






1 Department of Biotechnology, All India Institute of Medical Sciences (AIIMS), 110029 New Delhi, India
Abstract
In 2020, a novel strain of coronavirus (COVID-19) has led to a significant
morbidity and mortality worldwide. As of the date of this writing, a total of 116
M cases has been diagnosed worldwide leading to 2.5 M deaths. The number of
mortalities is directly correlated with the rise of innate immune cells
(especially macrophages) in the lungs that secrete inflammatory cytokines
(IL-1
Keywords
- COVID-19
- Cytokine Storm Syndrome
- CSS
- Immune system
- Phytotherapeutics
- Review
In December 2019, uncommon pneumonia like symptoms began to appear in citizens
of Wuhan city, China. A research group at virology institute of China started
identifying the agent responsible for the development of these atypical symptoms.
Next generation sequencing (NGS) data of nucleic acids isolated from the
bronchoalveolar lavage fluid (BALF) of diseased patients revealed the presence of
a novel coronavirus strain which was provisionally named as 2019-nCoV [1]. The
NGS data analysis revealed that the RNA genome of 2019-nCoV resembles the Severe
Acute Respiratory Syndrome-Coronavirus (SARS-CoV) genome responsible for the
SARS-epidemic during 2003–2004 [2]. Thus, 2019-nCoV was ultimately renamed as
SARS-CoV-2. Later, the disease caused by the novel SARS-CoV-2 was designated as
COVID-19 by World Health Organization (WHO) which also declared it as an
international public health emergency in March 2020 (Fig. 1). The early clinical
symptoms associated with SARS-CoV-2 infection are fever, muscle pain, chills,
sore throat, loss of taste, dry cough, dyspnea (difficulties in breathing),
headache and pneumonia. As of March 2021, 116 million confirmed cases of COVID-19
have been reported worldwide (https://coronavirus.jhu.edu/map.html). The onset of
COVID-19 leads to respiratory failure that further causes alveolar damage [3].
This novel strain of coronavirus has become a source of augmenting morbidity and
mortality worldwide for the last few months. An extreme inflammatory immune
response is believed to be the major cause of disease severity and death in
COVID-19 patients which involves elevated level of inflammatory cytokines and
chemokines (Interleukin (IL)-1
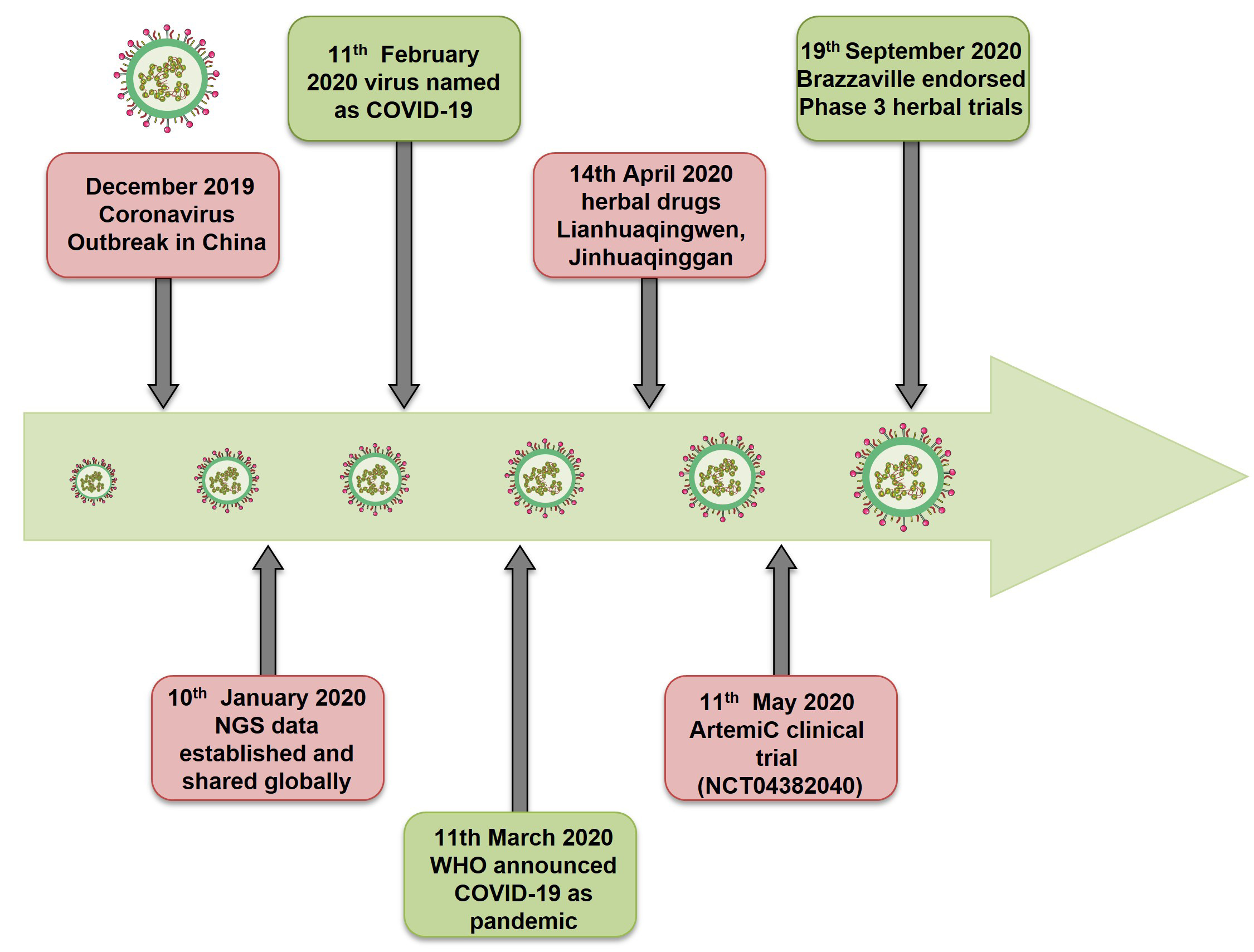 Fig. 1.
Fig. 1.Timeline of COVID-19. Revisiting the events of Coronavirus pandemic and therapeutics.
In the case of viral infections, the most commonly adopted therapeutic strategies include the blockage of the viral entry and replication or modulation of humoral and cellular immunity in the uninfected population. Unfortunately, till date, no FDA approved drug is available that can target or limit the entry and replication of SARS-CoV-2. Till date, the pertinent factor involved in disease severity relates to either activation or suppression of immune responses in the host. Several drugs have been repurposed for the treatment of COVID-19 patients like chloroquine, hydroxychloroquine, remdesivir, favipiravir, ritonavir, lopinavir, ribavirin, dexamethasone, arbidol, anti-inflammatory medicines viz. glucocorticoids, tocilizumab and siltuximab (IL-6 cytokine inhibitors) and anakinra (IL-1 cytokine inhibitor) [12]. Currently, convalescent plasma therapy showed effective results against COVID-19 [13]. But these synthetic drugs and therapies exhibit various adverse effects like heart failure, permanent damage to retina as observed in case of hydroxychloroquine and liver damage in case of remdesivir etc. [14]. Thus, there is an exigent need to identify safe and effective therapies with negligible side effects against COVID-19. Several in silico analysis showed that plant derived natural compounds could be effective therapeutic agents against SARS-CoV-2. Plants produce an unbelievable variety of natural compounds with immense therapeutic potential. However, the limited awareness regarding the mechanism of action of natural compounds is a shortcoming that basically prevents the acceptance of plants as phytotherapy by medical professionals. The Ministry of AYUSH, Government of India has recently recommended various immunity promoting measures based on the Ayurveda (the oldest healing science, 5000 years ago) for the prevention and management of COVID-19. Recently, WHO too had endorsed the use of traditional herbs in tackling COVID-19 induced health complications (https://www.afro.who.int/news/expert-panel-endorses-protocol-COVID-19-herbal-medicine-clinical-trials). In the present review we have discussed some of the most potent phytotherapeutics available from natural traditional medicines that can be employed by clinicians as therapeutic agents to control viral titer as well as toxicity associated with SARS-CoV-2 infection. Also, we offer suggestions to understand the significance of natural herbs and their utilization to enhance anti-viral immunity in the host.
Coronavirus belongs to a large family of enveloped, non-segmented, positive (+) sense single stranded RNA viruses that circulate in animals including bats, cats, dogs, camels etc. These viruses are named as coronaviruses due to the resemblance of their structure to a crown or corona. Coronavirus are known to cause disease in both animals and humans. Among all coronaviruses, four human coronavirus strains (OC43, NL63, HKU1 and 229E) are known to infect upper respiratory tract with minor symptoms [15]. On the other hand, three human coronavirus strains (SARS-CoV, SARS-CoV-2 and MERS) are known to infect lower respiratory tract that can develop pneumonia and lead to deadly condition. SARS-CoV-2 is a beta coronavirus that shares 79% genetic sequence similarity with SARS-CoV and 98% homology with RaTG13 coronaviruses residing in chrysanthemum bats [1]. Moreover, immunopathogenesis of SARS-CoV-2 that leads to airways destruction closely resembles with SARS-CoV [16]. In severe cases of infection with SARS-CoV-2, ARDS condition develops that causes respiratory failure which is found to be the major cause of mortality [17]. In addition to this, mounting of immune responses against viral infection causes release of enhanced levels of inflammatory cytokines from the innate immune cells thereby leading to the development of “Cytokine Storm Syndrome” (CSS) [17]. This condition results in uncontrollable inflammation that further impose multiple-organ failure eventually leading to death. Thus, we can infer from these findings that viral infections are not solely responsible for airways destruction, but the host immune response also plays a vital role in the advancement of disease. Furthermore, severity of the disease is found to be correlated with advanced ageing and with the presence of comorbidities [18].
The first step involved in the pathogenesis of COVID-19 is the binding of virus to the host cells (such as airway epithelial cells, endothelial cells, alveolar macrophages, and alveolar epithelial cells) in the lungs. All these cells express Angiotensin Converting Enzyme 2 (ACE2); the target receptor that makes these cells susceptible to coronavirus infection [5]. After infection, a reduction in the pulmonary ACE2 expression is observed and loss of ACE2 might be responsible for enhanced severity of the disease. Generally, ACE2 is known to regulate the Renin-Angiotensin-System (RAS) which in turn regulates the blood pressure and electrolytes/fluid levels and dysfunction of RAS system has been observed in patients infected with SARS-CoV-2. The corona virion comprises of four proteins (viz. spike-S, envelop-E, membrane-M and nucleocapsid-N) and a single stranded RNA genome having a length of 29,900 nucleotides (Fig. 2). Spike (S) protein of SARS-CoV-2 aids the interaction of the virus with target cells expressing ACE2 on their surface. In addition to ACE2, serine protease TMPRSS2 known to be the priming factor for S protein that facilitates the entry of SARS-CoV-2 into the host cell [19]. However, the reason behind the broad dissemination of SARS-CoV-2 is not reported till date. Recently, it is observed that host protease furin which results in cleavage of full-length S glycoprotein into polybasic S1 and S2 polypeptides resulting in exposure of Arg-Arg-Ala-Arg C-terminal sequence (CendR motif) in S1 polypeptide that aids in binding of virus to host cells via Neuropilin-1 (NRP1) receptor. NRP-1 is expressed on various human tissues such as respiratory tract, neurons and blood vessels. Thus, we can conclude that NRP serves as a second key host receptor that involves in SARS-CoV-2 infection and may provide a therapeutic target for COVID-19 [20].
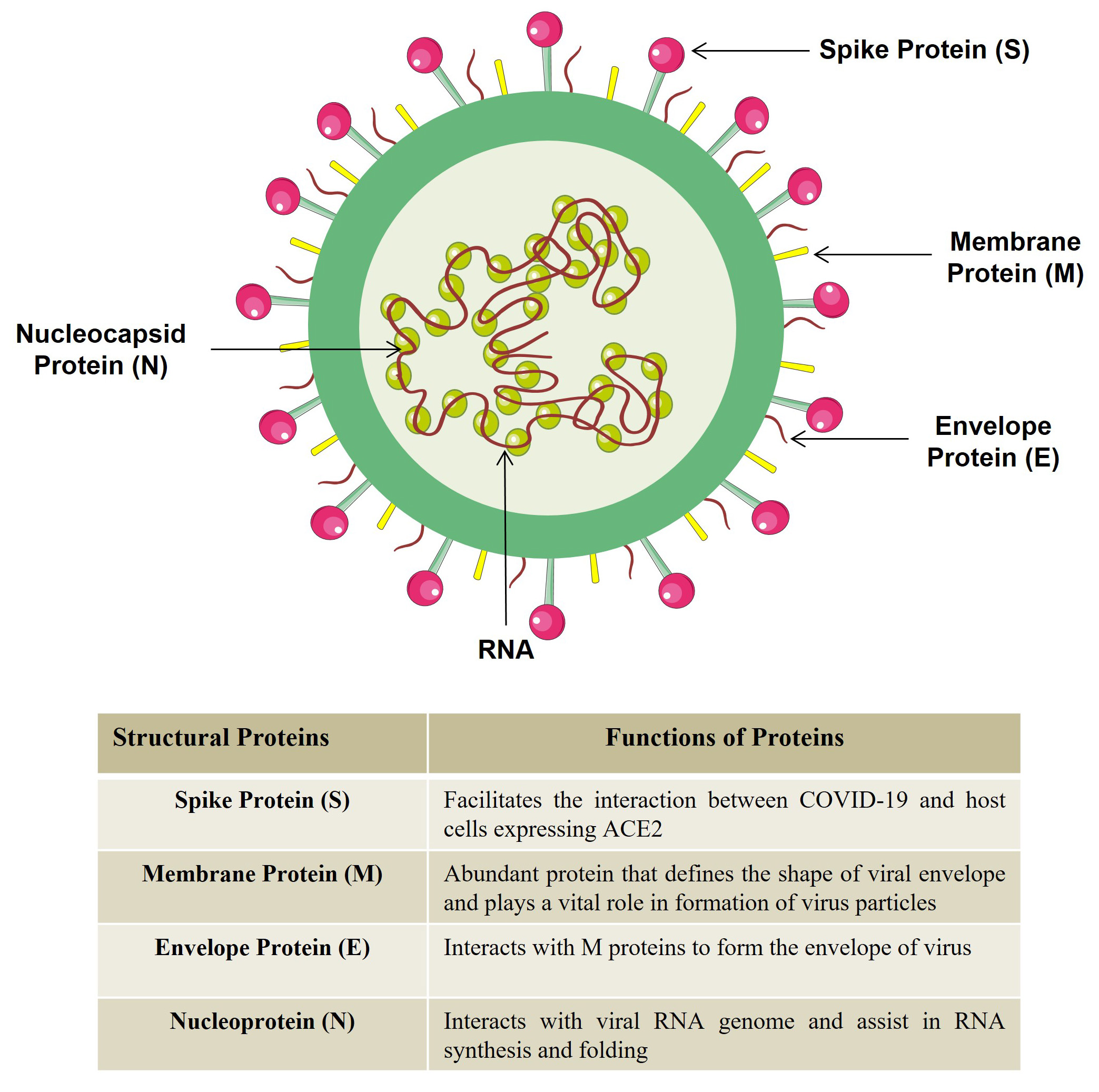 Fig. 2.
Fig. 2.Structure of COVID-19. COVID-19 structure is comprised of four proteins and single stranded RNA genome. (Figure illustrated with the help of https://smart.servier.com/).
Surprisingly, it has also been observed that males are highly susceptible to SARS-CoV-2 infection as compared to females [21]. Further, the sex-hormones like estrogen and testosterone exhibit different immunomodulatory properties [22, 23, 24]. Thus, highlighting the significance of sex steroid hormones for their possible role in disease severity among males in comparison to females. In case of SARS-CoV, a study reported that treatment with estrogen receptor antagonist or estrogen deficiency in ovariectomized female mice dramatically enhanced the morbidity and mortality in female mice due to virus infection [21, 25]. Recently, a study highlighted the association between Kawasaki disease (KD) and COVID-19 in infants [26]. Furthermore, a study revealed that the developing human embryo also expresses ACE2 receptor and TMPRSS2 protease necessary for virus internalization [27]. Thus, these findings abolish the misconception that only the aged population is highly susceptible to SARS-CoV-2 infection.
Viral infections induce the host innate and adaptive immune responses. These
responses are initiated with the activation of the innate immune system that
recognizes various molecular patterns such as pathogen associated molecular
patterns (PAMPs) and death associated molecular patterns (DAMPs) like ATP,
nucleic acids and ASC oligomers. Adaptive immune system is activated with the
induction of T cells and the release of various antigen specific antibodies by
the B cells. Activation of innate and adaptive immune response stimulates the
release of various proinflammatory cytokines and chemokines that further promote
the recruitment of macrophages and neutrophils at the site of infection. These
cells secrete various cytotoxic substances which are required to clear the
infection. These well-coordinated adaptive and innate immune responses generally
eliminate the viral infection but sometimes viral infection results in activation
of intense inflammatory response resulting in excessive destruction of host
tissue. This disproportionate immune response has been hypothesized to be the
reason of immunopathogenesis observed in COVID-19. SARS-CoV-2 infection induced
extreme inflammation causing the production of large amount of pro-inflammatory
cytokines and chemokines. Remarkably, it has been observed that COVID-19 patients
who were admitted to intensive care unit (ICU) had higher levels of inflammatory
cytokines such as IL-2, IL-7, IL-10, IFN-
Moreover, insufficient activation of the anti-viral cytokines viz. type I and type III interferon is found to be the crucial factor contributing to the failure of innate immune response thereby enhancing viral dissemination and persistence. Thus, the identification of molecular mechanisms responsible for reduced expression of anti-viral cytokines would be crucial for the development of directed immunomodulatory strategies for COVID-19 treatment.
Along with the cytokine storm, lymphopenia is one of the critical clinical
manifestations observed in COVID-19 patients [31]. Lymphopenia is defined as the
condition in which a substantial reduction in the lymphocyte counts mainly
CD4
Notably, a study reported that the degree of lymphopenia (lymphocyte count
Traditional medicines have long antiquity in playing a central role in the
prevention as well as in the management of several diseased conditions. They have
a holistic concept that works by strengthening the body resistance to eradicate
pathogenic factors. Recently, the significance of traditional medicines has also
been proven in the management of COVID-19 pandemic. A study reported that early
involvement of traditional medicine improves the recovery rate and reduces the
mortality rate in SARS-CoV-2 infected patients [35]. Traditional medicines not
only work by inhibiting virus replication but also dampen the inflammatory storm
by regulating the immune response [35]. Moreover, various studies have clearly
specified that phytoconstituents exhibit immunomodulatory properties and are
known to augment the expression level of anti-inflammatory cytokines along with
reducing the expression of inflammatory cytokines such as IL-6, IL-17, and
TNF-
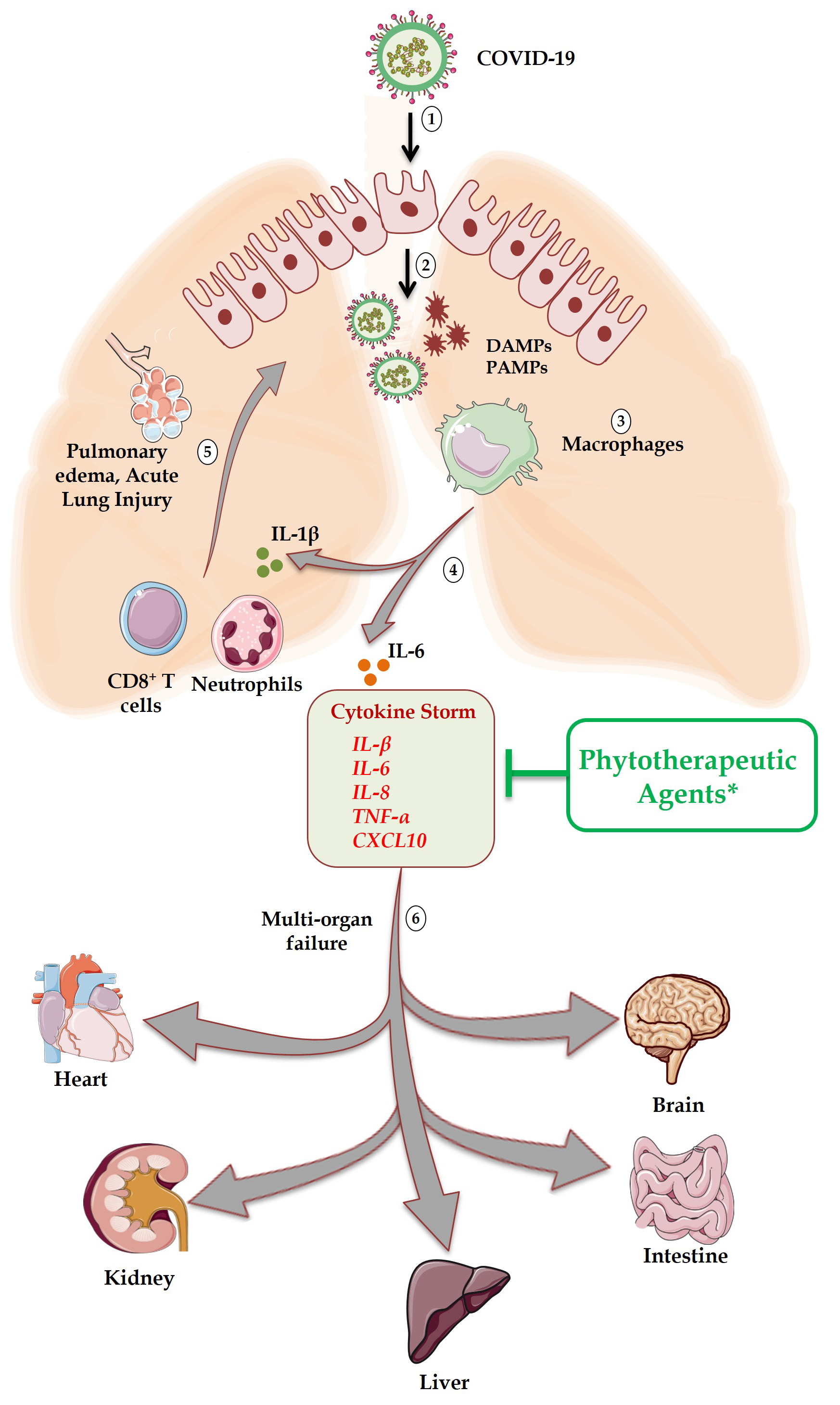 Fig. 3.
Fig. 3.Schematic representation of immune response and associated
tissues toxicity in sequential manner. Step 1: Internalization of COVID-19 into
target cells expressing ACE2 receptor; Step 2: After pyroptosis waning cells
generate death associated molecular patterns (DAMPs) and pathogen associated
molecular patterns (PAMPs); Step 3: Virally derived DAMPs and PAMPs engulfed by
macrophages in turn produce pro-inflammatory cytokines; Step 4: Interleukin
(IL)-1
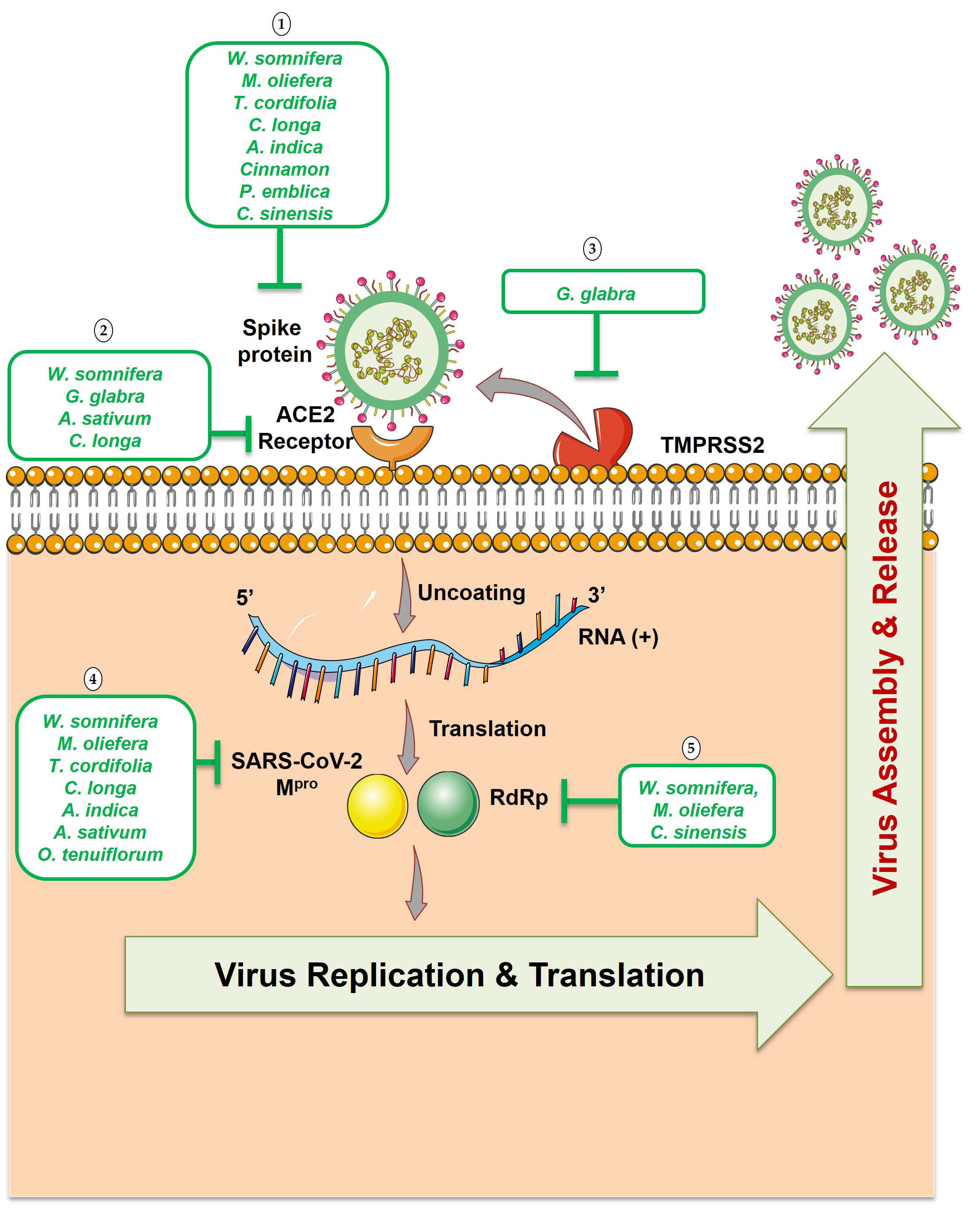 Fig. 4.
Fig. 4.Potential phytotherapeutic approaches against SARS-CoV-2. (1)
Natural compounds that targets spike proteins could block SARS-CoV-2 from
interacting with the ACE2 receptor. (2) Natural compounds that targets ACE2 host
receptor could block SARS-CoV-2 attachment to the cells. (3) Natural compounds
against the serine protease (TMPRSS2) prevent the spike protein cleavage which is
required for the viral fusion to the host cells. (4) Natural compounds that
targets M
| Phytotherapeutic agents | Common names | Family | Biologically active components | Pharmacological activities | Predicted targets in COVID-19 | Tissue protective activities | Ref. |
| Commiphora wightii | Gugal, Guggul, Indian Bdellium | Burseraceae | Guggulsteron, lignans, ketosterols, flavanones, guggulipid | Anti-inflammatory ( |
Cytokine storm (IL-1 |
Neuroprotective, Cardioprotective, Hepatoprotective, Nephroprotective, Lung protective | [37] |
| Salvia officinalis | Sage | Lamiaceae | Alkaloids, phenolic compounds, steroids polyacetylen, essential oils | Anti-inflammatory ( |
Cytokine storm (IL-1 |
Neuroprotective, Cardioprotective, Lung protective | [38] |
| Foeniculum vulgre | Fennel | Apiaceae | Coumarins | Anti-inflammatory ( |
Inflammatory cytokine (IL-17) | Lung protective, Skin protective | [39, 40] |
| Mentha balsamea | Peppermint | Lamiaceae | Ursolic acid (triterpenoid compound), phenolic acids (rosmarinic and caffeic acids), flavones, flavanones | Anti-inflammatory ( |
Cytokine storm (IL-1 |
Skin protective, Improve lung function | [41, 42] |
| Salvia rosmarinus | Rosemary | Lamiaceae | Carnosic acid, Carnosol, | Anti-inflammatory ( |
IL-1 |
Lung protective, Neuroprotective | [43, 44] |
| Echinacea purpurea | Cone flower | Asteraceae | Phylloxanthobilins (tetrapyrrolic compounds), caffeic acids | Anti-inflammatory, Anti-bacterial, Anti-oxidant, Anti-Diabetic, Treat upper respiratory illness, Common cold, Anti-viral, Immunomodulatory properties ( |
Lymphopenia (Increased CD4 |
Lung protective, Hepatoprotective, Neuroprotective | [45, 46, 47, 48, 49] |
| Sambucus nigra | Elderberry | Adoxaceae | Phenolic acids, flavonols, flavonoids, Total phenols | Anti-inflammatory ( |
Cytokine storm (IL-1 |
Lung protective, Hepatoprotective, Nephroprotective | [50, 51, 52] |
| Panax ginseng | Man-root | Araliaceae | Ginsenosides, Panax notoginseng saponin (PNS) | Immunomodulatory properties ( |
Cytokine storm (IL-1 |
Neuroprotective, Myocardial protection, Hepatoprotective, Intestinal protection, Lung protective | [53, 54, 55] |
| Taraxacum officinale | Dandelion | Asteraceae | Polysaccharide, Taraxasterol | Anti-inflammatory ( |
Cytokine storm (IL-1 |
Hepatoprotective, Lung protective, Neuroprotective, Nephroprotective | [56, 57, 58] |
| Tanacetum vulgare | Tansy | Asteraceae | Flavonoids | Anti-inflammatory ( |
Cytokine storm (IL-1 |
Nephroprotective, Neuroprotective | [59, 60, 61] |
| Drug | Composition | Clinical trial phase/type of study/No. of participants | Mechanism of action | Reference identifier |
| ArtemiC (Micellar formulation) | Artemisinin (6 mg), Curcumin (20 mg), Frankincense (15 mg), Vitamin C (60 mg) | Phase II/ To evaluate the safety and efficacy of ArtemiC on patients diagnosed with COVID-19/50 P | Diminish IL-6 and TNF-α levels | NCT04382040 |
| Previfenon® | Epigallocatechin-3-Gallate (EGCG) (250 mg) | Phase II/ To determine the efficacy of Previfenon® (EGCG) to prevent COVID-19/524 P | Exhibits anti-viral chemoprophylaxis of COVID-19 | NCT04446065 |
| Guduchi Ghan Vati | Giloy (500 mg) | NA/ To evaluate the safety and efficacy of Guduchi Ghan Vati for COVID-19 asymptomatic patients/18-75 years age | Immunomodulatory potential | NCT04480398 |
| Gargles (Mouthwash) | Neem | NA/ To reduce intraoral viral load in COVID-19 infected patients/50 P | Anti-viral | NCT04341688 |
| Omega 3 Viruxide | Neem oil & Wort oil | NA/ To study the viruxal oral and nasal spray for treating the symptoms of COVID-19/128 P | Reduce symptoms associated with COVID-19 infection | NCT04357990 |
| Nigella sativa (Black Cumin) | Black seed (500 mg) | Phase II/ To study Nigella sativa As a treatment option for patients having upper respiratory infection caused by SARS-CoV-2/200 P | NA | NCT04401202 |
| Nigella sativa (HNS-COVID-PK) | Cumin seed powder (1 gm) | Phase III / Role of honey and Nigella sativa in the management of COVID-19/ 30 P | NA | NCT04347382 |
| Traditional Chinese medicine | NA | Phase III/ To evaluate the safety and efficacy of TCM as an adjuvant for the patients with SARS-CoV-2 COVID-19/50 P | NA | NCT04323332 |
| Individualized Ayurveda | Ginger/Turmeric/Honey/Lemon | NA/ Ayurveda self-management for flu like symptoms during the COVID-19 outbreak/18-6 years age | —- | NCT04345549 |
| NA* denotes Not Available at clinical trial government site (https://clinicaltrials.gov/). | ||||
Withania somnifera belongs to Solanaceae family and is commonly known as “Indian Ginseng” or “Indian Winter cherry”. It is also known as “Sattvic Kapha Rasayana” that holds a prominent place in Ayurvedic Rasayana herb [62]. It is among the most esteemed herb of the Indian Ayurvedic system which is prominently used as a Rasayana (tonic) with wide range of health benefits. The pharmacologically active components of W. somnifera are alkaloids, steroidal lactones, and saponins [63] that are crucial for imparting these health benefits. Withaferin (WA) is the first withanolides compound that was isolated from the leaves of W. somnifera. Withanolides are a group of 28 carbon-containing steroidal lactones isolated from solanaceous plant. Along with anti-microbial activity, anti-arthritic activity and anti-tumorigenic activity, these withanolides also possess hepato-protective properties [64].
In addition to several beneficial and health promoting effects, W.
Somnifera root extract also possesses anti-viral and immunomodulatory properties
as treatment of PBMCs from HIV patients with W. Somnifera reduced the
expression of CD38 on CD8
Entry of SARS-CoV-2 virus into the host cells cause development of cytokine
storm that involves excessive production of inflammatory cytokines which further
results in pulmonary edema and acute lung injury which may results in shock and
multiple organ failure eventually leading to death in COVID-19 infected patients.
Of note, W. somnifera reported to suppress various pro-inflammatory
cytokines viz. IL-2, IL-6, TNF-
Tinospora cordifolia commonly named as “Guduchi” belongs to Menispermaceae family. Across the globe, this traditional herb has gained researcher’s interest because of its diverse medicinal properties e.g., anti-oxidant, anti-arthritic, anti-diabetic, anti-inflammatory, anti-malarial, anti-stress, anti-allergic and immunomodulatory etc. [80]. The biologically active components of Guduchi responsible for its medicinal properties are alkaloids, aliphatic, glycosides, steroids and diterpenoid lactones [81].
Recently, in silico study screened the anti-viral potential of chemical
constituents of Tinospora cordifolia (e.g., berberine
(C
The anti-inflammatory properties of Tinospora cordifolia extract (TCE)
is mediated via suppression of pro-inflammatory cytokines such as IL-1
Glycyrrhiza glabra belongs to Fabaceae family and is commonly known as Liquorice or Licorice. Since, 1950s one of the triterpene glycosides called Glycyrrhizin (GL) isolated from Glycyrrhiza glabra (root and rhizomes) has been studied widely because of its pharmacological properties such as anti-inflammatory, hepatoprotective, anti-carcinogenic and anti-viral properties.
In addition to various health promoting effects of Glycyrrhiza glabra,
it also possesses anti-viral activities: GC7 (18
Recently, it is proven that GL mediates anti-inflammatory effects by suppressing
the levels of inflammatory cytokines (IL-1
Allium sativum belongs to Liliaceae family and is commonly known as “Garlic” with a well-established historical relevance as a favorable traditional medicine in the ancient years. Notably, garlic has been in use for the treatment of several conditions such as cardiovascular diseases [103], hyperlipidemia [104] and as anti-fungal [105] or anti-diabetic for impediment of nephropathy [106] etc. The biologically active ingredients of garlic that are responsible for its medicinal properties are: sulphur containing compounds like alliin, diallyl-sulfide, diallyl-disulfide, diallyl-trisulfide, S-allylcysteine (SAC), enzymes (alliinase) and the compounds produced from alliin (allicin) [107, 108]. Garlic exhibits potent anti-microbial activity due to the production of allicin in damaged garlic tissues which inhibits bacterial infection in vapor form [109]. It also exhibits anti-bacterial and anti-biofilm potential against antibiotic resistant Shiga-Toxin producing Escherichia coli (STEC) isolates from food sources as well as from patient samples [110]. Since no volatile antibiotic is available till date that can treat pulmonary pathogenic infections, allicin could make a valuable addition to the presently available therapies.
Recently, the anti-viral property of garlic essential oil has been investigated by a group in which they examined the anti-coronavirus activity of garlic oil by molecular docking technique and observed that some biologically active ingredients of garlic such as diallyl-disulfide and diallyl-trisulfide have the potential to suppress ACE2 host receptor and main protease PDB6LU7 of SARS-CoV-2 virus [111]. Thus, the results suggest that garlic essential oil (GEO) may act as a valuable natural anti-viral source that contributes towards the restricted entry of coronavirus into the human body. A study suggested that Allium sativum possess the potential of reducing inflammatory cytokines and adipose tissue derive hormone i.e., leptin having inflammatory nature and may be utilize as a preventative measure in the population before being infected with COVID-19 infection [112]. Organosulfurs and flavonoids are found to be the predominant bioactives behind the immunomodulatory nature of Allium sativum. By forming the hydrogen bonds with the active sites of serine protease these bioactives may inhibit the COVID-19 outbreak [113]. Furthermore, a study demonstrated that alliin, SAC and other bioactive compounds of Allium sativum, can be employed as a potential inhibitor candidate for COVID-19 and could be beneficial in combating this pandemic [114].
Several in vitro and in vivo studies reported that water
fraction of garlic exhibits anti-inflammatory properties by which it ameliorated
the infiltration of inflammatory cells, mucus hypersecretion and lung
based-goblet formation in allergic asthma mouse model. Moreover, water fraction
of garlic enhanced the levels of Th1 cytokines (IL-12 and IFN
Zingiber officinale belongs to Zingiberaceae family which is not only utilized as a spice or flavouring agent in food but also considered as a traditional medicine from ancient times. It is adapted as a traditional herb due to its potential to treat various health problems such as nausea [117], migraine [118], diabetes etc. [119, 120]. The biologically active ingredients that are responsible for its pharmacological properties are paradols, 5-acetoxy-6-gingerol, gingerdiones, gingerdiols, 6-gingerol, 10-gingerol, 12-gingerol, 6-dehydrogingerols, 6-shogaol and 3,5-diacetoxy-6-gingerdioal. Among all the two primary compounds that are found to be associated with medicinal properties are 6-gingerol and 6-shogaol [121]. It also exhibits anti-microbial [122], anti-inflammatory [123], anti-ageing [124], anti-oxidant [125], neuroprotective [126] and anti-carcinogenic [127, 128, 130, 131] properties. Both hyperoxia and inflammatory conditions cause lung injury that further leads to the progression of broncho-pulmonary dysplasia (BPD) condition.
Moreover, it has also been found that ginger essential oil (GEO) constituents also exhibit the virucidal activity [131]. Recently, molecular docking study revealed that rhizome extract of ginger possesses higher affinity for the SARS-CoV-2 papain-like protease (PLpro) which is essential for the survival and replication of SARS-CoV-2 [132]. Recently, in silico study demonstrated that phytochemical compounds of Zingiber officinale showed significant affinity with the S-spike protein of SARS-CoV-2 and ACE2 receptor in the host and may have a great potential in reducing the viral load along with decreasing the shedding of SARS-CoV-2 in the nasal passage [133]. Hence, it may be of interest to consider garlic for anti-viral therapy.
Administration of ginger at a dose 1000 mg/kg abridged the levels of
inflammatory cytokines (IL-1
Curcumin belongs to curcuminoids group that accounts for the majority of fraction of curcuminoids as compared to bisdemethoxycurcumin and demethoxycurcumin. It is also known as diferuloylmethane and found to be the chief polyphenol observed in rhizome of Curcuma longa (Turmeric) and other curcuma species [135]. In the past 50 years, most of the beneficial properties of Curcuma longa are predominantly attributed to curcumin. Curcuma longa has traditionally been used as a medicinal herb for pathological conditions due to its anti-microbial, anti-inflammatory, anti-carcinogenic and anti-oxidant properties.
Curcumin is also known to possess anti-viral properties against dengue virus
(serotype 2) [136], human simplex viruses (HSV) [137], HIV [138], Zika and
chikungunya viruses [139]. Moreover, in vitro and in vivo
studies revealed that treatment with curcumin ameliorated the influenza virus
associated pneumonia by attenuating the lung injury and regulating the levels of
inflammatory cytokines in macrophages [140]. Recently, a study discussed the
potential ways by which curcumin (poly-phenolic compound) can be employed for
treating SARS-CoV-2 infection [141]. Furthermore, docking results suggest that
curcumin exhibits the highest interaction with the spike protein (-141.36
kcal/mol) and with the ACE2 receptor (-142.647 kcal/mole) [142]. In consistence
with this, a study suggested that curcumin also displayed significant binding
with the M
Inflammasomes plays a vital role in maturation and secretion of inflammatory
cytokines (IL-1
Till date different cinnamon species e.g. Cinnamomum verum, Cinnamomum cassia, Cinnamomum burmanii, Cinnamomum zeylanicum, Cinnamomum tamala, Cinnamomum loureirii and Cinnamomum cordatum have been reported that can be used in food industries as food additives. Cinnamon belongs to Lauraceae family and recently, cinnamon has gained tremendous attention due to its medicinal properties. The pharmacologically active chemical components of cinnamon are: cinnamaldehyde, trans-cinnamaldehyde and cinnamic acid. These active components confer various health benefits when administered adequately including antioxidants, anti-microbial, anti-inflammatory, anti-gastric ulcer, anti-yeast etc. [152].
In most of the viral infections including SARS-CoV-2 the expression of HSPA5
protein is upregulated due to ER stress conditions [153]. Interestingly, it has
been reported that SARS-CoV-2 with the aid of spike protein enters into the host
cells by using more than one host cell receptor and among them one of the
receptors is HSPA5 also known as Bip or GRP78 [154]. Under stress conditions,
HSPA5 is exposed on the cell surface that permits pathogen entry into the cell.
Surprisingly, cinnamon administration has been reported to reduce ER stress in
rat obesity mouse model [155] thus it is plausible to suggest that cinnamon
administration may also inhibit the translocation of HSPA5 to the cell membrane
from cytoplasm and reduce the interaction between virus and host cells thereby,
inhibiting virus entry. Recently, it has been demonstrated that cinnamon exhibits
the potential to bind HSPA5 substrate binding domain
The bark of Cinnamomum verum has well established anti-inflammatory
potential and it has been demonstrated that the ethanolic fraction of cinnamon
showed anti-inflammatory properties by inhibiting the production of
IL-1
Moringa oliefera belongs to Moringaceae family and commonly known as drumstick or miracle tree, considered as nature’s healthiest and nutritious plant. Its origin is in the northern side of India and its pharmacological properties lie in the different parts of tree such as leaf, bark, sap, roots and flowers that have been utilized as a traditional herb for years [161]. M. oliefera exhibits ethnomedicinal characteristics, attributed to various biologically active ingredients of the plant such as flavonoids, polyphenol, vitamins, tannins, isothiocyanates and saponins [162, 163].
Moreover, seeds of the M. oliefera have also been utilized as reducing
and stabilizing agents for the biosynthesis of silver nanoparticles (AgNP) to
fight against dengue virus (DEN-2) and its primary vector Aedes aegypti [164]. Further, leaves of M. oliefera tree facilitated the
manufacturing of gold nanoparticles that further aided the cancer cells
recognition and possess anti-proliferative and apoptotic activities [165].
Recently, some evidences suggest that plants not only provide macro and
micro-nutrients as food but also deliver distinct molecules with medicinal
properties such as microRNAs and developed a phenomenon known as “cross-kingdom
regulation” [166]. Thus, it is plausible to suggest that M. oliefera
being nutritious plants follows this cross-kingdom regulation and modulates the
expression of human genes responsible for immunomodulatory properties. Recently,
a study attempted to identify the natural compounds from M. oliefera
that exhibit the potential to inhibit COVID-19 [167]. The molecular docking
results suggested that four compounds such as quercetin, kaempferol, morphine and
pterygospermin displayed higher binding efficacy with M
Several studies demonstrated that different extracts (alcoholic or aqueous) of
M. oliefera leaves possess diverse range of biological activities such
as immunomodulatory, analgesic, radioprotective, tissue protective (liver,
kidney, testes, lungs and heart) and antioxidant properties [162, 170]. In
comparison to moringa seed extract (MSE), extract enriched with isothiocyanate
showed enhanced anti-inflammatory potential under both in vitro and
in vivo conditions making it a promising phytotherapeutic agent [171].
Furthermore, one of the novel polysaccharides (MRP-1) isolated from the roots of
M. oliefera showed enhanced anti-inflammatory properties by suppressing
NO and TNF-
Azadirachta indica belongs to Mahogany family, commonly known as
Muarubaini; and possess the potential to treat 40 different diseases. Due to its
astonishing activities this plant is also known as Heal All, Nature’s Drugstore,
Village Pharmacy and Panacea [173]. United Nations has declared Neem plant as
“Tree of 21
Recently, using computational and experimental method it has been observed that
bioflavonoids of A. indica inhibited Dengue virus type-2 (DENV-2)
infectivity by interacting with NS2B-NS3; DENV-2 serine viral protease. Further,
by using molecular docking, molecular dynamics simulation and binding energy
calculations, it was observed that the compounds derived from neem such as
Nimbolin A, Nimocin and Cycloartanols displayed stable and efficient binding with
the vital regions of E and M proteins required for the assembly of SARS-CoV-2
[176]. Moreover, a study revealed that several compounds present in the leaves of
neem exhibit the potential to suppress the M
Furthermore, a study revealed that treatment with neem leaf extract suppressed
the infiltration of neutrophils and monocytes along with reduction of IL-6, MCP-1
and TNF-
Ocimum tenuiflorum belongs to Lamiaceae family and is commonly known as holy basil or Tulsi. Within Ayurveda this plant is known as “The Queen of Herbs” and “Mother Medicine of Nature”. Various scientific studies till date have examined the medicinal properties of tulsi under both in vitro and in vivo conditions. These studies demonstrated that tulsi has unique blend of actions such as anti-inflammatory, anti-stress, anti-cancer, anti-microbial, anti-oxidant, anti-ulcer, anti-thyroid, adaptogenic, anti-cataract etc. [183].
Furthermore, it has been found that different fractions of O. tenuiflorum (crude extract, polyphenol and terpenoid) exhibited anti-viral properties against avian influenza H9N2 infection [184]. Molecular docking studies have proved that several phytochemicals isolated from O. sanctum showed anti-viral activity by interacting with proteins involved in viral replication and metabolic pathways of H1N1 influenza virus [185]. Anti-viral based cytopathic effects (CPE) studies showed that methanolic extracts of O. sanctum inhibited the cell death of Vero E6 cells infected with DENV-1 serotype [186]. Moreover, molecular modelling studies revealed that natural products derived from O. sanctum leaves possess inhibitory activity against ribonuclease-H and DNA polymerase activity of Reverse Transcriptase (RT) enzyme of HIV [187]. Recently, a study via in silico approach showed that flavonoids and polyphenolic compounds predominantly luteolin-7-O-glucuronide and chlorogenic acid, exhibits the ability to covalently bind to the main protease of SARS-CoV-2 and results in irreversible inhibition of viral enzyme [188]. Moreover, a study suggested that Ocimum sanctum extract bioactive compounds viz. tulsinol showed significant interaction with the ACE2 receptor, main proteases and papain like proteases [189]. Thus, these studies suggests that Ocimum sanctum can be employed as a preventative measure against COVID-19 due to its inherent potential to inhibit virus entry, virus replication etc.
A study revealed that leaf extract of O. tenuiflorum and its phenolic
compound eugenol attenuated the activation of human monocyte THP1 cells
by suppressing the LPS induced IL-6, MIP-1
Camellia sinensis belongs to Theaceae family and is commonly known as green tea. Tea is one of the widely used beverages worldwide and mainly comprises of minerals, vitamins, carbohydrates, polyphenols, caffeine, and theanine [193]. Among other polyphenols, catechins and flavonols are found to be the predominant constituents. Further, catechins include epicatechin (EC), epigallocatechin (EGC), gallocatechin (GC), epicatechin gallate (ECG), gallocatechin gallate (GCG), and epigallocatechin gallate (EGCG). Green tea exhibits various health benefits against different diseases such as diabetes, obesity, cancer, cardiovascular diseases, neurodegenerative disorders etc. [193, 194].
Furthermore, Zhou et al. stated that SARS-CoV-2 infected patients are
prone towards secondary infections that lead to multi-organ failure [3].
Interestingly, a study showed that administration of L-theanine alleviated the
organ damage in sepsis induced liver and kidney injury mice model in a dose
dependent manner [195]. Catechin polyphenol also known to exhibit anti-viral
properties against HIV, influenza virus, herpes simplex virus type-1 and
hepatitis B and C viruses [155, 196]. In order to investigate the anti-viral
potential of Camellia sinensis against coronavirus, a recent study
demonstrated that EGCG displayed higher binding energy (-8.3 kcal/mol) with
M
Remarkably, various animal and human studies showed that EGCG (predominant
catechins); possess anti-inflammatory potential [202, 203]. A study reported that
pre-treatment with theanine (20 mg/kg/day) (bioactive component of green tea
leaves) suppressed the LPS induced secretion of IL-1
Astragalus is an important traditional herb in Chinese medicine. Primarily, it is used as a lung protective tonic that enhances resistance against pulmonary infections. From in vitro studies, it was observed that treatment with Astragalus augmented the cytotoxic T cells function; B cells proliferation, activation and antibody mediated responses [206]. Various studies till date have reported anti-viral properties of Astragalus polysaccharide (APS) against viruses [207, 208].
A study reported that in case of COVID-19, endoplasmic reticulum (ER) stress
plays a crucial role in the upregulation of receptors required for SARS-CoV-2
adherence to the host cells. Remarkably, it has been observed that APS attenuated
porcine circovirus type 2 (PCV2) infections by inhibiting ER stress both
in vitro and in vivo [209] thereby highlighting the
significance of APS consumption in limiting virus entry into the host cells
through the inhibition of ER stress. These studies therefore clearly highlight
the potential of Astragalus in the management of SARS-CoV-2 mediated
pathologies. Recent studies have shown that multiple organ failure that occurs
during COVID-19 infection results from the enhancement in inflammatory cytokine
storm and accumulation of free radicals. A study revealed that Astragalus
membranaceus exhibits anti-inflammatory property and suppressed the activation
of MAPK/NF-
A study showed that treatment with APS suppressed the replication of avian
infectious bronchitis coronavirus in chicken embryo kidney (CEK) cells by
reducing the expression of inflammatory cytokines (IL-1, IL-6, IL-8,
TNF-
Piper longum belongs to Piperaceae family, commonly known as long pepper (pippali). Piperlongumine (PL) an amide compound extracted from the roots of P. longum, is a biologically active component known for its medicinal properties. It possesses various physiological and pharmacological properties such as anti-tumorigenic, anti-inflammatory, anti-depressant, analgesic, sedative etc. [220, 221, 222].
Recently a study demonstrated that piperolactam A from P. longum shows higher affinity for Mpro and ACE2 receptor [223]. It has been observed that by targeting these proteins, piperolactam A blocks the replication and entry of virus into the host cells [223].
Recently, a study revealed that pre-treatment with PL suppressed the OVA induced
airway inflammation in asthma mouse model. PL administration inhibits
inflammatory reaction in both BALF and lung tissue by suppressing the
infiltration of inflammatory cells and T
Phyllanthus emblica or Emblica officinalis belongs to Phyllanthaceae family and is commonly known as Indian gooseberry. The fruits are utilized widely in Ayurvedic medicine as a potent “Rasayana” and also found to be the major component of the Ayurvedic preparation “Chyawanprash” [231, 232]. Increasing evidences suggest that P. emblica extract (PEE) possess lung protective activities.
Recently, a study demonstrated that various chemical constituents of
Phyllanthus emblica such as phyllaemblicin-B, phyllaemblinol and
phyllaemblicin G7 exhibited higher binding affinity against helicase and spike
proteins of SARS-CoV-2 [233]. In case of SARS, MERS and SARS-CoV-2 virus,
3-chymotrypsin like cysteine proteases (3CL
Studies showed that administration of PE extract protected the lung from the
inflammatory injury by altering the IL-1
In the present review, we have summarized the pharmacological potential of
various traditional medicines that can be employed for the management and
treatment of cytokine storm induced in COVID-19. Phytotherapeutic agents play a
vital role in the prevention and treatment of several diseases including viral
diseases. Traditional plants having medicinal properties are used as foremost
resources for alternative medicine including several modern medicines for the
treatment of various human disorders including chronic inflammatory conditions.
The traditional medicines are also found to be effective in treating various
viral infections. The scientific society has already proven the efficacy of
various plants harboring anti-viral potential for such conditions and also
accomplished relevant position for them in the health care system across the
globe for both human and animals. Currently, there are no proven treatment
options available for the deadly acute respiratory distress syndrome caused by
SARS-CoV-2 virus. Various countries including India and China have already
resorted to their respective traditional medicine for the management of COVID-19
pandemic which has been of great help in alleviating various symptoms leading to
reduced mortality associated with the viral infection. In April 2020, China
approved the usage of three traditional herbs (Liahuaqingwen, Jinhuaqinggan and
Xuebijing) in treating mild and severe cases of COVID-19 [237]. However, when we
employ them with antibiotics, anti-virals and immunosuppressive drugs; safety
should be cautiously monitored. Several in silico studies have also
proven the relevance of traditional medicines in inhibiting the viral entry,
virus replication and assembly (Figs. 4,5). A study demonstrated that EGYVIR
(herbal drug comprised of black pepper and curcumin) antagonize the
NF-
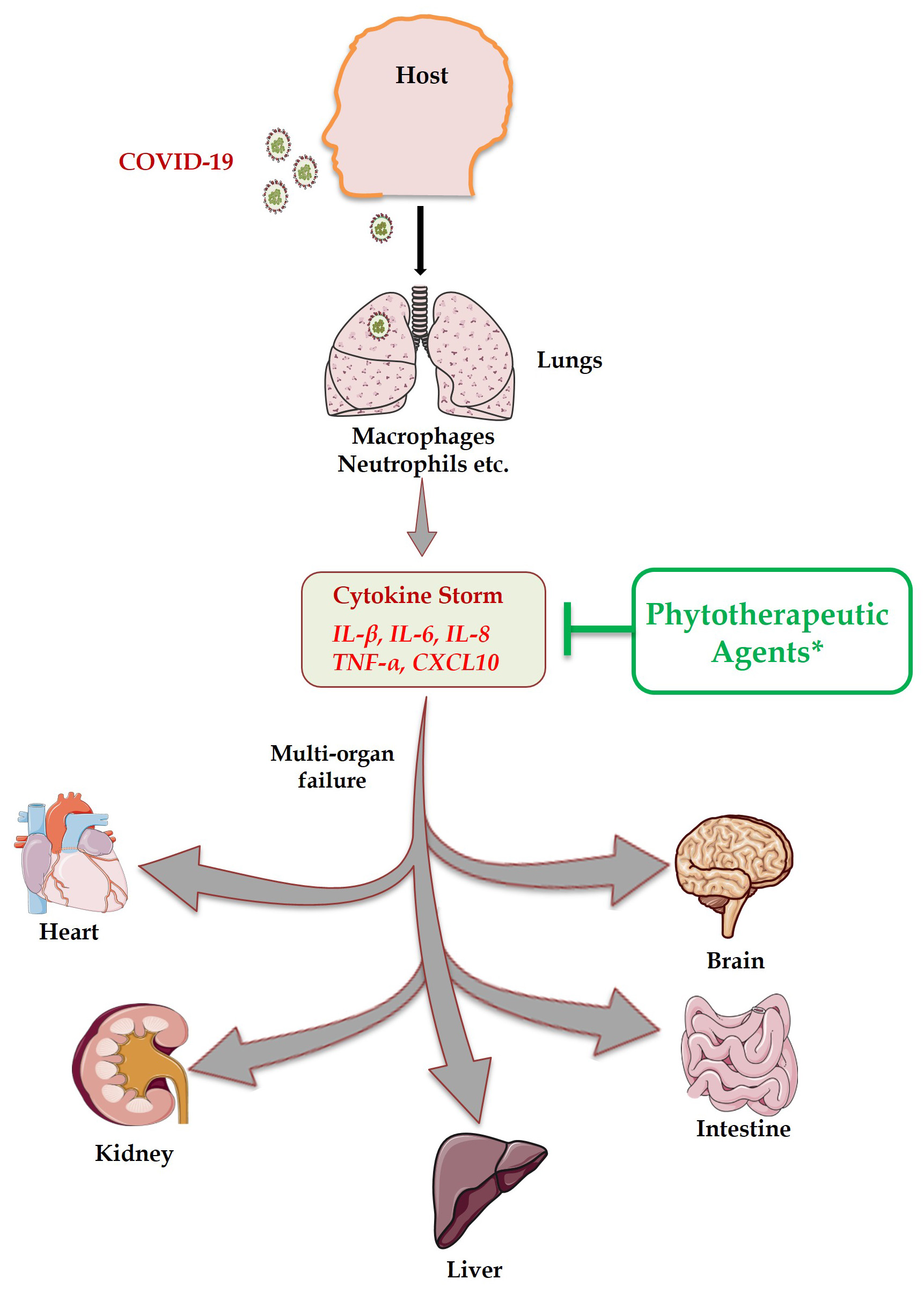 Fig. 5.
Fig. 5.Transmission of COVID-19 occurs mainly from one person to other through droplets during sneezing and coughing, touching contaminated surfaces, personal contacts etc. After entering host, it attacks the lung tissue and results in massive production of inflammatory cytokines (Cytokine Storm Syndrome) that in turn causes multi-organ failure. Phytotherapeutic agents exhibit the potential to suppress inflammatory cytokines along with tissue protective properties.
RKS conceptualized and wrote the manuscript. LS drafted and revised the manuscript. AB, ZA, DM, BV and SR provided valuable suggestions during manuscript preparation and critically revised the manuscript. RKS suggested and LS created the illustrations.
Not Applicable.
LS and ZA thank UGC for research fellowship, AB thank DST SERB project for research fellowship and DM thanks CSIR for research fellowship.
This work was financially supported by projects: DST-SERB (EMR/2016/007158), Govt. of India and intramural project from All India Institute of Medical Sciences (AIIMS) (A-596, AI-15), New Delhi-India sanctioned to RKS. LS, ZA, AB, DM, BV, SR and RKS acknowledge the Department of Biotechnology, AIIMS, New Delhi-India for providing infrastructural facilities.
The authors declare no conflicts of interest.
ACE2, angiotensin converting enzyme 2; ARDS, acute respiratory distress
syndrome; BALF, bronchoalveolar lavage fluid; CD, cluster of differentiation;
COX, cyclooxygenase; CSS, cytokine storm syndrome; ER, endoplasmic reticulum;
IFN, interferon; IL, interleukin; iNOS, induced nitric oxide synthase; MCP-1,
monocyte chemoattractant protein-1; MMP, matrix-metallo protease; NF-