











 , Demin Duan 5,*
, Demin Duan 5,*1 Faculty of Environment and Life, Beijing University of Technology, 100124 Beijing, China
2 College of Bioscience and Biotechnology, Shenyang Agricultural University, 110866 Shenyang, Liaoning, China
3 Nanopeptide (Qingdao) Biotechnology Ltd., 266000 Qingdao, Shandong, China
4 Chinese Academy of Inspection and Quarantine, 100176 Beijing, China
5 CAS Engineering Laboratory for Nanozyme, Institute of Biophysics, Chinese Academic of Science, 100101 Beijing, China
Academic Editor: Elena Levantini
Abstract
Background: This attractive and intriguing Ribonucleic acid (RNA) nanotechnology has been conceptualized over the last two decades and with our increasing understanding of RNA structure and function and improvements of RNA nanotechnology it is now possible to use this in clinical settings. Methods: Here we review the unique properties and the recent advances in RNA nanotechnology and then look at its scientific and preclinical applications for tumor diagnosis and targeted delivery and RNA-based therapy using RNA nanoparticles with diverse structures and functions. Finally, we discuss the future perspectives and challenges to RNA nanotechnology. Results: RNA can be designed and manipulated in a similar way to DNA while having different rules for base-pairing and displaying functions similar to proteins. Rationally designed RNA nanoparticles based on the three-way junction (3WJ) motif as the core scaffold have been extensively explored in the field of nanomedicine and targeted cancer diagnosis and therapy. Conclusions: RNA nanostructures based on 3WJs demonstrate promising future applications due to their thermal stability, molecular-level plasticity, multifunctional chemotherapeutic drug delivery and other intrinsic characteristics, which will greatly improve the treatment of cancer and promote further major breakthroughs in this field.
Keywords
- RNA nanotechnology
- 3WJ motif
- Bacteriophage phi29 pRNA
- RNA nanoparticles
- targeted cancer therapy
Ribonucleic acid (RNA) nanotechnology is an emerging field because RNA can be used as a unique polymeric material to construct a variety of nanostructures, such as nanoparticles, bundles, membranes, and polygons [1]. Recent advancements in RNA nanotechnology have made this application possible to various fields including RNA therapeutics for potential cancer treatment and the RNA nanoparticle is one candidate for targeted delivery of therapeutic drugs to cancer cells and tumors. Specific targeted delivery and negligible toxicity are the two major advantages of RNA nanoparticles in cancer therapy when compared to other nanovesicles [2].
Due to its enzymatic instability and susceptibility to ribonuclease (RNase)
degradation, RNA has been overlooked as a potential nanovesicle for therapeutic
drug delivery. Recent studies however, have found that modifications introduced
at the ribose 2
Three-way junctions are common structural and functional motifs in biological RNAs, which are stabilized via extensive tertiary structures and non-canonical base interactions including base pairing and base stacking [5]. The 3WJ motifs are distributed in ribosomes, ribozymes, and ribozyme P or signal recognition particles, which play important roles in the structure and function of RNA or the ribonucleoprotein system [6]. In 2011, Shu’s group thoroughly investigated 25 different 3WJ motifs in RNA [4, 5, 7]. As a platform, the 3WJ can be used as a target recognition system, with the three branches extended to carry versatile functional modules, which can be used as therapeutic, imaging, and targeting modules without affecting overall structure and stability. The unique enzymatic and thermal stability of pRNA-3WJ with its large payloads, makes it an important tool for RNA nanotechnology and nanomaterials design.
RNA nanotechnology not only shares common characteristics to DNA, but also possesses a range of unique properties. One of the great advantages of RNA nanotechnology is that different RNA motifs such as riboswitches, catalytic domains and DNA packaging units which play vital roles in genetic and metabolic processes, can be used as building modules individually, or simultaneously conjugated to the core scaffold to achieve their physiological functions. These well-known biological RNA motifs can be spliced and assembled with synthetic RNA fragments to enable those structures to possess more complex and diverse functions [8]. In addition, the inherent complementary interaction of genetic translational mechanisms between RNA and DNA leads to the hybridization of DNA and RNA molecules naturally, which plays a synergistic role between DNA with relative structural stability and RNA with abundant chemical diversity thus, enabling nanostructure design with different functions [9].
Nanotechnology typically uses “top-down” or “bottom-up” approaches to create nanoscale materials. The “top-down” approaches generally take materials of larger sizes (microns to centimeters) and use traditional techniques such as cutting, etching, and grinding to produce the desired nanostructure. The “bottom-up” approach uses some smaller structural units (such as atoms, molecules, or nanoparticles) to self-assemble through weak interactions to form a relatively large and complex nanostructure. Biomacromolecules such as DNA, RNA and proteins have inherent characteristics of nanoscale and can be used as powerful building blocks for “bottom-up” assembly of nanostructures and nanodevices [10]. DNA and protein have been studied extensively for applications in the field of nanotechnology [11, 12, 13, 14] but RNA, a different natural building block, has been overlooked for some time.
The first proof-of-concept demonstration of the feasibility of RNA nanotechnology was published in 1998 by Guo, where RNA nanoparticles were constructed through self-assembly of multiple reengineered natural packaging RNAs (pRNA) from the bacteriophage phi29 DNA packaging motor molecules [15]. RNA nanotechnology has gained more attention from the nanoparticle field in the last decade as people have gradually realized the potential of RNA nanotechnology for the treatment of cancer, viral infections, genetic diseases and other nanomedical applications [16, 17]. In 2010, Nature Nanotechnology published a review on RNA nanotechnology [10] and soon afterwards, a subsequent article showed that bacteriophage phi29 pRNA-3WJ could be used to construct multi-functional nanoparticles with therapeutic effects due to their thermodynamic stability [4]. These studies have given a tremendous boost to the progress of RNA nanotechnology and currently, the concept of RNA nanotechnology is defined as a specific research field for the design, processing, fabrication, and application of RNA nanoparticles. The main framework, targeted ligand, regulatory components, and therapeutic modules of the resulting nanoparticles can be composed of RNAs [2].
Each nucleotide is composed of a ribose sugar, phosphate group, and a nitrogenous base. The four nitrogenous bases that make up the ribonucleotides are adenine (A), uracil (U), guanine (G), and cytosine (C). Much like DNA, RNA molecules also exhibit complementary base pairs. Thymine (T) in DNA is replaced by uracil (U) in RNA to form A-U and G-C base pairs, which makes RNA nanostructures programmable like DNA. At the same time, unlike the DNA double helix structures which are usually formed by strictly following Watson-Crick base pairing, naturally occurring RNA usually exists in the form of single strands and possesses versatile sequences, secondary structures, and tertiary/quaternary interactions [2, 10].
Some non-canonical base pairs often appear in RNA secondary structures, which do not conform to the Watson-Crick principle [18], such as G-U wobbles, G-A pairing, A-U reverse Hoogsteen pairing. The tertiary structures of RNA, however, are more diverse, including helices, rings, bulges and stems, which can form special motifs and interact with each other [19, 20]. Non-Watson-Crick base pairs, loop-receptor interactions, base stacking and complex tertiary structural interaction systems not only increase the versatility of RNA structure, but also enhance the thermodynamic stability [16].
In addition, many RNA sequences are chemically reactive, and are especially catalytic. In the past, it was generally believed that only proteins had catalytic activity. However, with the discovery of ribozymes, the study of RNA structure revealed they had catalytic properties [21], and this revolutionary discovery suggested a new understanding of the function of RNA. Now it is well known that RNA has many functions such as protein synthesis, gene regulation and catalytic activity.
RNA is extremely sensitive to RNase degradation, which is responsible for its
very short half-life and therefore, the pharmacokinetic performance of most RNA
molecules is poor, which limits their application as a drug therapy in
vivo. However, chemical modification of RNA can overcome this shortcoming, for
example, fluorine, methoxy and amino groups are used to replace 2
RNA nanotechnology is unique and different from traditional RNA structural and functional research. The latter focuses on RNA intramolecular interactions and secondary or tertiary structure, while RNA nanotechnology emphasizes RNA molecular interactions and quaternary structure between motifs to construct the system structure and has medical application [8, 24, 25]. Based on previous studies, functional RNAs, such as ribozymes, riboswitches, miRNAs, and siRNAs, all have been used in the construction of RNA nanoparticles, as well as the discovery of various RNA motifs through structural biology.
To date there have been two clear milestones in the pharmaceutical field throughout medical history, one is based on chemical drugs, and the other based on protein drugs. It is expected that the third milestone will be RNA based drugs/therapeutics or drug based targeting by RNA [2]. In the field of nanobiomedicine, the structure, and chemical properties of RNA, which show unique properties, are conducive to the construction of nanoparticles.
First, RNA nanoparticles tend to be less cytotoxic than other nanovesicles as the negatively charged phosphate skeleton makes RNA nanoparticles accumulate many negative charges, which repel other similarly negatively charged structures such as cell membranes and effectively prevent non-specific targeted binding [26]. Systemic administration of RNA nanoparticles in murine models has shown that they tend to accumulate more in targeted tumors but less in organs, especially the liver and spleen [3]. In terms of beyond liver delivery, their unique biodistribution is superior to other nanovesicles, such as lipid nanoparticles and polymers, which tend to primarily accumulate in the liver and cause hepatic toxicity. Moreover, this high-density anion can be used to prime the protein corona on nanodrugs for targeted delivery, maximizing the RNA nanoparticle efficacy and improving their pharmacokinetics [27, 28, 29, 30].
Second, RNA nanoparticles can be conjugated to multifunctional groups without affecting their native folding or function in both the core scaffold and the individual modules [31]. Because RNA nanoparticles are modular in nature, each component can self-assemble into a multifunctional structure where the pRNA-3WJ motif is a very typical example. Each of the three helical branches of the 3WJ motif can be modified with different subunits, which can load three functional modules at the same time [4, 31]. Chemical ligands, such as fluorescent dyes, chemotherapeutic drugs, or biotin, can be easily coupled to RNA chains through crosslinkers and click chemistry strategies [32]. In this way, multiple functional units for targeting, treatment and tracing can be integrated into one nanoparticle.
Third, the size and morphology of RNA nanoparticles can be designed and controlled, showing good pharmacological effects. They are designed to be larger than 5 nm in diameter which can effectively avoid rapid excretion by the kidney, but they are still small enough to enter the target cells through enhanced permeability and retention (EPR) effects and receptor-mediated endocytosis, and thus avoid capture by immune cells [16]. The favorable size of RNA nanoparticles not only has the potential to improve pharmacokinetics, pharmacodynamics, biodistribution, and toxicology profiles but also can avoid non-specific cell penetration [33]. In addition, RNA nanoparticles also exhibit strong elastic properties, which enable them to squeeze through tumor blood vessels, while maintaining thermodynamic stability in the tumor microenvironment [34]. RNA can also be manipulated to form versatile structures, including twins, rods, triangles, loops, and other 3D shapes [35].
The diversity of structure and function, low energy consumption of folding, the tendency of “bottom-up” self-assembly, controllable size and structure design, and good biocompatibility in vivo, all have helped to make RNA increasingly attractive for nano-biomedical applications.
Although RNA molecules like DNA can be designed and manipulated easily, it remains challenging to construct RNA nanoparticles for in vivo application due to their structural flexibility, RNase sensitivity, and serum instability. One of the main challenges in the design of functional RNA nanoparticles is how to successfully combine “bottom-up” assembly with “top-down” methodology. Shu’s group used palindromic sequences introduced into the 3 prime end of pRNA in 2004, which served as a link for bridging and intermolecular interactions [35]. The formation of RNA nanostructures shows the most advanced self-assembly technology.
There are two basic methods for the construction of RNA nanoparticles. One is to
use the spontaneous self-folding properties of RNA to form a specific structure
through base interaction. Based on Gibbs free energy (
Many types of 3WJs have been found in nature for the fabrication of RNA
nanoparticles and among these, bacteriophage phi29 packaging RNA (pRNA) is
attractive for bottom-up assembly with three RNA fragments and shows strong
thermodynamic stability. Piao et al. [40] found that the thermal stability of phi29
pRNA-3WJs was gradually improved in the following order as DNA/DNA
It is essential to ensure the correct folding of RNA structures in
multifunctional RNA nanoparticles, and a reasonably designed system needs to
ensure that the designed nucleic acid sequence folds into the expected secondary
structure in the presence of its substrate. For each substituted, inserted, or
deleted nucleotide, the secondary structure of the whole system must be
reconfirmed to ensure that it remains folded into the expected active
conformation [41]. RNA molecules sometimes alternate between two conformations
with small
Some software applications and online resources, including Mfold, Sfold,
Nanofolder and Hyperfold, have been developed to predict RNA secondary structure,
however, to predict the accurate structure of unknown RNA nanoparticles remains a
great challenge [16]. The most popular method is to calculate the minimal free
energy of the structural motif formed by base pairing in RNA. The secondary
structure with the smallest change of
In 1987, Guo and co-workers discovered an RNA molecular motor in bacteriophage phi29, which can be powered by ATP to package the viral genomic DNA into a preformed protein shell termed a procapsid [15, 46, 47, 48]. It is unusual that an RNA molecule is an integral component of the packaging motor, because the same roles are always played by subdomains of the larger protein subunits of packaging motors in other double-stranded (ds) DNA phages [49]. This special RNA with packaging function was termed pRNA (packaging RNA), and contains 174-residues. Cleavage of this 174-residue form to a 120-residue form renders it fully functional in vitro [50, 51]. The first proof-of-concept exhibiting the technical feasibility of RNA nanotechnology was published by Guo’s group in 1998 [15]. In the phi29 DNA packaging motor, the pRNA forms dimers as the building blocks for the hexameric conformation. Six phi29 pRNAs can assemble into a hexameric pRNA ring through intermolecular base-pairing, which can be observed by transmission electron microscopy with nanogold-labeled pRNA [52, 53, 54] and this phi29 DNA packaging motor represents one of the strongest biomotors studied to date.
The pRNA monomer contains a 3WJ motif in the central domain and this 3WJ represents a flexible, dynamic region in pRNA [5, 55] capable of helping to place helices correctly in the spatial orientation required for packaging [56, 57, 58]. The fact that different pRNAs only retain 12% sequence similarity, but conserve secondary structure and function, suggests that certain motifs play a compensatory role in the stabilization of pRNA [59]. Studies have also found that pRNA can self-assemble into dimers, trimers, and higher order multimers under different conditions in vitro [4, 15, 60], where the 3WJ represents an important motif and contributes to pRNA self-assembly. Fig. 1 (Ref. [60]) shows the pRNA ball and stick model, where the ball represents the ring and the stick represents the helix and Fig. 2 (Ref. [55]) represents a schematic diagram of the primary and secondary structure of wild type pRNA, and where the 3WJ motif is found within the red box.
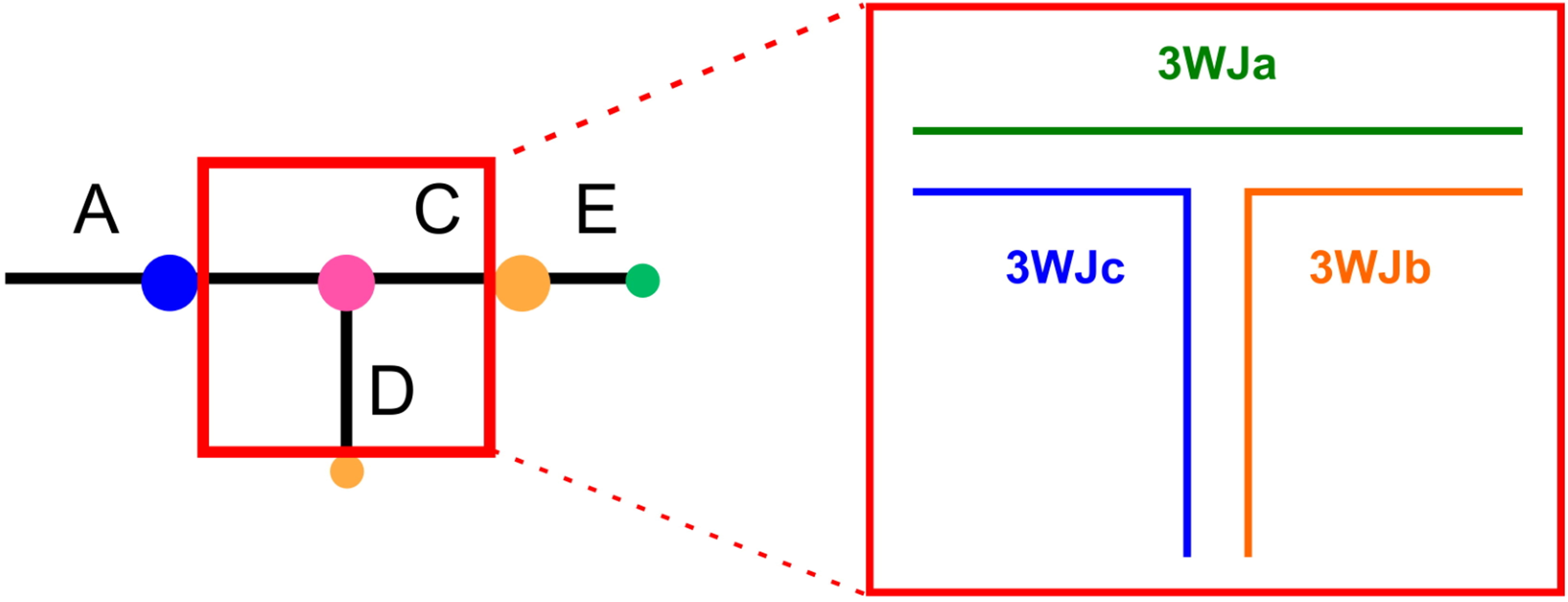 Fig. 1.
Fig. 1.Ball-and-stick model of pRNA. Balls represent loops and sticks represent helices. The pRNA 3WJ nano motif, inset, comprises strands 3WJa (green), 3WJb (orange), and 3WJc (blue). Reproduced with permission from (ref. [60]).
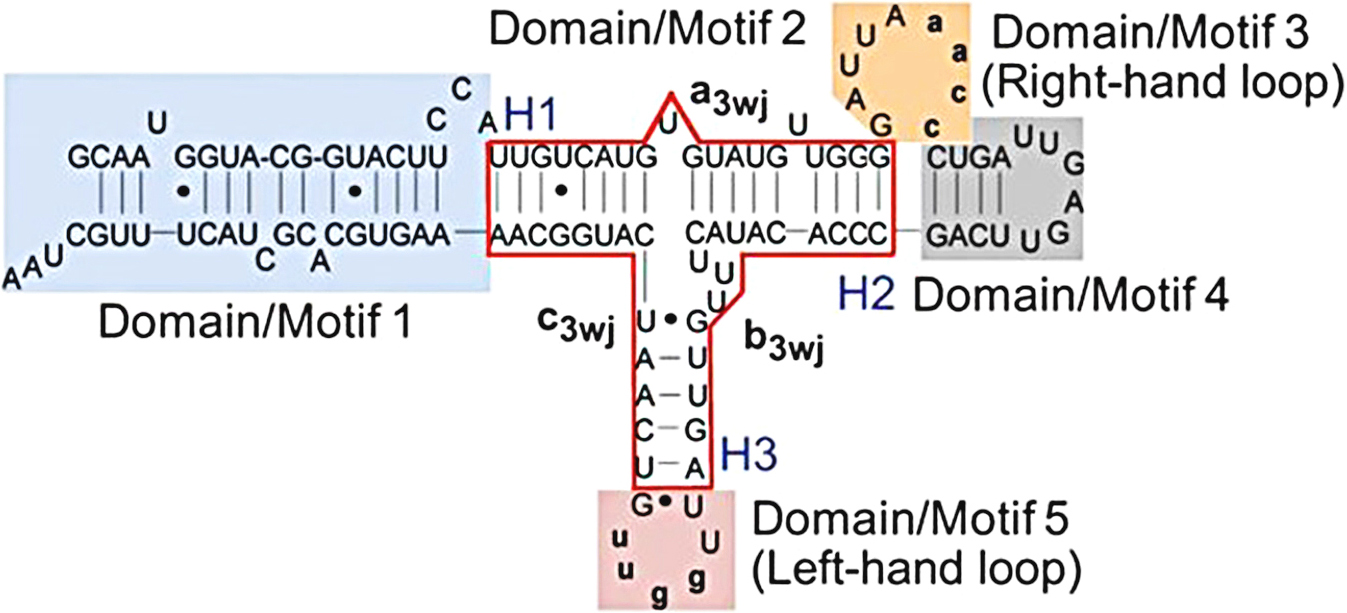 Fig. 2.
Fig. 2.Full-length Wild type pRNA with the 3WJ motif boxed in red. The
three individual RNA strands of the 3WJ are shown as a3wj, b3wj, and c3wj. The
helical segments are designated as H1, H2, and H3, respectively. The interlocking
loop sequences of 5
The crystal structure of the bacteriophage phi29 pRNA-3WJ and the published
structural data were used to construct the model structure. The 3
3WJa 5
3WJb 5
3WJc 5
The underlined nucleotides in the above sequences are unpaired, forming a bulge region. Fig. 3 (Ref. [60]) represents a schematic diagram of the primary and secondary structure of pRNA-3WJ. Among the three strands, 3WJa, 3WJb, and 3WJc have partial pairing complementarity and the unpaired nucleotides in phi29 pRNA-3WJ form bulge regions.
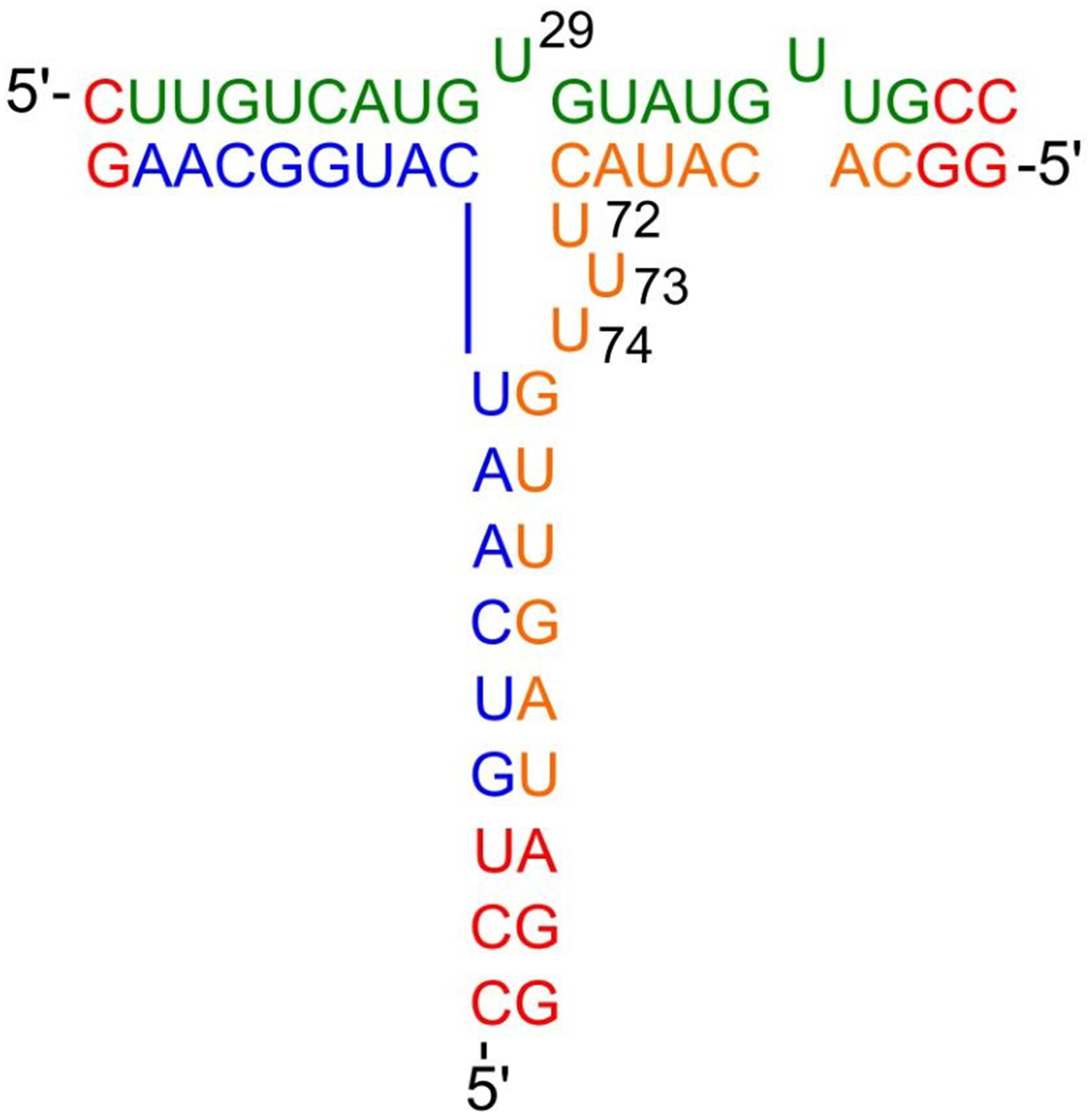 Fig. 3.
Fig. 3.pRNA-3WJ motif primary and secondary structures. Strand 3WJa is in green, 3WJb is in orange, and 3WJc is in blue. Changes to WT sequences are indicated in red. Nucleotides unpaired in the phi29 pRNA 3WJs are labeled. Reproduced with permission from (ref. [60]).
Zhao’s research showed that four Uridine (U) residues in the phi29 pRNA-3WJ (U29 and U72–74) could simultaneously be replaced with Adenine (A) residues without loss of biological function, meaning that the size of the bulge rather than the sequence may be the key to its activity [61]. They also found that, the deletion of one or two U residues showed activity similar to that of wild-type pRNA and the deletion of more than two U residues abolished functional DNA packaging, suggesting that the minimal tolerance for function is two residues; either two residues retained in the tri-U linker or the U29 residue in conjunction with one tri-U linker residue. The phi29 3WJ crystal structure displayed the formation of a cis U-U mismatched base pairing between the Watson–Crick edges of U29 and U72, and at the same time base stacking between U29 and U74 [55, 60, 62] (Fig. 4, Ref. [60]). Research has further suggested that the flexible U residues are very important to pRNA multimerization because no intermolecular base pairing occurs without the flexible U residues [15, 61].
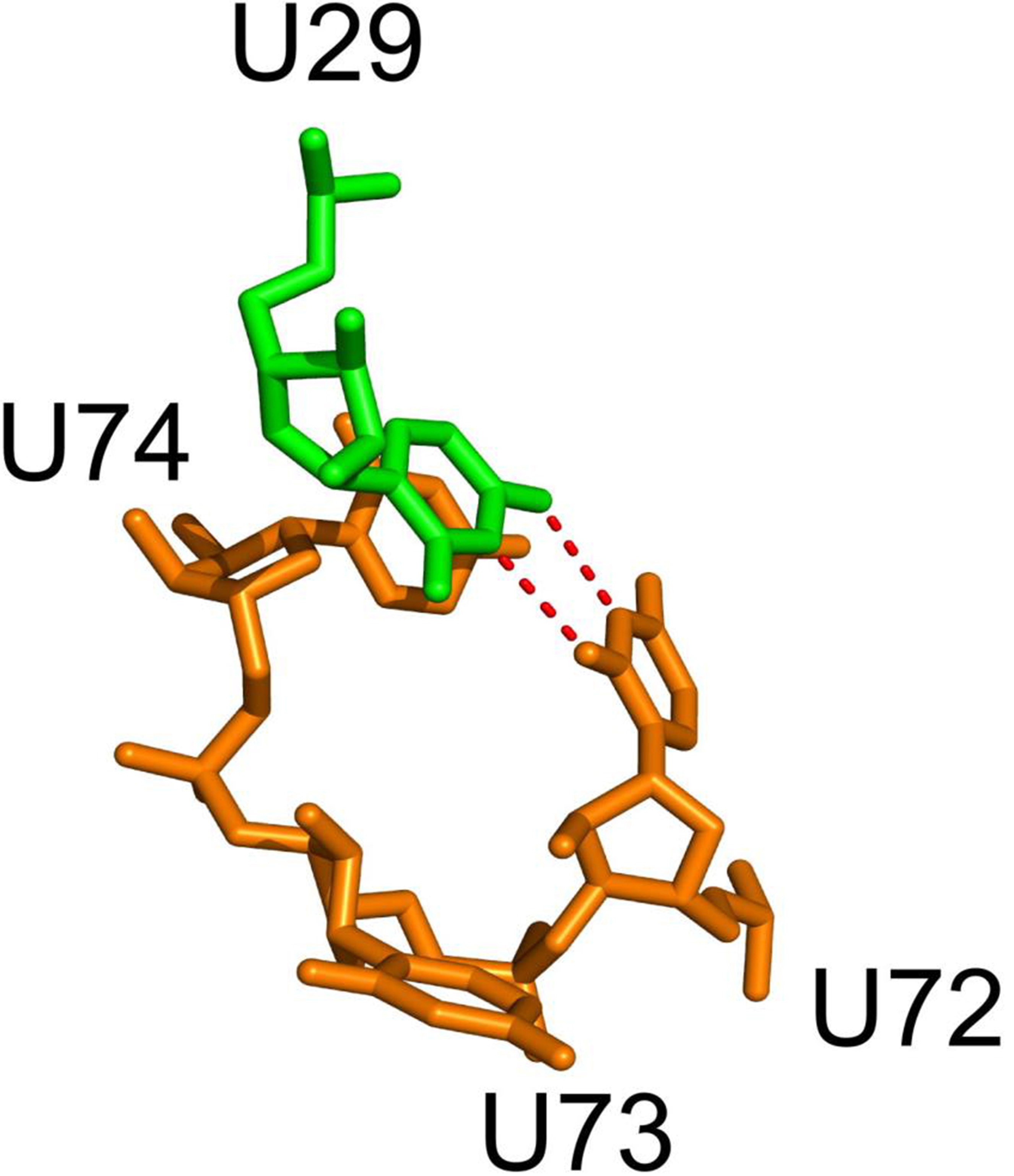 Fig. 4.
Fig. 4.Bulge residues at the 3WJ. Bulge residues at the 3WJ (i.e., U29/U72–73–74) are shown as sticks. A cis base pair forms between the Watson–Crick edges of U29 and U72. U29 and U74 forms a base stack. Reproduced with permission from (ref. [60]).
Divalent metal ions play a critical role in structural stabilization and
function of the pRNA 3WJ and pRNA hexamers. Metal ions such as Mg
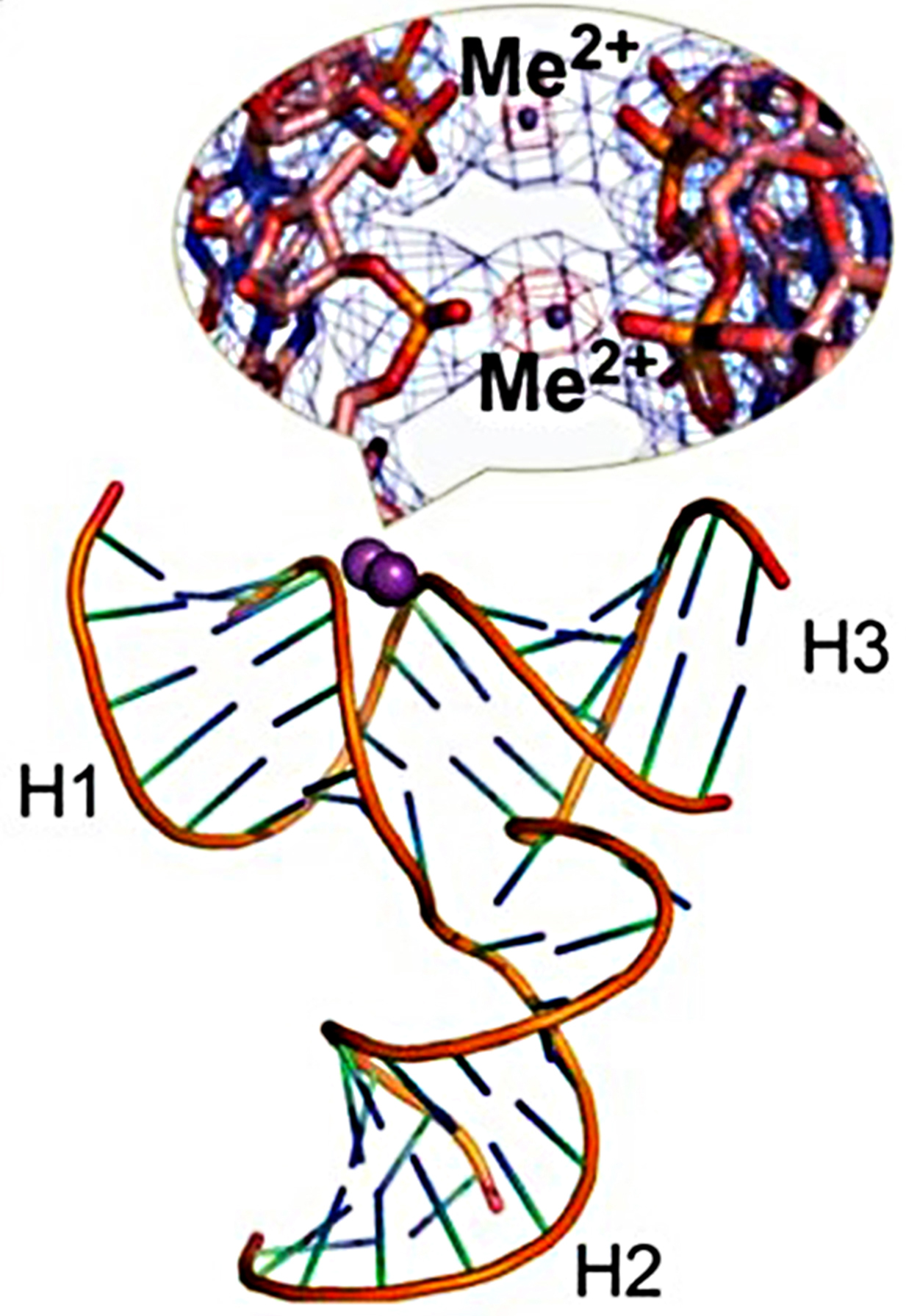 Fig. 5.
Fig. 5.Crystal structure of the pRNA 3WJ with two metal binding sites
(magenta). A close-up view of the metal binding sites superposed on the 2Fo-Fc
electron density map (blue mesh contoured at 1.0
RNA molecules assemble hierarchically from modules or building blocks to form complex three-dimensional structures and these spontaneously and repeatedly formed building blocks and can be packed together to form the molecule [65]. High level modules, like RNA 3WJs, are made of smaller and lower level modules, where Watson-Crick helices are linked together by single strands [66].
In 2006, Lescoute and Westhof investigated the structure of 3WJs in the published crystal structures of 33 folded RNAs [5]. Their findings showed that in 3WJs, three Watson-Crick paired helices were linked by at most three single-stranded segments. Two of the helices P1 and P2 form a coaxial stack, with the third helix P3 at an angle to the stack and three junctions, J12 is between P1 and P2, J23 is between P2 and P3, and J31 is between P3 and P1 (Fig. 6, Ref. [5]). According to the average number of nucleotides in each junction (J12, J23, J31) and the distribution of the three helix branches (P1, P2, P3), 3WJ motifs can be divided in three families A, B and C.
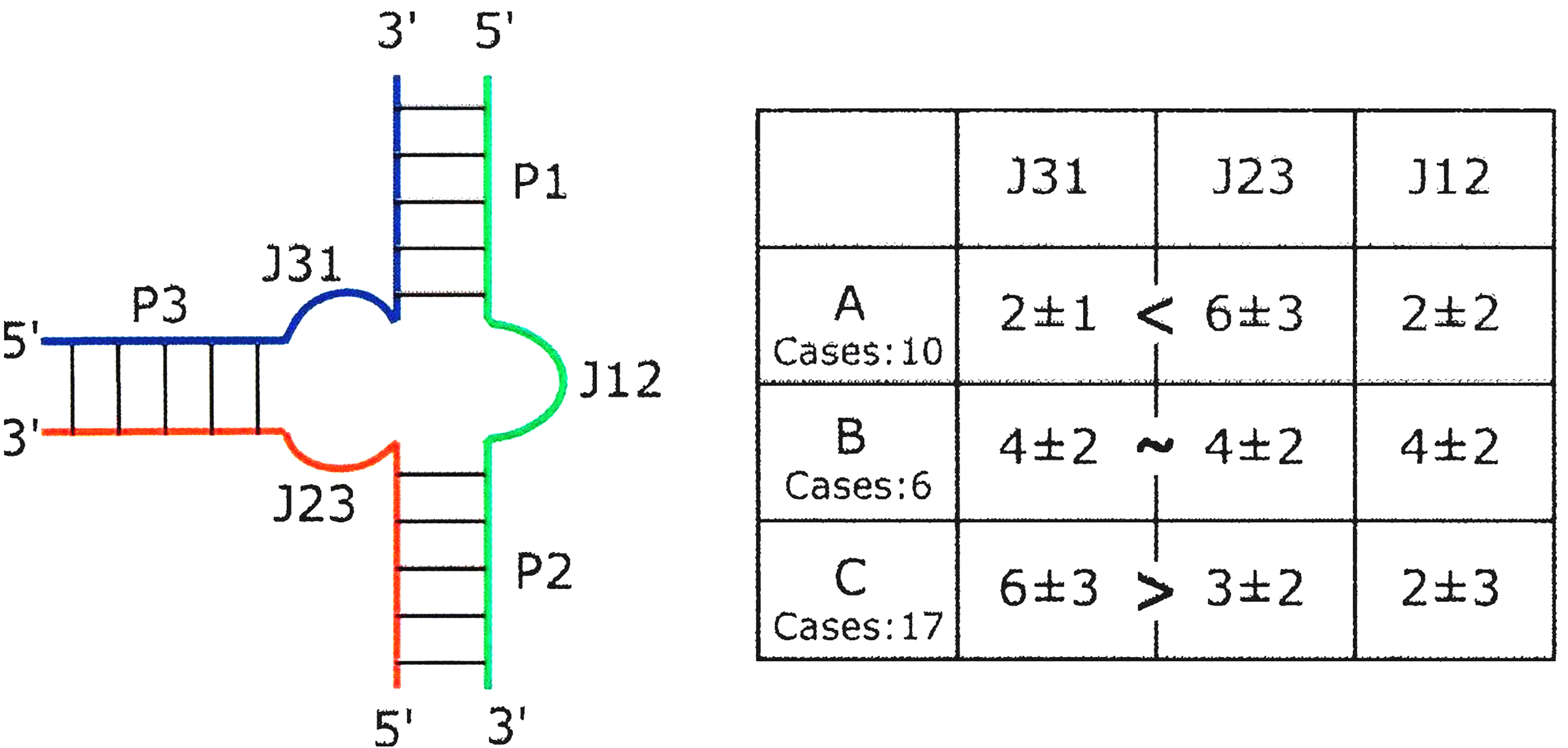 Fig. 6.
Fig. 6.Schematic diagram of the three-way junction (3WJ) structure. The nomenclature used for 3WJs with the average numbers of nucleotides in each junction in the table at the right. The number of instances in each of the three families is also indicated. Reproduced with permission from (ref. [5]).
Lescoute and Westhof found that all 3WJ motifs are presented in a similar way, P3 is always on the left side of stacked helices of P1 and P2 (Fig. 7, Ref. [5]). In most cases, the axes of the three helices are coplanar. There are always three families, where helix P3 is nearly perpendicular to the P1/P2 coaxial stack in family A, helix P3 bends toward helix P2 in family B, and helix P3 bends toward helix P1 in family C.
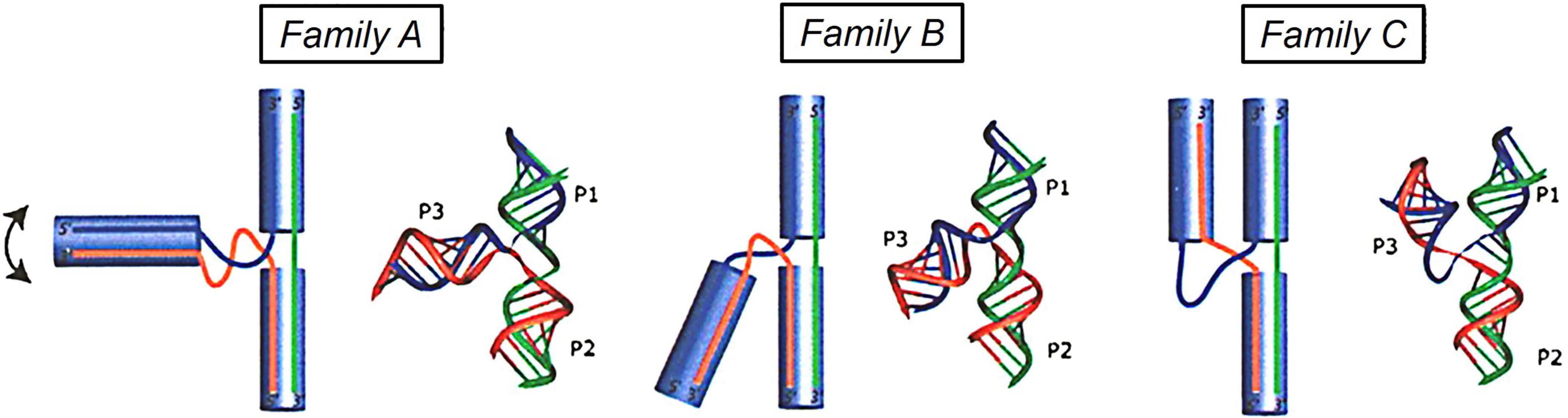 Fig. 7.
Fig. 7.Schematic diagram representing the families A, B and C in the three-way junctions. In family A, helix P3 is nearly perpendicular to the P1/P2 coaxial stack; In family B, helix P3 bends toward helix P2 in family B; and in family C, helix P3 bends toward helix P1. Reproduced with permission from (ref. [5]).
The past decades have witnessed successful RNA nanotechnology used in cancer therapy and other disease treatments, and it can also be used as a vehicle to deliver therapeutics [10, 67]. RNA nanoparticles have been constructed with diverse functions, good biocompatibility, and target specificity, suitable for in vivo drug delivery [31]. The different 3D structures of RNA help us identify useful RNA motifs, such as the pRNA-3WJ, used as building blocks [19]. The pRNA-3WJ has shown great potential as a useful platform for the generation of RNA nanoparticles for cancer targeting and therapy [4]. Based on rational designs, information of geometry derived from 3WJs is essential to produce self-assembling, stable, and polyvalent RNA nanoparticles [20, 33, 37, 44]. The crystal structures of 3WJs give insights into the designs of more effective therapeutic RNA nanoparticles and plays a pivotal role in the construction of RNA nanostructures. Based on previous reports, several techniques have been developed for the construction of RNA nanoparticles with various sizes, shapes, and function.
The size of nanoparticle can play a key role in the delivery, efficacy,
biodistribution and clearance in vivo [68] and de novo design
of RNA nanoparticles is difficult because of the complexity of RNA tertiary
structure. However, by utilizing the robust 3WJ motif as the building block, this
has become much easier. The pRNA-3WJ of bacteriophage phi29 DNA packaging motor,
folds intrinsically into a planar construction with three angles of 60
Adopting a “divergent growth” method and self-assembly strategy, RNA globular
dendrimers can be initiated form a single pRNA-3WJ planar square core and then
grown in a radial manner layer by layer to generate defined sizes ranging from 12
nm to 65 nm, which can be made into ideal drugs; not too small (
The shape of the nanoparticle is of great importance for their interaction with
cell membranes in vivo [71]. Previous work looking at RNA nanotechnology
has enabled the construction of RNA nanoparticles with precisely controlled
shapes. Each different shape has its own unique characteristics, for example,
discoidal-shaped nanoparticles are more prone to migrate to vessel walls and
interact with endothelial cells in blood vessels when compared to spheres [72, 73]. The pRNA-3WJ motif has been extensively developed to form elegant RNA
triangles, squares, and pentagons by stretching the interior angle of the RNA
strands from 60
As seen in images using Atomic Force Microscopy (AFM) (Fig. 8, Ref. [31]), as well; as the traditional trimers, tetramers, pentamers, hexamers, and heptamers, RNA nanoparticles based on pRNA-3WJ also include triangles and quadrilaterals, pentagons, and more complex 3D structures such as tetrahedrons, triangular prisms, and dendrimers. The tetrahedral structure of RNA with high mechanical rigidity and structural stability are envisioned to have a major impact on cancer targeting and disease therapy [39]. Furthermore, 3D RNA nanocages resembling triangular prisms based on pRNA-3WJs with fine control of geometry have been created to successfully bind and deliver drugs into cancer cells, tremendously benefiting the advancement of nanomedicine [74].
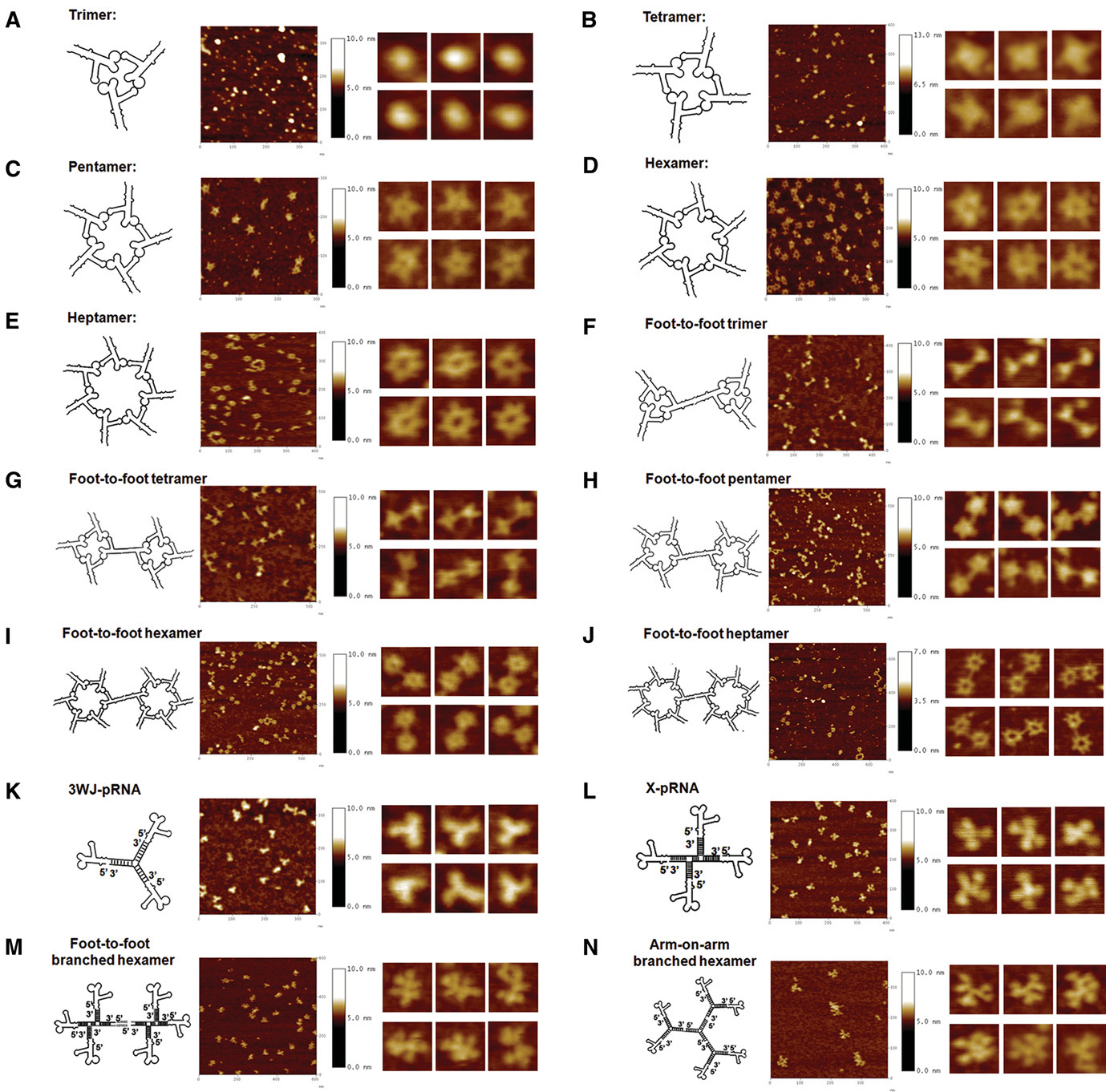 Fig. 8.
Fig. 8.AFM images of diverse pRNA nanoparticles. A-N are different pRNA nanoparticles with tunable shapes. The second column represents magnified images of individual nanoparticles. Reproduced with permission from (ref. [31]).
The pRNA-3WJ carries a branched construction allowing various RNA functional
moieties to conjugate at each branch end. Each of the three branches of the
pRNA-3WJ offers an overhanging, reverse fragment that serves as a sticky end to
carry functional groups. Therefore, targeting aptamers, therapeutic modules, such
as small interfering RNAs (siRNAs) or miRNA, and regulatory ribozymes can be
designed to link to the 5
How the problem of dealing with non-specific accumulation of therapeutic nanoparticles in healthy vital organs which increase their toxicity and side effects, remains the main challenge for cancer research. With the development of the systematic evolution of ligands by exponential enrichment (SELEX) to discover the oligonucleotides that specifically bind to cell surface receptors in order to reach tumors, tens of RNA aptamers have been identified, for example EGFR and CD4 [69]. Incorporation of RNA aptamers to pRNA-3WJ nanoparticles, where functional sequences are fused into core strands without affecting their original folding, has helped aptamers internalize into cancer cells via receptor-mediated endocytosis and this then lowers the off-target effect of the RNA nanoparticles [75].
RNA nanoparticles have an unusually high tumor targeting efficiency [76] and in vivo animal studies have revealed that RNA nanoparticles can specifically target tumors with favorable pharmacokinetic and pharmacodynamic parameters without unwanted accumulation in healthy organs [2]. Because they can be rapidly cleared from the body via renal excretion with little accumulation, it is expected that therapeutic application of RNA nanoparticles will increase for the detection and targeting of cancer. Here, we summarize some typical applications for RNA nanoparticles based on 3WJs used for cancer therapy.
MicroRNAs (miRNAs) are single-stranded non-coding RNAs composed of 18–24 nucleotides, which are critical for the regulation of gene expression and the translation of messenger RNAs. Irregular expression of miRNAs is always associated with many diseases including tumors, indicating that they may represent potential biomarkers for cancer diagnostics and as targets for treatment. These properties inspired Luo’s group to develop a fluorescence resonance energy transfer (FRET)-based assay for the accurate determination of the composition and concentration of miRNAs in total RNA extracts obtained from clinical samples, such as blood or tissue biopsies [77].
It is well-known that FRET-based assays rely on the energy transfer from a fluorescence donor to an acceptor at very close distances, within 10 nm. In this regard, conventional Cyanine3 (Cy3) and Cyanine5 (Cy5) dyes were utilized, where a Cy3-labeled DNA probe 1 and a Cy5-labeled DNA probe 2 were used as the donor and the acceptor, respectively. The target miRNA partially hybridized with both complementary sequences from probes 1 and 2, and these two probes also formed a short duplex to assemble a 3WJ structure, which implemented the optimal distance between the FRET pair [77].
The mutual hybridization of the duplexes within the 3WJ enabled thermal stability of the duplex at room temperature and this strategy can be operated within the linear range from 500 pM to 100 nM concentrations of miRNAs. After application in real biological samples, it showed extremely high sensitivity and specificity and was able to recognize even a single base mismatch, which is extremely efficient when compared to traditional approaches [77].
Carcinoembryonic antigen (CEA), is a type of glycoprotein, and represents a broad-spectrum tumor marker associated with colon, breast, and lung cancers, and its concentration in serum from cancer patients is significantly higher than serum from healthy subjects [78]. In 2017, a label-free assay for the detection of CEA in serum was established, using a 3WJ pocket and a fluorescent agent, which could be used in clinical cancer screening and early diagnostic applications. Aptamers (Apts) of CEA were isolated from nucleic acid libraries via the SELEX process, which could bind with high affinity and specificity to small dyes. Two complementary chains with GC repeated nucleotides CS1 and CS2 were synthesized, and 5,6,7-trimethyl-1,8-naphthyridin-2-amine (ATMND) was used as a fluorescent dye. It has been shown that ATMND can be trapped on the 3WJ structures especially those with GC nucleotide repeats, resulting in fluorescence quenching of ATMND. All of those structures exhibited the formation of 3WJ pockets and had ATMND entrapped within the pocket [79].
In absence of CEA, aptamers and their complementary strands CS1 and CS2 formed a 3WJ structure and when ATMND was added to it was locked into the 3WJ structure, leading to a weak fluorescence intensity. When CEA was present in the detection environment, the Apt/CEA complex was formed, and this was unable to construct a 3WJ structure. Thus, after addition of ATMND to the sample, a strong fluorescence intensity was detected [79]. The aptasensor showed good performance for the detection of CEA with a wide linear range of 4.5 pg/mL–30 ng/mL. The detection limit (LOD) was defined as 1.5 pg/mL under a signal-to-noise ratio of 3:1, which was lower than other detection methods. Therefore, this method shows great potential for real life sample analysis and can be extended to other types of aptamers for the detection of a wide range of targets.
Specific targeting and drug delivery applications using RNA nanotechnology can offer accurate delivery to cancer cells. The pRNA-3WJ of phage phi29 has the inherent property of spontaneously folding into an “arrow” plane and by adding a targeting ligand, where three classes of targeting ligands are always used—folate (FA), prostate-specific membrane antigen (PSMA) RNA aptamer, and epidermal growth-factor receptor (EGFR) RNA aptamer, the pRNA-3WJ can specifically bind to overexpressed receptors on cancer cells. At the nanometer-scale extracellular vesicles (EVs) play important roles in intercellular communication. They can deliver biomolecules into cells by direct membrane fusion through tetraspanin domains [80, 81], which have broad applications as therapeutic carriers. However, EVs lack selectivity and can randomly and hazardously fuse to healthy cells. To construct specific targeting EVs, many groups are currently engaged in this research and have made outstanding contributions.
The engineered pRNA-3WJ was used by Pi’s team to decorate EVs purified from
HEK293T cell culture supernatants, applying the specific binding of EVs to target
cells and transport the small interfering RNA (siRNA) carried by 3WJs to tumor
cells, effectively blocking tumor growth [82]. The engineered pRNA-3WJ
carried different functional groups on the three spiral branches, the 3
Survivin is an inhibitor of cell apoptosis and an attractive target for cancer therapy, as its knockdown can decrease tumorigenicity and inhibit metastases. Some siRNAs can silence the gene encoding Survivin, thereby causing cancer cell apoptosis. After loading specific siRNAs into EVs and decorating them with PSMAapt/3WJ/cholesterol, the RNA nanoparticles showed perfect targeting to the corresponding receptor overexpressing cancer cells. The presence of PSMA aptamer greatly enhanced the targeting of EVs to prostate cancer cells, enabling siRNA to reach the target effectively.
In this study, if the end of 3WJ c
Pi also conducted further studies on cholesterol loading positions between the
arrowhead and arrow tail of the 3WJ. It was found that cholesterol on the
arrowhead promoted the entry of RNA nanoparticles into EVs and in the arrow tail
configuration, RNA passing through the membrane was prevented. It seems
as if the two 60
Based on the above work, ginger derived exosome-like nanovesicles (GDENs) were used as an siRNA delivery vesicle. In comparison to EVs derived from mammalian cells, plant exosome-like vesicles do not require large scale cell culture have increased yields and are lower in cost. However, GDENs may not be able to carry ligands and cargos that are recognized by human cells for targeting. The 3WJ functional group anchored on the surface of GDENs was used as a target guide to transport siRNA to tumor cells, and the tumor suppressive effect was achieved by intravenous injection [83].
The arrow tail pRNA-3WJ was designed for end-labeling with a fluorescent dye
Alexa647 on the 5
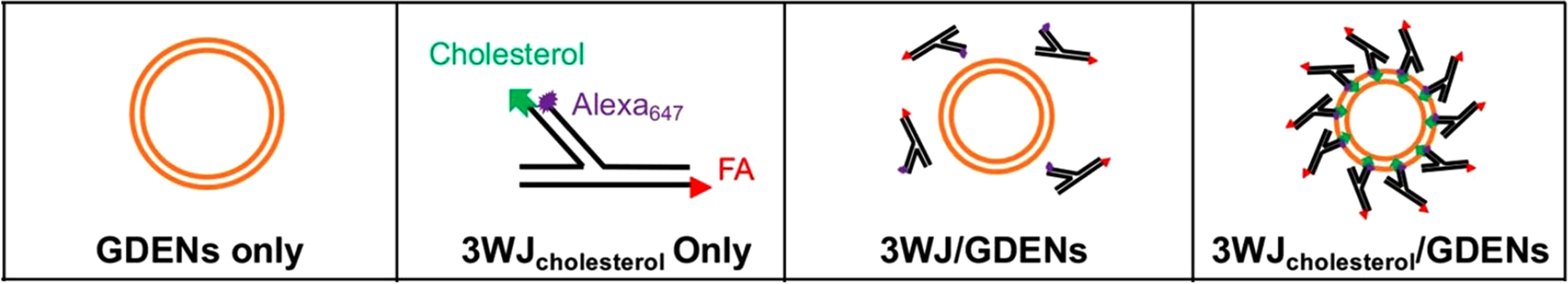 Fig. 9.
Fig. 9.Assembly mechanism for ligand displaying GDENs. The arrow tail
pRNA-3WJ was designed as a FRET accepter by end-labeling with a fluorescent dye
Alexa647 on the 5
In terms of cancer treatment, differentiation therapy is an important method as an adjunct to traditional chemotherapy or radiotherapy. Differentiation therapy results in irreversible changes in cancer cell phenotypes, which not only reverse the cell maturation process with less toxicity but also reverts the cancer cell back to normal cells without damaging the non-target cells [84, 85]. Nanoparticle (NP)-based cell differentiation therapy has attracted increasing research interest and has become a very promising alternative to traditional cancer treatment methods.
A silver Bi
The 3WJ designed in the above study integrated four functions into the three legs of the 3WJ (Fig. 10, Ref. [86]). One leg of the junction was conjugated with Ag@Bi2Se3 and to increase the colloidal stability of Ag@Bi2Se3, the surface was treated with polyvinylpyrrolidone (PVP). The second leg harbored a cell-penetrating RNA and a florescent tag and the third was designed to inhibit micro-RNA-17 (miR-17) and to further release retinoic acid (RA), which had a therapeutic role. Considering the cytotoxicity of RA, precise control of its slow release is crucial to avoid induction of apoptosis. A new drug delivery methodology using RA conjugated with an RNA strand (RA/R) through a stimulus-responsive cross-linker strategy in which the RA release requires the miR-17 inhibition step, can not only achieve the slow release of RA inside the cytosol, based on the inhibition of miR-17, but also resolve the hydrophobicity challenges of RA.
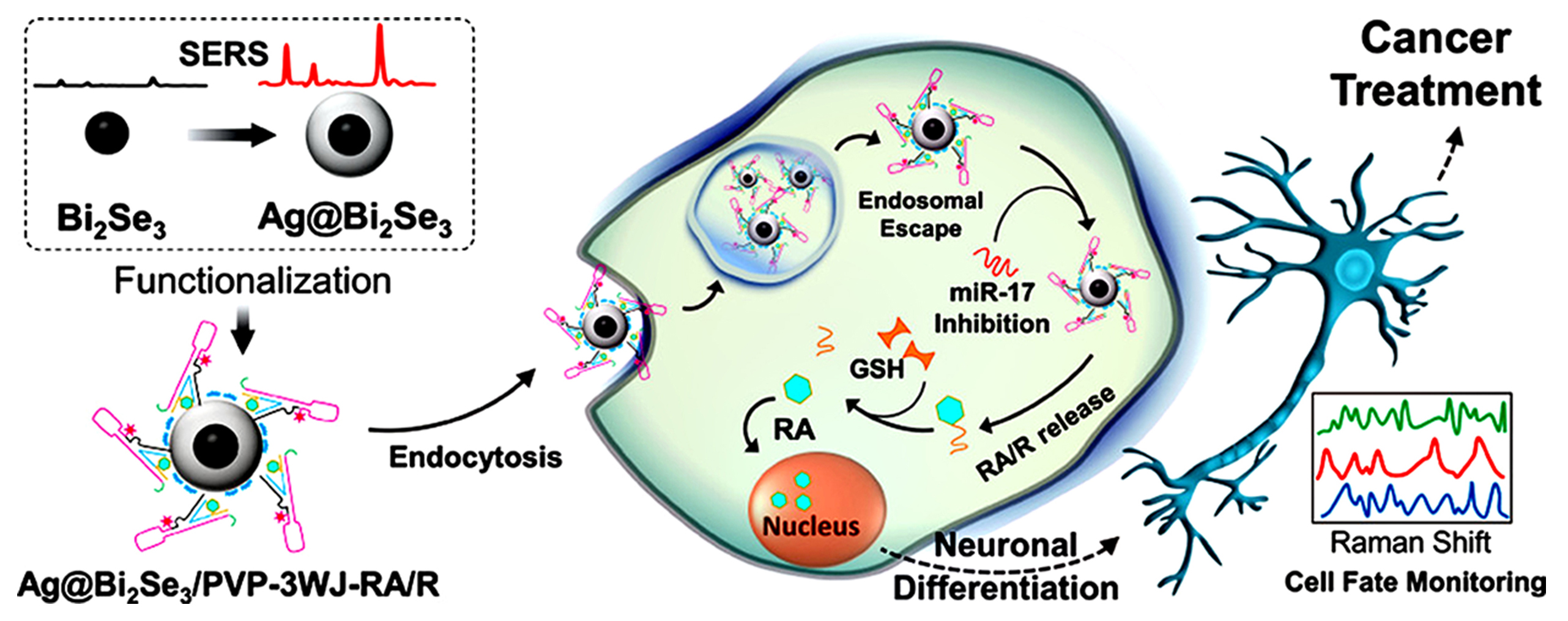 Fig. 10.
Fig. 10.Schematic diagram of the Ag@Bi2Se3/PVP-3WJ-RA/R construction and its application in NB cell differentiation. R represents the small RNA strand conjugated with RA. Reproduced with permission from (ref. [86]).
During this research, interestingly cell-to-cell differentiation was observed in which the differentiated cells containing NPs were attempting to locate and differentiate the undifferentiated cells within their vicinity by growing their axons toward them [86]. Soon with the help of RNA nanotechnology, NPs with controllable multifunctionalities will be developed for any type of cancer to produce robust cancer differentiation. Currently, this phenomenon of the cell-to-cell differentiation is interesting to many researchers attempting to develop novel devices and apparatus in the field of bioelectronic medicine.
During the past few years, immunotherapy has made a breakthrough in cancer therapy based on the discovery that monoclonal antibodies (mAbs) targeting co-inhibitory immune checkpoint proteins can stimulate effector functions and can induce the body’s immune response against cancer cells [87]. However, some immunotherapies based on proteins displayed nonspecific interactions, immunogenic side effects, and unfavorable biodistribution [88]. Antibody-like (Ab-like) RNA nanoparticles with various immune checkpoint-targeting aptamers can be formed carrying multiple ligands to bind different cell receptors with little accumulation in vital organs. The ultra-stable pRNA-3WJ was used to harbor RNA aptamers for immune checkpoint binding and showed great potential for cancer immunotherapy.
Using a bottom-up self-assembly approach, the pRNA-3WJ core can be assembled
from three single-stranded RNA (ssRNA) oligonucleotides, where the sequence was
derived from the central domain of bacteriophage phi29 pRNA. Then, the extended
3WJ nano-scaffold was equipped with anti-CD28 (selectively binding CD8
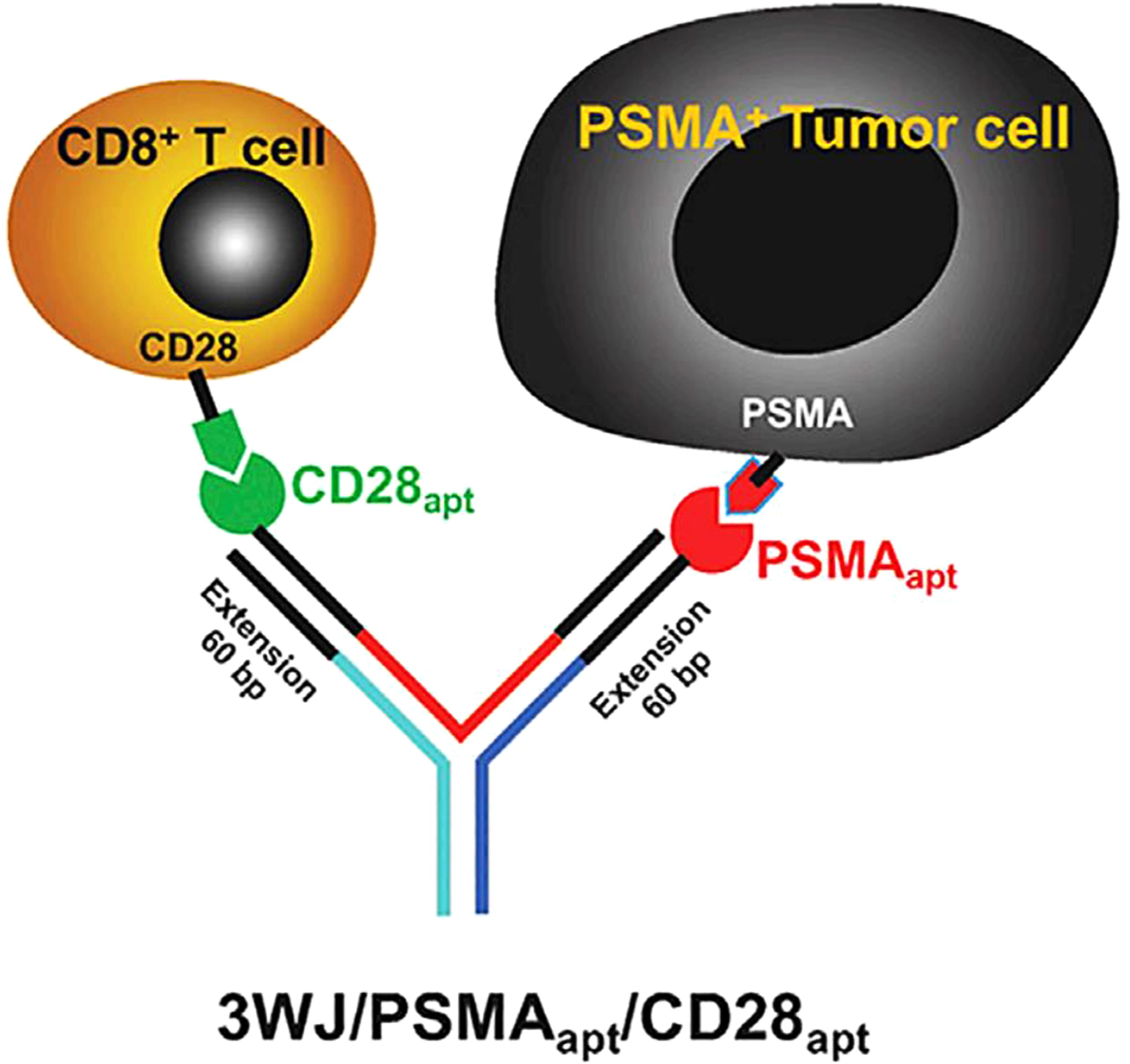 Fig. 11.
Fig. 11.Schematic diagram of bi-specific Ab-like RNA NPs. Reproduced with permission from (ref. [89]).
As a biocompatible material, RNA has great advantages in nanotechnology due to its versatility in structure and function. RNA nanotechnology makes it possible to develop various RNA nanostructures with defined shape and size, tunable thermodynamic, physicochemical properties for diverse applications in nanobiotechnology. RNA-based drugs have achieved much attention recently on account of their enormous potential as potential treatment for many diseases, especially cancer. Although RNA nanotechnology has made great progress in biomedical applications, there are still some challenges remaining, which need further consideration and careful design to overcome, when applying nanotechnology and bringing its products to the market. The effects of nucleic acid toxicity of the RNA nanoparticles used as therapeutic agents should be further tested in preclinical studies using non-human primates before used in humans. Herein some of the challenges and possible solutions are given.
One of the main obstacles for the use of RNA nanostructures in therapeutics is
the chemical instability of RNA itself. Natural RNA is very sensitive to RNase
(RNase) degradation and is especially unstable in the body or serum. Studies
have shown that chemical modifications can offer some advantages to increase
RNase resistance. Replacing the 2
The thermodynamic stability is essential for the use of RNA nanoparticles as therapeutics. The application of bottom-up assembled RNA nanoparticles in the body often dissociates when the concentration in the body is very low after systemic injection. However, this situation is no longer worrying. Studies have found that the pRNA-3WJ complex was thermodynamically stable in serum and remained intact at ultra-low concentrations [4, 33]. A type of synergistic tetravalent RNA nanoparticle has been injected into the tail-vein of mice and revealed that the RNA nanoparticles remained intact and strongly bound to cancers without entering any other organs or tissues, while remaining in cancer tissue for more than 8 h [33]. A different RNA 3WJ nanoparticle was reported to solubilize hydrophobic drugs for targeted delivery in human tumor xenograft models, which showed that it not only improved drug solubility in physiological conditions but also maintained high thermal and chemical stability [95]. As well as the phi29 pRNA, there are many other types of RNA molecules that could be used for nanotechnology-based therapy such as small interfering RNAs, and ribozymes [96]. However, only the 3WJ core of phi29 pRNA incorporating different therapeutic functional structures exhibits all the original functions in vivo and in vitro and therefore, represent potential therapeutic agents.
When using RNA nanostructures for treatment, side effects and toxicity to
healthy organs have always been major concerns. It is very important to ensure that
the nanoparticles have favorable pharmacological profiles such as
biodistribution, half-life and clearance rate, immune response, and specific
targeting. Recent research has reported that the half-life of the pRNA
nanoparticles was determined to be 6.5–12.6 h, when compared to control
2
Predicting complex RNA structures remains a huge challenge. New computational methods need to be developed to balance the kinetic aspects of RNA folding and the thermodynamic control mediated by rigid motifs. The ab initio calculation of RNA nanostructures may provide more diversity in structure and function than the applications we expect in nature, and it also provides the possibility to construct complex RNA structures [45]. The RNA motif database is much smaller than the database of small molecules and proteins and as a greater number of RNA structures are published, they need be catalogued with available thermodynamics, kinetics, and folding properties. While progress has been made in utilizing the thermodynamically stable pRNA-3WJ, endogenous production of programable functional RNA nanoparticles will bring great leaps in synthetic biology.
The pRNA-3WJ motif with extraordinary thermal stability, as the basic component of RNA nanostructure, has greatly promoted the development and application of RNA nanotechnology. The anionic nature, thermodynamic stability, self-assembly ability, and other characteristics of RNA, such as versatility, molecular-level plasticity, and potential semiconductor properties, will enable RNA to be used in therapy, detection, sensing, and other applications. More importantly, the pRNA-3WJ demonstrates a promising nano-platform for multifunctional chemotherapeutic drug delivery for cancer therapy with a high-yielding drug payload, tumor-specific targeting capabilities, and reduced adverse effects.
XL was at the origin of the text, collected the literature and wrote the paper. DD (Dihan Duan) helped to analyze the data and edit the manuscript drafts. YW made suggestions and partial revisions to the manuscript. JL and DD (Demin Duan) conceptualized, planned, edited, and finalized the manuscript. All authors read and approved the final manuscript.
Not applicable.
We thank Xinjiang Uygur Autonomous Region Product Quality Supervision and Inspection Institute for providing research platform and financial support. Demin Duan is thankful to CAS Engineering Laboratory for Nanozyme, Institute of Biophysics, Chinese Academic of Science, Beijing for providing the infrastructure and facility for research.
This research was funded by Special Project of Xinjiang Autonomous Region innovation environment (talent, base) construction, grant number PT2106; National Natural Science Foundation of China, grant number 82072324; Special fund support project of basic scientific research business expenses of Chinese Academy of inspection and quarantine, grant number 2016JK013; National Science and technology major project, grant number 2018ZX10101004002004; open project of Beijing field evidence inspection Engineering Technology Research Center, grant number 2016CSEEKFKT04.
The authors declare no conflict of interest.