

Frontiers in Bioscience-Landmark (FBL) is published by IMR Press from Volume 26 Issue 5 (2021). Previous articles were published by another publisher on a subscription basis, and they are hosted by IMR Press on imrpress.com as a courtesy and upon agreement with Frontiers in Bioscience.

1 Health Profession Awareness Program, Health Facilities Development, Hamad Medical Corporation (HMC), Doha, Qatar
2 Precision Medicine, College of Science and Engineering, Hamad Bin Khalifa University (HBKU), Doha, Qatar
3 Developmental Psychology Research Group, Department of Psychology, Kingston University-London, Kingston upon Thames, Surrey, United Kingdom
4 Molecular Genetics Laboratory, Hamad Medical Corporation (HMC), Doha, Qatar
5 Qatar Rehabilitation Institute (QRI), Hamad Medical Corporation (HMC), Doha, Qatar
6 Research and Policy Department, World Innovation Summit for Health (WISH), Qatar Foundation, Doha, Qatar
7 Clinical and Metabolic Genetics, Pediatrics Department, Hamad General Hospital (HGH), Hamad Medical Corporation (HMC), Doha, Qatar
Abstract
The prevalence rate of Autism Spectrum Disorder (ASD) has reached over 1% world-wide prompting governments, health providers and schools to develop programs and policies to address this challenging disorder. Here, we review the cause(s), as well as environmental factors, genetic mutations, and neural pathways that are implicated in ASD. We also discuss the criteria that are commonly used for the diagnosis of ASD and future clinical genetic testing that can aid in the diagnosis of this disorder. Finally, we provide practical steps that can be used to reduce the incidence and severity of ASD, as well as prognosis and treatment of autism.
Keywords
- Autism Spectrum Disorder
- ASD
- Autism
- Environment
- Genetics
- Risk Factors
- Prevelance
- Clinical Diagnosis Review
Autism Spectrum Disorder (ASDs; MIM 209850) is one of the most common and challenging neurodevelopmental disorders in children. ASD is characterized by deficits in communication and social interaction, as well as the presence of repetitive and restrictive behaviors. ASD often manifests with a wide range of comorbidities including morphological (macrocephaly), physiological (gastrointestinal and/or sleep problems) and psychiatric (anxiety) conditions (1). The most common proposed causes of ASD are physiological and metabolic disorders, involving immunity, oxidative stress and mitochondrial dysfunction (2). There is no pharmacological cure to ASD; however, early diagnosis (12 to 18 months) and intervention coupled with remedial services is highly beneficial to patients (3).
The number of individuals diagnosed with ASD has increased dramatically over the past 40 years, affecting roughly 1:60 children (3). This increase in diagnosis has led to large-scale research initiatives, awareness campaigns, and the need for government support. Currently, ASD is diagnosed clinically based on the severity of a heterogeneous list of social, communicative, and behavioral deficits; however, there is no universal standardized assessment for ASD. The broad clinical diagnosis provides practical means to obtain intervention and services, but may adversely impact research, prevalence rates and public understanding of the disorder. This article aims to provide the reader with a succinct understanding of what a current diagnosis of ASD entails, the multidisciplinary research (environmental and genetic) surrounding its causes, as well as, recommendations for future diagnostic methods to improve prognosis.
The prevalence of ASD and the marked increase of diagnosis have been a cause for debate. Roughly, one in 60 individuals are estimated to suffer from deficits falling under ASD. As a result, this is triggering a tremendous strain on the education, health and social service sectors (4). This incidence number appears to be consistent globally and across ethnic and socioeconomic groups (3). A pressing question in recent literature addresses whether the prevalence of autism between the 1970s and the 1990s was underdiagnosed, or if there is a true recent increase in individuals with ASD. While it is hard to quantify the level of influx in individuals with ASD phenotypes, several known factors have been attributed to the increase in clinical diagnosis (Table 1). These factors include: (a) the broadening of diagnostic criteria, (b) the increased efficiency over time in case identification methods used in surveys, (c) changes in diagnostic practices, and (d) the diagnostic substitution (or switching) when some diagnostic categories become increasingly familiar to health professionals and/or when access to better services are insured by using a new diagnostic category (5). Studies analyzing prevalence are vital in assisting providers and governments in their service planning; however, precaution needs to be taken when linking causation with prevalence due to the heterogeneous nature of diagnostic criteria. Interestingly, in a recent US study (6) an association between geographical location and significantly decreased ASD odds suggests the influence of social and environmental factors affecting etiology or diagnosis.
| The impact in measuring the prevalence of ASD |
|---|
| • Awareness |
| • Expanded diagnostic criteria (reclassification) |
| • Diagnostic substitution |
| • To qualify for support services |
| • Reporting practices |
Outside High-Income Countries (HICs), public health investigation into the prevalence of ASD, and initiatives concerning the impact of ASD remain poorly implemented. To date, 86.5% of all cases of ASD have been reported in HICs representing only 20% of the world population. Insufficient population-based studies have been conducted in Low to Medium Income Countries (LMICs), which may well underrepresent the impact of ASD. There are also difficulties in measuring the prevalence of ASD, stemming from population awareness, selection of studies and diagnostic capabilities, as well as cross-cultural appropriateness and comparability of ASD screening, measurement, and epidemiological data. The uneven rates of diagnosis have also led to variations in ASD prevalence by race and ethnicity including the MENA and Gulf regions as stated in the 2016 WISH Report on autism (7). Therefore, the knowledge gap between evidence and action in the care of ASD in LMICs has remained considerably wide.
In searching for a cure to ASD, the emphasis is that it is not a single medical condition, but a heterogeneous array of phenotypes. The co-occurrence of two or more disorders in the same individual was noticed in ASD. Comorbid conditions such as multiple psychiatric disorders anxiety, depression, attention deficit/hyperactivity disorder (AD/HD), epilepsy, gastrointestinal symptoms/problems, sleep disorders, learning disability, obsessive compulsive disorder (OCD), intellectual disability, sensory problems and Immune Disorders were found in ASDs, (1, 3, 8-12)
ASD can be part of known genetic syndromes. ASD was also found to overlap with other genetic disorders such as Rett syndrome (RTT, MIM 312750), Angelman syndrome (AS; MIM 105830), Adenylosuccinate Lyase deficiency (ADSLD; MIM 103050), Tuberous Sclerosis, Tourette’s syndrome (TS; MIM 137580), Fragile X syndrome (FXS; MIM 300624), Timothy syndrome (TS; MIM 601005), PTEN-Related Disorders, Neurofibromatosis type 1 (NF1; MIM 162200), Down syndrome (DS; MIM 190685), Smith–Lemli–Opitz syndrome (SLOS; MIM 270400), Cohen syndrome (CS; MIM 216550), Cornelia de Lange syndrome (CDLS; MIM122470), Prader–Willi Syndrome (PWS; MIM 176270) and untreated phenylketonuria see Figure 1. (1, 3, 8-12). These syndromes share overlapping clinical features with autism including developmental delay, language impairment, seizures and stereotypic behaviors. The root cause of all ASD phenotypes they do not arise from the same origin (mutation). The cause(s) when determined requires personalized treatments, the development of early detection diagnostics, and the establishment of preventive measures.
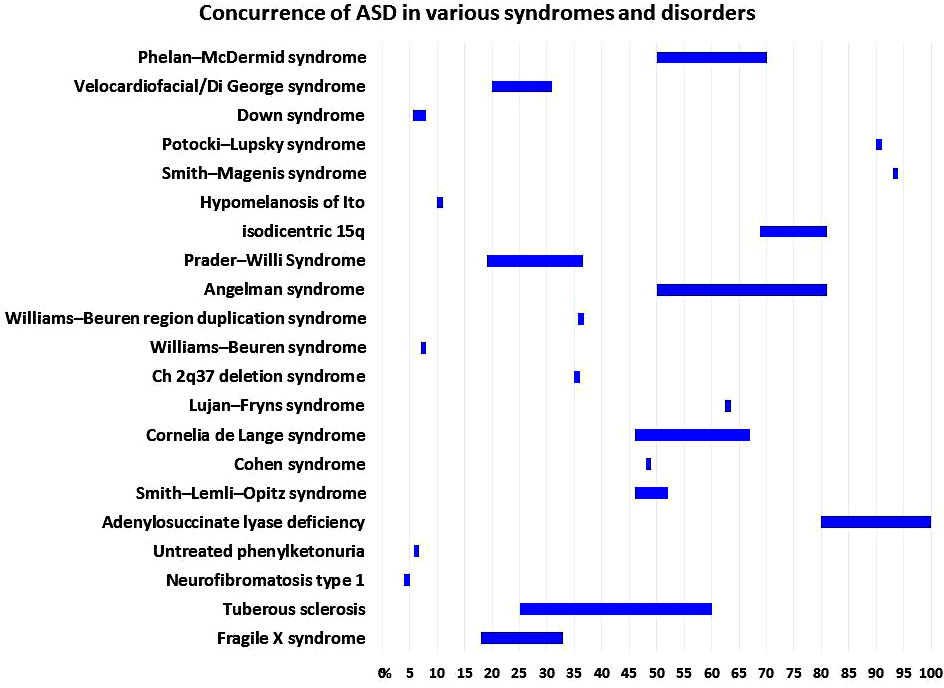 Figure 1
Figure 1Concurrence of ASD in various syndromes and disorders.
In a vast majority of cases, the cause of ASD is idiopathic, and researchers are examining comorbidities, neural pathways, environmental and genetic factors. The most prevalent comorbidity, at roughly 50%, is intellectual disability (3). It has also been proposed that in some individuals’ neural damage associated with seizures leads to deficits associated with ASD. An estimated 20% of individuals diagnosed with ASD also have epilepsy (1). A full list of common and rare comorbidities is shown in Table 2.
| Most common comorbidities |
|---|
| • Anxiety |
| • Sleep disorders |
| • Learning disability |
| • Obsessive compulsive disorder |
| • Developmental coordination disorder |
| • Intellectual disability (50%) |
| • ADHD |
| • Sensory problems |
| • Speech |
| • Stereotypies |
| • Epilepsy (25%) |
| • astrointestinal problems |
| • Immune disorders |
| Rare genetic comorbidities |
| • Tourette’s Syndrome |
| • Tuberous Sclerosis |
| • Fragile X |
| • Rett's Syndrome |
| • Fox 1 Gene |
| • Timothy Syndrome |
| Other risk factors for the development of ASD |
| • In vitro fertilization |
| • Prenatal exposure to air pollution |
| • Prenatal exposure to medications: Valproate (anti-seizure)1, Thalidomide, Phthalates, Pesticides |
| • Prenatal exposure to chemicals and plastics including PCBs and PBDEs |
| • Pre-eclampsia |
| • Gestational diabetes |
| • Infections |
| • Fever (late trimesters, without drug treatment) |
| • Metabolic conditions |
| • Obesity |
| • Advanced maternal age due to increased risk of chromosomal abnormalities in ova of increased age or/and because of unstable trinucleotide repeats |
| • Gestational hemorrhage by causing fetal hypoxia |
| • Prenatal drug use (psychoactive drugs) |
| • Prenatal stress |
| • Maternal mental diseases |
| • Maternal nutrition: folic acid, iron, fatty acids, vitamins |
| • Paternal Age2 |
| • Interpregnancy Interval (time between pregnancies) |
| • Prematurity and postmaturity |
| • Twin birth |
| • Suboptimal breast-feeding |
| • Birth order |
| • Familial socioeconmoic status |
| • Infant gender |
| • Ceserean delivery |
| • Prenatal exposure to hypoxia |
| • Congnital anomolies |
| • Fetal respiratory distress |
| • Post natal complications |
The etiology of ASD is commonly described as a genetic predisposition combined with an environmental impact these factors are delineated below.
The genetic contribution to ASD has been known since the 1970s, after two identical twins were found to have the same condition (13). It has since been determined that the heritable rate is around 80% in identical twins and the conforming rate for sibling twins is around 40%.
Some autistic characteristics can be observed to be part of several well-known syndromes as discussed in comorbidities section. Around 10% of all ASDs are classically associated with dysmorphic features and/or deformities and named Syndromic autism (14-16). The overall incidence of ASD in the syndrome was documented to be significantly higher than incidence of syndrome in the ASD cases. For instance, the highest was observed in ADSLD (80-100%) and the lowest was in Neurofibromatosis type 1 (17). On the other hand, incidence of the syndrome in ASD was found to be less frequent and not exceeding 5% (17). Remarkably the clinical presentations of the syndromic autism are extremely heterogeneous that could be attributed to differences in genetic background and epigenetic influences.
In addition, searches into the genetics of ASD have accelerated in recent years due to the rapid advancement of DNA-decoding technologies (18). With the identification of genetic variants related to ASD, testing at birth or in vitro may become a risk factor identification tool that leads to early intervention. A gender gap also emerges when deciphering etiology of ASD. There is recent support for a ‘female protective effect’ as an explanation for males being four to five times more likely to have the disorder (4, 7, 19).
As technology advances, the list of genes linked to autism is growing. Many of these genes are vital for communication between neurons or they control the expression of other genes. Although over 400 genes have been strongly linked and 200 weakly linked, the contribution of each particular gene within the ASD population is very minor, with none found in more than 2% of patients. It is clear that genetic architecture of ASD is extremely diverse, with contributions from alleles (variant regions within a gene that an individual has two copies of, one being inherited from each parent) of varying frequencies. The most common inheritance patterns of alleles associated with ASD are a dominant variant type, while recessive inheritance patterns are rare, and X-linked or de novo inheritance patterns are very rare. The variant types may include large chromosomal rearrangements, copy number variants (CNV), small insertions/deletions (indels), and single-nucleotide variants (SNVs) (20, 21). Although abundant variants have been associated with ASD, it is difficult to relate them with disease mechanisms.
The largest class of genetic risk for ASD accounts for around 50% of the total, and is estimated to derive from common variants of an additive effect; almost all of which have yet to be identified (22). A further 5–10% of genetic risk is estimated from rare inherited and de novo variants, leaving 40% of the genetic risk, currently, undetermined. Forthcoming whole-genome sequencing (WGS) of complex families will extensively expand our knowledge.
Using a classified approach to analyze autism-related mutations from two large whole-exome sequencing studies, it has been found that while those mutations did not duplicate at the single-nucleotide level, some do at the gene and pathway levels. These modifications combine to affect pathways involved in synaptic function, morphology, and plasticity. The results suggest that isolated rare mutation events are connected and recurrent at higher (gene and pathway) levels. So far, approximately 400–1000 ASD risk genes have been identified and found to be involved in pathways that include synaptic formation, transcriptional regulation, and chromatin remodeling (18, 23, 24). Three explanations have been proposed for increasing replication at the gene and pathway levels: a smaller background space (small sample sizes), high local mutation rates, or true and recurrent disease mechanisms. Based on these findings, it was postulated that a gene-plus-pathway dual-hit model could describe autism genetic associations. This model depends on the effect of the disrupted genes and whether that effect also influences related pathways (25).
Most interestingly, probands (the affected individual) did not simply harbor more variants overall, but more gene-disrupting ones and ones that affected selected pathways. Notably, probands are 55% more likely to have gene-disrupting, and 11% more likely to have missense mutations, than their siblings. These variants have a tendency to repeatedly affect a number of signaling pathways. While at the gene level, 92 genes mapped to these pathways, at the variant level, there were 43 likely gene disrupting, and 75 missense mutations.
These pathways included the, Wnt signaling, GABAergic signaling, synaptic pathways such as neuroligin 3 (NLGN3), NLGN4X, neurexin 1 (NRXN1), and SH3, and multiple ankyrin repeat domains 3 (SHANK3) and glutamatergic synapse pathways, among others. In particular, it was noted that the canonical branch of the Wnt signaling pathway and the whole of the GABAergic signaling pathway are involved. It was also noted that the pathways implicated are consistent yet are separated into two modules. One module, through its inclusion of Wnt signaling, cell adhesion, junction, and cytoskeletal-associated pathways, highlights the role of synapse morphology in autism. Meanwhile, the other module underscores the role of synapse function with its inclusion of glutamatergic synapse, GABAergic synapse, and related processes. This suggests that the synaptic transmission and function and neuronal wiring and morphology have key roles in ASD. Evidence indicates that many of the genes that are mutated in ASD are crucial components of the activity-dependent signaling networks that regulate synapse development and plasticity. Dysregulation of activity-dependent signaling pathways in neurons may therefore, have a key role in the etiology of autism spectrum disorders (26) .
Synaptic genes such as neuroligin 3 (NLGN3), NLGN4X, neurexin 1 (NRXN1), and SH3, and multiple ankyrin repeat domains 3 (SHANK3) have been involved in ASD. For instance, Neurexins act mainly at the presynaptic terminal in neurons and play essential roles in neurotransmission and differentiation of synapses. Some of these roles need the formation of trans-synaptic complexes with postsynaptic proteins such as neuroligins, LRRTM proteins or cerebellin and scaffolding proteins SHANK2 and SHANK3.
Rare mutations and CNVs in the neurexin genes have been connected to ASD and psychiatric disorders, indicating that impairments of synaptic function sustained by neurexins and their binding partners possibly related to the pathomechanism of the ASD (27, 28).
Some view autism to be a synaptic disorder. Some uncommon mutations may cause autism by interrupting some synaptic pathways, such as those involved with cell-cell interaction. Gene substitute studies show that autism is due to changes in activity of synapses and also on activity-dependent modifications. Chromosomal abnormalities and epigenetic changes deserve to be discussed and referenced.
In addition, evidence shows that many of the genes that are mutated in ASD are vital components of the activity-dependent signaling networks that regulate synapse development and plasticity. Dysregulation of activity-dependent signaling pathways in neurons may therefore, have a key role in the etiology of ASD (26).
Several studies showed that impairment of neural connectivity is the most common theory for the pathogenesis of ASDs. For instance, irregular number of neurons as well as abnormal brain and head growth, possibly due to impaired pruning mechanisms, have been documented in ASDs patients (29, 30). Courchesne et al reported that 67% more neurons in the PFC of children with ASDs compared to controls. A subsequent study by Courchesne et al. also found that excess neurons in the PFC, that are eliminated via apoptosis during the prenatal and perinatal periods in just a few months in healthy controls, take years to be eliminated in patients with ASD, thus indicating a relationship between an increase in neurons and ASD pathology.
Another region that is affected by irregular neural connectivity and important in ASD pathology is the amygdala. Children with ASDs have an 11% increase in the numbers of amygdalar neurons compared to controls. However, by adulthood, neurons in the amygdala decrease by 17% in patients with ASDs, while in controls, there is an increase by 11% (31). Abnormal populations of amygdalar neurons, similar to PFC, indicate dysfunctional apoptotic function (32). Clinical studies also revealed significant reduction of neurons in the fusiformgyrus, striatum, and cerebellum of ASD patients (33, 34). While the nature of this relationship is not confirmed, there is growing evidence that proteins and pathways regulating apoptosis are dysfunctional in ASDs (35). As these pathways are explored further, candidates for genetic manipulation arise that may provide answers about the role of apoptotic dysregulation in ASDs.
Thus, ASD is not solely a multi-genetic, but also a multi-pathway disease, and it is eventually a synapse disease. The affected pathways could be a collection of genetic risk factors for autism.
A de novo mutation is one that is not inherited from either parent, these types of spontaneous mutations are estimated to be the underlying genetic cause in between 15% and 25% of cases of ASD (36). They exhibit vast heterogeneity in size – some are small enough to affect a single gene or large enough to encompass many genes and clinical presentations; patients with multiple de novo CNVs usually have more severe phenotypes (37-44). All of these studies found that de novo SNVs have a predominantly paternal origin and the rate of de novo mutation frequency increases with paternal age, concordant with other studies on de novo mutation frequencies (45, 46).
In sum, as research progresses it has become clear that autism is a multifactorial genetic disease that does not follow classical Mendelian inheritance (Table 3).
| Genetic risk factors for ASD | Description of genetic factors | Reference |
|---|---|---|
| De novo large CNVs duplications, such as duplication of 15q, deletions of 22q11.2., deletion of Xp22.3., and duplication or deletion of 16p11.2. (∼600 kb, comprises around 29 genes), along of de novo submicroscopic CNVs such as duplication of 7q11.2.3. and microdeletion of 16p11.2. | The structural chromosomal variations contribution is estimated to be between 15% and 25% and considered as are the underlying cause for many cases of ASD. | (24, 37-44) |
| De novo SNVs in DYRK1A, POGZ, CHD8, NTNG1, GRIN2B, KATNAL2, and SCN2A. | These genes have a predominantly paternal origin and their frequency increases with paternal age. | (45, 46) |
| Regions/loci on chromosomes 20p13 and 7q35 (CNTNAP2) and two loci mapped on chromosome 8. | Regions of genetic susceptibility, these loci are associated with speech delay and social responsiveness | (152, 153) |
| 1-Intergenic variants between CDH9 and CDH10 on chromosome 5p14.1 such as rs4307059, rs10513025, |
These are common variants in ASD. | (44, 156) |
| Neuroligin family NLGN3 and NLGN4 | Rare X-linked mutations in ASD males and mental retardation in several families. | (157) |
| CNTNAP2, SLC9A9, BCKDK, AMT, PEX7, SYNE1, VPS13B, PAH, and POMGNT1 | Rare recessive mutations in consanguineous families were identified in Amish families as well as Middle Eastern families with ASD and epilepsy. | (158-160) |
| The scaffolding proteins family (SHANK1, SHANK2, and SHANK3) as well as neurexin family (NRXN1 and NRXN3) | Rare inherited and de novo variants have been documented in the ASD. | (161-166) |
| ANDP | Novel syndromes associated with ASD such as ASD with facial dysmorphism Rare inherited truncating mutations in genes that are infrequently mutated in the population | (167) |
| FOXP1, MAL and C11orf30 | The dysregulation of these genes have been found to be implicated in the ASDs pathogenesis as several variants have been found to be associated to language delay and autism. | (168-171) |
| AMT, PEX7, SYNE1, VPS13B, PAH, and POMGNT1 | Inherited recessive variants in consanguineous families | (172) |
| noncoding variants in DNAse I hypersensitivity sites located within 50 kb | Variants were found in consanguineous families | (173) |
| RELN, PRRT1, ZFP57, TSPAN32/C11orf21, OXTR, EN2, MTHFR and MECP2 | ASD-specific methylation biomarkers |
(174-186) |
| Non-coding RNA: LncRNA: MSNP1AS, Evf2, RPPH1, NEAT1, and MALAT1and MiRNA hsa-miR-21-3p, hsa-miR-29b, hsa-miR-219-5p, miR-146a, miR-221, miR-654-5p, and miR-656 miR-133b/miR-206 | LncRNA were found to be more likely involved in the molecular function and/or regulation of the specific ASD risk genes | (172, 187-198) |
During the first eight weeks from conception some teratogens or exposure to polluted air like fine/ultrafine particulate matter (PM2.5), dangerous plastic items, hazardous type solvents, metals, and chemicals (e.g., PCBs, Phthalates, Phenols, fire-resistant brominated material, etc.), or detrimental behavior such as alcohol consumption, smoking, and use of illicit medicines, or adverse reaction to some vaccines as well as prenatal stress/tension increase the risk of autism. In animal studies, prenatal exposure to viruses like rubella or cytomegalovirus activates the mother's immune response and may greatly increase the incidence of autism in mice. It seems that congenital rubella syndrome is the most convincing environmental cause of autism. However, not all of these risk factors are evidence-based or have been substantiated to be involved.
In addition to the social, behavioral and communicative deficits, a requirement for an ASD diagnosis is the presence of these phenotypes in early childhood. This time period, from gestation through the first three years, corresponds with significant neurodevelopment. Through ongoing work in defining typical trajectories for maturation of neural circuits utilizing neural imaging, researchers have created a backdrop against which genetic alterations and the effects of adverse experience on brain maturation can be compared. Researchers continue to investigate periods of critical vulnerability during the development of neural circuits and their sensitivity to experiences and environmental exposures (9).
Investigation into consequences of exposure to specific environmental factors at critical moments in time, is now a major field in ASD research. Examples of environmental risk factors include how low levels of folic acid may be linked to ASD in early but not late pregnancy, or that exposure to harmful chemicals and infections have greatest impact during the second and third trimester when brain growth and synapse formation is largest (9). A comprehensive list of environmental risk factors including prenatal, natal, and postnatal can be found in Table 2.
Recent reviews point to an important role for metal neurotoxins in ASD (47, 48) and neurotoxic chemicals interaction with neurotransmitters (49). Both Aluminum (Al) and Mercury (Hg) emerge as the predominant neurotoxic elements. Experimental results indicate that the chemical form can penetrate the blood/brain barrier. Ingestion of these neurotoxic compounds by women of child-bearing years increases ASD risk. Exposure to neurotoxic chemicals have been shown to lead to alterations in neuro-receptor systems related to ASD. Intriguingly, an association was found between air pollution and ASD (50, 51). The research group found that high maternal exposure to particulate matter during pregnancy, especially during the third trimester, was associated with greater odds of a child having ASD. In a separate study, postnatal exposure to nitrogen dioxide (a traffic related air pollutant), was associated with increased odds of ASD.
The exposure itself does not directly cause ASD, but when compounded with other risk factors may trigger changes, which affect neural development, and result in ASD phenotypes. It is the toxic load of chemicals, infectious, psychological, nutritional, and other stressors that can erode an individual’s resilience through effects on the epigenome, endocrine homeostasis, cell signaling, etc. ultimately overwhelming adaptive capacity (9). In searching for environmental factors, the focus is on finding those most prevalent, so that by reducing exposure, the greatest positive impact can occur. Thus, numerous other factor including environmental factors are considered to be responsible for susceptibility to autism through interaction of susceptible genes (Table 2).
Physical, mental, and psychological well-being and the family financial situation throughout the pregnancy are important influencers affecting fetal development and health. An unhealthy mother who is not mentally and/or physically healthy and undernourished is implausible to have a healthy neonate. A set of prenatal risk factors which increase a child's susceptibility to autism is discussed below.
Advanced parental age (particularly paternal age) has been identified as one of the most significant risk factors of autism (52-56). Several studies have showed that maternal and paternal age older than or equal to 34 years their offspring has greater risk of autism; however, in other studies, the relationship between child autism and the age of both parents (57) or even the age of one parent is ambiguous and unclear (58-62). Intriguingly, the relationship between increased risk of autism and elevated paternal age has been shown in few studies (55, 63-69). Particularly, a study that was conducted amongst the Iranian people in 2010 explored the relationship between parental age and autism risk had affirmed such association (68). Based on this study, autism risk increases by 29% for every 10-years elevation in the fathers’ age. In other words, fathers’ aged between 34 and 39 had nearly a two-fold greater risk, and those who are older than 40 have more than two-fold (2.58) greater risk to have an affected child in comparison to those ones who are aged 25-29 years old (68). In other studies in Japan (62) and China (67), similar relationships were explored between paternal age and increased risk of autism. The lack of any correlation between maternal age and susceptibility to autism in these two mentioned studies has critical importance. The probable explanation for this phenomenon is the formation of de novo mutations in germline cells and modifications in DNA methylation, which can result in general epigenetic alterations in the expression of neural developmental genes and, finally, disorders in sperm genomic imprinting. As a result, the probability of neural impairments, such as autism, would be increased (46, 55, 64). Advanced paternal age also affects immune system functions, consequently, the development of the nervous system (63).
In autism studies investigating increase in maternal age, they revealed a correlation between autism chromosomal abnormalities and trinucleotide repeat expansion in the ovule (57-59, 70-72); and a connection between autism and the increased obstetric intervention at advanced maternal age (71). On the other hand, being young at gestational age potentially increases the risk of autism due to the lack of physical maturity, and the inability or poor maternal care. Mothers who are younger than 20 may be exposed to intrauterine growth retardation of fetus and preterm birth, which both are compelling factors associated with increased risk of autism (57, 59, 73).
Infection, a metabolic syndrome or bleeding during pregnancy are among the conditions tied to autism. Maternal bleeding during pregnancy is significantly associated with an elevated risk of autism, nearly 81% (53). Metabolic syndromes, including diabetes (53, 74) hypertension (59, 75) and obesity (75, 76) pave the way for hypoxia (oxygen deficiency) in utero (74) leading to retarded brain development, induction of myelination changes, alteration in membrane adhesion, and deficiency in hippocampal neurons (a brain area which is highly involved in autism) (77, 78). Maternal viral infections including rubella (79-81), measles, mumps, chicken pox (79, 82), influenza (79, 82, 83), herpes (79, 84), varicella zoster (84) and cytomegalovirus (82, 85), and bacterial infections such as pneumonia and syphilis in the first trimester or in the second trimester if they require hospitalization, distress the embryo and increase the risk of autism (79, 86). Such outcome is due to abnormal maternal immune activation and, consequently, elevated levels of inflammatory cytokines which affect the embryonic brain development and alter neuropathophysiological status ergo increasing the risk of autism (86, 87).
The family unit well-being, parental behavior, and communication patterns affect the formation of personality and emotions. There is an association between parental psychiatric history and the threat of developing a child with mental disorders, especially autism. For example, the association of parental psychiatric history such as schizophrenia causes the odd of autism to nearly three-fold (57, 88-91). Moreover, the relationship between the anxiety (67, 90, 92-97), depression (88-91, 98) and personality disorders (88) of the mother, and the susceptibility to autism has been demonstrated in many studies.
Besides mothers that are recognized as mentally ill, those who undergo considerable stress during the 21–32 weeks of gestation, a period of heightened plasticity for fetal formation and development (71, 99), may have irreversible effects through epigenetic mechanisms, expression of fetus stress response genes, and genes involved in neurodevelopment, metabolism, and physiology persisting across lifespan. The psychological state of the mother, especially periods of great and long-lasting stresses result in personality disorders such as aggression (100) causing changes in cortisol levels thus disrupting the HPA axis in the mother, therefore, amplifying adrenal steroids and increasing placental permeability to hormones further impacting the fetus. Consequently, fetal developmental epigenetic programming would be highly affected because of interrupting the fetal stress response system, paving the way for different physical and mental impairments including autism (98).
Use of maternal prenatal medication was suggested to increase the risk of autism. Use of prenatal psychiatric medication cause 68% increase in the risk of autism, likely due to the fact that such drugs pass the placenta and disrupt the fetal development (53). For example, the use of antiepileptic drugs such as valproic acid leads to fetal valproate syndrome due to increased oxidative stress and altered gene expression patterns, subsequently resulting in developmental delays, deficient motor activities and postnatal growth aberrations (82, 101-102). Moreover, it is documented that paracetamol (acetaminophen), which is widely used as an analgesic/antipyretic drug, induces apoptosis and necrosis similar to the legions observed in autistic brains. Additionally, paracetamol (acetaminophen) induces oxidative stress and immune dysregulation in humans (103). Furthermore, positive correlation between antidepressant medications and autism has been demonstrated in many studies (104-106). Other medications display increased susceptibility to autism such as thalidomide, a painkiller, (107), misoprostol, a prostaglandin analog drug for the prevention and treatment of gastric ulcers (108) in the first trimester, and β2-adrenergic agonists like terbutaline to treat asthma (109, 110).
Considering economic, social, educational, and psychological aspects of a family's life, autistic children and their families are mainly of poor socioeconomic status (111). Basically, these families inevitably experience unhealthy, inappropriate sociality and unrehabilitated life conditions because of their financial problems, and occupational and psychological stresses (112-114). The inaccessibility to health care and recreational facilities compound issues and impair physical health (115). Furthermore, exposure to stress and anxiety (such as shared living quarters with other couples or families) imposes continued psychological tension for the parents, especially pregnant mother, thus increasing susceptibility to child autism during pregnancy (67, 93, 94).
Isolation of a mother and breakdown in communications and social interactions negatively affect both the mother and the fetus (116). There are also reportd of existence of a relationship between the parental education and the risk of autism (111, 117-119).
Results in Table 2 shows the prenatal risk factors that may increase the fetal risk of autism. Abnormal gestational age, pre-term (<35 weeks) and post-term pregnancy (>42 weeks), are associated with a significantly increased risk of autism (53, 57, 67, 118, 120, 121). Prenatal risk factors such as bleeding during pregnancy and natal risk factors such as fetal complications including fetal distress, umbilical-cord complications such as fetal nuchal cord and cesarean delivery (26% increased risk of autism) (53) are all involved in hypoxia (lack of oxygen), consequently increasing the susceptibility to autism (53, 59, 70, 121). Fetal nuchal cord occurrence was significantly more frequent among children with autism (23.2%) when compared to controls (6.3%). This complication causes fetal blood deficiency, oxygen, and nutrition. If the inadequate blood flow is severe or long-lasting it would affect fetal brain development and damage the newborn central nervous system (110). Three brain regions, including basal ganglia, hippocampus, and lateral ventricles, are highly known to be highly vulnerable to hypoxia (70). Autistic children's brain exhibit larger lateral ventricles, morphological hippocampal abnormalities, and increased dopaminergic activity, which are all inducible by hypoxia (122).
Postnatal risk factors play a crucial role in susceptibility to autism. A set of such risk factors is included in Table 2. Low birth weight, jaundice, and postnatal infections are some of the most significant risk factors. A neonate with a birth weight, which is determined based on three potential factors (genetic growth potential, duration of pregnancy and rate of fetal growth), less than 2500 g is considered a low birth weight and has a two-fold increased odds ratio of autism (53, 57, 59, 119, 123).
Postnatal jaundice is the result of high bilirubin production caused by increased breakdown of fetal erythrocytes and a low hepatic excretory capacity resulting from general immaturity of the liver. Postnatal jaundice can be associated with death during a sensitive period (around the 40 weeks of pregnancy) or susceptibility to mental disorders, especially a four-fold increase in autism is noted if it survives (53, 57, 67, 124, 125). In addition to prenatal maternal infection during pregnancy, postnatal infections such as meningitis, mumps, varicella, fever of unknown origin, and ear infections (126) during the first 30 days of life are correlated with a high risk of autism (127, 128).
In 1943, the psychiatrist Leo Kanner published a paper describing “children whose condition differed so markedly and uniquely from anything reported so far” (4, p. 217). The author believed that the children suffer from a yet unnamed disorder. The severity varied amongst the cases, but from birth all displayed an “extreme autistic aloneness” (4, p. 242), a powerful desire for sameness, and the ability to interact intelligently with objects, but not with people (129). It is from Kanner and, later psychiatrist, Hans Asperger’s observations that the current classification of ASD arose.
Today, the paths to an ASD diagnosis vary by country, practitioner training, as well as the age of the patient. Children around the world are routinely screened during wellness visits, or in school settings. A list of popular global screening tools is provided in Table 4. A positive diagnosis via screening tool is sufficient in many countries to access treatment services. In the United Kingdom and the United States, diagnosis is made by a developmental pediatrician, psychologist, or psychiatrist using either the International Classification of Diseases Version 10 (ICD-10) (130), or the Diagnostic and Statistical Manual of Mental Disorders, 5th Edition (DSM-5) (131). In the Arab world, the use of ADI-R or ADOS as diagnostic tools, with clinical observations based on DSM-4 and DSM-5 criteria is typical. Amongst the Gulf States, Qatar uses ADOS as the primary diagnostic tool.
| Popular global screening tools | References |
|---|---|
| • Modified Checklist for Autism in Toddlers, Revised (M-CHAT) | (132, 145-147) |
| • Autism Diagnostic Interview-Revised (ADI) | |
| • GARS-2: Gilliam Autism Rating Scale-Second Edition (GARS) | |
| • Childhood Autism Rating Scale (CARS) |
The DSM-5 is considered to be the gold standard in ASD diagnosis and the ICD closely mirrors it. Similar to the children Kanner first described, the DSM-5 classifies autism spectrum disorder based on deficits in social communication, and the presence of restricted, repetitive behaviors, giving each of these two categories a rating between 1-3 based on severity. A rating of 1 is the lowest and indicates that the child has milder symptoms and ‘requires support’ and a rating of 3 indicates that the child’s symptoms are severe and ‘requires very substantial support’. A diagnosis also includes clinical notes and specifies any accompanying intellectual impairments, language impairments, medical or genetic conditions, or environmental factors. The DSM and the ICD, have expanded criteria of diagnosis in subsequent editions; notably, the DSM-5’s reclassification of Asperger’s and pervasive developmental disorder not otherwise specified (PDD-NOS) now falls under a single diagnosis of ASD (131).
While developmental concerns are noted in approximately 85% of children before the age of three, the average age for diagnosis is four (3). Concerns may include lack of eye contact, not meeting developmental milestones, lack of affection and reciprocal social interaction, as well as poor language development. Popular screening tools such as the Modified Checklist for Autism in Toddlers (M-CHAT) (132) are available online for free, and provide an early resource to caregivers when initially noticing concerning behavior, before seeking professional help. A growing awareness of autism has led many adults, who were not diagnosed as children, to seek treatment. In adults, the diagnosis generally relies on self-report or informant questionnaires, observation guides, and clinical interviews (9, 133). Although, early diagnosis remains a challenge. Nevertheless, a team of investigators from the Cleveland Clinic Children’s Center for Autism has developed what may become ASD’s first objective diagnostic aid: an autism risk index based on remote eye-gaze tracking, which is proving to be remarkably accurate (134) (Table 4).
As of 2010, there were an estimated 52 million cases of ASD worldwide. Given the current prevalence estimates of ASD when comorbidities such as learning disabilities (dyslexia), epilepsy and intellectual disability are factored in, the diagnosis rate jumps. The causes of ASD are not clear-cut, nonetheless evidence suggests that a number of environmental and genetic factors are at play (7, 19).
Once a diagnosis of ASD has been established, deficits are treated with a multifaceted team approach. (4)Treatments regularly include occupational, behavioral, speech, and play therapies. There are no pharmacological treatments for ASD, though many individuals receive medications to address comorbidities, such as seizures, and attention deficit hyperactivity disorder (ADHD). In some areas, children may receive support services in schools, including special education programs that target common comorbidities (discussed below) including learning disorders, and intellectual disabilities (4).
The prognosis for an individual with ASD is as heterogeneous as the diagnosis. Those on the mild end of the spectrum, commonly referred to as Aspergers, with average to high IQ are often able to enter mainstream education, progressing into the workplace and through therapeutic support improving skills in social communication. It is not uncommon for individuals at the highest end, many of whom would have previously received a diagnosis of Aspergers, to embrace and capitalize on their atypical neurodevelopment. For example, while an interest in objects, rigid desire for sameness, routine, and detail can hinder social interaction, creativity and flexibility, those same qualities may be very useful in a field that requires a restricted focus on objects, precision, and strict adherence to routine.
For those individuals with a severe diagnosis, prognosis is far less positive. Severe cases may never learn to communicate, and they remain in a world that is withdrawn from peers and family. The focus of treatment for ASD is personalized to the needs of the individual, with a goal to improve quality life of the patient and those around them. Family and caregivers often struggle to relate due to barriers in communicating with those suffering with ASD, and are encouraged frequently as part of therapy to find shared enjoyment in activities that promote bonding.
In addition to traditional therapies, families and the individuals with ASD may seek out support from the community. This can include support through online groups or websites. Autism Speaks (135) is one such popular website, allowing people to connect, access the latest research, and receive moral support. Community and business support for those on the spectrum is becoming more widely available, whether in the form of activities such as playgroups, gymnastics or music classes, that specifically target young ASD individuals, or job programs such as ‘The Microsoft Autism Hiring Program’ (136), which helps qualified ASD adults integrate in the workplace.
The broad criteria for ASD diagnosis leads to a heterogeneous pool of research participants. Lynn Waterhouse in her book, Rethinking Autism (137) proposes the abandonment of the DSM-5 diagnosis of ASD for research. Other researchers recommend a change to the DSM-5 to include more subtypes of ASD (138). Even without a reclassification, one of the largest ASD research projects to date, SPARK by the Simons Foundation (139), narrows the pool of participants through a series of questionnaires, removing those who have known genetic conditions, or epilepsy.
It would seem that abandoning the umbrella classification of ASD would assist research, but the abandonment also creates few obstacles. Due to the prevalence of the current ASD diagnosis, it has become a priority health initiative in many countries. One such country is Qatar where a national plan was launched in April 2017 (19). Widespread public and financial support may not be as readily available if diagnostic criteria changes and the prevalence rates drop. The current umbrella classification of ASD also allows individuals in need to easily qualify for support services, and finally it brings together caregivers and individuals who are dealing with similar phenotypic behaviors and gives them support, regardless of cause (4, 7). National plans put an issue on the policy agenda. In moving forward, the priority is then given to ensure that autistic people, their families and caregivers have access to effective support through the diagnostic process and post-diagnosis, and that requisite educational support is available, as in Qatar’s case (4).
The standard of practice for diagnosis of ASD is first by microarray for detection of large and submicroscopic CNV, followed by WES or WGS. Other genomic techniques such as transcriptomics and epigenomics could also be employed to lead to an enhanced understanding of the molecular mechanisms involved in ASD, and ultimately inform clinical care. Cell-free DNA (cf DNA) testing is becoming an indispensable diagnostic tool. This type of testing or screening is often called noninvasive prenatal testing (NIPT). With next-generation sequencing techniques or single nucleotide polymorphism-based approaches, fetal cf DNA in maternal plasma can be analyzed to screen for testing genetic disorders or related conditions. As stated previously, the use of genetic testing or clinical genomics as an in vitro screen tool, may be a promising way to identify individuals at risk for autism.
Progress in clinical care for ASD will continue to be contingent on multidisciplinary, collaborative research efforts. Efforts are currently underway to identify functional, genetic-ontological subtypes that may provide additional utility with regard to clinical intervention. This includes investigation of broader phenotypes associated with gene disruptions that share molecular properties, for example, genes regulated by the CHD8 protein (140, 141). Identification of measurable neurological effects of gene disruptions, such as an electrophysiological (EEG) signature, could translate to meaningful ASD biomarkers that are essential for clinical treatment trials (142). Meanwhile, development of pharmacotherapies for genetic subtypes, such as SCN2A, will require increased knowledge about the timing of genetic expression and reversibility of neurodevelopmental impairment. Thus, continued comprehensive phenotyping will be essential to the success of clinical trials for genetic subtypes of ASD (140).
Gene therapy seems to hold promise as potential treatment for ASD. Recently, the first preclinical studies in monogenic ASD, involving both gene replacement and silencing have been reported. With increased understanding of ASD etiology, this work can be extended to polygenic ASD (143).
Another possible futuristic treatment is the use of genome editing. We believe the CRISPR-gene editing strategies will soon be increasingly utilized in ASD models to both recapitulate pathophysiology and alleviate neurodevelopmental conditions. For example, a recent study utilized CRISPR-Gold, a novel nanoparticle delivery method, to target striatal mGluR5 levels in FXS mice. In addition to a successful reduction in mGluR5 levels, the study revealed subsequent rescue of repetitive behaviors in FXS mice (144).
The economic impact associated with ASD is substantial, and includes direct medical and non-medical, and indirect productivity costs. Studies estimate the lifetime cost of caring for an individual with ASD to be $2.2 million in the US, and £1.5 million in the UK; though the cost drops to $1.4 million in the US and £0.92 million in the UK for ASD without comorbid conditions. In addition, if unrecognized or untreated, ASD can contribute to poor educational attainment and difficulty with employment, leading to negative economic implications. Recent estimates of the total economic impact of ASD in the US in 2015, based on direct medical, non-medical and productivity costs combined, totaled $268 billion – ranging from 0.9 to 2% of gross domestic product (GDP). This amount is expected to rise to $461 billion (ranging from 0.99 to 3.6% of GDP) by 2025. These costs are on par with recent estimates for ‘silent epidemics’ such as diabetes. Indeed, the burden of ASD appears to exceed the cost of the traditional enemies of health. However, what sets ASD apart from other non-communicable diseases – such as heart disease, cancer, stroke and hypertension – are the significantly higher non-medical costs when compared to direct medical costs. There is also suggestion that comorbidities of ASD tend to amplify burden to the society and afflicted individuals alike (7, 19).
In this review, we offered a summary of ASD clinical diagnostic criteria and provided an insight into ongoing research toward environmental factors, genetic mutations, and neural pathways. We also explained trends in clinical genetic testing. The future for individuals suffering from less severe forms of ASD is positive. It is hoped that through early behavioral screenings, genetic testing, identification of environmental risk factors as well as a better understanding of neural development, the number of individuals suffering from autistic phenotypes may be greatly reduced. While there is much work still to be done in understanding and treating ASD, there are important steps that can be taken now. First, continued awareness programs so that children are identified and treated as early as possible. Second, pre-natal and pre-pregnancy awareness of environmental factors, including recommendations against consanguineous marriages, information regarding optimal maternal nutrition and the importance of limiting exposure to toxins and pollutants. Finally, the expansion of genetic screening, and early post-natal monitoring of infant feeding, nutrition, eye contact, will help to provide the earliest treatment possible.
The authors want to thank their respective institutions for their continued support. The authors declare no conflict of interest.