

Frontiers in Bioscience-Landmark (FBL) is published by IMR Press from Volume 26 Issue 5 (2021). Previous articles were published by another publisher on a subscription basis, and they are hosted by IMR Press on imrpress.com as a courtesy and upon agreement with Frontiers in Bioscience.

1 Department of Applied Biology, Kyoto Institute of Technology, Sakyo-ku, Kyoto 606-8585, Japan
2 Faculty of Agriculture, Bangladesh Jute Research Institute, Manik Mia Ave., Dhaka-1207, Bangladesh
Abstract
In some adult holometabolous insects, specific epithelial tissues form imaginal discs, small sac-like clusters of cells that form in the larval body. During metamorphosis, in response to hormones, the discs undergo dramatic changes including cell proliferation and differentiation. In Bombyx mori insects, the cuticular protein (CP) genes, which are expressed at prepupal stage in the wing discs, are divided into six groups according to their developmental expression profile and responsiveness to steroid hormones. In this article, we discuss the expression of CP genes in the wing disc, and examine the molecular mechanisms by which metamorphosis and cell transformations are regulated by hormones in insects.
Keywords
- Cuticle proteins
- Imaginal disc
- Bombyx mori
- Ecdysone
- Review
The insect epidermis is covered with an exoskeleton made of cuticle layers, which is comprised of the outermost epicuticle, exocuticle, and innermost endocuticle. The exoskeleton is rigid, therefore, insects must shed their exoskeleton in order to grow, a process known as “molting”. The first step of molting is to break the connection between epidermal cells and the endocuticle, called apolysis. After apolysis, the interspace is filled with inactive molting gel-containing enzymes. The epidermal cells are covered with newly-produced epicuticles, followed by activation of molting gel. Old endocuticle is digested by activated molting gel, whereas the epidermal cells are protected by the new epicuticle layer from digestion. Furthermore, epidermal cells produce a new exocuticle layer under the new epicuticle. After digestion is completed, the remnants of the old cuticles are removed (called ecdysis). The epidermal cell layer expands after ecdysis and the cells produce new cuticle layers by secretion of new endocuticle. Finally, insects obtain a new exoskeleton composed of epi-, exo-, and endocuticle layers. The exo- and endocuticle layers are thick, and constructed of chitin polysaccharides and cuticle proteins having an R&R consensus sequence (CPR). The R&R consensus sequence in CPRs can bind to chitin (1, 2). The term R&R consensus, derived from Rebers and Riddiford who identified the sequences, had an original motif of G-x(8)-G-x(6)-Y-x(2)-A-x-E-x-G-F-x(7)-P-x-P (where x represents any amino acid) (3). Three types of CPRs have been identified: RR1 are soft cuticular proteins, RR2 are hard cuticular proteins, and RR3 is composed of a few groups (4-6). Other types of cuticular protein (CP) genes can be grouped according to their characteristics in Bombyx mori and Anopheles gambiae (7-9). Over 200 CP genes have been identified in A. gambiae and B. mori (10-12), with several distinct families of CPs recognized (5). For example, the CPF family comprises four CPs identified with two conserved regions (13, 14). The CPF and CPF-like (CPFL) proteins cannot bind to chitin, thus, it is still unknown whether they form the epicuticle or exocuticle. The CPT family is named from a mutation of body shape, and the glycine-rich CP family is named CPG (15). Additionally, the cuticular protein hypothetical (CPH) family contains putative CPs (12). Thus, it can be said that many different types of CP genes have appeared during insect evolution.
Insect molting and metamorphosis are regulated by two principal hormones; juvenile hormone (JH) and ecdysteroid (ecdysone). The active form of ecdysone, 20-hydroxyecdysone (20E), produced by a P450 enzyme reaction, induces the molting process by binding to a heterodimeric nuclear receptor (EcR/USP). The regulation of ecdysone response genes is explained in the classical Ashburner’s model based on puffing patterns of polytene chromosomes of salivary glands (16, 17). They are generally classified into early response genes, early late genes, and late genes according to their temporal appearance. Genes induced directly by EcR/USP are called early genes: E74, E75, hormone receptor 3 (HR3), and Broad Complex (BR-C). These encode DNA binding proteins that regulate cascades of other gene expression patterns. Following early gene expression, the expression levels of HR3, HR4, and βFTZ-F1 are increased (18). These genes are involved in the molting process induced by 20E. In the presence of JH in hemolymph, ecdysone induces larval molting, molting from larvae to next instar larvae. In other words, a high concentration of JH during the larval period prevents induction of pupal metamorphosis. A decrease in JH in hemolymph after the last larval molt allows ecdysone to induce metamorphosis. JH also regulates transcription factors via its receptor, Methoprene-tolerant (Met) (19, 20). In the absence of JH, a small elevation in ecdysone is required for the reprograming of larval tissues to be ready for metamorphosis (21, 22). This elevation of ecdysone to determine the future characteristics of cells or tissues is called a “commitment peak”. Thus the tissue is committed to a pupa, in which tissues become unable to produce larval characteristics. Commitment ensures the precise development of animals, observed not only in insects, but also in mammals. In the case of mammals, commitment mainly occurs during embryogenesis. Investigating hormonal control of insect commitment could contribute to establishing a general concept of commitment in developmental biology. Insect epidermis or imaginal discs are a good model to identify commitment, because CP genes are stage-specifically regulated by ecdysteroid. Furthermore, the expression of CPs can be observed very easily. We have focused on the wing disc of B. mori, because the developmental profiles of the disc corresponding to ecdysteroid expression are well studied. For better understanding of the wing disc, understanding the regulatory mechanisms of CP gene expression is necessary. Thus, we will further discuss the regulatory mechanisms of CP genes. CP gene mRNAs are abundant, short in size, and transcribed in a short period of time. For instance, CP11 is expressed at very low ecdysone levels at the beginning of the last larval instar, through to CP5, which is expressed after the peak of the ecdysone surge just before pupation. Understanding the expression of CP genes in relation to ecdysone signaling could enable us to gain a better understanding of insect metamorphosis. In order to clarify the relevance of CPs to endocrine signaling during metamorphosis, this review summarizes CP gene expression at the prepupal stage in wing discs of B. mori, according to their developmental expression and ecdysone responsiveness.
Physiological events in each cell and tissue of holometabolous insects are well-orchestrated in an endocrine-dependent manner. An increase in ecdysone in the presence of JH at the end of a larval instar can force the larva to molt into the next instar larva. JH is called the “status quo hormone” because its function is to prevent metamorphosis by binding to its receptor, Met (19, 23). The JH-Met complex induces the expression of transcriptional factor Krüppel homolog 1 (Kr-h1), which suppresses the expression of BR-C, the “initiator of metamorphosis” (20, 24, 25). BR-C is considered an initiator of metamorphosis because mutations produce a phenotype with defective imaginal discs and loss of ability to undergo metamorphosis (26-29). At the beginning of last larval instar, ecdysone synthesis in prothoracic glands are inactivated. This stage-specific absence of ecdysone suppresses JH synthesis in the corpora allata (30, 31). In other words, a stage-specific decrease in JH titer is observed at the beginning of the last larval instar. As a result of the last instar-specific hormonal environment, JH-Met-mediated suppression of BR-C by Kr-h1 maintained is removed.
Expression of some ecdysone “early response genes”, such as BR-C, E75, and HR3, is regulated directly by ecdysone via the EcR receptor, identified as a member of the steroid hormone receptor family in 1991 (32). EcR can form a heterodimer with ultraspiracle protein (USP), a vertebrate retinoid X receptor, which then binds to a specific DNA sequence called the ecdysone response element (EcRE) (32, 33). Orphan nuclear receptor βFTZ-F1 is classified as an early late ecdysone response gene, first isolated from a homeotic mutant of Drosophila melanogaster (34). Expression of βFTZ-F1 is observed after the peak of ecdysone, shortly before larval or pupal molting. As a decline of ecdysone in hemolymph is necessary for normal molting, it is inferred that βFTZ-F1 is a downstream response to the decline in ecdysone in hemolymph. In the prepupal stage of D. melanogaster, induction of βFTZ-F1 enhances the expression of early ecdysone response genes, including BR-C, E74, and E75, responding to the ecdysone pulse just before pupal metamorphosis (35, 36). Premature expression of βFTZ-F1 induces ecdysis defects, while knockdown of βFTZ-F1 prevents molting by suppressing Inka cells to release ecdysis-triggering hormone, emphasizing the importance of βFTZ-F1 as a key molecule in the regulation of molting and metamorphosis (37). Expression of βFTZ-F1 is upregulated by HR3; a Drosophila HR3 (DHR3) mutant showed that DHR3 is essential for the regulation of βFTZ-F1 in the prepupal stage (38, 39). The roles of βFTZ-F1, HR3 and some other ecdysone response genes in the regulation of cuticle genes will be discussed in the next section.
Alterations in holometabolous insect cells during metamorphosis can be generally divided into three types: unnecessary cells after metamorphosis are removed (i.e., larval-specific structures undergo programmed cell death); other tissues, imaginal disc or primordial cells, form pupal or adult structures (e.g., adult appendages of B. mori or D. melanogaster, wings, antenna, and legs); and tissues undergo remodeling of their cell characteristics into pupa or adult. In the case of the Lepidopteran epidermis, parts of it change characteristics to produce the pupal cuticle. Such "actual” alterations of tissues are induced by the ecdysone peak at the end of the final larval instar, but these cells or tissues are determined to undergo metamorphosis before their “actual” alteration. The determination, called “commitment”, takes place without any change in cell appearance. In the case of Manduca sexta, the commitment of epidermal cells is induced by a small elevation in ecdysone in the absence of JH, at the beginning of the last larval instar (21, 22). Wing discs of B. mori are committed on the first day after the last larval molting, becoming competent to receive ecdysone signals and to express pupal CP genes (40).
After the wing discs are committed to metamorphosis, these cells are able to produce CPs depending on the ecdysteroid titer. CP genes are expressed and produced sequentially at the pre-pupal stage to construct pupal cuticle layers. We report here that induction of CP gene expression in the wing disc of B. mori was regulated by the successive appearance of ecdysone response genes, as indicated in Figure 1. Furthermore, we classifiy CPs into six groups according to their developmental expression profiles and ecdysone responsiveness, as described as below briefly.
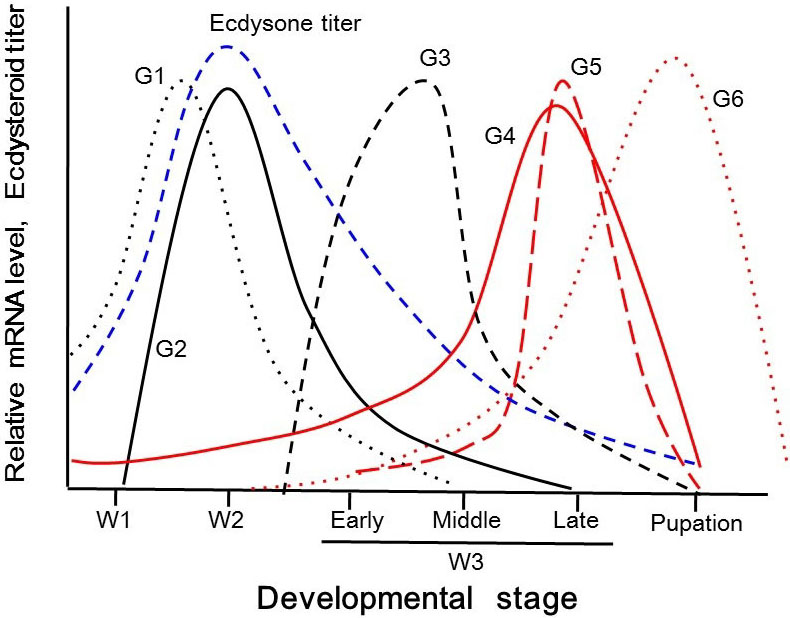 Figure 1
Figure 1Schematic representation of cuticular protein genes and related ecdysone-responsive transcription factors in parentheses expressed in wing discs of B. mori in the late fifth larval instar. Ecdysone titer is represented as blue broken line. G1: Group1 (EcR, BmorCPR21), G2: Group2 (BHR3, BmorCPH5), G3: Group3 (BHR4, BmorCPR34), G4: Group4 (BR-C Z2, BmorCPG11, BmorCPG13 and BmorCPH30), G5: Group5 (E74A, BmorCPR23, BmorCP93) and G6: Group6 (FTZ-F1, BmorCPR45, BmorCPR55, BmorCPR92, BmorCPR99) are represented as black dotted line, black line, black broken line, red line, red broken line and red dotted line, respectively.
Ecdysone titer in hemolymph increases around the first day of the “wandering” stage (W0) in B. mori. (It is customary to set the first day of wandering stage as day 0, the second day as day 1.) Wandering is a last-instar larval-specific action induced by ecdysone in order to look for a place to pupate. The titer reaches a peak on the third day of the wandering stage (W2) in B. mori (41). “Group 1” genes BmorCPR21 and BmorCPR28 are induced directly by 20E, with BmorCPR21 expression increasing from W0 and peaking at W2 (43). “Group 2” only consists of BmorCPH5, reported to be induced by 20E accompanied with the involvement of the “ecdysone early response gene” BHR3 (Bombyx-HR3) (44). The peak expression of Group 2 in vivo is observed a little after the peak of Group 1 (44). Expression patterns of the “Group 3” genes BmorCPR34 and BmorCPH33 resemble the pattern of the ecdysone early late gene BHR4, the transcript levels of which increase early on the fourth day of the wandering stage (W3E) and show a broad expression peak around W3E and late in W3 (W3L) (44-46). Expression of “Group 4” genes (BmorCPG11, BmorCPG13, and BmorCPH30) increases from W0 and reaches a peak on W3L (44, 47, 48). This pattern corresponds to the expression pattern of the BR-C-Z2 isoform. In fact, it was reported that the promoter of BmorCPG11 is primarily regulated by BR-C (48). Expression of “Group 5” CP genes BmorCPR23, BmorCPR93, BmorCPR95, and BmorCPT3 is up-regulated by E74A, peaking at stage W3L (44, 47, 48, 49). Expression pattern of “Group 6” CP genes, BmorCPR45, BmorCPR55, BmorCPR92, and BmorCPR99, in vivo correspond to the pattern of βFTZ-F1, and their levels increase gradually from W2, reaching their highest levels around the peak period of pupal molting (42, 44, 50, 51). Furthermore, Group 6 genes BmorCPT2, CPH1, and CPH2 also show similar expression patterns to βFTZ-F1 (45, 47, 51). In order to consider whether this grouping method according to developmental expression profiles is reasonable to investigate the mechanism of regulation of wing disc development, the next section will discuss the in vitro response of CPs to 20E, and the involvement of ecdysone-related genes.
The responsiveness of CP genes to ecdysone can be examined in vitro and compared with the expression patterns in vivo. We challenged exposed wing discs with an ecdysone pulse in culture medium. Transcripts of Group 3 and Group 4 genes were induced by 6 h and reached their peak at 12h and 18h after the pulse, respectively (46, 48). Group 5 genes were increased 12h after incubation, peaking at 18h (44, 52). All the genes in Group 6 peaked at 24h after the 20E pulse (48, 52). Taken together, Group 3 gene expression peaks first, followed by Group 4, then Group 5 genes, and finally Group 6. Notably, the order of the CP group expression peaks in vitro is consistent with the order of these peaks in vivo (shown in Figure 1). These findings indicate that the responsiveness of each CP group in vitro may be reflected in the developmental regulation of these genes in vivo.
As discussed above, each group of CP genes has a specific developmental expression pattern. In this section, we describe how CP gene expression levels are regulated by ecdysone-response transcription factors. Furthermore, we discuss how the temporal expression of these transcription factors ensures the timing of each CP expression. Ecdysone can regulate gene expression via the EcR/USP receptor complex, which binds to the ecdysone response element (EcRE), an upstream DNA element. Some transcription factors induced by the 20E/EcR/USP complex can indirectly pass on the ecdysone signal to other genes, as described in Section 2. The network of ecdysone-response genes involved in the onset of pupal metamorphosis is well examined in D. melanogaster (54-56). Ecdysone induces DHR3 (Drosophila-HR3) and DHR4 via EcR, which in turn induce expression of E74A and βFTZ-F1. Furthermore, our results indicate that treatment with 20E on wing discs of B. mori is a model for investigating the gene expression network during metamorphosis: ecdysone induces BHR3, the expression of which induces BHR4. BHR4 then in turn induces E74A, BR-C, and βFTZ-F1 (48, see Figure 1).
Group 1 genes are regulated directly by ecdysone. Electrophoretic mobility shift assays (EMSAs) revealed that the EcRE is directly upstream of BmorCPR21 (57). Furthermore, reporter assays indicated that EcR and BR-C Z2 activate the promoters of the Group 1 genes (42, 43, 57, 58). Reporter assays have also revealed that the Group 2 gene BmorCPH5 is regulated by ecdysone in an “indirect” manner through the expression of BHR3. HR3 is an orphan nuclear receptor induced by EcR (44). The developmental expression pattern of BmorCPH5 indicates that it occurs after the expression peak of Group 1 and EcR, as shown in Figure 1. Developmental profiles and reporter assays also suggest that the expression of Group 3 CP genes BmorCPH33 and BmorCPR34 is regulated by BHR4 (44, 59). Group 4 expression is believed to be regulated by BR-C, as the expression profiles of BmorCPG11, BmorCPG13, and BmorCPH30 are similar to the expression profile of BR-C, and a BR-C binding site is located upstream of BmorCPG11 and BmorCPG13 (47, 48, 60) (48 Ref3). The expression of the Group 5 genes can be observed after the wandering stage. Reporter assays suggest that BmorCPR23, BmorCPR93, and BmorCPR95 are directly regulated by E74A (44, 49, 52). Lastly, reporter assays and EMSAs demonstrated that βFTZ-F1 can bind to putative βFTZ-F1 binding sites upstream of Group 6 genes BmorCPR55, BmorCPR92, and BmorCPR99 (42, 48, 50, 51, 52). βFTZ-F1 is key trasnscription factor in the prepupal-to-pupal transition (56, 61, 62), the expression of which is induced by HR3, an early ecdysone response gene. Thus, it is suggested that ecdysone induces BHR3 expression followed by expression of βFTZ-F1, which then induces Group 6 expression. βFTZ-F1 and Group 6 expression levels peak at the end of the larval period.
Taken together, we conclude that the expression patterns of all six groups of CP genes are strictly regulated by ecdysone, either directly or indirectly. The temporal expression pattern of each transcription factor (EcR, HR3, HR4, BR-C, E74A, and βFTZ-F1), involved in ecdysone signaling corresponds to the expression of each group of CP genes, as shown in Figure 1. Thus we can conclude that these molecules ensure the timing of CP gene expression. We further conclude that our grouping of CP genes reflects the physiological characteristics of those genes.
Clarification of insect metamorphosis has been challenging for many decades. Ecdysone and a network of transcription factors induced by ecdysone play principal roles in metamorphosis. In this review, we demonstrate that wing discs can be a good model to examine pupal metamorphosis with the involvement of ecdysone and a cascade of ecdysone response genes. Classification of CP genes into 6 groups according to their responsiveness to ecdysone in vitro proposed by Dr Ali and colleagues reflects physiological characteristics well. The suggested gene groupings may allow for molecular analysis of wing discs during metamorphosis.
It is a question to be addressed in the future whether the expression timing of CP genes contributes to the structure of the cuticle layers. Groups 2, 3, and 4, containing CPH and CPG genes, are thought to be components of the epi- and procuticle layers. However, Group 4 genes are expressed at the beginning of the last larval instar. We consider that the beginning of the last larval instar is too early to produce pupal cuticles, therefore we still do not know why Group 4 must be expressed early in the last instar. Cuticle proteins containing an RR motif (Groups 1, 5, and 6) are expressed around W3L and the pupal stage, and are thought to be a component of the pro-cuticle. It will be a challenge in the future to determine the relationship between temporal expression patterns and position of each group of CPs in the cuticle layers.
This work was partially supported by a grant-in-aid for scientific research from the Ministry of Education, Science, and Culture of Japan (to HK; 16580036, 19380033). We acknowledge Dr. Kawasaki and collaborators in the projects on CPs.
cuticle protein
cuticle proteins containing R& R consensus
Cuticular protein hypothetical family
juvenile hormone; Ecdysone; ecdysteroids;
20-hydroxyecdysone
hormone receptor
Broad complex
ecdysone receptor
ultraspiracle
Methoprene-torelant
Krüppel homolog 1
ecdysone response element
wandering stage
Electrophoretic Mobility Shit Assay