






1 Department of Animal Science, Texas A&M University, College Station, TX 77843, USA
2 Department of Veterinary Integrative Biosciences, College Station, TX 77843, USA
§The current address of Xilong Li is the Institute of Feed Research, The Chinese Academy of Agricultural Sciences, 100081 Beijing, China.
Academic Editors: Graham Pawelec and Brenda M. Alexander
Abstract
Background: Fetal-placental development depends on a continuous and efficient supply of nutrients from maternal blood that are acquired by exchange through the placenta. However, the placenta is a low permeability barrier, and effective transport of substances depends on specific transport mechanisms. Active transport requires that ions or nutrients be moved against an electrical and/or concentration gradient. In pigs, active transport of ions occurs across the chorioallantois placenta to produce an electrochemical gradient that changes throughout gestation. The aim of this study was to utilize Ussing chambers to detect regulation of ion transport across the porcine chorioallantois by a factor(s) within the uterine-placental environment of pigs. Methods: For the measurement of transchorioallantoic voltage potential as an index of ion transport across the placenta, pieces of chorioallantoic tissue from Day 60 of gestation were mounted into the cassettes of Ussing chambers, and treatments were added to the mucasal side of the tissue. Treatments included: (1) media incubated with Day 60 chorioallantois (placenta-conditioned media); (2) osteopontin/secreted phosphoprotein 1 (OPN/SPP1) purified from cow’s milk; (3) placenta-conditioned media from which OPN/SPP1 was removed; and (4) recombinant rat OPN with an intact RGD integrin binding sequence or a mutated RAD sequence. Ouabain was added to both sides of the chamber. Immunofluorescence was utilized to localize beta 3 integrin, aquaporin 8 and OPN/SPP1 in porcine placental tissues, and OPN/SPP1 within porcine lung, kidney and small intestine. Results: Day 60 chorioallantoic membranes had greater transepithelial voltage in the presence of porcine placenta-conditioned media, indicating that a molecule(s) released from the placenta increased ion transport across the placenta. OPN/SPP1 purified from cow’s milk increased ion transport across the placenta. When OPN/SPP1 was removed from placenta-conditioned media, ion transport across the placenta did not increase. Recombinant rat OPN/SPP1 with a mutated RGD sequence that does not bind integrins (RAD) did not increase ion transport across the placenta. Ouabain, an inhibiter of the sodium-potassium ion pump, ablated ion transport across the placenta. Conclusions: The present study documents a novel pericellular matrix role for OPN/SPP1 to bind integrins and increase ion transport across the porcine chorioallantoic placenta.
Keywords
- pig
- placenta
- ion transport
- ussing chambers
- osteopontin
- secreted phosphoprotein 1
Fetal growth and development depends on a continuous and efficient supply of nutrients from maternal blood. Most nutrients required by the fetus are acquired by exchange through the placenta [1, 2]. However, the placenta is a low permeability barrier, and the effective transport of nutrients and substrates for fetal growth depends less on intrinsic diffusion, and more on a host of specific transport mechanisms [1]. As such, the placenta resembles the intestinal mucosa or renal epithelium [2]. Active transport requires that ions or nutrients be moved against an electrical and/or concentration gradient.
In pigs, active transport of ions occurs across the chorioallantois to produce
an electrochemical gradient (potential difference) that changes throughout
gestation [3, 4]. Isolated chorioallantoic membranes mounted in Ussing chambers
produce an electrical potential at 20 days of gestation that increases 4-fold by
days 45–67 [5], and is maintained through the 107th day of gestation [4]. This
electrical potential across the porcine chorioallantois is likely maintained by a
Na
Nutrients that pass from the uterus through the chorion can directly access allantoic vasculature for transport to the fetus, but nutrients can also diffuse across the allantoic mesenchyme for active transport across the allantoic epithelium into the allantoic sac. Allantoic fluid is rich in electrolytes, sugars, amino acids, and proteins, and serves as a nutrient reservoir. Because the allantoic epithelium is derived from the hindgut and is, therefore, absorptive in nature, these stored nutrients can be transported back across this epithelium to the allantoic vasculature for transport to the fetus. The mechanistic trigger(s) for these active transport events across the maternal and fetal sides of the chorioallantois is unknown. The regulation of transport may be affected by intrinsic mechanisms, in which intracellular proteins interact with transporters, or by extracellular influences such as effectors and their receptors in the chorionic and allantoic membranes [2].
Active transport requires that ions or nutrients be moved against an electrical and/or concentration gradient. Ussing chambers are a physiological tool to distinguish between active transport of ions, nutrients and drugs across epithelia and passive movement of ions through paracellular pathways [9]. This method utilizes short-term tissue culture that enables precise measurement of electrical and transport parameters of intact, polarized epithelia in a physiological context [9, 10]. The aim of this study was to utilize these chambers to detect regulation of ion transport across the porcine chorioallantois placenta by a factor(s) within the uterine-placental environment of pigs. Osteopontin (OPN)/Secreted Phosphoprotein 1 (SPP1) is a secreted matricellular effector molecule perfectly localized to bind integrin receptors on chorion and allantoic epithelium to increase ion and nutrient transport, e.g., glucose, fructose and amino acids, across the pig placenta [11, 12, 13, 14, 15].
Briefly, seven gilts (F1 crosses of
Yorkshire
For the measurement of transchorioallantoic voltage potential as an index of ion
flux across the placenta [9], pieces of chorioallantoic placental tissue (1
cm
Immunoprecipitation of OPN/SPP1 from placenta-conditioned media was performed by
combining placenta-conditioned media with Protein A-Sepharose (Pierce, Rockford,
IL, USA), 0.5% Triton X-100 in TBS, and a pool of two rabbit antibodies directed
against human OPN/SPP1 (LF-123 and LF-124) [20] and a rabbit anti-bovine OPN/SPP1
IgG (generously provided by Dr. George J. Killian) [21], or irrelevant rabbit
IgG. This mixture was rotated gently overnight at 4
For immunofluorescence staining, primary antibodies included rabbit anti-beta 3 integrin (ITGB3) IgG (AB1932; EMD Millipore, Burlington, MA, USA; 1/1000 dilution for OCT-embedded tissue sections), mouse anti-AQP8 IgG (Sigma Aldrich, St. Louis, MO, USA; 1/200 dilution for paraffin-embedded tissue sections), rabbit anti-OPN/SPP1 IgG (AB10910; EMD Millipore; 1/1000 dilution for OCT-embedded tissue sections, and 1/200 dilution for paraffin-embedded tissue sections, boiling citrate antigen retrieval for paraffin-embedded tissue sections), and mouse anti-E-cadherin monoclonal IgG (BD Biosciences; San Jose, CA, USA; 610182; 1/200 dilution for paraffin-embedded tissue sections, boiling citrate antigen retrieval). The secondary antibodies included goat anti-rabbit-Alexa Fluor 488-conjugated IgG, goat anti-rabbit Alexa Fluor 594-conjugated IgG, and goat anti-mouse-Alexa Fluor 594-conjugated IgG (Life Technologies, Grand Island, NY, USA; 1/250 dilution). In addition an FITC-conjugated lectin from Dolichos biflorus (Sigma-Aldrich; L9142-1MG; 1/200 dilution) was used to co-localize with OPN/SPP1 immunofluorescence.
Immunostaining of frozen tissue sections was performed to localize ITGB3 to
placental areolae and OPN/SPP1 to allantoic epithelium as previously described
[23]. Briefly, frozen sections (~10
Immunostaining of paraffin-embedded tissue sections was performed to localize
aquaporin 8 (AQP8) to placental areolae as previously described [25]. Briefly,
sections (5
Dual immunofluorescence staining of OPN/SPP1 and E-cadherin proteins in paraffin-embedded tissue sections followed the same procedures as described for normal immunofluorescence staining except that the two primary antibodies were added simultaneously on the first day and the two secondary antibodies (goat anti-rabbit-Alexa Fluor 488-conjugated IgG and goat anti-mouse-Alexa Fluor 594-conjugated IgG) were added simultaneously on the second day [25]. Images were taken using an Axioplan 2 microscope (Carl Zeiss, Thornwood, NY, USA) interfaced with an Axioplan HR digital camera.
All statistical analyses were performed using GraphPad Prism (GraphPad Software,
La Jolla, CA, USA). Data for the effect of conditioned media and OPN/SPP1 on ion
transport were subjected to one-way ANOVA followed by a post-hoc Tuckey analysis.
For comparison of two treatment groups over time, a two-way ANOVA with Bonferroni
multiple comparison test was used. All data are presented as mean
Day 60 chorioallantoic membranes were placed into Ussing chamber cassettes and
0.2 mL of placenta-conditioned media added to the mucosal side of the tissue. A
statistically significant increase in the transepithelial voltage, as an index of
ion transport across the placenta, was observed between 0 and 5 min after the
addition of 0.2 mL placental solution to the “mucosal side” of the using
chamber, and this increase was maintained through the remaining 15 min (Fig. 1).
When 100
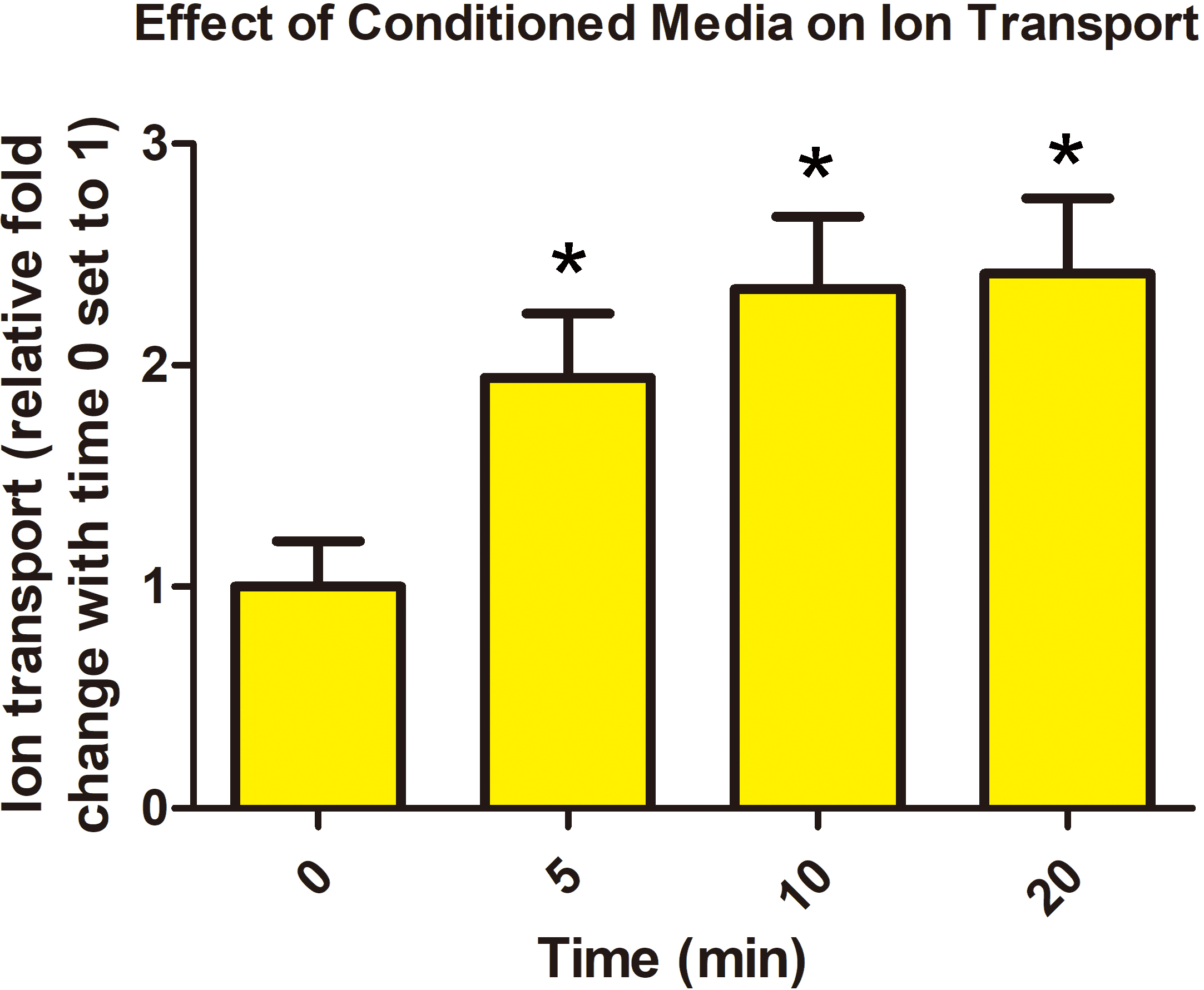 Fig. 1.
Fig. 1.Porcine chorioallantois releases a factor(s) that increases ion
transport across the porcine placenta. Addition of 0.2 mL of
placenta-conditioned media to the mucosal side of the Ussing chamber increased
transepithelial voltage across Day 60 chorioallantois by 10 min of
incubation. Values are
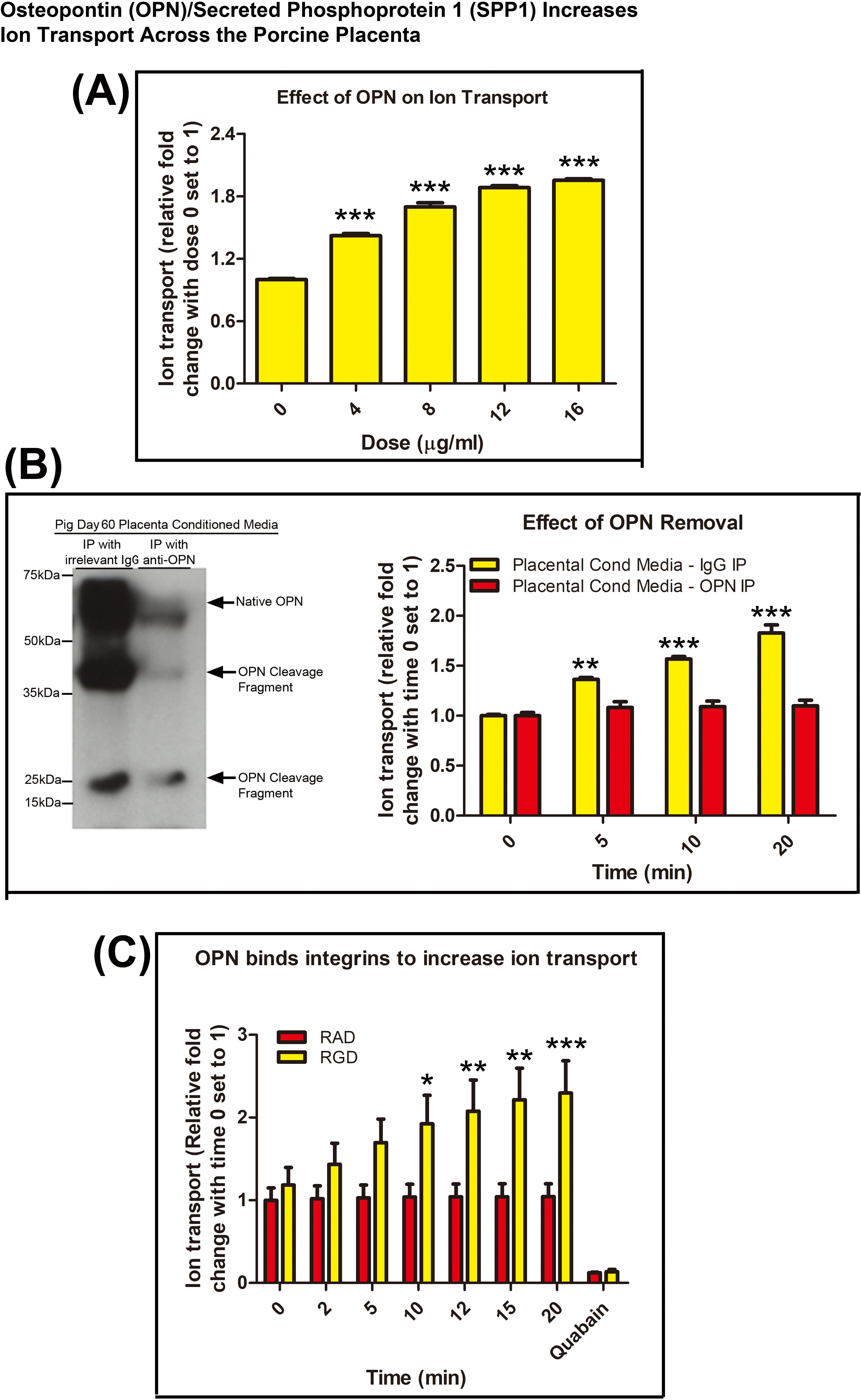 Fig. 2.
Fig. 2.Osteopontin (OPN)/Secreted Phosphoprotein 1 (SPP1) increases ion
transport across the porcine placenta. (A) Effect of addition of OPN/SPP1
purified from bovine milk to the mucosal side of a Ussing chamber. OPN/SPP1 from
bovine milk increased transepithelial voltage across Day 60 chorioallantois in a
dose dependent manner. (B) Western blot demonstrating removal of OPN/SPP1 from
placenta-conditioned media via immunoprecipitation. Rabbit IgG controls for these
antibodies in porcine tissue have previously been published [11]. Addition of
placenta-conditioned media with OPN/SPP1 removed (immunoprecipitation with
anti-OPN) to the mucosal side of a Ussing chamber did not increase
transepithelial voltage across Day 60 chorioallantois. (C) Effect of addition of
recombinant rat OPN/SPP1 with an intact integrin binding RGD sequence and
recombinant rat OPN/SPP1 with an integrin binding sequence mutated to RAD to the
mucosal side of a Ussing chamber. Addition of OPN/SPP1 containing an RAD sequence
did not increase transepithelial voltage across Day 60 chorioallantois. 100
Day 60 chorioallantoic membranes were placed into Ussing chamber cassettes and
OPN/SPP1 purified from cow’s milk was added to the mucosal side of the tissue. A
statistically signifant increase in the transepithelial voltage was observed in
response to treatment with 4
Chorionic areolae are composed of specialized tall columnar epithelial cells that are not closely apposed to the uterine luminal epithelium (LE), and form a pouch at the openings of the mouths of uterine glands to receive the secretions of uterine glandular epithelium (GE). Areolae transport glandular secretions across the placenta and into the fetal-placental circulation [26]. ITGB3 can noncovalently link to the alpha v integrin subunit (ITGA3) to form an integrin receptor that binds to OPN/SPP1 and has been demonstrated to engage OPN/SPP1 on porcine uterine LE and conceptus trophectoderm cells [13, 14]. Aquaporins (AQPs) are water-selective channels that function as pores for water transport through the plasma membrane [27]. Fig. 3A,B demonstrate expression of ITGB3 and AQP8 protein at the apical surface of the tall columnar cells of porcine areolae on Day 60 of gestation. In addition AQP8 is expressed at the apical surface of uterine GE and allantoic epithelial cells (Fig. 3B).
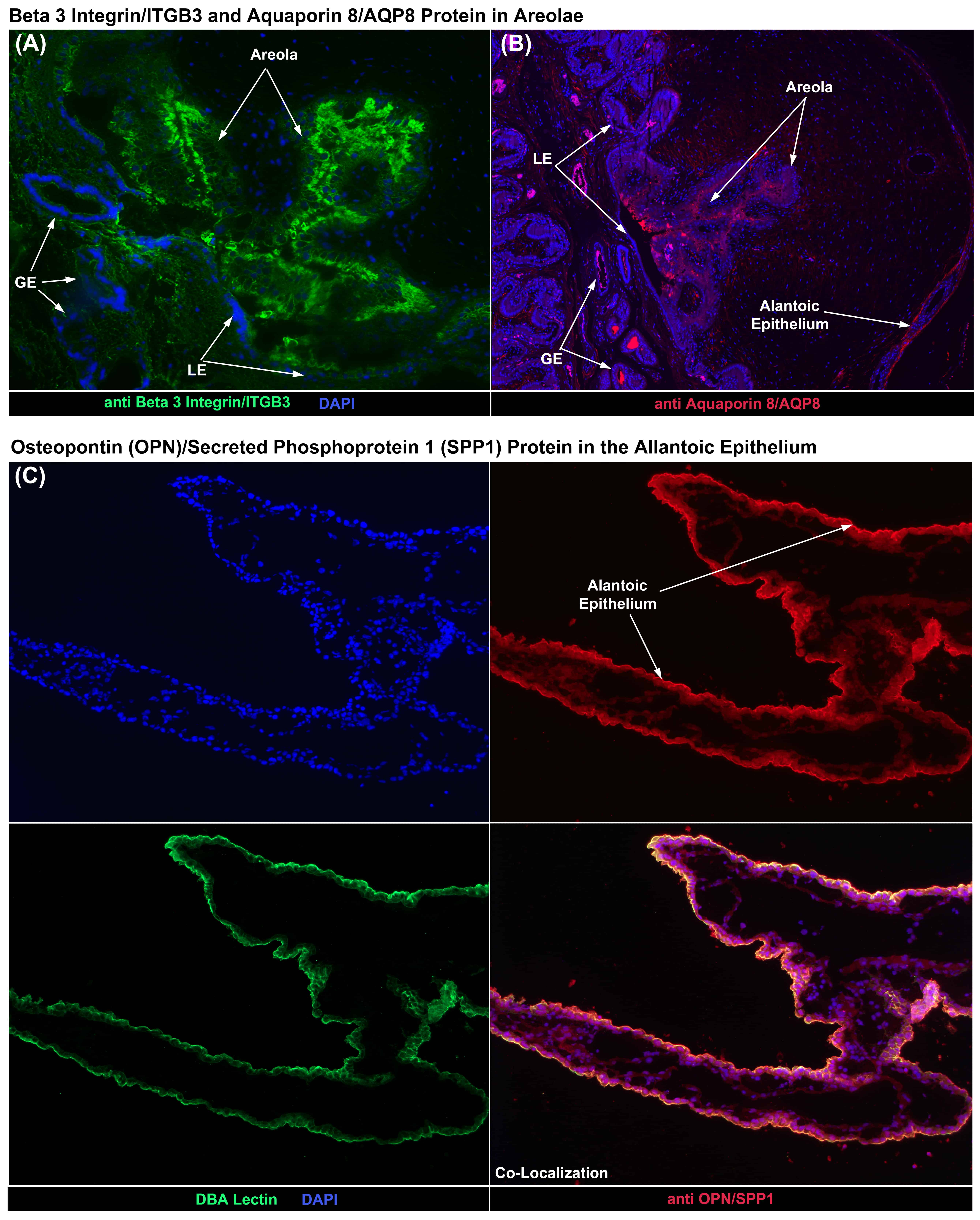 Fig. 3.
Fig. 3.Immunofluorescence (IF) staining for proteins in the porcine
placenta. (A) IF localization of the beta 3 integrin subunit (ITGB3) to the tall
columnar cells of a Day 60 chorionic areola. (B) IF localization of aquaporin 8
(AQP8) to the tall columnar cells of a Day 60 chorionic areola, as well as to the
allantoic epithelium. (C) IF co-localization of OPN/SPP1 and DBA lectin to the
apical surface of allantoic epithelial cells on Day 60 of gestation. LE, uterine
luminal epithelium; GE, uterine glandular epithelium. Rabbit and mouse IgG
controls are shown in Fig. 4. Width of fields is 895
The allantoic epithelium lines the allantoic cavity and is a tissue conduit for transport of ions, water and nutrients from the allantois of the placenta to the allantoic cavity for storage in allantoic fluid [26]. Fig. 3C demonstrates the localization of OPN/SPP1 protein to the allantoic epithelium, and this expression is at the apical surface of the allantoic epithelium because OPN/SPP1 expression co-localizes with DBA lectin, which is only expressed at the apical surface of epithelial cells (Fig. 3C).
Results strongly suggest that OPN/SPP1 mediates ion transport across the porcine choriollantois. The porcine placenta, indeed all placentae, are highly adapted for transport of ions, water, nutrients, etc. across the tissue, and OPN/SPP1 is expressed by porcine uterine LE and GE [11, 12]. Therefore we examined the cell-type specific expression of OPN/SPP1 in other tissues known to be adapted for transport including the lung, kidney and small intestine. Within the lung, OPN/SPP1 protein was localized to a small subset of bronchioles with a punctate pattern of immunostaining (Fig. 4A). In addition OPN/SPP1 protein was detected in the bronchial cartilage, which is to be expected, and suggests effective immunostaining (Fig. 4A). Within the kidney, OPN/SPP1 protein was localized to the collecting ducts of both the cortical medullary rays and the renal medulla (Fig. 4B). Within the small intestine OPN/SPP1 protein was localized to a subpopulation of goblet cells (Fig. 4C).
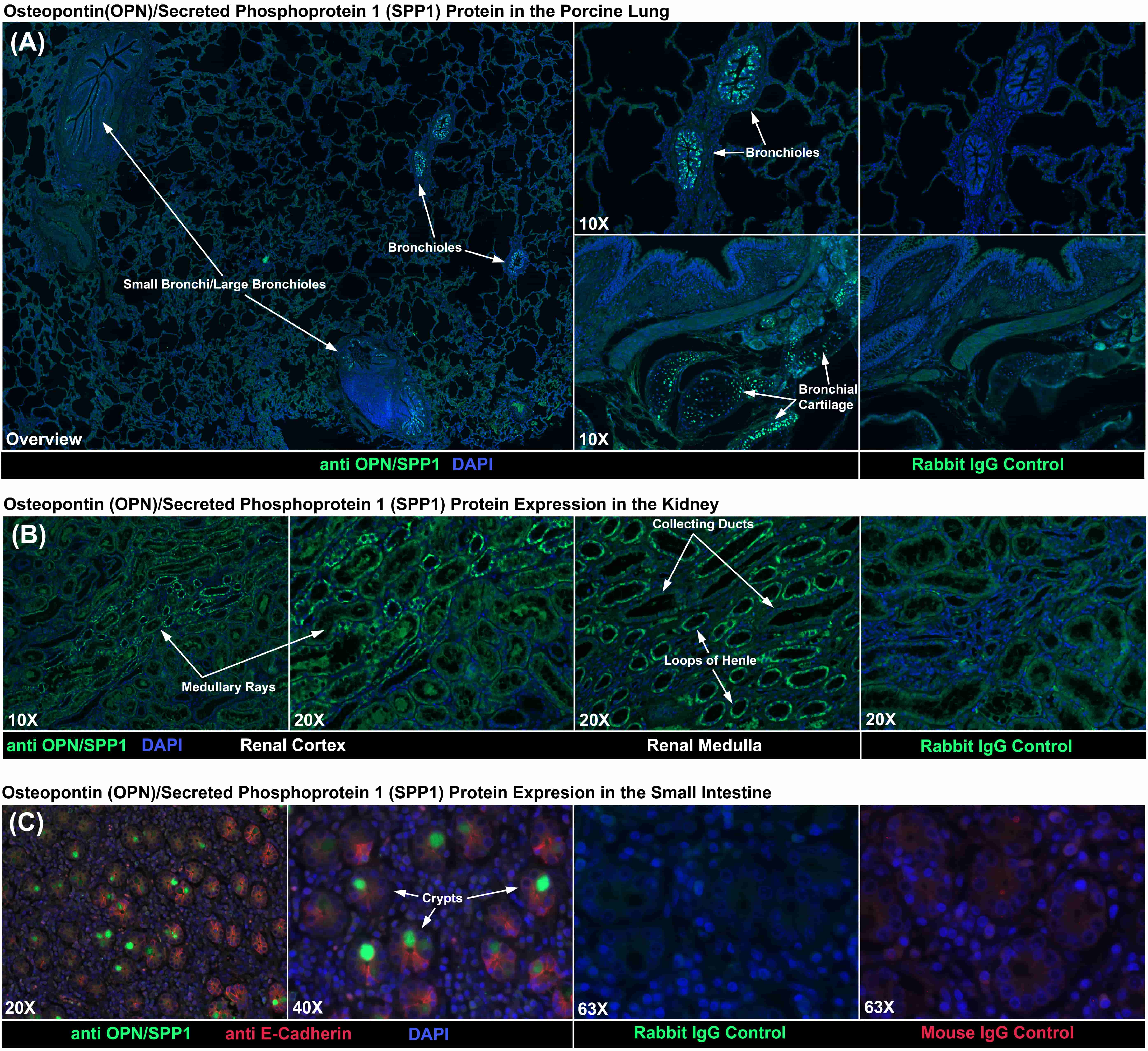 Fig. 4.
Fig. 4.Immunofluorescence (IF) staining for OPN/SPP1 protein. (A) IF
localized OPN/SPP1 protein to some bronchioles and the cartilage of bronchi
within the lung. (B) IF localized OPN/SPP1 protein to collecting ducts within the
kidney. (C) IF localized OPN/SPP1 protein to some goblet cellswithin the small
intestine. Rabbit and mouse IgG negative controls are shown. Width of fields for
overview image is 4120
Prevailing dogma asserts that OPN/SPP1 binds integrins to mediate attachment and migration of placental epithelia during implantation and placentation [28, 29, 30, 31]. However, in the present study we have documented a novel pericellular matrix role for OPN/SPP1 as it binds integrins to increase ion transport across the porcine chorioallantoic placenta. Day 60 chorioallantoic membranes from pigs were placed in Ussing chambers and the transepithelial voltage potential was measured as an index of ion flux across the placenta. Addition of Day 60 placenta-conditioned media doubled the transepithelial voltage, indicating that a molecule(s) released from the pig placenta increases ion transport across the placenta. We tested the ability of OPN/SPP1 to affect ion transport. OPN/SPP1 purified from cow’s milk increased ion transport, and maximum transepithelial voltage plateaued at levels similar to those for placenta-conditioned media. To confirm that OPN/SPP1 is the factor in placenta-conditioned media responsible for increasing ion transport, we removed OPN/SPP1 from placenta-conditioned media by immunoprecipitation using a mixture of antibodies developed against OPN/SPP1. We also performed immunoprecipitation using an irrelevant rabbit IgG. Placenta-conditioned media depletion of OPN/SPP1 did not stimulate ion transport across the placenta. To confirm that OPN/SPP1 binds integrins to mediate ion transport, Day 60 chorioallantoic membranes from pigs were placed into Ussing chambers, recombinant rat OPN with an intact RGD integrin binding sequence was added to the mucosal side of the chamber, and transepithelial voltage and current were measured. Addition of increasing amounts of rat OPN/SPP1 with an intact RGD dose-dependently increased ion transport across the placenta. When the same experiment was performed using recombinant rat OPN/SPP1 with a mutated RGD sequence that does not bind integrins (RAD), no increase in placental ion transport was observed.
OPN/SPP1 is highly expressed within the intrauterine environment of pregnant
pigs [32]. During pregnancy, in pigs, OPN/SPP1 mRNA is induced initially by
estrogens secreted by the conceptus (embryo and associated placental membranes)
in discrete regions of the uterine LE juxtaposed to the conceptus during the
apposition and adhesion phases of implantation. OPN/SPP1 mRNA expands to the
entire uterine LE by Day 20 when firm adhesion of conceptus trophectoderm to
uterine LE occurs [11, 12], and OPN/SPP1 protein remains abundant along the
apical surfaces of uterine LE and trophectoderm in all areas of direct contact
throughout pregnancy. In addition, there are specialized cells of the chorionic
areolae at the openings of the mouths of uterine glands. Indeed, the open space
between the chorionic areolae and uterine GE is filled with secretions of the
uterine GE collectively termed histotroph [33]. The areolae transport secretions
of uterine GE such as macromolecules, particularly proteins, across the placenta.
Total uterine OPN/SPP1 mRNA increases 20-fold between Days 25 and 85 of gestation
in the uterine GE [11], which results in accumulation of OPN/SPP1 in allantoic
fluid (unpublished results). Therefore, large amounts of soluble OPN/SPP1 are
present at both the maternal and fetal surfaces of the chorioallantois. OPN/SPP1
expression in uterine GE during the later stages of pregnancy is similar in sheep
[34], and a microarray study in rats showed that OPN/SPP1 expression increased
60-fold between Day 0 of the estrous cycle and Day 20 of pregnancy [35], likely
within the decidua because OPN is expressed by uterine natural killer cells of
the mouse decidua [24]. In humans, OPN/SPP1 is expressed by the cytotrophoblasts
of chorionic villi, and its
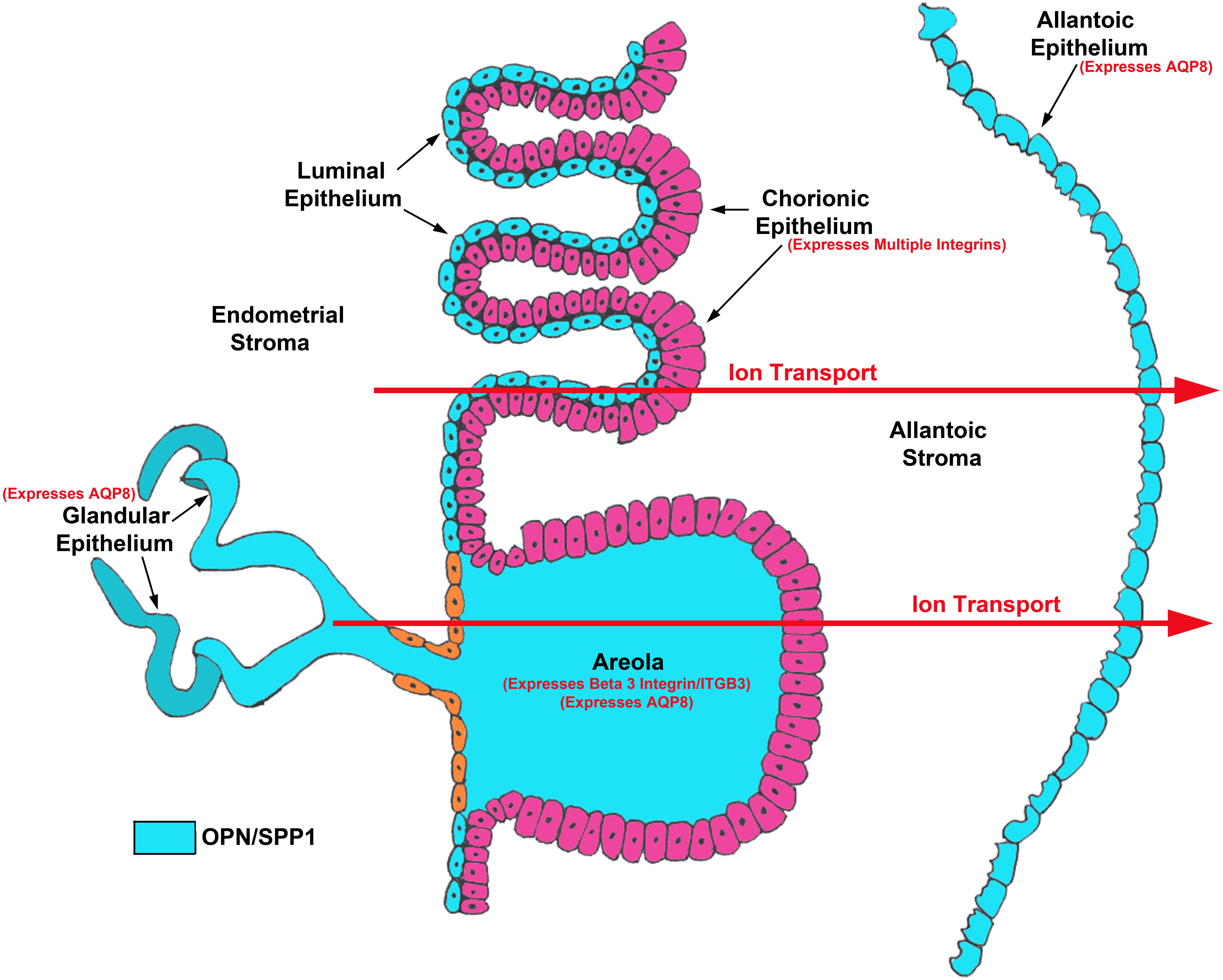 Fig. 5.
Fig. 5.Working model for OPN/SPP1 mediated ion transport across the porcine placenta. In the pig, for an ion to transport from the mucosal to the serosal side of the chorioallantois it must pass through the cytoplasm and basement membrane of the chorionic epithelial cells of the folded inter-areolar regions of placentation and areolae, across the remaining allantoic stroma, and through the basement membrane and cytoplasm of the allantoic epithelial cells [26]. OPN/SPP1 is highly expressed by both the uterine LE and GE of pigs which potentially supply secreted OPN/SPP1 protein to the to areolar and inter-areolar chorionic epithelium [11]. OPN/SPP1 has also been localized within the porcine allantoic stroma [15], and Fig. 3C demonstrates OPN/SPP1 expression in the allantoic epithelium. Multiple integrin subunits that could assemble into integrin receptors that bind OPN/SPP1 have been localized to the apical surface of inter-areolar chorionic epithelium [14], and Fig. 3A demonstrates beta 3 integrin (ITGB3) subunit expression in areolar chorionic epithelial cells. Transporters have been localized to key tissues within the chorion of pigs including SLC2A3 and SLC2A8 [24, 41], and Fig. 3B demonstrates aquaporin 8 (AQP8) expression in the uterine GE, areolar chorionic epithelium and allantoic epithelium.
Integrins are transmembrane proteins composed of
Integrins mediate cell-cell and cell-ECM adhesion to regulate cell motility,
proliferation and differentiation [42, 47]. These events are often accompanied by
changes in ion flow. Integrins form macromolecular complexes that localize ion
channels to the plasma membrane [51, 52]. Integrins regulate K
We identified a here-to-fore unknown role of OPN/SPP1 and integrins at the uterine-placental interface of pregnancy in enhancing placental ion transport. We hypothesize that in pigs OPN/SPP1 is synthesized and secreted from uterine GE and LE, binds to integrins on the chorionic epithelium and activates ion transporters that alter the magnitude of and/or cellular localization of nutrient transporters and/or the activity of those transporters to increase nutrient transport across the chorionic and allantoic membranes to the placental vasculature and allantoic cavity for transfer to the embryo/fetus (Fig. 5). This novel finding of our current study may provide key insight into why OPN/SPP1 is highly expressed at sites of active nutrient transport in a variety of placentae including the uterine-placental interface of species with epitheliochorial and synepitheliochorial placentae, uterine decidua of rodents, and cytotrophoblasts of human chorionic villi.
GW contributed funding to support the studies, performed the experiments to measure ion transport using Ussing chambers, contributed to the interpretation of data, and contributed to manuscript preparation. XL contributed extensively to animal husbandry including feeding, pen care, heat checking and breeding. HS contributed to immunohistochemical studies and manuscript prepartation. BAM contributed immunohistochemical studies. ACK contributed to immunohistochemical studies. FWB contributed funding to support the studies, contributed to the surgical procedures to procure placentae, contributed to the interpretation of data, and contributed to manuscript preparation. GAJ contributed funding to support the studies, contributed to animal husbandry and the surgical procedures to procure placentae, directed the studies, contributed to interpretation of data, and contributed to manuscript preparation.
This research protocol was approved by the Institutional Animal Care and Use Committee of Texas A&M University (Animal Use Protocol number 2015-0396).
The authors would like to thank the staff, graduate students, and undergraduate students in the laboratories of GAJ, GW and FWB for their significant assistance in animal husbandry, animal surgeries and sample collection.
This project was supported by Agriculture and Food Research Initiative Competitive Grant no. 2016-67015-24955 to GAJ and FWB and Grant no 2015-67015-23276 to GW, FWB and GAJ from the USDA National Institute of Food and Agriculture.
The authors declare no conflict of interest. GW is serving as the editorial board member of this journal. We declare that GW had no involvement in the peer review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to BMA and GP.