










1 Laboratory of Molecular Immunology, Federal State Budgetary Scientific Institution “Research Institute of Fundamental and Clinical Immunology" (RIFCI), 630099 Novosibirsk, Russia
2 First Therapy Department, Federal State Budgetary Institution “Novosibirsk Research Institute of Tuberculosis", Ministry of Health of Russian Federation, 630099 Novosibirsk, Russia
Academic Editor: Hongwei Yao
Abstract
Background: Tumor necrosis factor (TNF) plays an important role in immune responses to the causative agent of tuberculosis, Mycobacterium tuberculosis. Additionally, TNF can also mediate many negative disease manifestations. The aim of this study was to assess the contribution of anti-TNF autoantibodies to the pathogenesis of active pulmonary tuberculosis (TB). Methods: The levels of anti-TNF autoantibody classes and subclasses were determined by applying enzyme-linked immunosorbent assays (ELISAs). The levels of TNF and of its soluble receptors were also evaluated using commercial ELISA kits. Results: The levels of both types of soluble TNF receptors were lower patients with TB than in healthy donors. Patients with TB had higher titers of immunoglobulin (Ig)G class and IgG3 subclass anti-TNF autoantibodies in comparison with healthy donors. Patients who had a disseminated TB infection had higher TNF level and IgG, IgG1 and IgG3 autoantibody titers compared with patients who had a localized TB infection. Conclusions: Changes in the titers of anti-TNF autoantibody classes and subclasses were noted in patients with TB, suggesting their possible contribution to the disease pathogenesis of TB.
Keywords
- anti-cytokine autoantibodies
- tumor necrosis factor
- pulmonary tuberculosis
- enzyme immunoassay
The pro-inflammatory cytokine tumor necrosis factor (TNF) plays a key role in
the host immune response to the tuberculosis pathogen Mycobacterium
tuberculosis (M. tuberculosis). TNF performs a number of important functions
during infection; for example, it stimulates T-lymphocyte and monocyte
chemotaxis, phagocyte activation, macrophage apoptosis, dendritic-cell maturation
followed by T-cell activation, interferon (IFN)-
Soluble type 1 (sTNFR1, p55) and 2 (sTNFR2, p75) TNF receptors are identical to
the extracellular cytokine-binding domains of membrane TNF receptors; they can
compete with membrane-associated TNF receptors for binding TNF and thus inhibit
TNF bioactivity [6]. Human anti-cytokine autoantibodies (ACAABs) are polyclonal
and belong mainly to the IgG class [7]. Their binding avidities range from 1
So, considering prominent role of tumor necrosis factor in protective immunity against M. tuberculosis, its involvement in negative manifestations of pulmonary tuberculosis and important role of anti-cytokine autoantibodies in modulating biological activity of corresponding mediators and in disease pathogenesis in a number of nosologies, the aim of this study was to obtain new data on the levels of various classes and subclasses of anti-TNF autoantibodies in patients with TB and healthy individuals.
The group of patients with TB included 45 individuals who were hospitalized in the First Therapy department of Novosibirsk Research Institute of Tuberculosis. The diagnosis is formed on the basis of complaints, anamnesis, physical examination, sputum culture for the presence of Mycobacterium tuberculosis in liquid and solid media, Xpert MTB/RIF test, positive skin test, X-ray studies and the dynamics of the process against the background of anti-tuberculosis therapy. HIV infection was the exclusion criterion for the study. Several patients were diagnosed with hepatitis B and C, while they had no clinical or laboratory manifestations. Most patients were receiving anti-tuberculous therapy, but instead of whether were patients receiving therapy at the time of blood collection or not, all of them had active pulmonary tuberculosis. There were no patients with miliary tuberculosis in the study. None of the patients included in the study received steroid therapy.
Blood samples were obtained from 150 healthy donors at Novosibirsk Blood Center. Blood sampling of patients with TB and healthy individuals was performed after obtaining their informed consent.
The study protocol was approved by the local ethics committee at the Research Institute of Fundamental and Clinical Immunology.
To obtain serum samples, 4 mL of venous blood per subject was collected into
vacuum tubes with Vacuette Z Serum Clot Activator (Greiner Bio-One GmbH,
Kremsmunster, Austria), decanted for 1 h, and centrifuged at 2.500 rpm for 20
min. Serum was then isolated and stored at –20
Serum titers of anti-TNF autoantibodies were evaluated by conducting solid-phase ELISAs using recombinant human TNF (Institute of Medical Biotechnology, The State Research Center of Virology and Biotechnology VECTOR, Berdsk, Russia) as described in our previous study [16]. Antibodies against immunoglobulin classes and subclasses, each conjugated with horseradish peroxidase, (Bialexa, Moscow, Russia) were diluted as follows: 1:499 for anti-IgM (clone 1H9c); 1:1.599 for anti-IgG (clone 2M9c); 1:499 for anti-IgA (clone 1A9c); 1:99 for anti-IgG1 (clone 2C11c); and 1:24 for anti-IgG2 (clone 52G1), anti-IgG3 (clone 5G12c), and anti-IgG4 (clone 5C7c).
Statistical data analysis was performed using GraphPad Prism 6 software
(GraphPad Software, San Diego, CA, USA). Independent samples were tested for
statistical significance using non-parametric Mann-Whitney test. Data are
expressed as medians and interquartile ranges. We evaluated correlation among
parameters with the Spearman test. Differences with p
The groups of patients with TB and healthy donors had comparable compositions, in terms of age and sex. The study group characteristics are presented in Table 1.
| Characteristics of the patients with TB and the healthy individuals groups | Patients with TB (n = 45), n (%) | Healthy donors (n = 150), n (%) | p-value |
| Sex, M/F | 27/18 (60/40) | 83/67 (55.3/44.7) | 0.611 |
| Age, median [IQR] | 31 [25–44] | 30.5 [23–45] | 0.512 |
| Disseminated infection | 26 (57.8) | - | - |
| MBT+ | 9 (34.6) | - | - |
| MBT− | 17 (65.4) | - | - |
| Localized infection | 19 (42.2) | - | - |
| MBT+ | 6 (31.6) | - | - |
| MBT− | 13 (68.4) | - | - |
| MBT+: patients with TB, from whom M. tuberculosis was isolated; MBT–: patients with TB, from whom M. tuberculosis was not isolated; Localized infection: patients with TB with infection occupying one or two segments of lungs; Disseminated infection: patients with TB with infection occupying more than two segments of lungs. | |||
The serum levels of TNF and of its soluble type 1 and 2 receptors in patients with TB and healthy individuals were assessed by ELISA. There was no difference in the TNF level between the patients with TB and the healthy individuals (Fig. 1A). A comparison of the serum level of TNF among patients with TB revealed higher TNF level in patients with disseminated infection (infection occupying more than two segments of lungs) than in patients who had a localized infection (infection occupying one or two segments of lungs) [21] (Fig. 1B) and in patients with disseminated infection from whom pathogen was isolated on bronchoscopy (the group of patients with most severe disease) than in other patients (Fig. 1C).
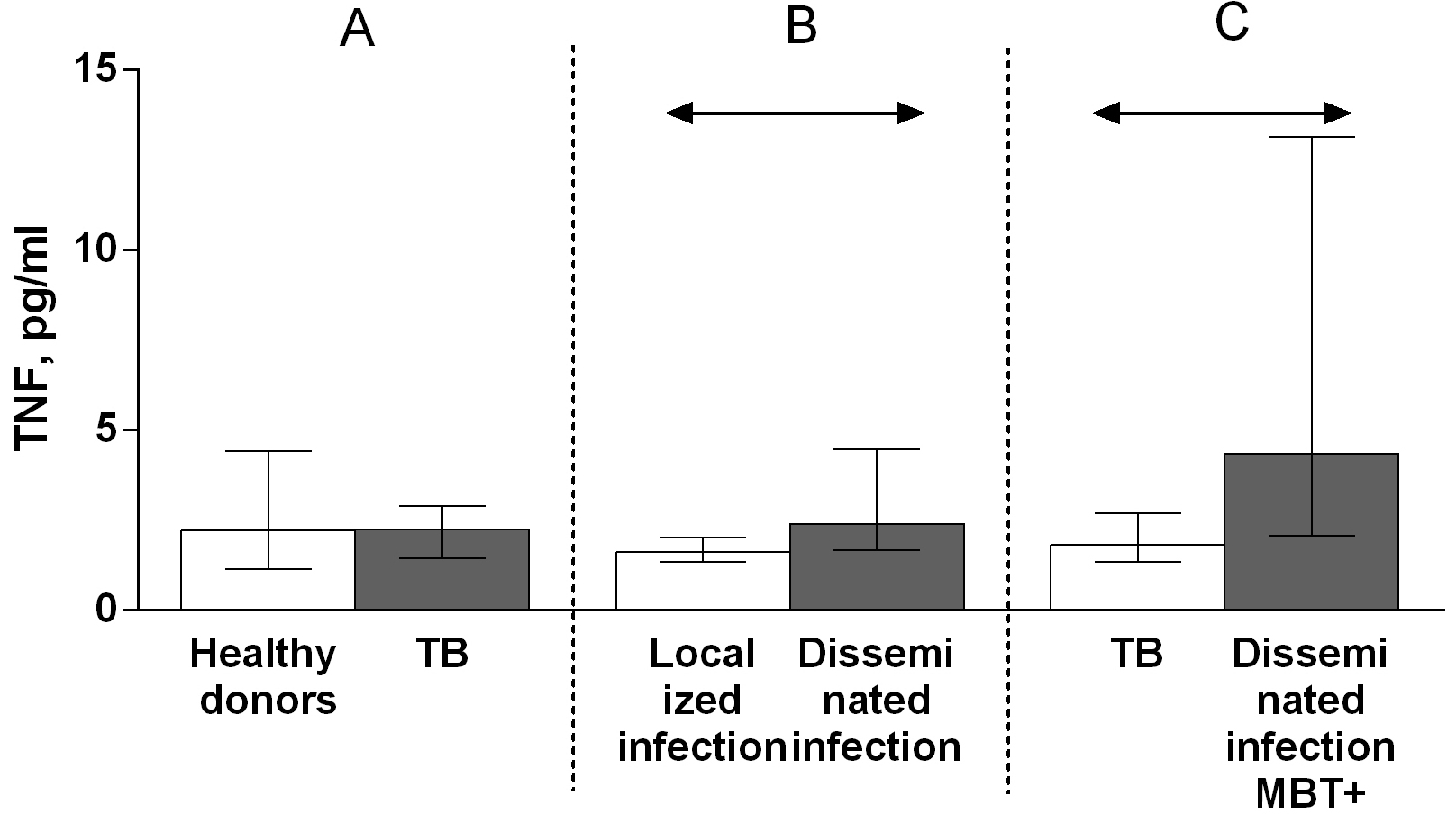 Fig. 1.
Fig. 1.Serum levels of TNF in patients with TB and healthy donors (A),
patients with TB who had localized or disseminated infection (B) and in MBT+ on
bronchoscopy patients with disseminated tuberculosis and other patients with TB
(C). Data are presented as the median (interquartile range). Double-arrow bar
indicate statistical significance. Mann–Whitney test, p
The serum levels of both types of soluble TNF receptors were lower in patients with TB than in healthy donors (Fig. 2A). There were no differences in sTNFR levels between patients with TB with disseminated and localized infection (Fig. 2B), at the same time sTNFR1 levels in patients with TB, from whom M. tuberculosis was isolated (MBT+) were higher in comparison with patients with TB, from whom M. tuberculosis was not isolated (MBT–) (Fig. 2C), sTNFR1 and sTNFR2 serum levels were higher in MBT+ on bronchoscopy patients with disseminated infection than in other patients with TB (Fig. 2D).
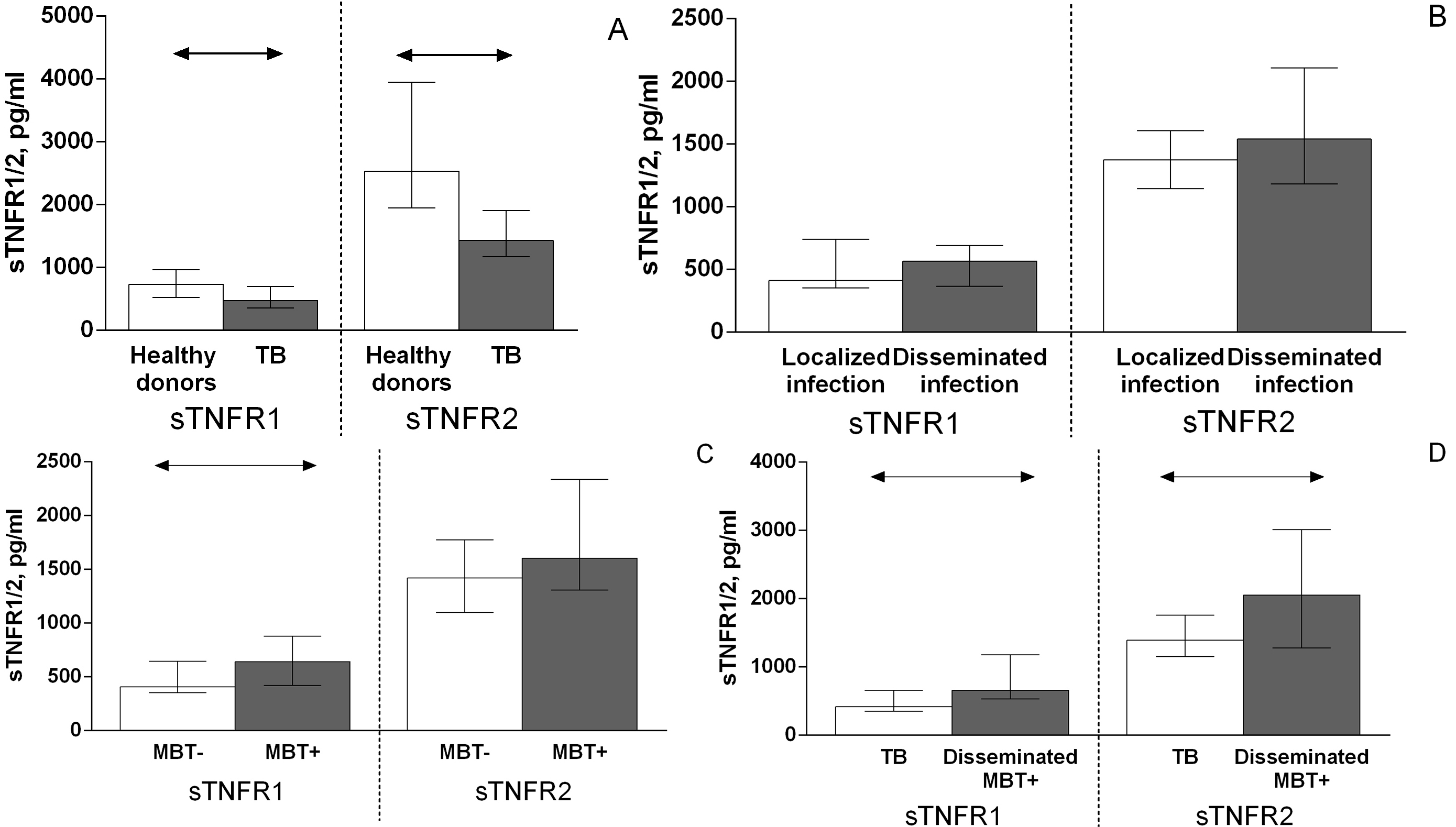 Fig. 2.
Fig. 2.Serum levels of TNF soluble type 1 and 2 receptors in patients
with TB and healthy donors (A), patients with TB who had localized or
disseminated infection (B), in patients with TB from whom Mycobacterium
tuberculosis was (MBT+) or was not (MBT–) isolated (C) and in MBT+ on
bronchoscopy patients with disseminated tuberculosis and other patients with TB
(D). Double-arrow bar indicate statistical significance. Data are presented as
the median (interquartile range). Mann–Whitney test, p
Serum titers of anti-TNF autoantibodies belonging to classes IgA, IgG, and IgM and subclasses IgG1, IgG2, IgG3, and IgG4 in patients with TB and healthy donors were assessed by performing ELISAs. The measured titers are expressed here in optical density international units.
The IgG autoantibody titer was significantly higher in patients with TB than in healthy donors (Fig. 3A). There were no significant differences in the IgM (Fig. 4A) and IgA (Fig. 5A) autoantibody titers between patients with TB and healthy individuals.
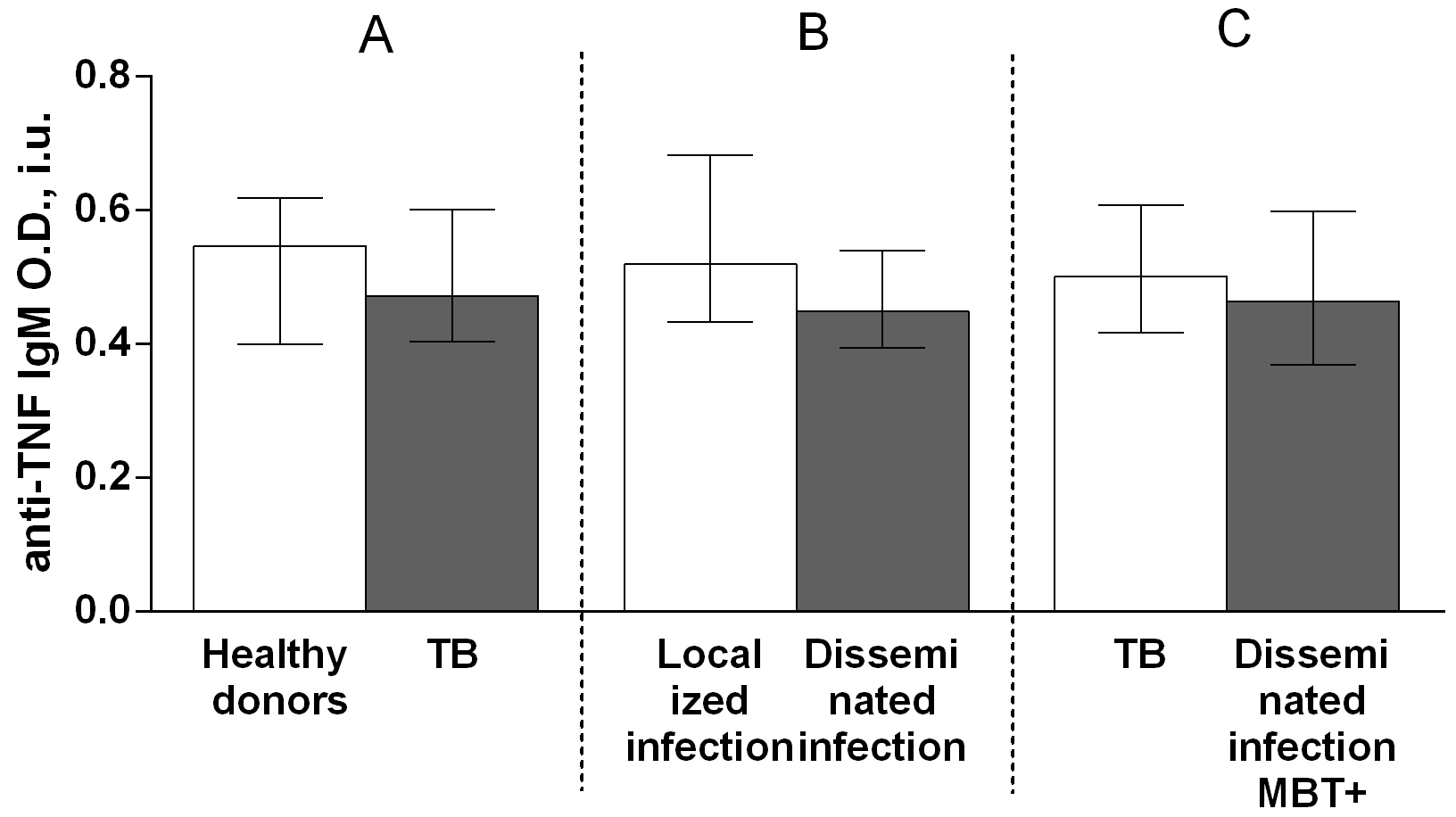 Fig. 3.
Fig. 3.Serum levels of anti-TNF IgG autoantibodies in patients with TB
and healthy donors (A), patients with TB who had localized or disseminated
infection (B) and in MBT+ on bronchoscopy patients with disseminated tuberculosis
and other patients with TB (C). Levels anti-TNF IgG autoantibodies are expressed
in international units of optical density (O.D., i.u.). Double-arrow bar indicate
statistical significance. Data are presented as the median (interquartile range).
Mann–Whitney test, p
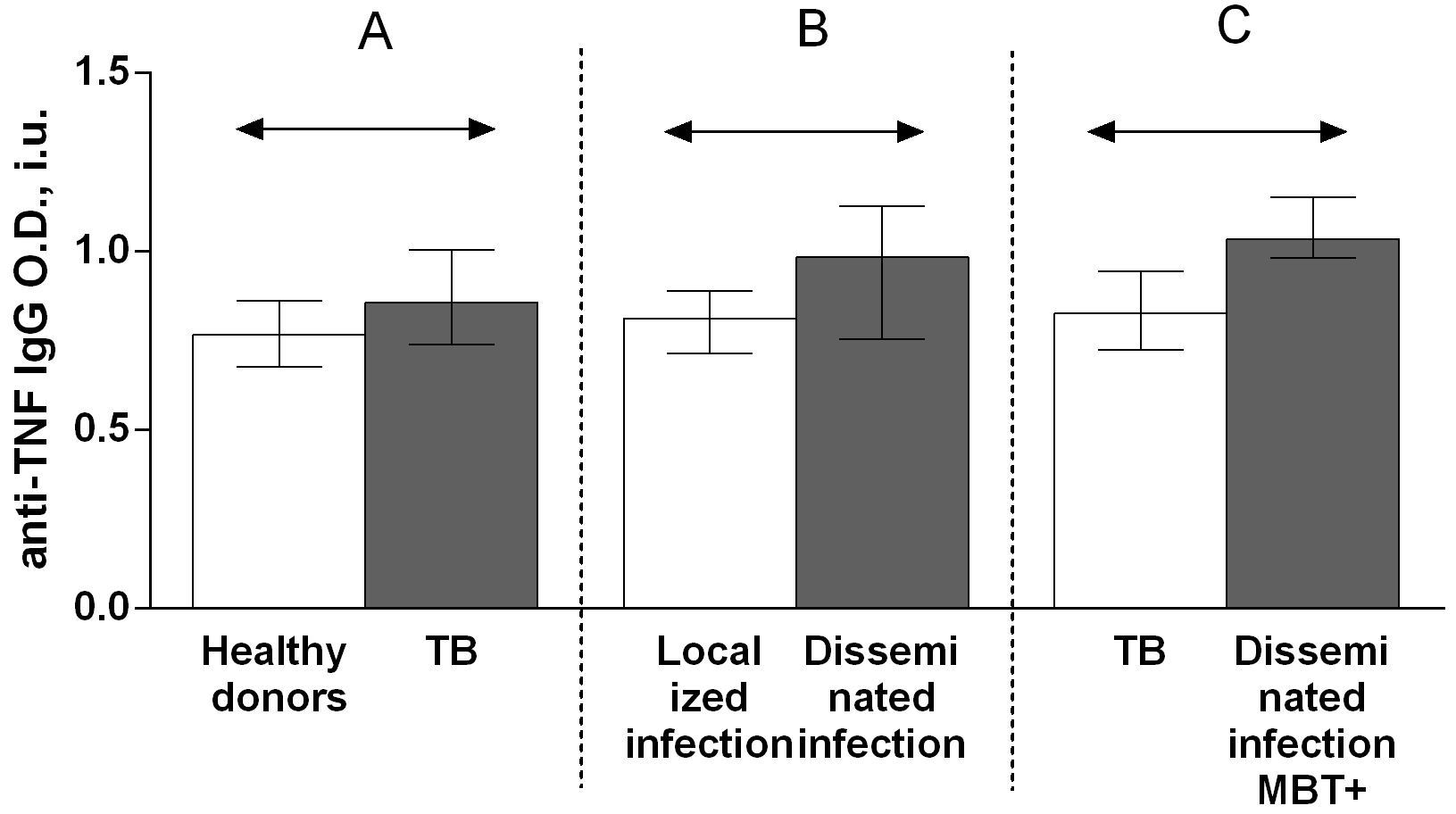 Fig. 4.
Fig. 4.Serum levels of anti-TNF IgM autoantibodies in patients with TB
and healthy donors (A), patients with TB who had localized or disseminated
infection (B) and in MBT+ on bronchoscopy patients with disseminated tuberculosis
and other patients with TB (C). Levels anti-TNF IgM autoantibodies are expressed
in international units of optical density (O.D., i.u.). Double-arrow bar indicate
statistical significance. Data are presented as the median (interquartile range).
Mann–Whitney test, p
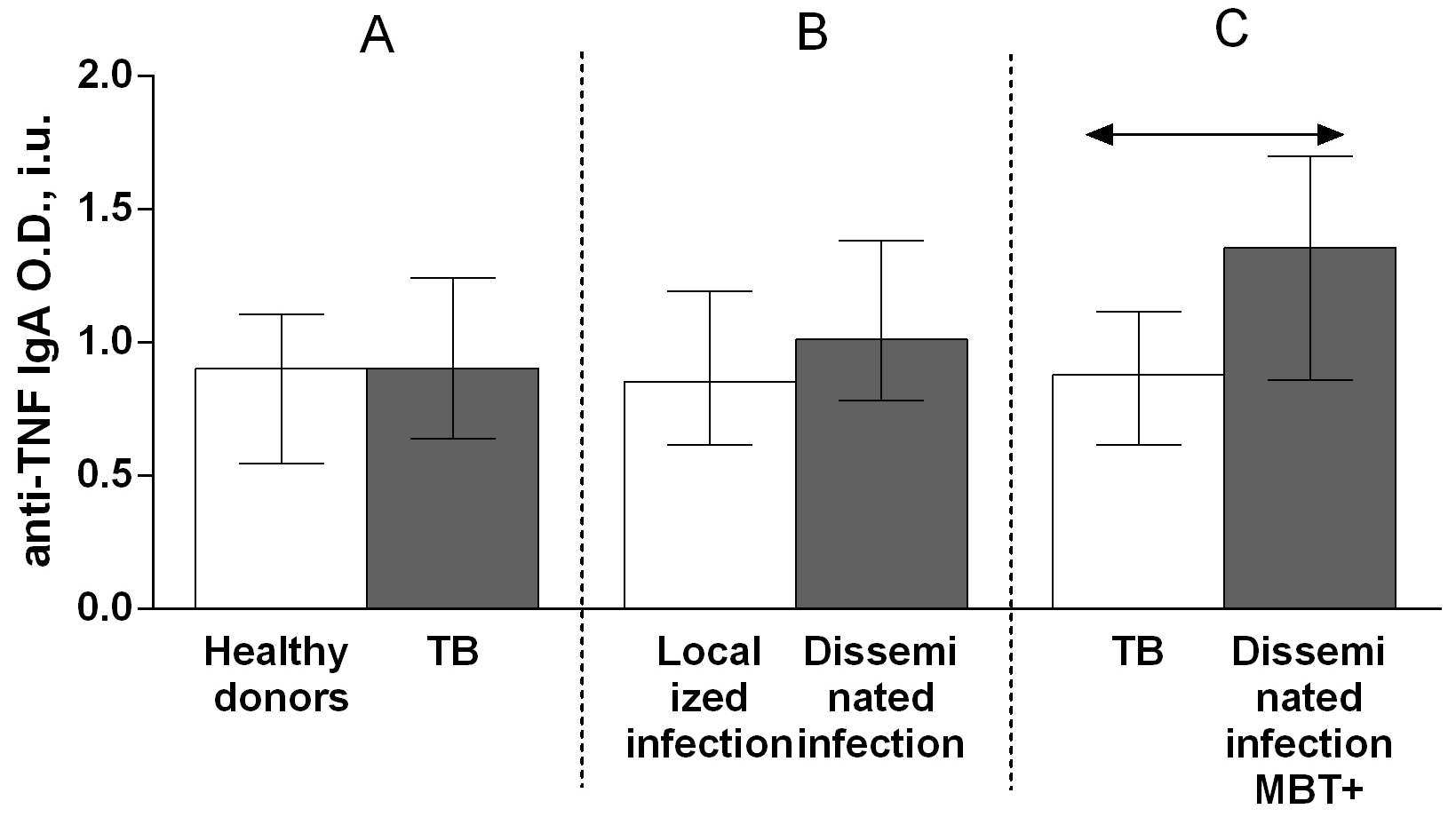 Fig. 5.
Fig. 5.Serum levels of anti-TNF IgA autoantibodies in patients with TB
and healthy donors (A), patients with TB who had localized or disseminated
infection (B) and in MBT+ on bronchoscopy patients with disseminated tuberculosis
and other patients with TB (C). Levels anti-TNF IgA autoantibodies are expressed
in international units of optical density (O.D., i.u.). Double-arrow bar indicate
statistical significance. Data are presented as the median (interquartile range).
Mann–Whitney test, p
Regarding anti-TNF autoantibody subclass, the IgG3 autoantibody titer was higher in patients with TB compared with that in healthy donors (Fig. 6A), whereas there were no significant differences in the titers of IgG1 (Fig. 7A), IgG2 (Fig. 8A), and IgG4 (Fig. 9A) autoantibodies between patients with TB and healthy individuals.
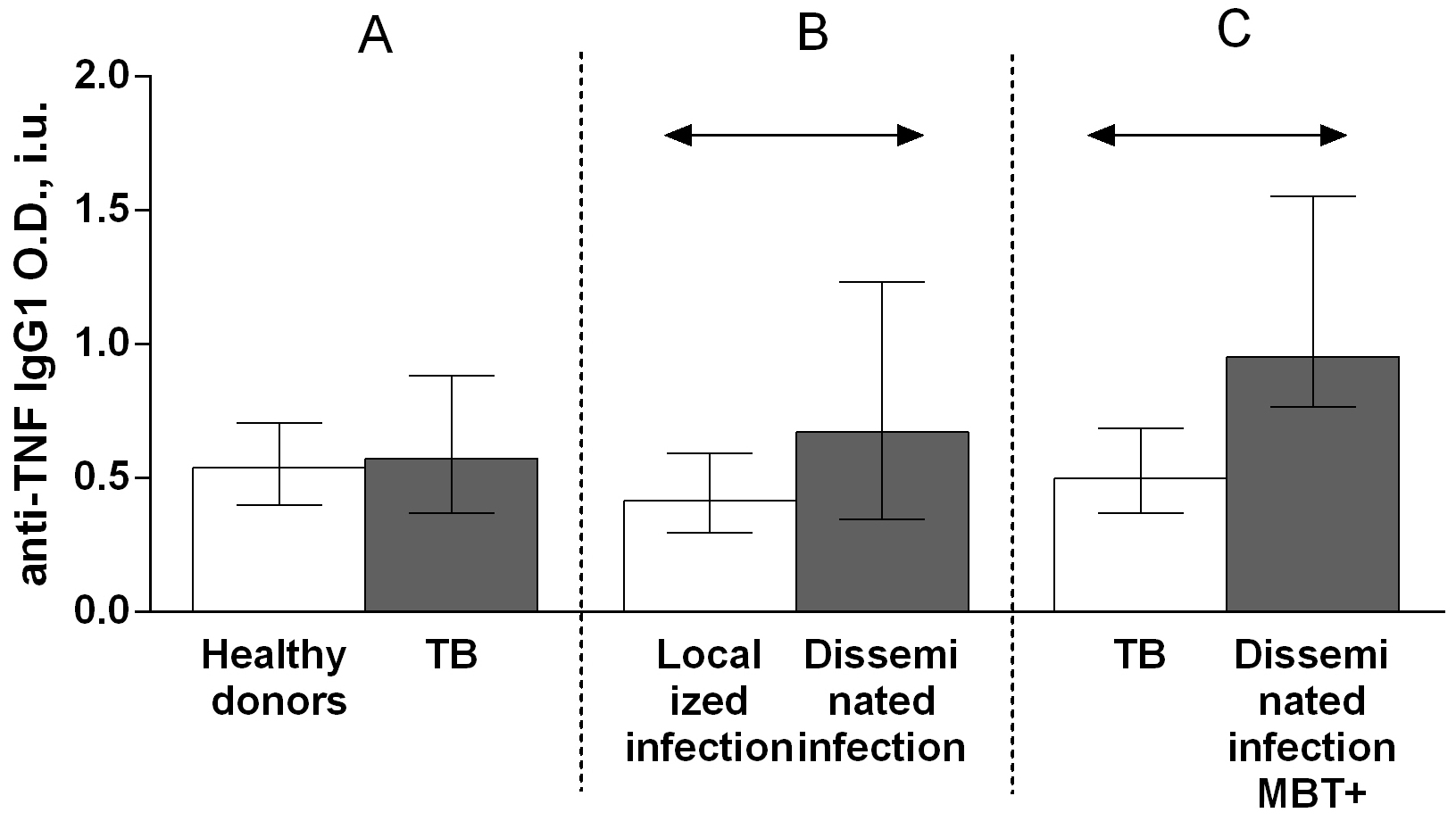 Fig. 6.
Fig. 6.Serum levels of anti-TNF IgG3 autoantibodies in patients with TB
and healthy donors (A), patients with TB who had localized or disseminated
infection (B) and in MBT+ on bronchoscopy patients with disseminated tuberculosis
and other patients with TB (C). Levels anti-TNF IgG3 autoantibodies are expressed
in international units of optical density (O.D., i.u.). Double-arrow bar indicate
statistical significance. Data are presented as the median (interquartile range).
Mann–Whitney test, p
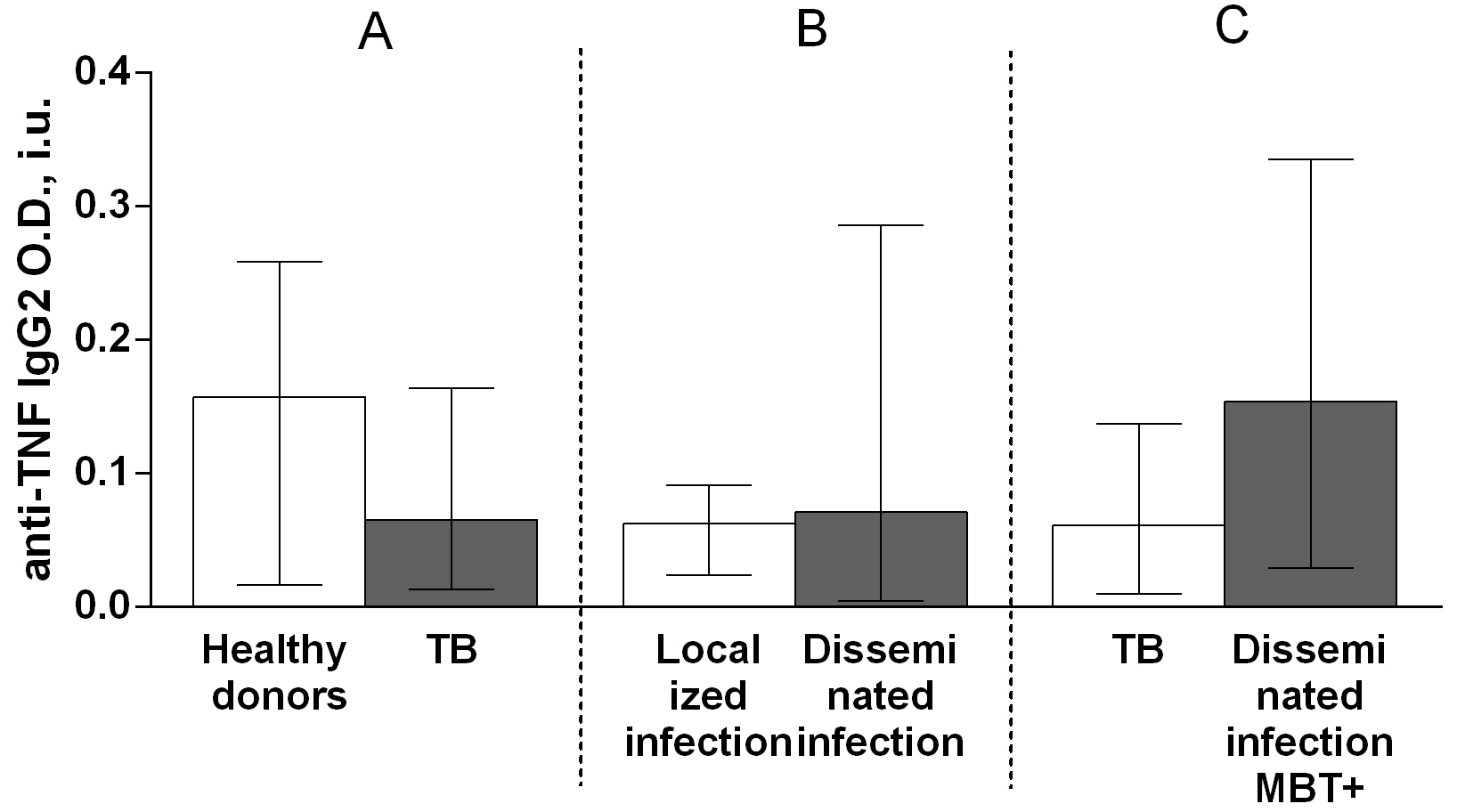 Fig. 7.
Fig. 7.Serum levels of anti-TNF IgG1 autoantibodies in patients with TB
and healthy donors (A), patients with TB who had localized or disseminated
infection (B) and in MBT+ on bronchoscopy patients with disseminated tuberculosis
and other patients with TB (C). Levels anti-TNF IgG1 autoantibodies are expressed
in international units of optical density (O.D., i.u.). Double-arrow bar indicate
statistical significance. Data are presented as the median (interquartile range).
Mann–Whitney test, p
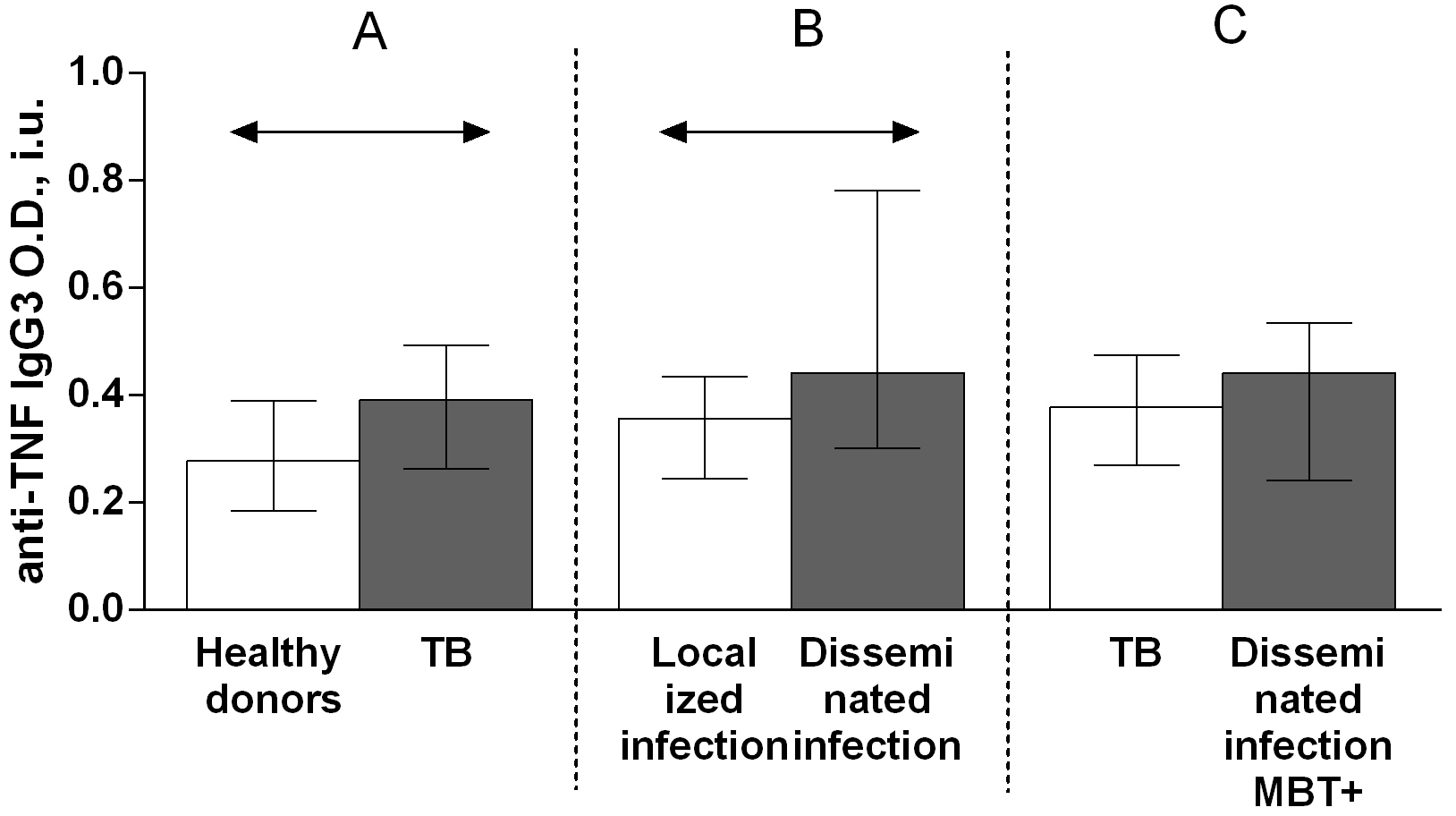 Fig. 8.
Fig. 8.Serum levels of anti-TNF IgG2 autoantibodies in patients with TB
and healthy donors (A), patients with TB who had localized or disseminated
infection (B) and in MBT+ on bronchoscopy patients with disseminated tuberculosis
and other patients with TB (C). Levels anti-TNF IgG2 autoantibodies are expressed
in international units of optical density (O.D., i.u.). Double-arrow bar indicate
statistical significance. Data are presented as the median (interquartile range).
Mann–Whitney test, p
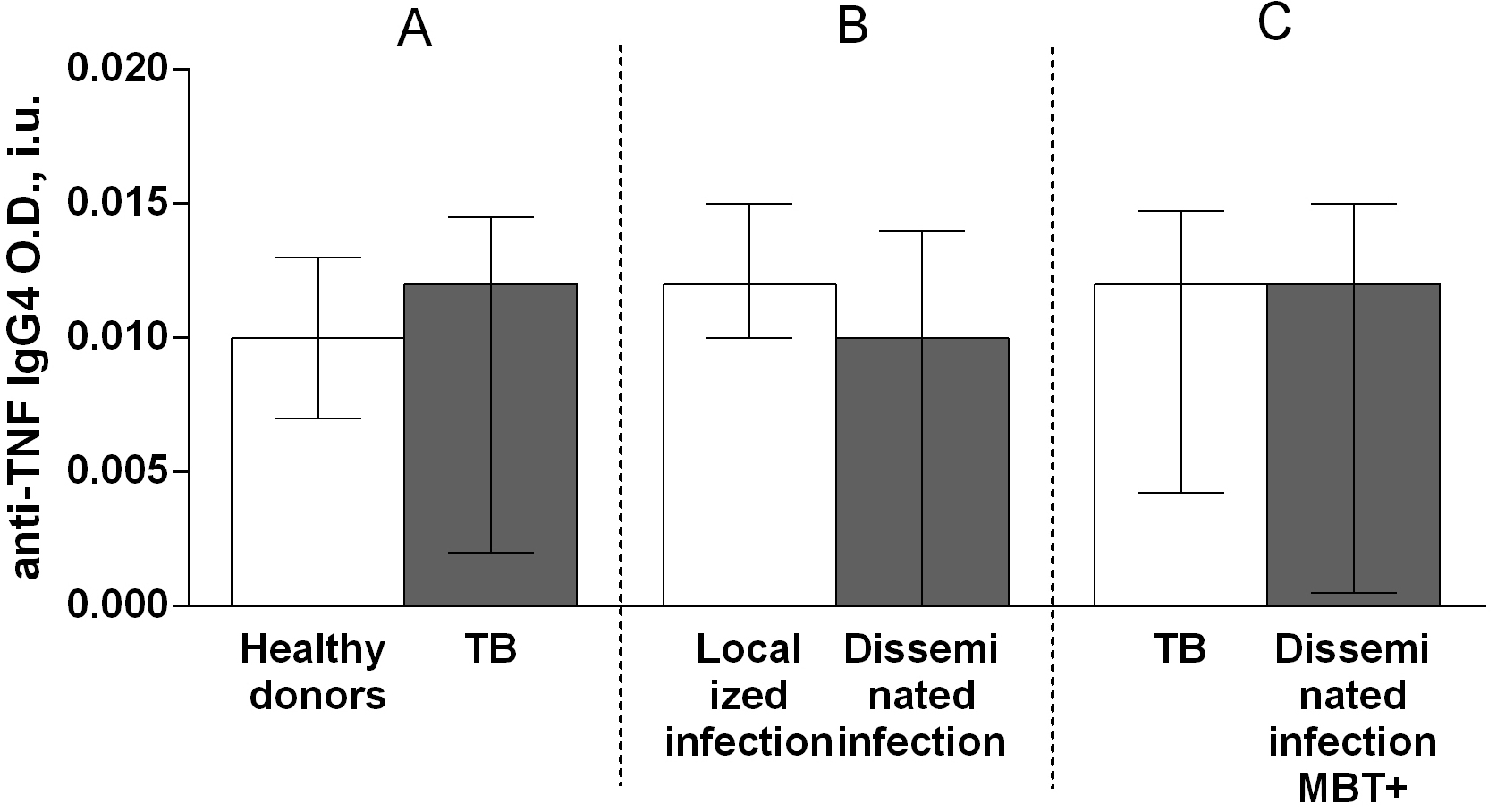 Fig. 9.
Fig. 9.Serum levels of anti-TNF IgG4 autoantibodies in patients with TB
and healthy donors (A), patients with TB who had localized or disseminated
infection (B) and in MBT+ on bronchoscopy patients with disseminated tuberculosis
and other patients with TB (C). Levels anti-TNF IgG4 autoantibodies are expressed
in international units of optical density (O.D., i.u.). Double-arrow bar indicate
statistical significance. Data are presented as the median (interquartile range).
Mann–Whitney test, p
A comparison of the serum titers of anti-TNF autoantibody classes and subclasses among patients with TB revealed higher IgG (Fig. 4B), IgG1 (Fig. 6B) and IgG3 (Fig. 8B) autoantibody titers in patients with disseminated infection than in patients who had a localized infection, and higher IgG (Fig. 4C), IgA (Fig. 5C) and IgG1 (Fig. 6C) in patients with disseminated infection from whom M. tuberculosis was isolated on bronchoscopy than in other patients.
In patients with TB, positive correlations were observed between the titers of anti-TNF autoantibodies of classes IgG and IgA and between those of subclasses IgG1 and IgG3, as well as between the titers of these class and subclass groups. Among healthy individuals, positive correlations were noted between the titers of anti-TNF autoantibodies of classes IgG and IgA, those of classes IgG or IgA and the subclass IgG3, and between the groups of subclasses IgG1 and IgG4, and IgG2 and IgG4.
TNF plays a key role in host defense against M. tuberculosis. However, it can also enhance the pathogenic effect of this disease and is associated with a number of negative manifestations of TB infection. In the literature, there are conflicting reports regarding differences in serum TNF levels between patients with TB and healthy donors; some studies have found an elevated serum TNF level in patients with TB [4], whereas others have observed no significant difference in the serum TNF titer between patients with TB and healthy donors [5]. Notably, the TNF level can decrease during anti-tuberculosis therapy [4]. One previous study evaluated the TNF production of peripheral blood mononuclear cells from patients with TB or healthy donors; mononuclear cells from patients with infiltrative and fibrous cavernous TB were shown to produce less TNF than those from healthy donors. Furthermore, although the levels of purified protein derivate-stimulated TNF production by these cells from patients with infiltrative TB did not differ from that of cells from healthy individuals, these levels were higher in cells from patients with fibrous cavernous TB than in those from healthy donors [22]. There is no generally accepted classification of the severity of pulmonary tuberculosis. In Russia, a classification is used according to the volume of lung damage: up to two segments inclusive — a limited process, more than two segments — a disseminated process [21]. A group of patients with disseminated infection from whom pathogen was isolated on bronchoscopy is the group with most severe disease because it is characterized by disseminated destructive process with cavitations [21]. In the present study, we did not observe significant differences in the TNF level between patients with TB and healthy individuals (Fig. 1A). However, we have found elevated level of tumor necrosis factor in patients characterized by disseminated infection compared with patients with localized infection (Fig. 1B) and in the group of patients with most severe disease with disseminated infection from whom M. tuberculosis was isolated on bronchoscopy than in other patients (Fig. 1C). These differences reflect the lesion volume of the lung tissue and the severity of infectious process during pulmonary tuberculosis, in which TNF plays key roles in the immune and pathological responses and mediator production is stimulated by multiple mechanisms [1, 2].
Soluble receptors can compete with membrane-bound receptors for TNF binding and consequently inhibit TNF bioactivity [6]; however, the functional interactions between TNF and its soluble receptors are complex. The addition of sTNFR1 and sTNFR2 to trimeric TNF leads to the formation of complexes between them, stabilizing the active TNF form [6]. TNF can dissociate from this complex and replace decayed TNF at the periphery, thus maintaining a constant concentration of unbound bioactive trimeric cytokine. This process seems to occur within TB lesions, as evidenced by the increased levels of TNF and of both types of its soluble receptors in bronchoalveolar fluid from patients with TB [23]. Factors sTNFR1 and sTNFR2 can act as TNF antagonists, TNF-carrier proteins, slow-release reservoirs for TNF, stabilizers of TNF bioactivity capable of increasing the cytokine half-life, and a TNF buffer by inhibiting the effect of high TNF concentrations and providing the effect of low and well-controlled cytokine levels on cells [6]. Thus, soluble TNF receptors affect local and systemic TNF bioavailability [6].
One previous study reported that patients with TB who have an elevated serum TNF level exhibit high levels of its soluble type 1 and 2 receptors [23]. However, we found that serum levels of soluble type 1 and 2 TNF receptors in patients with TB were lower compared with those in healthy donors (Fig. 2A). This difference is probably due to the lack of an increased systemic level of the cytokine itself in the serum of patients with TB. Similar to tumor necrosis factor level, sTNFR1 and sTNFR2 levels are elevated in the group of patients with disseminated infection from whom M. tuberculosis was isolated on bronchoscopy than compared other patients (Fig. 2D). This is, obviously, due to the regulatory mechanism, by which high levels of soluble TNF receptors are produced to neutralize cytokine excess and maintain a constant concentration of unbound bioactive trimeric cytokine and to reduce tissue damage.
The study of autoantibodies as molecules reacting with the body’s own antigens
is an important area of research in modern molecular immunology [24]. ACAAB
induction is one of the mechanisms of regulating cytokines in health and
pathology. The reason why autoantibodies lead or do not lead to the development
of pathology is not completely clear. The main point of view is that if the
antibody has sufficient affinity and its content is above a certain threshold
concentration, the autoantibody can neutralize the biological activity of the
cytokine by binding to it and preventing its interaction with its receptor on the
cell. However, a number of studies have shown the ability of different antibodies
to bind to the same molecule of toxins [25, 26] or cytokines [27, 28], which
suggests that in some cases neutralization depends on the production of
antibodies to several antigenic sites, which leads to the formation of immune
complexes that can be effectively eliminated in vivo [27, 28]. Moreover,
monoclonal antibodies to interleukine (IL)-2, which neutralize the effects of a
cytokine in vitro, can enhance and prolong the biological activity of
IL-2 when administered in vivo [19]. Furthermore, there are data on
different biological activity of IgG isolated from the blood serum of patients
with various autoimmune diseases and healthy donors. It has been shown that IgG
isolated from the blood serum of patients with different autoimmune diseases and
healthy individuals has a different effect on the biological activity of
granulocyte colony-stimulating factor, in addition, in patients with one
nosology, the effect of IgG obtained from individual patients may also differ
[13]. In another work, incubation of mononuclear cells from healthy donors,
thyroid peroxidase, and blood sera of patients with Hashimoto’s thyroiditis and
healthy donors was carried out. Wherein, patients had different levels of
anti-thyroid peroxidase autoantibodies. It has been shown that when incubated
with serum containing high levels of autoantibodies to peroxidase an increased
production of TNF, IL-6 and IFN-
The presence of high-affinity IgG autoantibodies against IFN-
Herein, to determine the possible contribution of anti-TNF autoantibodies to the pathogenesis of TB, serum titers of anti-TNF autoantibodies belonging to classes IgA, IgG, or IgM and subclasses IgG1, IgG2, IgG3, or IgG4, as well as the levels of TNF and of its soluble receptors, were assessed in both patients with TB and healthy donors. These autoantibodies were detected in serum from both patients with TB and healthy individuals. The IgG autoantibody titer was higher in patients with TB than in healthy donors (Fig. 4A), but no differences were observed in the titers of IgM (Fig. 3A) and IgA (Fig. 5A) autoantibodies between patients with TB and healthy individuals. Furthermore, the level of IgG3 autoantibodies was higher in patients with TB compared with healthy donors (Fig. 8A), whereas the titers of IgG1 (Fig. 6A), IgG2 (Fig. 7A), and IgG4 (Fig. 9A) autoantibodies in patients with TB did not differ from those in healthy individuals.
A comparison of the anti-TNF autoantibody titers among patients with TB revealed higher IgG (Fig. 4B), IgG1 (Fig. 6B) and IgG3 (Fig. 8B) titers in patients who had a disseminated infection compared with individuals who had a localized infection, as well as IgG (Fig. 4C), IgA (Fig. 5C) and IgG1 (Fig. 6C) in MBT+ on bronchoscopy patients with disseminated infection than in other patients with TB. These data point on a possible significant role for autoantibodies belonging to the IgG class, particularly those belonging to the IgG1 or IgG3 subclasses, in immune responses to TB. Various research groups have previously shown that specific autoantibodies in autoimmune disease patients are often composed mainly of antibodies belonging to the IgG1 or IgG3 subclass [37].
The observed higher levels of IgG (Fig. 4A) and IgG3 (Fig. 8A) anti-TNF autoantibodies in patients with TB compared with healthy donors, of IgG (Fig. 4B), IgG1 (Fig. 6B) and IgG3 (Fig. 8B) anti-TNF autoantibodies in patients with a disseminated infection compared with patients with a localized manifestation of TB and IgG (Fig. 4C) and IgG1 (Fig. 6C) anti-TNF autoantibodies in MBT+ on bronchoscopy patients with disseminated infection suggest an active involvement of anti-TNF autoantibodies in complement system activation. IgG1 and IgG3 immunoglobulins have the strongest ability to interact with the C1q complement component, while IgG2 is capable of weak binding, and IgG4 is unable to bind C1q [37].
Elevated IgG class (Fig. 4A) and IgG3 (Fig. 8A) subclass of anti-TNF autoantibody titers in patients with TB, as well as higher IgG (Fig. 4B), IgG1 (Fig. 6B) and IgG3 (Fig. 8B) anti-TNF autoantibody titers in individuals with a disseminated infection, and IgG (Fig. 4C) and IgG1 (Fig. 6C) anti-TNF autoantibody titers in the group of patients with most severe disease with disseminated infection from whom pathogen was isolated on bronchoscopy may indicate the damaging effect of these antibodies in pathology [20]. This effect might be caused by enhanced TNF action via the formation of a complex with TNF that increases the expression of pro-inflammatory mediators through an Fc-receptor-dependent mechanism, as is the case with anti-IL-8 autoantibodies during acute respiratory distress syndrome [20]. Moreover, some anti-cytokine antibodies can enhance the bioactivity and increase the half-life of cytokines in vivo, acting as a reservoir of cytokines (carrier proteins) [38, 39]. It has been shown in a mouse model that the administration of a cytokine with the corresponding monoclonal antibody in a low molar ratio (cytokine:mAb = 2:1) leads to an increase in the biological activity of the mediator and an increase in the half-life [38]. In a number of studies, a high level of antibodies enhances the function of the corresponding cytokines. For example, mAb to IL-3 or IL-4 increase the stimulation of proliferation of mouse mucosal mast cells [39]. However, to clarify the likely damaging role of anti-TNF autoantibodies in the pathogenesis of pulmonary tuberculosis, it is necessary to conduct tests to determine the functional activity of such antibodies and their influence on the biological effects of tumor necrosis factor.
In conclusion, our findings on the anti-TNF autoantibody titers in patients with TB and healthy donors suggest that these autoantibodies may play an important role in the immunopathogenesis of pulmonary tuberculosis.
ACAAB, anti-cytokine autoantibody; ELISA, enzyme-linked immunosorbent assay; Ig, immunoglobulin; IFN, interferon; IL, interleukine; TB, active pulmonary tuberculosis; TNF, tumor necrosis factor.
FDK, JAL, ZAL and SVS contributed to the conception, design, analysis and data interpretation, and drafting of the manuscript; FDK and JAL contributed to experimental work; ZAL performed patient recruiting; SVS contributed to revision and final approval of the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript.
The study protocol was approved by the local ethics committee at the Research Institute of Fundamental and Clinical Immunology.
We thank the staff of the Laboratory of Molecular Immunology, Federal State Budgetary Scientific Institution “Research Institute of Fundamental and Clinical Immunology”, Novosibirsk, Russia, for their assistance. We are also grateful to the staff of the First Therapy Department of Federal State Budgetary Institution “Novosibirsk Research Institute of Tuberculosis”, Ministry of Health of Russian Federation, Novosibirsk, Russia, for their help. We thank Katie Oakley, from Edanz Group (https://en-author-services.edanz.com/ac) for editing a draft of this manuscript.
This work was supported by the Ministry of Science and Higher Education of the Russian Federation.
The authors declare no conflict of interest. SVS is serving as one of the Editorial Board members of this journal. We declare that SVS had no involvement in the peer review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to HY.