




Frontiers in Bioscience-Landmark (FBL) is published by IMR Press from Volume 26 Issue 5 (2021). Previous articles were published by another publisher on a subscription basis, and they are hosted by IMR Press on imrpress.com as a courtesy and upon agreement with Frontiers in Bioscience.

1 Departamento de Fisiologia, Facultad de Medicina, Universidad Nacional Autonoma de Mexico, Mexico City, Mexico
2 Programa de Doctorado en Ciencias Biomedicas, Universidad Veracruzana, Xalapa, Veracruz, Mexico
3 Licenciatura en Ciencias Genomicas, Universidad Autonoma de la Ciudad de Mexico, Mexico City, Mexico
Abstract
Bisphenol A (BPA), a compound used in the manufacturing of plastics and epoxy resins, is an endocrine disruptor with significant adverse impact on the human's health. Here, we review the animal models and clinical studies as well as the molecular and cellular mechanisms that show that BPA alters the normal function of the reproductive system, metabolism, brain function and behavior and contributes to the development of certain neurodevelopmental disorders including autism spectrum and attention-deficit and hyperactivity disorders. BPA also causes aberrant cognitive function, behavioral disturbances, and neurodegenerative diseases, including Parkinson's disease, amyotrophic lateral sclerosis (ALS), and multiple sclerosis. It has recently been proposed that exposure to BPA may be associated with the development of certain neurodegenerative diseases and neurodevelopmental disorders; however, it is a line of research that is just emerging. This work aims to review the available information about the association between exposure to BPA and cognitive function, behavioral disturbances, neurodegenerative diseases (Parkinson´s Disease, Amyotrophic lateral sclerosis, Multiple Sclerosis), and neurodevelopmental disorders (Autism Spectrum and Attention-Deficit/Hyperactivity Disorders). Likewise, the molecular and cellular mechanisms that may be involved with these pathological conditions will be analyzed.
Keywords
- Bisphenol A
- BPA-A
- Cognition
- Behavior
- Neurological diseases
- Neurodevelopment disorders
- Mechanisms
- Review
In recent years, the importance of studying the mechanisms of action of bisphenol A (BPA; 2,2-bis- (4- hydroxyphenyl) propane), and its relationship with various diseases has been highlighted. BPA is an endocrine disruptor compound (EDC), a chemical that mimics, alters, or blocks endogenous hormonal activity. Exposure to EDCs can modify specific molecular mechanisms or signaling pathways, causing alterations in the structure and function of the Central Nervous System (CNS) (1).
BPA is an industrial chemical compound produced from one molecule of acetone and two molecules of phenol. It was first synthesized by the Russian chemist AP Dianin in 1891 (2). Years later (3), the estrogenic properties of this compound were found when looking for compounds with estrogenic activity without a phenanthrene nucleus.
BPA is a water-soluble-polymer. Therefore, it is stable and inactive, although, when this EDC is subjected to heat, pH changes, and under UV light, its ester bonds hydrolyze. BPA monomers can be dissolved in food and beverages for human and animal consumption, as well as in water, soil, and, to a lesser extent, in the air. Therefore, BPA is potentially a food and environmental contaminant (4). For this reason, it is thought that oral exposure is the primary source of BPA-exposure in both humans, plants, and animals; however, air, soil, and dermal routes can also contribute (5).
It is estimated that 3 million tons of BPA are produced annually and used to manufacture plastics used for industrial, agricultural, and medical purposes. It is also used for epoxy resins that cover canned food and beverages, polycarbonate plastic bottles, food containers, dental sealants, water supply pipes, toys, and cigarette filters (6). After intake through diet, BPA is absorbed from the gastrointestinal tract and transported to the liver, where it is converted into BPA glucuronide, and to a lesser extent, to BPA sulfate, which is eventually eliminated via the renal route (7, 8). Hence, BPA has been detected in human and animal urine and blood, amniotic fluid, fetal serum, and breast milk (9-12). The Environmental Protection Agency (EPA) has estimated that more than one billion pounds of BPA are not filtered into the environment each year, and it is estimated that humans ingest approximately 1–5 μg of BPA/kg a day (13). These data show that humans are widely exposed to BPA.
Numerous studies have reported a variety of effects of BPA in animals, including alterations in the reproductive system such as infertility problems (ovarian cysts, damage to testicular histoarchitecture, prostate cancer), metabolism (obesity and hyperinsulinemia), brain functions and, behavior (14-16). On the other hand, recently, exposure to BPA may be associated with the development of certain neurodegenerative diseases. However, it is an emerging field of research, with many questions still to be resolved.
This work aims to review the available information about the association between exposure to BPA and several pathological conditions within the CNS (neurodevelopmental, memory/learning, behavioral and neurodegenerative disorders). Furthermore, the molecular and cellular mechanisms that may be involved in this association will be analyzed.
As previously reviewed, EDCs, such as BPA, can affect neurodevelopment, causing various effects. Several studies have examined the effects of BPA in vivo or in vitro on the developing and adult brain. To mention some of them, embryonic exposure to BPA has shown to disrupt healthy neocortical development, the cortical organization in adult subjects (17), and altered spatial learning (18). In vitro studies have shown that when BPA is applied in cell cultures, several alterations are provoked. For example, in fetal rat hypothalamic cell cultures, it induces the expression of two proteins related to dendritic and synaptic development (19). Moreover, it decreases cell proliferation and differentiation of hippocampal oligodendrocyte progenitor cells and neural stem cells (20, 21).
Studies performed with experimental animals (most of them utilizing rodents) have shown that exposure to low doses of BPA, which are environmentally relevant during critical periods (such as gestation and lactation), alter sex-specific structural and behavioral patterns. Behavioral disturbances (which can be defined as a disruption of "normal" behavioral patterns) in experimental animals induced by the exposure to this EDC have been amply investigated. The excellent study of Wolstenholme and colleagues (22) revised some of these aspects almost ten years ago. However, we are actualizing this information and considering some other issues that were not revised in literature before. The following sections will review the animal and epidemiological studies that associate BPA exposure with behavioral effects.
Memory and learning are defined as the processes by which an individual acquires the information that encodes, stores, and retrieves. One of the main structures involved in these processes is the hippocampus, which is associated with a wide variety of cognitive functions, such as navigation and spatial planning, coding, and recovery of memory, processing, and novel contexts (23).
It has been reported that murine BPA exposure causes alterations in learning and memory. For example, in the work of Carr et al., oral administration of low (100 mg/kg) and high doses (250 mg/kg) of BPA from postnatal day (PND) 1 through PND 14 disrupted a typical gender-dependent pattern of acquisition (low dose) and retention (high dose) of spatial information (24). Moreover, Xu and collaborators demonstrated that perinatal maternal exposure to BPA at 0.5, 5, and 50 mg/kg/day in ICR mice significantly extended the escape length to find the hidden platform in the Morris water maze. Furthermore, this treatment (at doses of 0.5 or 5 mg/kg/day) markedly decreased the percentage of time spent in the quadrant where the platform had been during training both in PND 21 and PND 56 mice. The results of these authors using the passive avoidance test showed that the error frequency to step down from a platform after receiving footshock was significantly increased, indicating that BPA administration produced an impairment of memory (25). In another study, Tian and colleagues reported that BPA-treated mice (100 and 500 mg/kg/day) showed decreased alternation behavior in the Y-maze at both doses, indicating working memory impairment. BPA-treated mice also showed decreased novel object recognition as expressed by central locomotion and frequency in the central zone (26).
It is worth to mention that BPA effects on learning and memory appear to be sex-dependent. Accordingly, Hass et al., reported that BPA exposure to 25 μg/kg/day in Wistar rats resulted in altered spatial learning in a Morris water maze (18) only in females. In contrast, male but not female deer mice exposed to an elevated dose of BPA demonstrated impairments in spatial learning in the same test (19). However, this latter result opposes the work of Jašarević et al. (20), where they report no significant changes between males and female deer mice in this spatial learning paradigm.
Although most of the available information suggests that BPA exposure induces alterations in memory and learning processes, there have been other studies where no effects have been found (27, 28). As mentioned earlier, the effects of BPA exposure on memory and learning have been amply studied in different behavioral paradigms; therefore, results vary from one study to another. Differences in the reported outcomes are due, to no small extent, to distinct methodologies utilized, which include: the life stage of exposure, the dose, frequency and duration of treatment, the species, strains and sex of the animals used and the route of administration. However, at the neuronal level, some of the effects of perinatal exposure to BPA have been amply described. The results are diverse, and some of the main findings will be described below. In murine models, BPA provoked a decreased signaling of the estrogen receptor (ER) α in the hippocampus of males and increased methylation of ERα gene in the hippocampus of both males and females (29). It has also been observed an upregulation of the mRNA of thyroid hormone (TH) transporter in prefrontal cortex and hippocampus (30); a decreased glucose metabolism (30); changes in the hippocampal metabotropic glutamate 2/3 receptors (mGlu2/3) (31); a downregulation of excitatory protein PSD95 and synaptophysin (a presynaptic density marker), and upregulation of inhibitory protein gephyrin in the cerebral cortex and hippocampus (32).
In a study in which the rats were exposed to BPA during the juvenile stage, it was reported that male animals exposed to a high dose of this EDC showed deficient spatial memory and anxiety. Such deficiency may be related to a downregulation of N-Methyl-D-aspartic acid (NMDA), subunit NR2, and α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) GluR1 subunit, and a decreased spine density of pyramidal neurons in the hippocampus (33). Furthermore, perinatal exposure to BPA (0.5, 5, and 50 mg/kg/day) significantly inhibited the expressions of N-methyl-D-aspartate receptor (NMDAR) subunits NR1, NR2A, and 2B in the hippocampus during the development stage, especially in PND 56 mice (25).
All of the BPA-induced molecular changes mentioned above may partially explain the observed deleterious effects in memory and learning. This idea arises from the fact that all of these cognitive functions are related to brain plasticity. Therefore, whether the expression or function of brain plasticity-related receptors and proteins is diminished as seen with the BPA exposure- the processes of long-term potentiation, synaptic plasticity, and the codification of the diverse memory types, such as spatial are negatively affected remains to be studied.
It has been demonstrated that in male and female rats exposed to this EDC, there are increases in anxiety and depressive-like behaviors. For example, in the predator odor stress paradigm, the subjects exposed to BPA showed an exacerbated avoidance behavior to an aversive stimulus (fox odor), suggesting enhanced levels of anxiety in these animals compared to vehicle-treated subjects (34). Moreover, in pubertal rats that received 40 mg/kg/day of BPA during a week, males but not females exhibited an anxiogenic-like profile (a reduction in the number of entries to the open arms of an elevated plus maze (EPM) (35), one of the most common paradigms used to study anxiety-like behaviors). This effect was also found when administering low doses of BPA (2 mg/kg/day) in gestating female rats until weaning. When their progeny became adults, they were evaluated and showed increased anxiety (36) and depressive-like behaviors (36–38). Curiously, this behavioral pattern was different depending on sex: females were more anxious than males, and males were more depressive than females, as compared with controls (rats not exposed to BPA) (37, 39). Anxiogenic-like effects have also been reported in some studies utilizing the mice version of the EPM (28).
Prenatal and postnatal treatment with BPA has also shown to impact depressive-like behaviors. An increase in immobility time (considered as a depressive-like index) has been observed by several groups utilizing mice (40–42) and rats (38, 42). Other authors have reported no changes in this behavior but have demonstrated that BPA administration eliminated sex differences found in vehicle-treated rats (43).
Controversial data have also been published by several authors that report anxiolytic (26) or lack of BPA effects on anxiety and depressive-like behaviors (37, 44, 45). These discrepancies, again, can be explained by methodological differences between study groups.
Although most studies regarding BPA exposure on anxiety and depressive-like behaviors have been performed using the maternal exposure model, the effects of paternal exposure to BPA have been recently explored (46). For this purpose, it was evaluated the behavior of progeny of male rats that were administered with 50 mg/kg/day BPA for 21 weeks and then mated with females that were not exposed to BPA. This study's results showed that BPA administration produced an anxiogenic profile in the progenitors (F0) and females within the progeny (F1). Moreover, animals of both sexes showed an increase in depressive-like behaviors. These data emphasize that paternal exposure to a low dose of BPA induces transgenerational sex-specific behavioral impairments (46).
It has also been demonstrated that the administration of BPA alters sociosexual behaviors during gestation or lactation. Accordingly, Farabollini and his group studied aggressive behaviors in the resident-intruder paradigm with adult female and male rats whose mothers were administered with BPA (40 mg/kg/day) during pregnancy or lactation. This study revealed an increase in males' defensive behavior, but not in females, with prenatal exposure to BPA (47). In another study, it was observed that aggression scores increased significantly at eight weeks of age in male mice exposed to BPA (2 or 20 ng/gram) compared to a control group, but no differences were found after 12 weeks. Thus, it can be assumed that BPA effects on aggression are transitory (48). These effects are in line with clinical data, where increased aggression has been reported in children (see below).
Motor hyperactivity is another behavioral disturbance reported by BPA administration during both, dark and light phases in rats (46-48). Interestingly, this effect appears to be chronic, considering that it is present in the newborn (47) and adult rodents (of 3, 6, and 9 months) after perinatal exposure through maternal diet (46). These alterations might be related to an abnormal neuronal distribution and layer formation, hypoplasia of layer 6b, and impaired dopaminergic neuronal projections in the neocortex (48) and might be related to pervasive developmental disorders, such as autism, and attention-deficit hyperactivity disorder (ADHD) (see below) (46).
Playing behavior is affected by the alteration of the hormonal environment during development (48); thus, it is a feature susceptible to be affected by BPA exposure. Farabollini and colleagues (2002) observed a masculinization of female behavior in two behavioral categories (play with females and sociosexual exploration), an effect that they suggested it is probably mediated by the estrogenic activity of BPA in the CNS (47). In another study, Porrini and her group utilized dams exposed to 40 mg/kg/day BPA during pregnancy and lactation, and their offspring were observed in a heterosexual social situation from 35 to 54 days of age. Their results show that the early administration of BPA was responsible for an increase of exploration (including social investigation), as well as a decrease of play with males and social grooming at 45 days of age. These observations indicate a general decrease in playful interactions. Together, these results suggest that BPA defeminize some aspects of female's playing behavior (48).
In mammals, maternal behavior is the result of a complex interaction between the lactating dam and her developing offspring. Slight perturbations in the mother-infant interaction may provoke alterations in the mother and the offspring (49, 50). Maternal behavior has been reported to be affected by BPA exposure, too (49). In the study of Palanza et al., they reported that maternal behavior of mice dams exposed to BPA (10 mg/kg) during gestation (days 14-18) was reduced; i.e., dams spent less time nursing their pups and more time out of the nest compared to the control group (50). Some other features of maternal behavior that are altered because of BPA treatment are increased time of the dams resting during pregnancy, an increase in the percentage of time dams resting alone, and reduced time of nest building (49). However, no significant differences in the number of live F2 pups/litter, sex ratio, or body weight at birth, or weight gain during the lactation period have been found between BPA or vehicle-treated offspring. The effects of BPA on maternal behavior have been attributed to estrogen-modulated differences in oxytocin (OT) receptors levels in specific brain areas (see below) and to long-term effects on offspring endocrine and behavioral profile in adulthood (49). Moreover, it has been suggested that these behavioral alterations might be due to an interference in the organization of the neuroendocrine substrates underlying the expression of maternal behavior later in life (49).
It is worth mentioning that BPA exposure also affects the development and functioning of reproductive organs and sexual behavior. It has been demonstrated that, in male mice, BPA administration during fetal development produces a reduction in testis relative weight (48) and sperm count (18). Moreover, plasmatic luteinizing hormone and testosterone are reduced (51) in males exposed to BPA. These features are closely related to fertility and the expression of sexual behavior in male rodents.
Considering that sexual behavior is usually displayed when the hypothalamus-hypophysis-gonadal axis functions adequately, it can be assumed that BPA interferes with the development and functioning of gonads or in this axis, and these alterations can be manifested in reproductive and sexual behavior disturbances. For example, BPA exposure produces a slight modification in male's sexual behavior temporal pattern, showing a discrete reduction in the frequency and latency to intromissions (47, 52). Nevertheless, this latter effect appears to depend on the doses of BPA administered because it occurs only with low doses (52).
Likewise, BPA neonatal administration in females causes an increase in body weight and hyperinsulinemia in adult animals (53–55). This condition can be associated with sexual and reproductive dysfunctions in females (56–58). Interestingly, it has been documented that BPA exposure induces premature puberty, ovarian malformations (for example, an increased number of atretic follicles and a reduced number of corpora lutea), and early reproductive senescence (54, 55, 59). Thus, these characteristics interfere with regular ovulation and fertility in female rodents. Moreover, it has been reported that BPA has effects on female's sexual behavior. Inline, Farabollini and colleagues reported a small increase in sexual motivation and lordosis (a behavioral feature in females denoting sexual receptivity) as a consequence of BPA administration (47). However, this latter result contrasts with other studies that have observed a reduction (60) or lack of effects (54, 55, 61–63) on sexual receptivity.
The finding that BPA exposure changes sociosexual interactions in infant and juvenile non-human primates is of particular interest because of their similarities with humans. Nagakami and colleagues (64) used Cynomolgus Macaque (Macaca fascicularis) to assess the influence of prenatal exposure to BPA (10 mg/kg/day) administered subcutaneously with implanted pumps and examined social behaviors between infants and their mothers during the suckling period. Their results demonstrated that exposure to BPA altered some maternal behaviors, mainly in male-nursing mothers. Furthermore, it affected the behaviors of male infants towards the mother (male infants behaved as females) and modified sociosexual interactions in male juvenile primates by reducing the environmental exploration and the gestures related to sexual reproduction. These features indicated demasculinizing key sexually dimorphic behaviors (65).
The effects of BPA exposure on the human brain and behavior is a relatively new field, and particular concerns have been raised about its potential impact on children. Nowadays, there are eleven articles related to this issue and have been systematically reviewed by Ejaredar and his group, and Mustieles and colleagues (66, 67). Epidemiological studies have shown that prenatal exposure to maternal BPA concentrations is related to higher levels of anxiety, depression, aggression, hyperactivity, conduct problems, inattention, and other cognitive function impairment in children (66). Some of these reports have shown that after sex-stratified analyses, the behavioral outcomes between boys and girls are different. For example, Braun et al. (2009 and 2011) published evidence of a significant association with BPA gestational exposure with externalizing behavior, increased hyperactivity, anxiety, depression, and more inadequate emotional control and inhibition only in girls (68). However, there are four articles in which, regardless of sex, no significant association was found between prenatal BPA exposure and neurodevelopment or behavioral outcomes (69–72).
Another essential factor that influences the results reported in clinical studies is the life stage of exposure to BPA (gestational/prenatal vs. childhood). Most of the studies have shown a stronger relationship between prenatal exposure of BPA than during childhood. The fetal period (especially between weeks 8 and 24 of gestation in humans) is a crucial stage in sexual differentiation of the CNS and neurodevelopment, and steroid hormones are vital factors in this process. The impact that EDC's exposure on human health appears to depend strongly on its timing and the effects of BPA exposure in utero are distinct from those of exposure during adulthood, in rats, non-human primates, and apparently, in humans too (62).
The controversial results found in epidemiological studies can be explained on the bases of many disparities in the study design, the approximation to assess exposure to BPA (proxy vs. direct measurements), the behavioral evaluation (distinct tests used, evaluation periods, and reporters), the sample size, adjustment for potential confounders, and socio-demographic characteristics of the populations. It is important to note that most of the existing epidemiological studies (10 out of 11) were conducted in the United States and only one in Korea (73) and that the results obtained cannot be compared due to their marked heterogeneity, as mentioned before. These aspects limit the possibility to make conclusions that can be drawn on the epidemiology of the potential health risks of BPA exposure on neurodevelopment. Thus, further investigation in this field is warranted (66, 67).
The evidence here presented indicate that BPA exposure exerts significant physiological and behavioral disturbances even in shallow doses, especially in critical periods of development, such as gestation and lactation thus, as it has been pointed out by other authors that it is necessary to be vigilant towards the potential adverse effects of BPA exposure and to take actions to inform women planning or undergoing pregnancy about measures to reduce or avoid exposure to BPA (4, 67).
Neurodegeneration is a pathological condition characterized by progressive damage of neurons, which includes the loss of structure or function, and even death of these cells (74). Among the most common neurodegenerative diseases studied are Parkinson's Disease (PD) and amyotrophic lateral sclerosis (ALS). Although multiple sclerosis (MS) has been classically classified as an autoimmune disease in which myelin sheaths are damaged, it has also been accepted as a neurodegenerative disease (75). In this section, we will explore the available evidence that shows an association between the exposure to BPA and the neurodegenerative diseases here mentioned.
PD is the second-most common neurodegenerative disorder, and its prevalence in adults with ≥65 years of age is around 2–3% (76). The hallmark of this disease is the loss of 50 to 70% of dopaminergic neurons located in the substantia nigra (SN) within the midbrain, and widespread intracellular protein (α-synuclein, a pathological marker of neurodegeneration, particularly in PD) accumulation, which leads to the onset of prominent motor symptoms (bradykinesia) that characterize this disease, and that is why we include it in the present study. Since BPA can bind to ER and estrogens increase dopamine (DA) release from GABAergic neurons (77), the potential indirect effects of BPA in the development of PD are of particular interest.
One of the first evidence that BPA may induce the development of PD was reported by Ishido et al. (78); they found that postnatal BPA exposure caused decreased tyrosine hydroxylase immune reactivity in the midbrain of 7-week old rats and downregulation of the gene expression of dopamine transporter (DAT) of adult rats. Furthermore, these transporters are active, as demonstrated by Alyea and Watson (79), where BPA-exposed PC12 rat pheochromocytoma cells expressing membrane DAT showed an increased DA efflux, which is induced in a non-genomic manner through guanine nucleotide-binding protein and N-type calcium channels (80). The increased function of DAT has caused particular interest among some research groups since this parameter can be used in the early diagnosis of PD (81).
On the other hand, it has been reported that 5-days-old rats exposed to BPA presented an increased spontaneous motor activity at eight weeks of age (82). This might be related to the accumulation of α-synuclein in SN. Another observation obtained from this study was that adult rats that received microinjection of BPA showed degeneration of striatal dopaminergic neurons (82). Also, the group of Nakamura et al. has vigorously studied the effects of maternal BPA exposure in neurodevelopment; in that sense, they have reported that prenatal exposure to BPA affected fetal murine neocortical development by accelerating neuronal differentiation and migration (17). They have also demonstrated that the prenatal and lactational exposure of murine to BPA, increases in adult mice the levels of DA and its metabolite 3,4-Dihydroxyphenylacetic acid in different brain regions, including SN (83). The increased DA levels represent an impairment in the regulation of this neurotransmitter; accordingly, some of its metabolites accumulate and may exert some neurotoxic and cytotoxic effects within the dopaminergic neurons, and mediate mitochondrial dysfunction that may lead to α-synuclein aberrant accumulation in the SN (84).
Altogether, this evidence shows that BPA modifies brain dopaminergic pathways and, therefore, it can be associated with PD. So far, few groups have explored the molecular mechanisms underlying the modulation of dopaminergic pathways by BPA. However, in an in vitro model of maternal exposure to BPA using embryonic bodies derived from human embryonic stem cells, it was determined that BPA induces the decrease of DA production, therefore, increasing the risks of fetal origin of PD through downregulation of the expression of fetal insulin growth factor-1 (IGF-1) (85). Thus, some studies have shown that IGF-1 is essential for dopaminergic neurons since it may protect these cells from oxidative stress (OS) (86) and modulates DA synthesis and DA neuron firing (87). Many other molecules might be associated with the mechanisms that underlie the BPA effects in the dopaminergic pathways; therefore, this is a wide-open field that should be explored.
There has not been extensive exploration regarding the association of exposure to BPA and PD in humans. Although, there is an epidemiological study conducted by Landolfi et al. (88) in which they quantified blood concentration of free and conjugated BPA (BPA conjugated to glucuronic acid) of patients with PD, and their spouses were used as controls. Both populations were homogenous in terms of BPA exposure; however, patients with PD showed significantly lower levels of conjugated BPA compared to controls (88). These results suggest that in PD, there is a diminished BPA metabolism. Although it is important to mention some limitations of this study. 1) The controls of this research work were the patients` wives. However, it was not considered whether sex might be a factor that influences the ability to metabolize BPA. 2) Probably, some drugs used in the treatment of PD (such as rasagiline, entacapone, and benserazide), could affect the efficiency of enzymes of the glucuronidation system that participate in the degradation of BPA. In later studies, PD patients free of the aforementioned anti-parkinsonian drugs should be included as controls (88). Finally, further studies must be done to understand the underlying mechanisms and the toxic effects of BPA that can favor PD development.
ALS is a neurodegenerative disease characterized by progressive degeneration of motor neurons in the cortex, brainstem, and spinal cord (89). Its name comes from "amyotrophic" that refers to muscular atrophy, and "lateral sclerosis" that is related to the scarred aspect of the spinal cord (90). The major clinical manifestations in ALS are cognitive and behavioral impairment, dysphagia, dysarthria, respiratory insufficiency, muscle cramps, weakness, and atrophy (89). Mutations in several genes have been associated with ALS' pathophysiology, as it has been demonstrated to exert neuronal motor injury. For this reason, most of the animal models of this disease have focused on genetic mutations, which are related to the familial form of ALS. However, some studies have suggested that there are some environmental risk factors (neurotoxins, heavy metals, pesticides, and others) that are associated with sporadic ALS cases (91).
Recently, Morrice et al. (92) developed a model of environmentally induced motor neuron degeneration in zebrafish based on exposure to BPA. In this study, it was observed that the zebrafish embryos exposed to BPA showed motor function impairment, reduced motor axon length and branching (markers of motor neurodegeneration in these animals), decreased neuromuscular junction integrity, motor neuron cell death and activation of microglia. These results are essential in the study of ALS pathophysiology as they reinforce the proposal that axonopathy precedes motor cell death in this disease (92). It is essential to highlight that this animal model has two relevant limitations: 1) BPA exposure does not cause motor neuron-specific cell death, and 2) the BPA doses that lead to motor neuron death are over the environmental exposure doses; that is, this model does not reflect the ability of BPA to induce the development of ALS. Although, this is a valuable study, as it can examine the microenvironment during motor neuron degeneration, which may have implications on early phase treatments of motor neuron diseases such as ALS (92).
Multiple sclerosis (MS) is a "chronic, inflammatory, demyelinating and neurodegenerative disease of the CNS" (93). MS is characterized by the destruction of myelin sheaths, axonal damage, and formation of a glial scar in the white and grey matter of the brain and spinal cord, which lead to the clinical features of MS: paralysis, muscle spasms, optic neuritis, and neuropathic pain (93). MS is caused by complex gene-environment interactions; in that sense, some environmental pollutants, such as BPA, could be implicated in the dysimmunity of MS.
To date, there is only one published study that associates exposure to BPA with the development of an autoimmune response similar to what occurs in MS (94). In this work, the authors performed an MS experimental model of autoimmune encephalomyelitis (EAE) in mice exposed to BPA. Their results showed that gestational exposure to this EDC increased susceptibility of adult male mice to EAE when a suboptimal disease-inducing immunization was used. It was observed that stimulated peritoneal macrophages from male mice gestationally exposed to BPA have an altered expression of cytokines and chemokines. For instance, these macrophages produced significantly more TNF-α and granulocyte-colony stimulating factor (G-CSF).
Moreover, when a post suboptimal immunization was performed, adult male mice exposed to BPA showed an altered plasmatic G-CSF and neutrophilic immunity which could be the basis for the development of EAE, as they demonstrated that neutralizing G-CSF in adult male mice mitigates EAE susceptibility caused by gestational BPA exposure (94). Some other evidence support this hypothesis. In mice, it has been shown that G-CSF is critical for the mobilization and maturation of neutrophils (95), and these cells are important for EAE development. Likewise, it has been suggested that neutrophils play an essential role in the development of MS, as it has been established a correlation between neutrophils cytokines and indices of brain lesion formation (96).
This study is the precursor of a new research line, which should be oriented to investigate the molecular mechanisms underlying the association between gestational exposure to BPA and the appearance of MS, as well as those that cause sex-related differences.
"Neurodevelopmental disorders (NDD) are highly prevalent and severely debilitating brain illnesses caused by aberrant brain growth and development" (97). They cause cognitive, social, motor, language, and affective disabilities. The most common NDD include autism spectrum disorder (ASD) and attention deficit hyperactivity disorder (ADHD) (97). The pathophysiology of these diseases is mainly characterized by deleterious effects on neurogenesis, glial and neuronal proliferation and migration, synapse formation and myelination, and neural development (97). NDD is complex and, therefore, their pathogenesis involves various genetic, epigenetic, and environmental factors. It has been reported that the development of the human brain is easily affected by the third trimester to at least 2–3 years after birth (98). During that period, any developmental neurotoxicant exposure can lead to NDD in humans (98). Accordingly, there are numbers of evidence that associate exposure to BPA with NDD, which we will explore in the next section.
"Autism spectrum disorder (ASD) is a set of NDD characterized by a deficit in social behaviors and non-verbal interactions such as reduced eye contact, facial expression, and body gestures in the first three years of life" (99). It is a multifactorial disorder resulting from genetic and non-genetic risk factors and their interaction. In that sense, specific gene mutations interfere with synaptogenesis and axonal motility. Neuroimaging studies have shown that patients with ASD show pathological changes in distinct brain structures, such as the amygdala and nucleus accumbens (99).
It has been reported that the prevalence of ASD is around 1 to 2% in infants up to 12 years of age. However, the number of cases has been increasing over the last two decades, and environmental factors are suspected to be involved in its pathogenesis (100).
Recently, some authors have established an association between prenatal exposure to BPA and ASD-related changes. In two different studies it was found that gestational exposure to BPA increases the externalizing scores (based on the Behavioral Assessment System for Children 2) in 2-years (101) and 7-years-old (102) girls, and internalizing problems in boys, including anxiety and depression, at age 7 (102). More recent studies found that children with ASD had higher serum (103) and urinary concentrations of BPA and its metabolite (104) compared with age-matched healthy controls.
Other groups have endeavored to elucidate the cellular and molecular mechanisms underlying the association mentioned above. One of the first studies to explore these mechanisms was conducted by Kaur et al. (105). This group reported that lymphoblastoid cells from subjects with ASD and age-matched unaffected sibling controls exposed to BPA showed increased levels of reactive oxygen species (ROS) and a decreased mitochondrial membrane potential (106). Although, in a further subdivision of the control group (unaffected non-twin siblings and twin siblings), it was observed significantly higher ROS levels in the autism vs. the unaffected non-twin siblings' group. Moreover, Metwally and collaborators (106) evaluated the effect of BPA on OS in children with ASD, compared to a control group of sex-matched children. This study's results showed that serum levels of BPA and 8-hydroxy-2'-deoxyguanosine (8-oxodG, an OS marker) were significantly higher in children with autism than in control children. Besides, positive correlations were found between both BPA and 8-oxodG with ASD severity (107). These studies suggest that BPA induces OS that may result in mitochondrial dysfunction, which is a risk factor to develop ASD behavior (107). One of the main limitations of this study is that it only evaluated the levels of BPA and OS at the peripheral level. However, it is probable that the ROS -derived from the state of the systemic OS- can enter the CNS and affect brain areas related to speech processing, memory, social interaction, and sensory and motor coordination, as it has already been observed in some studies (107).
Another intriguing question that has arisen in this field is whether BPA dysregulates ASD-related genes in the developing brain. To answer that, Thongkorn et al. (108) made a "transcriptome profiling of hippocampi isolated from neonatal pups prenatally exposed to BPA and revealed a list of differentially expressed genes (DEGs) associated with ASD." Further analysis showed significant associations between the DEGs in males (stronger than in females) and neurological function disorders associated with ASD (108). This work suggests that BPA is associated with dysregulation of genes related to the development of ASD. However, this gene evaluation should be done in other brain sites related to specific symptoms of ASD (such as the frontal lobe, middle frontal gyrus, dorsal prefrontal cortex, and orbitofrontal cortex), in different stages of the neurodevelopment. Moreover, explore if there are sex-related differences, that could partially explain the higher incidence of ASD in males (109).
Attention-deficit/hyperactivity disorder (ADHD) is an NDD that affects 5% of children and adolescents, and 2.5% of adults worldwide (110). It is characterized by impulsivity, hyperactivity, and inattention. ADHD is multifactorial; hence, genetic and environmental factors play a role in its etiology (111). ADHD has a heterogeneous profile of psychopathology, neurocognitive deficits, and abnormalities in the anatomy and function of frontal–subcortical–cerebellar pathways that control attention, response to reward, salience thresholds, inhibitory control and motor behavior (110).
As mentioned above, exposure to toxic substances during the perinatal stage is injurious for neurodevelopment. Accordingly, BPA has been associated with the development of ADHD. However, direct clinical evidence is scarce. So far, there are only two cohort studies that evaluated this association; the first one was performed in a US population of children aged 7-15 years (112), while the second one corresponded to children aged 6-12 years from Guangzhou, China (113). Additionally, it was reported that urinary 8-OhdG concentrations were significantly higher in ADHD children than in the control group (113), which reflects an increase in OS within the ADHD population. In addition, it has been reported that children who received food based on milk formula, are more likely to develop ADHD than those who were breastfeeding and that this is mainly due to the content of BPA present in these formulas (114).
Regarding the mechanisms related to prenatal exposure to BPA and the occurrence of ADHD, as mentioned earlier, it has been suggested that the amygdala is one of the brain structures that are affected by this substance. Inline, a study examined the synaptic properties of the GABAergic/dopaminergic systems in the basolateral amygdala (BLA) of 28-days-old rats, perinatally exposed to BPA (115). The results derived from this study showed that the perinatal exposure to BPA causes disinhibition of the BLA GABAergic system and enhances the dopaminergic system in BPA-exposed rats. These changes alter the synaptic transmission and plasticity of the cortical BLA, which may be associated with the appearance of behavioral features similar to those present in human ADHD (115). These changes are likely due to alterations in the expression of specific synaptic receptors and transporters. This idea is supported by a previous study in which 5-days-old mice were intracisternal injected with BPA, and further evaluated for spontaneous motor activity and gene expression (116). In this study, it was observed that adult mice administered with BPA developed motor hyperactivity, as well as a reduced expression of several genes involved in GABAergic and dopaminergic pathways within midbrain: dopamine transporter 1, GABA transporter three and glutamate/aspartate transporter (117). These observations suggest that BPA induces alterations in the dopaminergic and GABAergic systems within the midbrain, which, at the same time, may cause a dysfunctional release of neurotransmitters in the prefrontal cortex (PFC) and striatum, thus contributing to increase the ADHD susceptibility (117, 118). Accordingly, it has been proposed that the hyperactive and inattentive behaviors that characterize ADHD may be caused by low levels of dopamine in the striatum and PFC, "leading to a more active glutamatergic output to the striatum, as well as an increased glutamate signal to the PFC" (117).
This evidence supports the idea that there is an association between perinatal exposure to BPA and ADHD's appearance. Future research should be aimed at understanding the underlying cellular and molecular mechanisms, and whether the reduction in perinatal BPA exposure decreases the prevalence of ADHD.
It has been well documented that BPA exposure produces its effects interfering with the action of estrogen and androgen receptors (4). These hormonal effects have been extensively revised and include various endocrine pathways (see below). Nevertheless, in the last decades, there have been described many other hormonal alterations as a consequence of BPA that may have effects in the neuroendocrine systems, leading to any of the neurodegenerative and neurodevelopmental disorders we have mentioned above. Below we will describe, in general, some of the endocrine mechanisms in which the BPA participates.
1. Estrogen receptors (ERs): BPA binds to nuclear ERs exerting distinct agonist or antagonist actions depending on the target tissues, cell types, ER subtypes, and differential cofactors recruited by ER-ligand complexes (4). It has a lower affinity for ERα and ERβ in comparison to estradiol (119); it has been reported to have an 80-fold higher affinity for ER (which is expressed in placenta and mammalian fetal brain) than for Erα in vitro (120) and in vivo (121). BPA has a half-maximal effective concentration (EC50) of 10–100 nM for the ERα (122). Also, BPA can bind to membranal ERs such as GPR30 (123) and exert rapid non-genomic estrogenic responses (124) even at deficient concentrations (10 FM to 10 nM) (125).
2. Androgen Receptors (ARs): BPA is an antagonist of ARs (125). It has a maximal inhibitory concentration (IC50) value of 1–2 mM against the AR. BPA inhibits nuclear translocation of the AR and interferes with its function via multiple mechanisms (122). Also, BPA can bind to sex hormone-binding globulin (126), altering the balance among androgens and estrogens (127).
3. Aryl hydrocarbon receptor (Ahr): Ahr is a transcription factor that mediates the effects of polyaromatic hydrocarbons. It forms a heterodimer with the Ahr-nuclear translocator (Arnt) to regulate cytochrome P450, xenobiotic metabolism, and immune suppression. BPA decreases Ahr's activity in vitro (128). In vivo, gestational exposure to BPA increases Ahr mRNA expression in the brain, testes, and ovaries (129). BPA also upregulated the mRNA level of the Ahr repressor (Ahr) and Arnt in mouse embryos, altering the expression of Ahr and related factors and xenobiotic-metabolizing enzymes (130). Thus, BPA may interfere with Ahr binding, translocation into the nucleus, and activation of co-transcription factors (131). BPA can also reduce aromatase activity in vitro (128) and the synthesis of testosterone and estradiol in vivo (132).
4. Thyroid hormone receptors (TRs): BPA binds to TRs (α and β subtypes) with relatively low affinity (133). It acts as an antagonist by suppressing transcriptional activity (134) and gene expression of TRs in vitro and in vivo (135,136). Likewise, this EDC can inhibit TH sulfotransferase activity (137). See Table 1 for more details regarding the effects of BPA on TRs within the CNS.
| Hormonal mechanism | BPA exposure protocol | Offspring`s age when evaluated | BPA-induced effects on the hormonal mechanism | Reference |
|---|---|---|---|---|
| Estrogen receptors |
-Gestational exposure (days 4-7 post-fertilization) in zebrafish | - Up-regulated expression of the estrogenic marker (cyp19a1b gene, Aromatase B) in the brain of developing zebrafish, through ERα. BPA did not show estrogenic activity | (181) | |
| -Perinatal BPA exposure (40 mg/Kg/day) | -PND 85 in Sprague-Dawley rats | - Decreased expression of ERα mRNA in the hippocampus, along with elevated DNA methylation of the ERα gene promoter | (44) | |
| -Gestational exposure, BPA (2.5, 25, 260, or 2700 μg BPA/Kg/day) | -Prepubertal and young adult female rats | - Decreased expression of ERα in the AVPV of the hypothalamus and ERb in the adult female rat AVPV and MPOA, relative to vehicle controls | (182) | |
| - Lactational exposure; dams were fed with BPA dissolved water (0.1mg/L), until weaning | -PND 7-21 rats | - Decreased ERα phosphorylation and impaired translocation to nucleus in male`s hippocampus | (29) | |
| -2.5 or 25 μg/Kg/day BPA from gestational days 6-21 | -PND 1 rats | -Altered levels of ERa and b within the anterior and mediobasal hypothalamic and amygdala in a sex-specific manner |
(183) | |
| -BPA (40 mg/Kg/day) | -Pregnant/ lactating or estrous cycling young adult rats | -Decrease in ERa-ir cells in the arcuate nucleus of the hyppthalamus in the lactating group | (184) | |
| Aryl hydrocarbon receptor (Ahr) | Gestational exposure, BPA |
-Embryonic days 14.5 and 18.5 mid-and late-developmental stage, respectively | - Increased mRNA expression of AhR and related factors (Ahrr and Arnt) in the brain and cerebellum of male and female embryos | (129, 130) |
| Thyroid hormone (TH) and thyroid hormone receptors (TRs) | -Perinatal BPA exposure (0.1 mg/L in drinking water) spanning gestation and lactation | -Sprague-Dawley rats at PND 21 and 90 | - Up–regulated mRNA expression of the TH transporter monocarboxylate 8 mRNA at PND 21 |
(30) |
| -Cerebellar cells treated with BPA (10-10 M) | - Decreased TR mRNA and protein expression (BPA alone) |
(185) | ||
| -GH3 cells (rat pituitary gland epithelial tumor cell line) treated with 10-11-10-7 M BPA diluted in ethanol | - BPA binds to TRs in a non-competitive pattern. |
(186) | ||
| 000-Pregnant mice were injected s.c. with 20 mg/kg of BPA daily from embryonic |
-Embryonic days 14.5 and 16.5 | - Morphological and molecular changes during development of the mouse forebrain. |
(17) | |
| -Gestational and lactational exposure, BPA (1,10,50 mg/kg) | -Pups of PND 15, Sprague- Dawley female rats | - Increased serum thyroxine levels. |
(175) | |
| -Purified oligodendrocyte precursor cells (OPCs) treated with 10-7 M |
-Inhibition in the differentiation of OPCs induced by exposure to TH (T3). The effect is mediated through various mechanisms via the TRβ1 (responsible for OPC differentiation) | (188) | ||
| Hypothalamic–pituitary–adrenal (HPA) axis and glucocorticoid receptor (GR) | -Perinatal BPA exposure. Female breeders were orally administered with 2 μg/kg/day BPA from gestation day 10 to lactation day 7 | -PND 80, male rats | -Reduced levels of the GR mRNA in the hippocampus. |
(36, 39) |
| Vasopressin (AVP)/ Oxitocin (OT) | -Gestational and lactational BPA exposure (2.5, 25, or 2500 |
-Brains obtained from juvenile rats | - Elimination of sex-differences observed in OT receptors expression in three different hypothalamic regions in control animals of both sexes. Males are more susceptible to these changes | (142) |
| - Dams were gavaged from gestational day 6 until parturition with BPA (2.5, 25, 250, 2500, or 25000 μg/kg/day) | -Sprague-Dawley rats, PND 1. | - Disruption of neonatal amygdala´s transcriptome in a sex-specific manner: females are more sensitive to BPA exposure during fetal development |
(141) | |
| -Gestational exposure | -Embryonic day 18.5 | -Lower brain gene transcript levels for AVP and OT. |
(22) | |
| -Beginning at birth to PND 3 (4 injections), BPA (50 or 50 μg/kg) | -Female Long-Evans adult rats (6 months old) | - Both doses of BPA increased the number of OT-ir neurons within the PVN of the hypothalamus | (54, 55) | |
| -Oral exposure of BPA (5,50 and 50,000 mg/kg/day) during 8-14 PNDs | -Prairie vole pups | -The highest dose of BPA increased the number of AVP-ir neurons in the anterior portion of the PVN and decreased OT-ir neurons in the posterior portion of this hypothalamic nucleus |
(8) |
5. Hypothalamic–pituitary–adrenal (HPA) axis and glucocorticoid receptor (GR): BPA has been found to alter the HPA axis in a sex-dependent manner. Under basal conditions, BPA treated females had higher corticosterone (CORT) levels than BPA-treated males. Moreover, when they were submitted to a mildly stressful experience, male and female rats exposed to BPA exhibit enhanced levels of CORT compared to controls. Also, in females, GR levels were increased when compared to males (138, 139). Another study reported basal levels of serum CORT, adrenocorticotropin (ACTH), and corticotropin-releasing hormone mRNA were increased in male BPA-treated rats, but not in females, compared to animals that received vehicle. Following mild-stressor, the elevation of CORT or ACTH levels was higher in male BPA-treated rats, whereas it was lower in females treated with BPA than in vehicle-injected controls (36, 39). Finally, BPA exposure led to abnormal adrenal gland histology, including reduced zona reticularis, especially in males, hyperplasia of zona fasciculata in both sexes, and increased adrenal weight in females (139). These alterations might be related to enhanced anxiety and depressive-like behaviors reported in BPA exposed subjects since the HPA axis, and GR are closely related to these psychiatric entities.
6. Neuropeptides (Vasopressin (AVP)/OT): Although the vast majority of the literature focuses on steroid hormone signaling related impacts, growing evidence demonstrates that AVP and OT are also BPA targets (140). BPA exposure can interfere with AVP and OT signaling pathways in the developing brain and alter signaling pathways critical for synaptic organization and transmission (141). Effects include the elimination or reversal of well-characterized sexually dimorphic aspects of the AVP system, including innervation of the lateral septum, and other brain regions critical for social and other non-reproductive behaviors. Disruption of magnocellular AVP function has also been reported in rats, suggesting possible effects on hemodynamics and cardiovascular function (142). Finally, it has been demonstrated that BPA interferes with uterine contractility, reducing its force (143, 144), suggesting an altered OT signaling. See Table 1 for more details regarding the effects of BPA on AVP and OT expression and functioning in the CNS.
It is essential to note that all of these mechanisms are involved in the modulation of distinct behavioral patterns, such as sexual behavior (as in the case of ER and AR), anxiety and depressive-like behavior (such as ERs, GR and AVP), memory and learning (ERs and GRs), and social behaviors (OT). Thus, it is easy to infer that if there are alterations in the expression of these molecules, it can provoke behavioral disturbances. For example, in the OT case, which expression is altered by BPA exposure, it can provoke fewer social interactions in the progeny, an effect that can be related to ASD.
It is also worthy of mentioning that the molecules listed before can exert a broad of physiological and behavioral effects depending on the receptors they activate. Besides, the expression of these molecules is modulated depending on age, sex endocrine condition, and brain region, making it more complex to understand its interactions with BPA. For example, the effects of estradiol when binding with its receptors can be contrasting; regarding anxiety-like behavior, it elicits anxiogenic effects when it interacts with ERα, whereas anxiolytic effects are observed when it activates ERβ receptors (145). Thus, the effects of BPA on the different systems involved in the regulation of behavioral features may consider biological and methodological issues when analyzed.
Here we present a table that summarizes the primary experimental evidence of the BPA-induced endocrine mechanisms within the SNC and in nervous cells (Table 1). As the reader will realize, some of these mechanisms have been scarcely explored within the SNC, which guarantees there will be future studies in this field.
Epigenetics is related to inheritable changes in DNA, which do not affect the sequence of genes but do alter their reading. Thus, epigenetic changes refer to chemical labels attached to DNA or histones: DNA methylation, acetylation, differential histone modifications, and modulation of the levels of noncoding RNAs (146, 147). Recently, the term "epigenetic toxicity" has emerged, and it is defined as "a phenomenon in which chemical substances affect epigenomes and exert undesirable effects on living organisms, which may explain long-term and transgenerational effects of chemical compounds and the predisposition to diseases due to environmental factors including chemicals" (148). Therefore, it has been proposed that prenatal exposure to BPA may cause epigenetic toxicity in the brain of children and lead to neurodevelopment impairment, affecting memory and learning, even in adult life. In that sense, several in vitro and in vivo studies have assessed the effect of BPA exposure on the brain epigenome.
For instance, Yeo and colleagues evaluated mice gestationally exposed to a high dose of BPA (administered in the diet) and BPA-exposed brain-derived primary cultured neurons (immediately after birth) (149). With this study, it was demonstrated that BPA leads to "a delay in the perinatal chloride shift caused by significant decrease in potassium chloride cotransporter 2 (Kcc2) mRNA expression in developing rat, mouse, and human cortical neurons" (149) and that this effect is accentuated in females. As the concentration of chloride in the neurons increases, the change to an inhibitory GABAergic activity is delayed. It is essential to highlight that BPA is likely to cause this effect because it produces Kcc2 methylation and interferes with brain development (148). It was reported that mice exposed to BPA in an embryonic stage showed an altered neocortical development by accelerating neuronal differentiation/migration, disturbed murine behavior, and disrupted neurotransmitter system (including monoamines) (150). These effects persisted in the adult stage of the BPA-exposed mice. The authors reported that the molecular mechanisms that partially underlay the observed changes are epigenetic alterations in promoter-associated CpG islands (150).
On the other hand, it was demonstrated that prenatal exposure of mice to BPA caused changes in the hippocampal and cerebral cortex expression of genes encoding ERs, altered mRNA levels of epigenetic regulators, such as DNA methyltransferase (DNMT) 1 and DNMT3A. It favored anxiety-like behaviors (151, 152). These changes were observed in a persistent, sex-specific, dose-dependent, and brain region-specific manner. Thus, changes in ERα and DNMT expression distinctly occurred in the males' cortex and females' hypothalamus, and, importantly, these alterations were associated with DNA methylation changes in the ERα gene (151, 152). Moreover, in a recent study, it was reported that perinatal BPA exposure in females resulted in hypermethylation of brain-derived neural factor (Bdnf) gene promoter, and a weak negative correlation was found between DNA methylation of this gene and spatial learning and memory scores (153). Together, these observations reveal that exposure to BPA in a critical period of brain development may be a risk factor for learning and memory later in life.
Interestingly, Alavian-Ghavanini and colleagues conducted one of the first studies that evaluated the effects of BPA in neurodevelopment (154). Accurately, they assessed the effects of "early life BPA exposure on methylation levels of the ionotropic glutamate receptor NMDA type subunit 2B (Grin2b) gene in brain tissue from rats and buccal samples from humans". Surprisingly, these authors found that early exposure to BPA induced an opposite-sex-dependent effect in humans and rats: while in the hippocampus of female rats, the methylation of Grin2b decreased, in 7-years-old girls, it increased. Although this study reflects that early exposure to BPA induces changes in the methylation of a regulatory region of the Grin2b/GRIN2B gene, it is crucial to consider that it presented some limitations and methodological biases: 1) The use of different animal species; 2) The age or stage on which the samples were taken (those of humans were in childhood, while those of rats were in adulthood); 3) Distinct types of samples.
It was reported that in humans, "Grin2b methylation was also associated with a low APGAR score, which has been documented as a risk factor for NDD" (154). However, future studies should deepen into behavioral and memory evaluation and their association with levels of exposure to BPA at different stages of life.
An epigenetic mechanism that has been associated with the age-related cognitive decline -which is exacerbated in neurodegenerative diseases- is the modification of chromatin, including histone acetylation. Thus, Jiang and colleagues orally exposed pubertal mice to BPA to assess if the exposure to this substance exacerbates the age-related impairment of spatial cognition in elderly, and "whether serum sex and thyroid hormones or hippocampal histone acetylation (H3K9ac and H4K8ac) are associated with cognitive effects" (155). The results showed that aging, per se, caused a marked decline of spatial learning and memory, and decreased levels of the hormones mentioned above, and hippocampal H3K9ac, and H4K8ac. Although BPA exacerbated all of these effects, it was also reported a positive correlation between free thyroxine, H3K9ac and H4K8ac, and spatial memory abilities. Similar results on the effects of BPA on histone acetylation at the hippocampal level were reported by Zhang et al. (156), who performed a murine study with slight methodological differences, showing similar results, reporting increased H3K14 acetylation and inhibition of histone deacetylase 2. These results suggest that pubertal exposure to BPA modifies histone acetylation related to an exacerbated age-related cognitive decline.
Finally, it has been proposed that BPA-induced epigenetic mechanisms may be related to stress-responsiveness and related behaviors. In that sense, Kitraki and colleagues (157) examined the epigenetic effects of perinatal BPA exposure in the GR gene and its regulator Fkbp5 in the hypothalamus and hippocampus of juvenile rats. In this study, it was observed that exposed male rats had increased DNA methylation of Fkbp5 and reduced GR protein levels in the hippocampus. Furthermore, these results suggest that these epigenetic changes may be regulated by the ER β (157). Therefore, these data show that developmental BPA exposure may have a long-lasting impact on some stress-responsibility pathways.
All the evidence presented in this section supports the idea that exposure to BPA causes epigenetic alterations that, in turn, can influence at a neurological level (Figure 1). However, future studies must focus on better understanding the associated molecular mechanisms and homogenizing methodological approaches. Likewise, it is crucial to study the relationship between these epigenetic mechanisms (induced by exposure to BPA) and the development of the diseases that have been presented in this work.
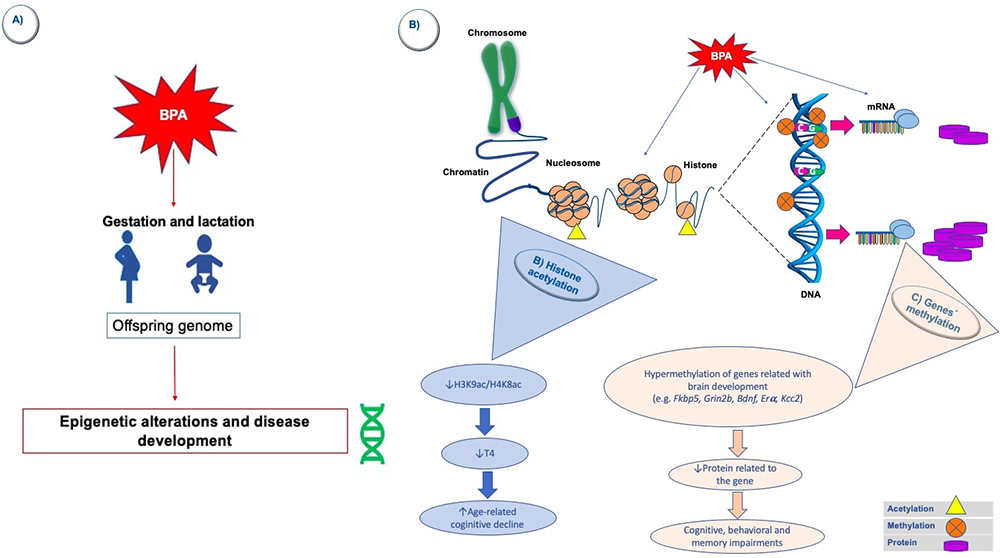 Figure 1
Figure 1BPA-induced epigenetic changes in the brain. A) Exposure to BPA in critical development stages (during gestation or lactation, for example) may induce some modifications in offsprings` epigenome. Specifically, in the brain, some in vivo and in vitro studies have revealed changes in some epigenetic marks caused by the exposure to this EDC. First, it has been observed that exposure to BPA can induce a reduction in hippocampal acetylation of some histones (H3K9/H4K8) (B); in consequence thyroxine levels decrease and this is associated to an accelerated age-related cognitive decline that may lead to the development of neurodegenerative diseases. Furthermore, some genes related to the neurodevelopment (e.g. Fkbp5, Grin2b, Bdnf, Era, Kcc2) are hypermethylated in the hippocampus or cerebral cortex after gestational or lactational BPA exposure (C); therefore, the proteins codified from these genes decrease and cognition, behavior and memory may impair as it occurs in some neurodevelopmental diseases.
Mitochondria are organelles specialized in energy production, as well as redox and calcium homeostasis. The energetic mitochondrial function is necessary for ATP-dependent intracellular reactions such as endocytosis, ion transport, and neurotransmitter biosynthesis. It has been recently discovered that mitochondria can change its shape by regulated processes of fusion and fission, which allow the active traffic between cell compartments (soma, axon, and presynaptic boutons) (158). While fusion has been related to the maintenance of mitochondrial functions (159), fission is associated with apoptosis (160). Several proteins are responsible for the mitocondria`s dynamic balance within the brain. For example, dynamin-related protein (Drp)-1 and fission protein (Fis)-1 are related to mitochondrial fission, whereas mitochondrial dynamin-like GTPase (Opa)-1, mitofusin (Mfn)-1, and Mfn-2 to mitochondrial fusion. Nevertheless, it has been suggested that the disruption of either fusion or fission could lead to the development of neurodegenerative (161) and behavioral disorders, such as ASD (162).
In the last years, it has emerged a growing interest in exploring the effects of exposure to BPA on brain mitochondrial functions and integrity. For example, Agarwal and colleagues' work showed that early postnatal short-term exposure to BPA activates autophagy, leading to an increase of OS, mitochondrial fragmentation, and apoptosis in hippocampal neural stem cells (NSCs). Furthermore, overexpression of autophagy-related genes occurs in response to BPA-induced neurotoxicity via AMP kinase (AMPK)/mammalian target of rapamycin (mTOR) pathways (163). In another study of this group, it was demonstrated that exposure to BPA (gestational, lactational, and during adulthood) caused hippocampal neurotoxic effects through mitochondrial dysfunction, provoking increased OS, apoptosis and reduced neurogenesis (164). The disruption of the mitochondrial dynamics produces these neurodegenerative effects induced by BPA (e.g., dysregulated fission) in the rat brain hippocampus. In that sense, dynamin-related proteins (e.g., Drp-1, which is upregulated after BPA exposure) have emerged as potential targets to mitigate the BPA-induced neurotoxicity. However, the effects of BPA exposure on dynamism and other mitochondrial functions should be studied in greater depth. The mechanisms here described can be seen as represented in Figure 2.
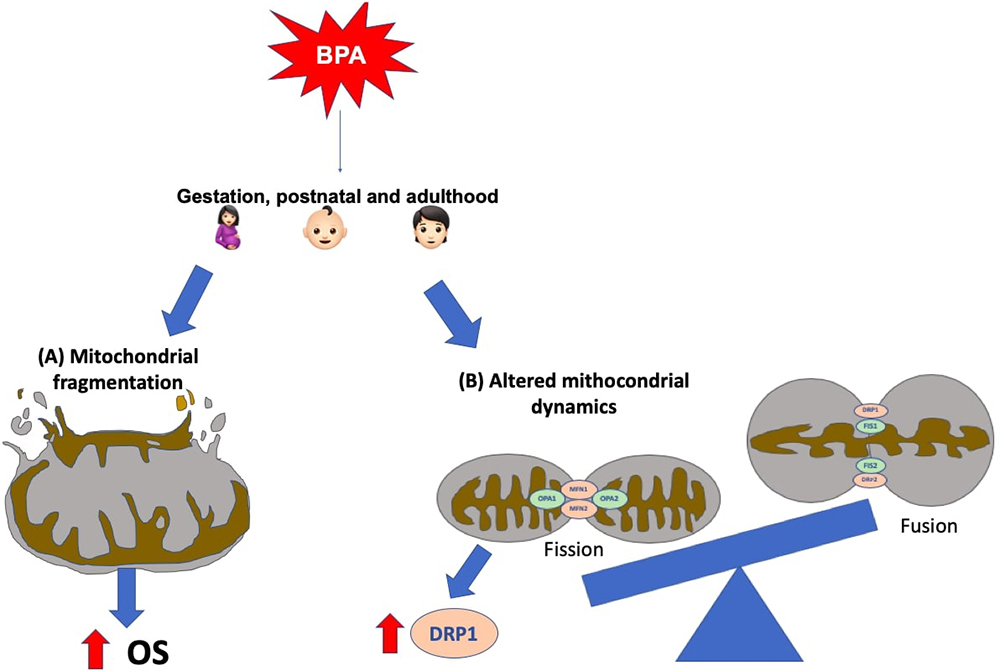 Figure 2
Figure 2Mitochondrial-related BPA-induced mechanisms in the brain. It has been observed both, in vivo and in vitro, that gestational, postnatal and adult BPA exposure leads to hippocampal mitochondrial alterations, such as: (A) fragmentation, that leads to and increased oxidative stress (OS), and (B) dysregulation in mitochondrial dynamics. The latter has been related to dynamin-related proteins, for example, Drp-1, which is up-regulated secondary to BPA exposure.
Calcium (Ca2+) ions are important cell signaling regulators implied in a wide variety of cellular processes (165). In the CNS, Ca2+ is critical for neuronal survival and function. Intracellular Ca2+ homeostasis depends on its rapid redistribution into the diverse subcellular organelles, which serve as stores of this ion (166). On the other hand, Ca2+ communicates with some other systems and pathways, for example, with ROS (166). Several sources can generate these reactive species as "by-products of mitochondrial respiratory chain activity and by several extramitochondrial enzymes such as NADPH oxidases, xanthine oxidase, uncoupled nitric oxide synthase, myeloperoxidase, cytochrome P450, cyclooxygenase, and lipoxygenase" (166).
Nowadays, it is widely accepted that ROS and Ca2+ have bidirectional interactions, while "ROS can regulate cellular calcium signaling. Calcium signaling is essential for ROS production" (166). For example, when intracellular Ca2+ levels increase, some of the ROS-generating enzymes activate and, consequently, free radicals are produced. Therefore, excessive Ca2+ influx and ROS levels can lead to neural disorders and psychiatric diseases (167).
Since BPA can cross the blood-brain barrier, much has been speculated about its ability to interfere with Ca2+ homeostasis and redox balance. It has been reported that mouse hippocampal HT-22 cells exposed to high doses of BPA showed lower viability than controls. This was because BPA induced an increased generation of ROS, caused by a higher intracellular concentration of Ca2+ (168). Furthermore, the pretreatment of BPA-exposed cells with inhibitors for Ca2+ leads to decreased cell death. Therefore, these results suggest that Ca2+ is involved in BPA-induced apoptotic cell death in HT-22 cells (168). Besides, it was found that BPA caused a transient Ca2+ and ROS generation in neuroblastoma cells (SH-SY5Y). It was also reported that these cells had a higher expression of Alzheimer's Disease-related proteins, such as beta-secretase (BACE)-1, amyloid protein precursor (APP), amyloid-beta (Aβ) 1–42 and phosphorylated tau proteins (169).
Recently, it was reported that after chronic exposure to BPA, human embryonic stem cells-derived human cortical neurons (hCNs) showed elevated intracellular Ca2+ levels through the activity of an NMDAR. BPA caused an increase in ROS generation, attenuating the antioxidant defense in hCNs (170). Other authors have also reported similar effects in vitro (171, 172), and in vivo (173–176). Other BPA-induced effects observed in the hCNs model were endoplasmic reticulum stress, overexpression of cytochrome c (caused by mitochondrial impairment), and increased cell apoptosis by regulating the Bcl-2 family and caspase-dependent signaling pathway. All these results suggest that BPA induces neurotoxic effects on hCNs by eliciting apoptosis, which may be caused by the disruption of Ca2+ homeostasis and the deleterious effects in some cell organelles (170).
All the evidence presented above shows that there is a relationship between chronic exposure to BPA and disruption of Ca2+ and redox homeostasis, which cause neurotoxic effects that probably favor the development of neurodegenerative diseases and behavioral disturbances (Figure 3).
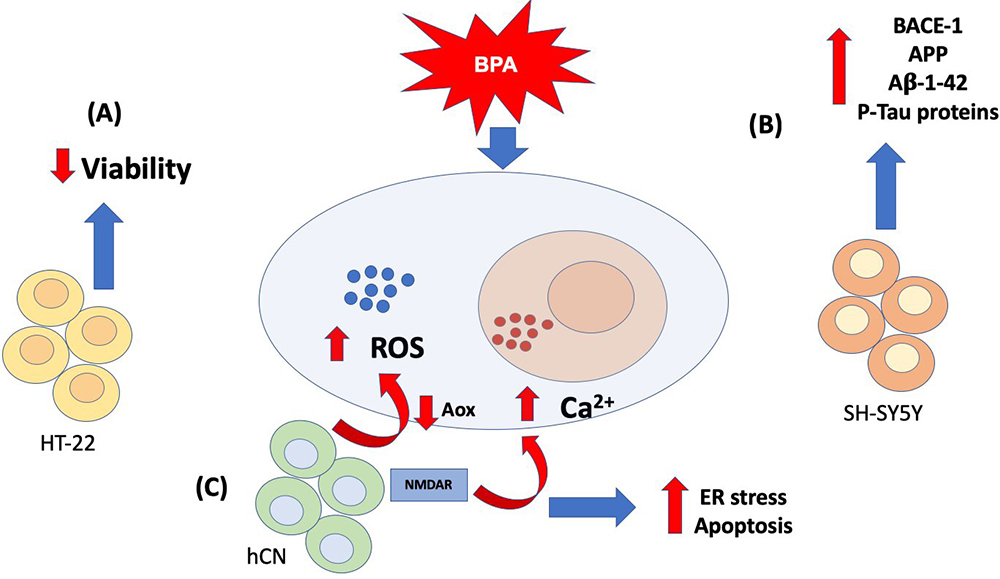 Figure 3
Figure 3Calcium and oxidative stress-related BPA-induced mechanisms in brain cell lines. So far, in vitro approaches have revealed that exposure to BPA increases reactive oxygen species (ROS) and intracellular calcium (Ca2+), although the effects of this altered homeostasis in different types of brain cell lines. In (A) mouse hippocampal HT-22 cells the increase of ROS and Ca2+ leads to a decreased cell viability. In (B) neuroblastoma cells (SH-SY5Y) it has been observed an increased expression of Alzheimer´s disease-related proteins, such as beta secretase (BACE)-1, amyloid protein precursor (APP), amyloid beta (Aβ) 1–42 and phosphorylated tau proteins (P-Tau). Finally, (C) human cortical neurons (hCNs) showed elevated intracellular Ca2+ levels through the activity of a N-methyl-d-aspartate receptor (NMDAR), while increased ROS may be related to a decreased antioxidant activity. Other observed effects related to the ROS and Ca2+ dysregulation are: endoplasmic reticulum (ER) stress and increased apoptosis. All of these neurotoxic effects may lead to the development of neurodegenerative diseases in humans.
"Inflammation is a biological response of the immune system that can be triggered by a variety of factors, including pathogens, damaged cells, and toxic compounds" (177). Consequently, an inflammatory response is initiated and is defined as "the coordinate activation of signaling pathways that regulate inflammatory mediator levels in resident tissue cells and inflammatory cells recruited from the blood" (177). It has been shown that exposure to BPA can alter the production of inflammatory cytokines and subsequently cause an immune dysfunction (176). Therefore, for this review, we will describe some of the existing evidence of how BPA causes an activation of the inflammatory response in the CNS, leading to the development of neurological diseases.
It has been reported that the chronic oral exposure of mice to BPA leads to an increased brain mRNA expression of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6), and decreased anti-inflammatory IL-10 (176). Therefore, this demonstrates the BPA capacity to induce a pro-inflammatory response within the CNS. This local inflammatory response must be regulated by the cells that reside in the CNS; therefore, some authors have proposed that the microglial cells could be those that orchestrate BPA-induced inflammatory responses. Inline, Zhu and colleagues reported that exposure of BV2 cells (a murine microglial cell line) to BPA caused its activation and increased IL-6 and TNF-α mRNA expression levels. Results suggest that this inflammatory reaction is mediated by ER signaling, MAPKs, and NF-κB pathways (178).
Nevertheless, this latter effect could also be mediated by the activation of extracellular signal-regulated kinase (ERK) and c-Jun N-terminal kinase (JNK), as it was observed after BPA exposure of HT-22 cells (178). Though, in this last study, the effects of BPA on the secretion of inflammatory mediators (such as cytokines) were not reported. Therefore, future studies in this regard must focus on generating in vivo models that evaluate a vast repertoire of inflammatory factors and the signaling pathways that might be involved.
Recently, it has been proposed that BPA may have the potential to trigger an autoimmune response in the CNS. Therefore, Vojdani and colleagues demonstrated that BPA could bind to human serum albumin (HSA), forming a new antigenic epitope that could induce autoimmune reactivity (179). In a later study, they found a high correlation between the levels of IgM antibodies against BPA-HSA with protein disulfide isomerase (PDI, a BPA target), myelin essential protein (MBP) and myelin oligodendrocyte glycoprotein (MOG) antibodies (180). This data suggest that BPA may trigger the production of antibodies against PDI, MBP, and MOG, which are usually found in some neurodegenerative diseases. Although this study shows that BPA indirectly induces the formation of these autoantibodies, it has the limitation that it was only evaluated at the serum level and in healthy subjects; hence, it does not reveal the interaction of these antibodies within the brain of patients with any neurological disorder. Future studies must focus on obtaining direct evidence on BPA's role in inducing an autoimmune response in the CNS. The mechanisms mentioned in this section are summarized in Figure 4.
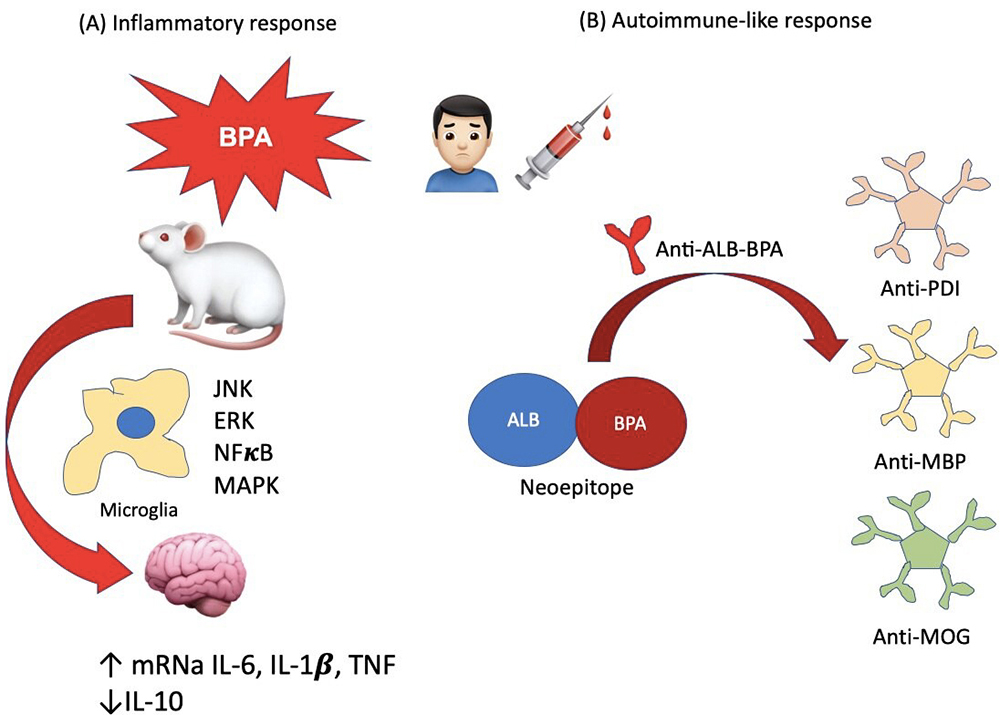 Figure 4
Figure 4Brain inflammatory and peripheral autoimmune-like BPA-induced pathways. (A) It has been reported that mice exposed to BPA increase their brain mRNA expression of some proinflammatory cytokines such as IL-6, IL-1 b and TNF-a; on the contrary, the mRNA of anti-inflammatory IL-10 is decreased. These effects may be regulated by activated microglia, via the activation of extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), some other mitogen-activated protein kinase (MAPK) and nuclear factor (NF) kB. On the other hand, (B) it has been observed that BPA can bind to serum albumin (Alb-BPA), thus forming a neoepitope, which can target B cells to secrete anti-Alb-BPA antibodies. These also may induce the secretion of auto-antibodies such as anti-protein disulfide isomerase (PDI, a BPA target), anti-myelin basic protein (MBP) and anti-myelin oligodendrocyte glycoprotein (MOG) antibodies.
BPA is a ubiquitous compound that has been classified as an endocrine disruptor due to its physicochemical and functional properties. Owing to this last characteristic, BPA can dysregulate various biological pathways and be a risk factor for human and animal health.
In that sense, there has been a particular interest in elucidating whether there is an association between exposure to BPA and neurological effects in recent years. Various pieces of evidence suggest that exposure to BPA in critical stages of development, influences in neurodevelopment. Therefore, BPA can be considered a risk factor for the onset of several cognitive and behavioral disturbances, memory, and learning. There is also evidence that BPA is a risk factor for the development of certain neurodegenerative diseases.
The cellular and molecular mechanisms that underlie the neurological effects of exposure to BPA are very diverse. They include neuroendocrine disruption (in hormonal axes, neurotransmitters, neuropeptides and others), mitochondrial, epigenetic and inflammatory effects, as well as imbalance in the redox state and of Ca2+.
As here reviewed, there are a large number of studies that support this association. However, there are certain discrepancies in the results due to methodological differences between one study and another. Therefore, it is vital to homogenize methodological issues in studies performed in animal models and clinical studies. It is also important to make methodological designs that evaluate the effects of several pollutants simultaneously, as it occurs in the environmental exposure of humans and animals.
Since sexual non-reproductive and dimorphic behaviors are sensitive to endocrine disruption during development, it has been pointed out by some authors that it is necessary to be vigilant towards the potential adverse effects of BPA exposure and, therefore, take actions to inform women planning or undergoing pregnancy about measures to reduce or avoid exposure to BPA (4, 67).
Drs. Daniela Rebolledo-Solleiro and Helena Solleiro-Villavicencio contributed equally to this work. We are thankful with Dirección General de Asuntos del Personal Académico (DGAPA) from the Universidad Nacional Autónoma de México (UNAM) for the grant given to Daniela Rebolledo-Solleiro (2017-2019) throughout her postdoctoral fellowship and with Consejo Nacional de Ciencia y Tecnología (CONACYT) for the support of this work through grant number 610805 given to Laura Yesenia Castillo Flores during her doctoral studies.
Aβ
amyloid beta
adrenocorticotropin
attention deficit hyperactivity disorder
Aryl hydrocarbon receptor
Ahr repressor
amyloid protein precursor
Ahr-nuclear translocator ;ALS: amyotrophic lateral sclerosis
α-amino-3-hydroxy-5-methyl-4-isoxazolpropionic acid
androgen receptor
autism-spectrum disorder
vasopressin
anteroventral periventricular nucleus
beta secretase
brain-derived neural factor
basolateral amygdala
bisphenol A
central nervous system
corticosterone
dopamine
dopamine transporter
differentially expressed genes
DNA methyltransferase
Dynamin-related protein
endocrine disruptor compound
estrogen receptor
extracellular signal-regulated kinase
glucocorticoid receptor
human cortical neurons
histone deacetylase 2
hypothalamic–pituitary–adrenal axis
human serum albumin
c-Jun N-terminal kinase
potassium chloride cotransporter 2
mitogen-activated protein kinase
myelin basic protein
myelin oligodendrocyte glycoprotein
medial preoptic area
multiple sclerosis
Neurodevelopmental disorders
N-methyl-d-aspartate
N-methyl-d-aspartate receptor
oxidative stress
oxytocin
Parkinson´s Disease
protein disulfide isomerase
prefrontal cortex
post-natal day
phosphorylated tau proteins
paraventricular nucleus of the hypothalamus
reactive oxygen species
substantia nigra
thyroid hormone
thyroid hormone receptor
8-hydroxy-2'-deoxyguanosine.