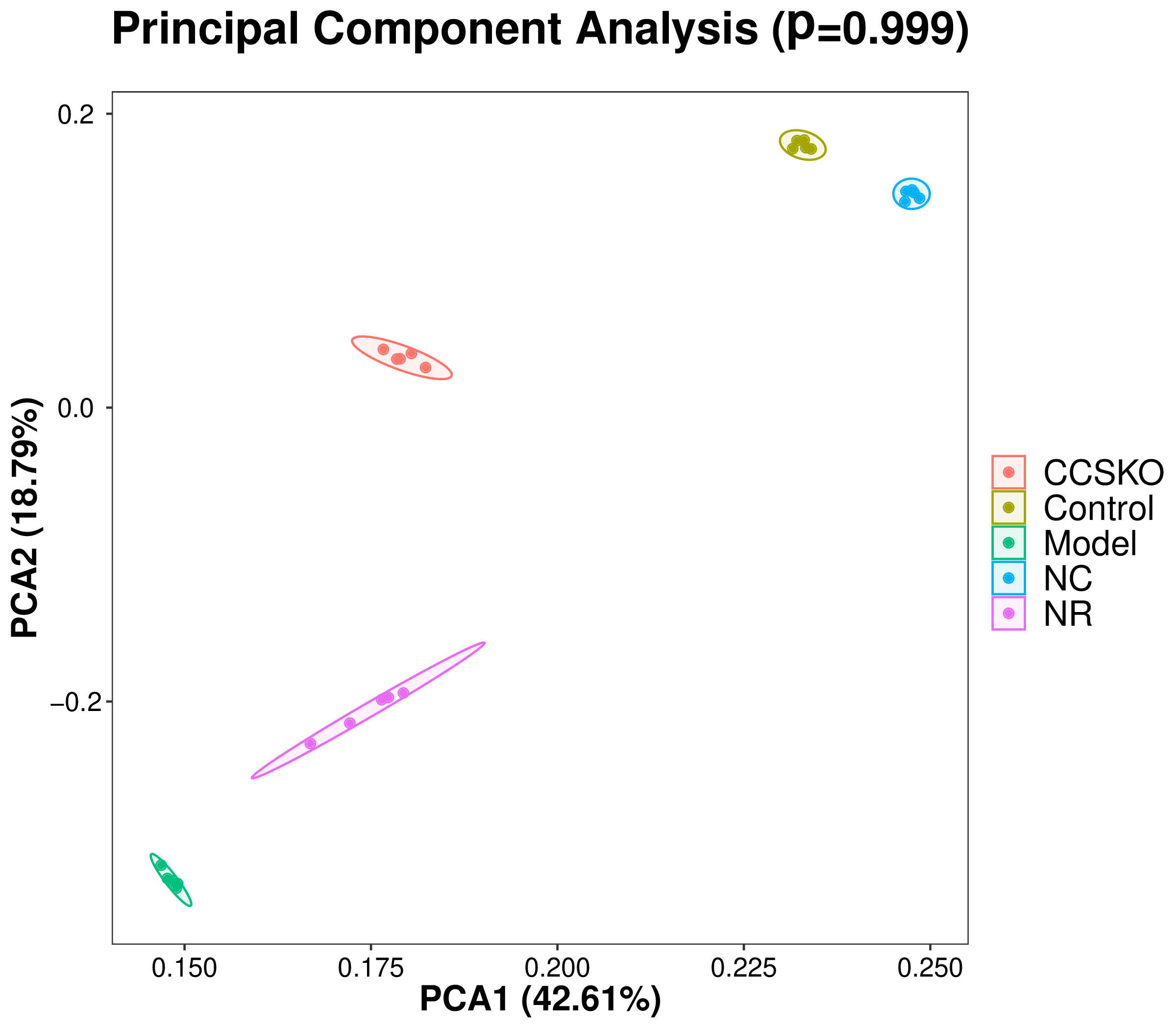 Fig. 6.
Fig. 6.Each point in the PCA analysis diagram of samples represents a sample, and samples of the same group are represented by the same color. The closer the distance between the points in the figure indicates that the species composition is more similar, and the closer the distance between the same group of samples and other groups of samples is far, indicating that the biological repetition of samples is better or the composition of the same sample is similar.
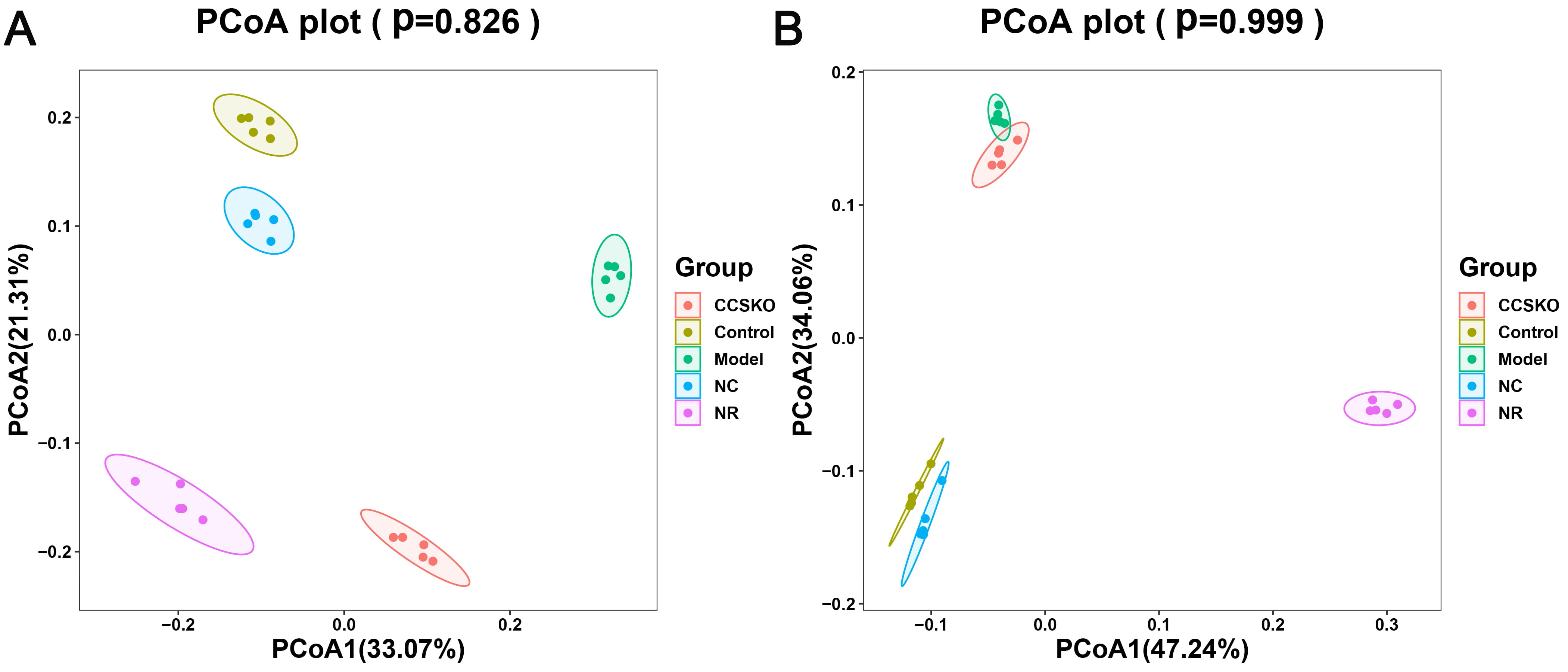 Fig. 7.
Fig. 7.PCoA analysis of samples Figure A: Analysis it based on the weighted Unifrac distance matrix B: Analysis of the distance between the two colors based on the unweighted Unifrac distance matrix, showing their similarities.
Camphor seed kernel oil (CSKO) has high nutritional value and contains a large number of biologically active compounds which have well-known effects on blood fat, blood sugar level, body fat deposition and quickly supply energy. The contents of decanoic acid and lauric acid in CSKO are relatively high, accounting for more than 90%, which is a natural medium carbon chain oil resource. Medium carbon chain triglyceride not only has the function of providing energy and nutrition to food, but also does not affect food flavor. And it has a unique metabolic mode and specific health care role, so it has been popular in recent years. However, it remained unclear whether MCT produces any effect on NAFLD and gut microbiota dysbiosis. In the present study, after oral administration of Camphor seed kernel oil, there was an obvious improvement in hepatic steatosis and gut microbiota dysbiosis in CSKO-fed mice.
The liver plays a core role in lipid metabolism throughout the body. NAFLD, a metabolic syndrome, is characterized by imbalance of hepatic lipid metabolism with accumulating fat in the liver. Its complex regulation mechanisms include high-fat diet, obesity and so on. Due to excessive intake of nutrients, new lipids are synthesized from the beginning and triglycerides (TG) tend to accumulate, which can cause liver cell damage [17]. In the present study, the liver index of the NR group increased, while CCSKO treatment exerted ameliorative effects on the problem. The level of TG and TC in NAFLD mice were higher than those of CCSKO group. These results suggested that CCSKO could be used to prevent fat deposition and reduce the level of TG and TC in the liver, indicating its role in lipid homeostasis. A previous study also obtained similar results and speculated that the effect of CCSKO might be associated with its unique fatty acid compositions [18].
A large number of the gut microbiota contribute to the regulation of nutrient metabolism and self-development of the body. However, the gut microbiota disorder is one of the important pathogeneses of NAFLD. It is reported that the Bacteroidetes and Firmicutes act as the main components of gut microbial community [19]. In the present study, NAFLD greatly influenced the diversity and richness of the gut bacteria compared with the Control group, resulting in dysregulation of the microbiota. Though we observed that Firmicutes and Bacteroidetes accounted for a large proportion in all groups, there was a difference of the gut microbiota composition between CCSKO group and NR group. Besides, accumulation of studies have demonstrated that the increased abundance of Verrucomicrobia is positively related with nutrient metabolism [19, 20], but we found that NR group and Model group had higher abundance of Verrucomicrobia compared with CCSKO group. With CCSKO treatment, the relative abundance of Proteobacteria increased to close to that of the Control group. A study showed that fat feeding could promote certain gut bacteria such as Lactobacillus [21]. The results of this experiment showed that the structure of the liver tissue in the CCSKO group was similar to that in normal control group, and the structure was intact without degeneration and necrosis. In the genus level, compared with the model group, CCSKO significantly reduced the relative abundance of Lactobacillus. As a result, CCSKO can change the gut microbiota structure of NAFLD mice. The Bacteroides was only dominated in the CCSKO group. The CCSKO treatment might significantly increase the abundance of Bacteroidetes. A previous study found that the Bacteroides were involved in many important metabolic activities in the human colon, including carbohydrate fermentation, utilization of nitrogenous substances, and biotransformation of bile acids and other steroids [22].
NAFLD is a challenging disease caused by multiple factors, which might partly explain why it still remains orphan of an adequate therapeutic strategy. Oxidative stress is a redox imbalance between the rate of production of reactive oxygen and nitrogen species and their consumption by antioxidants, which is quite strictly linked to the lipotoxic liver injury from fatty acids. The interaction between the gut epithelia and some commensal bacteria induces the rapid generation of reactive oxygen species (ROS) [23]. The gut-liver axis may enhance the natural interactions between intestinal bacteria/bacterial products and hepatic receptors, thus promoting the following cascade of events: oxidative [24, 25]. Our experimental results showed that CCSKO significantly changed the bacterial flora structure, including increasing the richness of the microflora and the proportion of probiotics (Bifidobacterium and Lactobacillus). Probiotic able to modulate the microbiota with benefits for the health of humans. Studies report that bacterial endotoxins such as lipopolysaccharides (LPS) produced by the growth of Gram-negative bacteria, plays an important role in the pathogenesis of NAFLD. Fukunishi’s research shows that LPS accelerate hepatic steatosis in the development of nonalcoholic fatty liver disease in Zucker rats [26], and, histological examination revealed the most severe hepatic steatosis in the LPS group compared with the other groups. EF Mostafa’s research shows that endotoxemia may contribute in the pathogenesis of NAFLD especially in non-obese patients [27]. LPS also further leads to the occurrence of oxidative stress as a consequence of enhanced intestinal permeability [28]. Oxidative stress produces lipid peroxidation and activates inflammatory cytokines resulting in NASH [29]. We all know that probiotics are bacteria which promote good health by reducing pathogens. In addition, probiotics diminish the development of NAFLD, suggesting a relationship between the enteric flora and liver injury [30]. The results of our previous research also suggested that MCFA treatment could alleviative oxidative stress-induced cytotoxicity [5]. CCSKO contained a high level of medium-chain fatty acids (MCFA, more than 90%), such as capric acid and lauric acid [31]. Therefore, it is very meaningful to investigate whether CCSKO able to protect hepatic cells from damaging effects caused by gut microbiota and oxidative stress, through an enhanced gut–liver axis in further experiments.
In summary, the results showed that being the natural resources of MCTs, CCSKO effectively reduced the level of TG and TC in the liver, maintained lipid homeostasis and modulated the dysregulation of the microbiota induced by NAFLD to some extent. These finding indicate a strong contribution in shaping the gut microbiota by treatment groups, shedding light on impact of camphor tree seed kernel oil on gut microbiota, which is associated with human growth and health.
Not applicable.
BW, SS, WD and JW collected and processed the data. All authors contributed to drafting and preparation of the manuscript.
All operations to mice in this study followed the standard practices of animal care and use. The practices related to the animals in this study were approved by the Binzhou University Animal Care and Use Committee (BZUCUC number 20180812).
Not applicable.
The authors gratefully acknowledge the financial supported by Shandong Provincial Natural Science Foundation ZR2019PH097 and ZR2021QD082.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.