







Frontiers in Bioscience-Landmark (FBL) is published by IMR Press from Volume 26 Issue 5 (2021). Previous articles were published by another publisher on a subscription basis, and they are hosted by IMR Press on imrpress.com as a courtesy and upon agreement with Frontiers in Bioscience.
Retraction published on 25 Dec 2024, see Frontiers in Bioscience-Landmark 2024, 29(12)

1 Department of oncoradiology, Rui’an People’s Hospital and The Third Affiliated Hospital of Wenzhou Medical University, rui’an, Zhejiang, 325200, P. R. China
Abstract
Radiation therapy is a major treatment in hepatocellular carcinoma (HCC). Yet, this treatment is ineffective in HCC due to lack of radiosensitivity. For this reason, we examined whether berberine (BBR) might modify the radioresistance of HCC cells. BBR enhances the radiation-induced oxidative stress and apoptosis in Huh7 and HepG2 cells while it protectes HHL-5 cells from radiation damage. To test the importance of Nrf2, a master transcription factor in oxidative damage, the effect of BBR is studied in irradiated Nrf2-deficient cells. BBR fails to induce the radiosensitivity in Nrf2-deficient cells suggesting that Nrf2 is required for the effect of BBR. BBR suppresses the expression of Nrf2 signaling-related proteins (Nrf2, HO-1 and NQO-1) in Huh7 and HepG2 cells, demonstrating that BBR strengthens radiosensitivity via suppressing Nrf2 signaling pathway in HCC cells. Furthermore, experiment using xenografts in nude mice indicated that BBR enhances the growth inhibitory effect of radiation in a Nrf2-dependent manner in vivo. In conclusion, these results suggest that BBR is a promising potential sensitizer for the radiotherapy of HCC.
Keywords
- Berberine
- Hepatocellular carcinoma
- Radioresistance
- Nuclear factor E2-related factor-2
- Radiosensitivity
Hepatocellular carcinoma (HCC) is the most common malignancy and is considered to be a viral-associated inflammatory transformation disease. HCC causes major cancer deaths (1, 2). To date, surgery combined with radiotherapy and chemotherapy is the mainstream treatment of HCC (3). However, due to the high metastatic rate of cancer cells and tolerance to chemotherapeutic drugs and radiation, the prognosis of patients after treatment is poor (4, 5). Therefore, finding new radiosensitizers to improve the radiosensitivity of liver cancer and its anti-radiation mechanism has become a hot spot in radiation oncology research.
Oxidative stress involves a variety of physiological and pathological processes and plays an important role in the development of tumors (6, 7). Oxidative stress is caused by an imbalance between reactive oxygen species (ROS) and antioxidant enzymes (8). Numerous studies have shown that ROS mediates the occurrence and development of tumors. Radiation therapy can effectively inhibit cell proliferation and induce apoptosis by producing excess ROS in tumor cells (9, 10). However, overexpression of antioxidant enzymes is associated with radiation resistance of tumors. Therefore, preventing the defense of these antioxidant enzymes can increase radiation sensitivity (11, 12). Oxidative stress also plays an important role in acute and chronic liver injury (13).
Oxidative stress in the liver triggers an antioxidant response by activating the Nrf2 pathway (14). It is known that Nrf2 is activated in oxidative stress. Under stress-free conditions, Nrf2 binds to the negative regulator Keap1 to form the Nrf2 / Keap1 complex. Under pressure, Nrf2 dissociates from the Nrf2 / Keap1 complex, and then the activated Nrf2 (p-Nrf2) is transferred to the nucleus and binds to the antioxidant response element (ARE), thereby activating various antioxidant genes (15). Compared with other antioxidant defense systems, Nrf2-mediated antioxidant defense system is the main defense system against oxidative stress, which can activate a variety of downstream cell protection genes, such as activation of GST, NQO-1, HO-1, SOD, GPX and other II-deoxygenases (16, 17) to maintain the homeostasis of the cells. In addition to activating antioxidants and Nrf2, Nrf2 also promotes Bcl-2 expression (18). In addition, recent studies have shown that inhibition of the Nrf2 signaling pathway is sensitive to ionizing radiation and chemotherapeutic drugs (19, 20). In the liver, the Nrf2 pathway is required for the prevention and treatment of liver failure (21, 22). Dushani et al reported that Nrf2 is activated by WithafinA and plays an effective role in reducing liver damage by regulating oxidative stress (23). Therefore, targeting Nrf2 and oxidative stress may be a potential way to explore radiosensitizers.
Berberine (BBR) is a natural alkaloid isolated from the stems and roots of Berberis Vulgaris and Coptis Chinensis. BBR inhibits tumor growth and induces apoptosis in a variety of tumor cells (24-27). In addition, BBR has been reported to have synergistic effects with anti-tumor drugs (28, 29). BBR also enhances radiosensitivity of esophageal squamous cell carcinoma, nasopharyngeal carcinoma, and prostate cancer (30-32). Under oxidative stress, BBR acts as a Nrf2 activator to reduce paclitaxel-induced oxidative stress by reducing lipid peroxidation and glutathione levels in the sciatic nerve (33) and increasing superoxide dismutase (SOD) expression. In addition, BBR can also protect against kidney damage caused by methotrexate by upregulating Nrf2 levels (34). However, the effects of BBR on the radiosensitivity of HCC and its molecular mechanisms have not been reported. Therefore, the purpose of this study is to investigate whether BBR can enhance the radiosensitivity of HCC and explore its related mechanisms.
Human hepatoma cell lines (Huh7, HepG2) and human hepatocyte line (HHL-5) are purchased from ATCC. (Manassas, USA) Cells are grown in DMEM medium containing 10% fetal bovine serum (FBS, LifeTechnologies, Grand Island, USA). All the cells are cultured in 5% CO2 incubator at 37 °C. HHL-5 cells in logarithmic growth phase are inoculated in 96-well plate and incubated with BBR (0, 0.5, 1, 5, 10, 20, 40, 60, 80 and 100 μ M) for 24 h. Huh7 and HepG2 cells are inoculated in 96-well plate and incubated with BBR (0, 10, 20 and 40 μ M) for 24 h, respectively. If necessary, the cells are irradiated with different doses (0, 2, 4 and 6 Gy) of 100KVP X-rays (35) using an X-ray generator (Philips, UK). The irradiation time is 72 h.
Cell viability is evaluated via MTT assay according to the manufacturer’s instructions. (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). The formazan crystals are dissolved in DMSO. Huh7, HepG2, and HHL-5 cells are incubated with 10 μ l MTT solution for 4 h after treated with different doses of BBR. The supernatant of cell culture is removed and MTT crystal is dissolved with dimethyl sulfoxide (DMSO). The absorbance at 570 nm is measured by a spectrophotometer (Bio-Rad Laboratories, Hercules, CA).
Huh7 and HepG2 cells irradiated by 2 Gy X-rays are treated with different doses of BBR (0, 10, 20, 40 μ M) for 24 h. Huh7 and HepG2 cells treated with 10 μ M BBR are irradiated with different doses of X-rays (0, 2, 4, 6 Gy) for 2 h. After 2 h of culture, the fresh medium is used instead of the medium without any drug treatment, and the culture medium is cultured for about 2 weeks. Replace the media every 2 days. Finally, Giemsa staining is used to count the colonies containing more than 50 cells. The data are expressed as a percentage of the comparison
HHL-5 cells, Huh7 cells and HepG2 cells in logarithmic growth phase are cultured in serum-free medium for 24 h, and the degree of cell fusion reached 50%. After irradiation with X-ray (2Gy) for 2 h, the cells are treated with different doses of BBR (0, 10, 20, 40 μ M) for 24 h. The cells are collected and resuscitated with 70% ice-ethanol. The suspension is incubated with propidium iodide (PI) and analyzed by flow cytometry (BD, CA, USA). The number of G0/G1, S, G2/M phase cells is recorded and calculated.
Huh7 and HepG2 cells are treated with BBR or radiation and in combination with radiation. The cells are ished with cold PBS for 3 times and then suspended in AnnexinV binding buffer. The cells are incubated with FITC bound AnnexinV antibody (HeatFisher Company) and PI. Then the samples are analyzed by flow cytometry (BD, CA, USA).
The total proteins of Huh7 and HepG2 cells are prepared by RIPA buffer. The same amount of protein samples is separated by 10% SDS-PAGE and transferred to PVDF membrane (Billerica, Millipore, USA). The membrane is blocked by 5% skim milk and incubated with primary antibodies (Bcl-2, #4223, 1:1000. Bax-6A7, #5023, 1:1000. Nrf2, #12721, 1:1000. LaminB, #12255. HO-1, #82206, 1:1000. NQO1, #62262, 1:1000. β-actin, #4970, 1:1000. Cytochrome c, #4280, 1:1000. Caspase-3, #9665, 1:1000) and corresponding secondary antibody Anti-rabbit IgG. (#7074, Cell Signaling Technology, USA) The targeted band in the membranes is visualized by enhanced chemical luminescence (ECL) and analyzed by ImageJ software.
HHL-5, Huh7, and HepG2 cells are grown in serum-free DMEM supplement with 10 μM DCFH-diacetate (CAS4091-99-0, Sigma) for 1 h. Then, the cells are collected and suspended in PBS and analyzed by flow cytometry. The content of malondialdehyde (MDA) and the activities of glutathione (GSH) and superoxide dismutase (SOD) in cell lysate are determined by a spectrophotometry according to the manufacturer’s instructions (Vazyme, China).
The sequence is described earlier (36). Si-Nrf2 or si-Ctrl are transfected into Huh7 cells with Lipofectamine3000 reagent (Invitrogen Company). After 24 hours of transfection, the cells are collected for further treatment.
All animal experiments are performed in accordance with the NIH Guide for the Care and Use of Laboratory Animals and are approved by The Third Affiliated Hospital of Wenzhou Medical University. Athymic BALB/c mice (nu/nu, 20–25 g body weight) are purchased from the Animal Center of The Third Affiliated Hospital of Wenzhou Medical University (Rui’an, China). Mice are housed in controlled conditions at 25 ± 2 °C, 70% humidity and 12-light-dark periods with and regular sterile chow diet and water. Huh7 cells transfected with si-Nrf2 or si-Ctrl are injected subcutaneously to mice. 9 days after inoculation, the tumor grew to 50 – 100 mm3 and is randomly divided into four groups (n = 6): Control group (solvent group), BBR group (mice are treated with 5 mg/kg/day BBR), Radiation (mice are radiated with 8 Gy X-ray) (32), Radiation plus BBR (mice are treated with 5 mg/kg/day BBR and radiated with 8 Gy X-ray). On the 25th day, the mice are killed and the tumor weight is calculated.
Tumor sections are prepared essentially by standard protocol (30). The sections are incubated with PCNA (sc-56, 1:500, Santa Cruz Biotech, CA, USA) and Ki67 antibodies (sc-23900, 1:500, Santa Cruz Biotech, CA, USA) overnight at 4°C, followed by incubation with fluorophore-conjugated secondary antibody (Invitrogen, Carlsbad, USA) for 1 h. Sections are visualized with a fluorescent microscope and analyzed by ImageJ software.
Data are analyzed by SPSS 19.0 software and the results are expressed as mean ± standard deviation (SD). The statistical significance of the studies is analyzed using ANOVA. The difference is considered significant at P < 0.05.
The antitumor activity of BBR is detected by MTT assay. First, HHL-5 cells are treated with different doses of BBR (0, 10, 20, 40, 60, 80 and 100 μM) for 24 hours. As shown in Figure 1, low doses of BBR (0, 10, 20 and 40 μM) significantly inhibits the growth of Huh7 and HepG2 cells compared to the control, but have no significant effect on the survival of HHL-5 cells. When the dose reaches 60 μM, BBR has certain toxic side effects on normal liver cells and liver cancer cells. Therefore, BBR (0, 10, 20 and 40 μM) is used as the subsequent treatment concentration in this study. (* P < 0.05, **P < 0.01) These results indicate that BBR reduces the viability of HCC cells.
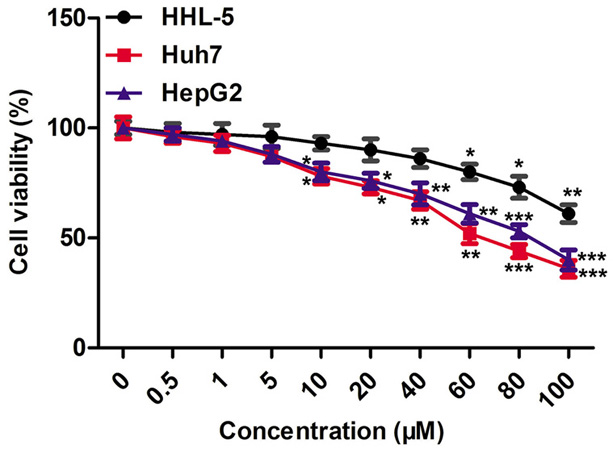 Figure 1.
Figure 1.BBR reduces viability of HCC cells. Huh7 cells are treated with diverse doses of BBR (0, 10, 20, 40 and 60 μM) for 24 h. HepG2 and HHL-5 cells are treated with diverse doses of BBR (0, 10, 20 and 40 μM) for 24 h, cell viability is measured by MTT assay. (A) HHL-5 cells. (B) Huh7 cells. (C) HepG2 cells. All the experiments are repeated at least three times. *P < 0.05, **P < 0.01 versus control.
Giemsa staining is used to explore the relationship between BBR and radiation. After 2 hours of X-ray irradiation, HHL-5, HuH7 and HepG2 cells are treated with different doses of BBR (0, 10, 20 and 40 μM) for 24 hours. The effects of different doses of BBR on the proliferation of HHL-5, HuH7 and HepG2 cells are observed. As shown in Figures 2A-C, BBR enhances the radiosensitivity of Huh7 and HepG2 cells in a dose-dependent manner, but had no significant effect on HHL-5 cells. (* P < 0.05, ** P < 0.01, #P < 0.05) Furthermore, in the presence of BBR, radiation also reduces the cell viability of Huh7 and HepG2 cells in an intensity-dependent manner. However, in HHL-5 cells, BBR reduces the damage of cell survival induced by radiation. (Figure 2D and F, *P < 0.05, **P < 0.01, #P < 0.05) BBR increases the proportion of G0 / G1 phase cells in a dose-dependent manner, but the proportion of G2 / M phase cells does not increase greatly. In HHL-5 cells, BBR significantly inhibits radiation-induced increases in G0/G1 phase cells, suggesting that BBR and radiation mutually promote inhibition of HCC cell survival. (Figure 2G-I, * P < 0.05, ** P < 0.01, #P < 0.05) According to our results, 20 μM BBR and 2 Gy radiation are good choices for further experiments. Taken together, these results indicate that BBR and radiation mutually enhance the inhibition of survival of HCC cells. Videlicet, BBR is a radiosensitizer for HCC cells. According to our results, 20 μM BBR and 2 Gy radiation are better choices for further experiments.
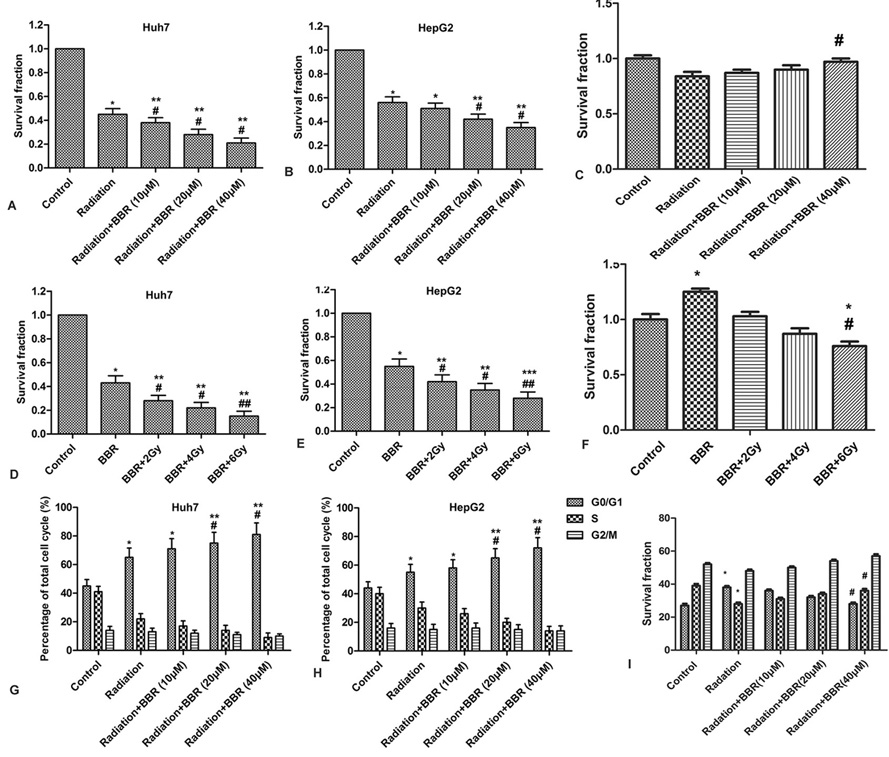 Figure 2.
Figure 2.Synergistic enhancement of BBR and radiation on the survival inhibition of HCC cells. (A-C) Huh7, HepG2 and HHL-5 cells irradiated with X-ray (2 Gy) are treated with diverse dosages of BBR (0, 10, 20 and 40 μM) for 24 h. Cell survival fractions are measured by colony score. *P < 0.05 versus control. #P < 0.05, ##P < 0.01 versus radiation. (D-F) Huh7, HepG2 and HHL-5 cells treated with BBR are irradiated with various doses of X-ray (0, 2, 4, and 6 Gy) for 2 h. Cell survival fractions are measured by colony score. *P < 0.05 versus control. #P < 0.05, ##P < 0.01 versus BBR. (G-I) Huh7, HepG2 and HHL-5 cells irradiated with X-ray (2 Gy) are treated with diverse dosages of BBR (0, 10, 20 and 40 μM) for 24 h. Cell cycle is monitored by flow cytometry. All the experiments are repeated at least three times. *P < 0.05 versus control. #P < 0.05, ##P < 0.01 versus radiation.
In order to investigate whether the apoptosis of Huh7 and HepG2 cells is related to the enhancement of radiation sensitivity, the apoptosis rate of Huh7 and HepG2 cells is detected by flow cytometry and western blot. As shown in Figure 3A, the apoptosis rate of Huh7 and HepG2 cells is significantly increased after 2 hours of X-ray irradiation, and BBR treatment significantly increases the rate of apoptosis induced by radiation. (* P < 0.05, ** P < 0.01, #P < 0.05). In addition, western blot and ELISA show that BBR could further enhance the expression of radiation-induced apoptosis-promoting proteins (Bax-6A7) and caspase-3, and reduce the level of anti-apoptotic protein (Bcl-2). (Figure 3B, * P < 0.05, ** P < 0.01, #P < 0.05) In addition, BBR also reduces the level of cytochrome c in mitochondria, thereby promoting apoptosis. (Figure 3C, * P < 0.05, ** P < 0.01, #P < 0.05) This study demonstrates that BBR enhances the radiosensitivity of Huh7 and HepG2 cells by promoting apoptosis.
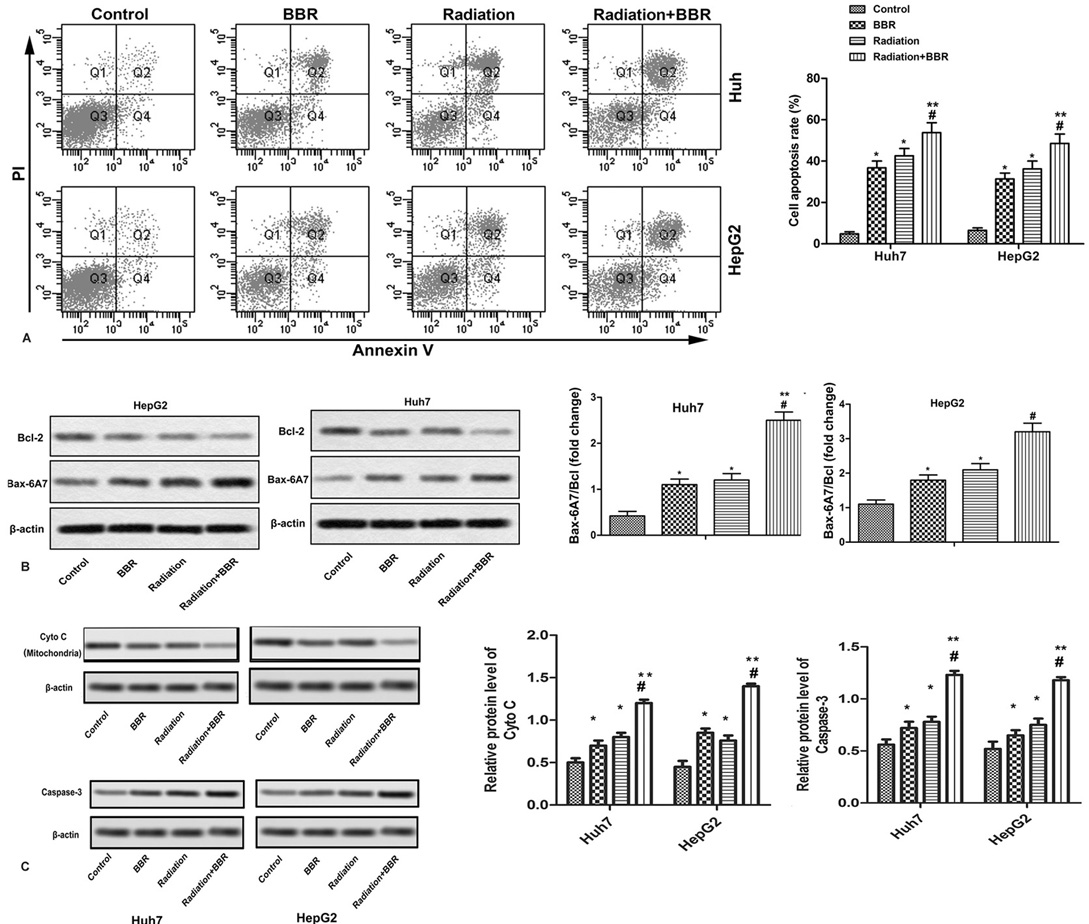 Figure 3.
Figure 3.BBR accelerates radiation-induced apoptosis of Huh7 and HepG2 cells. Huh7 and HepG2 cells are treated with BBR (20 μM) for 24 h or X-ray radiation (2 Gy) for 2 h and in combination with X-ray radiation (2 Gy). (A) Cell apoptosis is analyzed by Annexin V flow cytometry in Huh7 and HepG2 cells. (B) Protein levels of Bax-6A7 and Bcl-2 are detected by Western blot in Huh7 and HepG2 cells. (C) Protein levels of cyto C in mitochondria and caspase-3 in Huh7 and HepG2 cells. All the experiments are repeated at least three times. β-actin is used as loading control. *P < 0.05 versus control, #P < 0.05 versus radiation.
Oxidative stress is the main mechanism of radiation-induced tumor cell death. This study measures the production of ROS by DCFH-DA. As shown in Figures 4A and B, BBR treatment further increased radiation-induced ROS production and MDA accumulation in Huh7 and HepG2 cells. In HHL-5 cells, BBR inhibits radiation-induced ROS production and MDA accumulation (* P < 0.05, **P < 0.01, #P < 0.05) and increases the activities of SOD and GPX in HHL-5, Huh7 and HepG2 cells. (Figure 4C and D, * P < 0.05, ** P < 0.01, #P < 0.05) Therefore, we conclude that BBR promotes the cytotoxicity of radiation to HCC cells by enhancing oxidative stress.
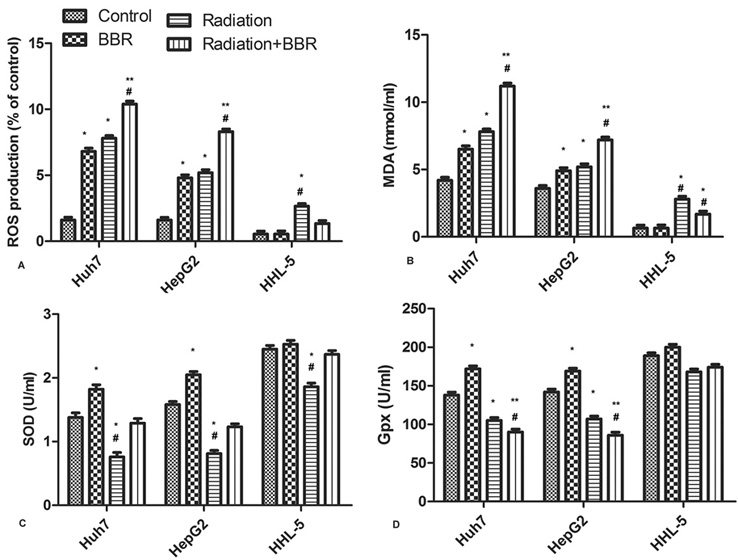 Figure 4.
Figure 4.BBR aggravates radiation-induced oxidative stress in Huh7 and HepG2 cells. Huh7, HepG2 and HHL-5 cells are treated with BBR (20 μM) for 24 h or X-ray radiation (2 Gy) for 2 h and in combination with X-ray radiation (2 Gy). (A) The intracellular ROS is measured with DCFH-DA in Huh7, HepG2 and HHL-5 cells. Data is obtained with flow cytometry. (B) The generation of MDA in Huh7, HepG2 and HHL-5 cells. (C-D) The activities of cellular SOD (C) and Gpx (D) in Huh7, HepG2 and HHL-5 cells. All the experiments are repeated at least three times. *P < 0.05, **P < 0.01 versus control, #P < 0.05 versus radiation.
Nrf2 is a global regulator of cellular oxidative stress, regulating the expression of various detoxifying enzymes, such as NQO-1 and HO-1. Further studies of this mechanism have shown that X-ray irradiation can reduce the levels of Nrf2, HO-1 and NQO-1 in Huh7 and HepG2 cells, and the treatment of BBR further enhances the inhibition of radiation-induced Huh7 and HepG2 cells. In HHL-5 cells, Nrf2, NQO-1 and HO-1 show lower levels, and radiation exposure increases the levels of Nrf2, NQO-1 and HO-1 in HHL-5. BBR treatment inhibites the increase of Nrf2 and other protein levels (Figure 5A-C, *P <0.05, **P <0.01, #P <0.05). These results indicate that the Nrf2 signaling pathway may be associated with BBR-mediated radiosensitivity of liver cancer cells.
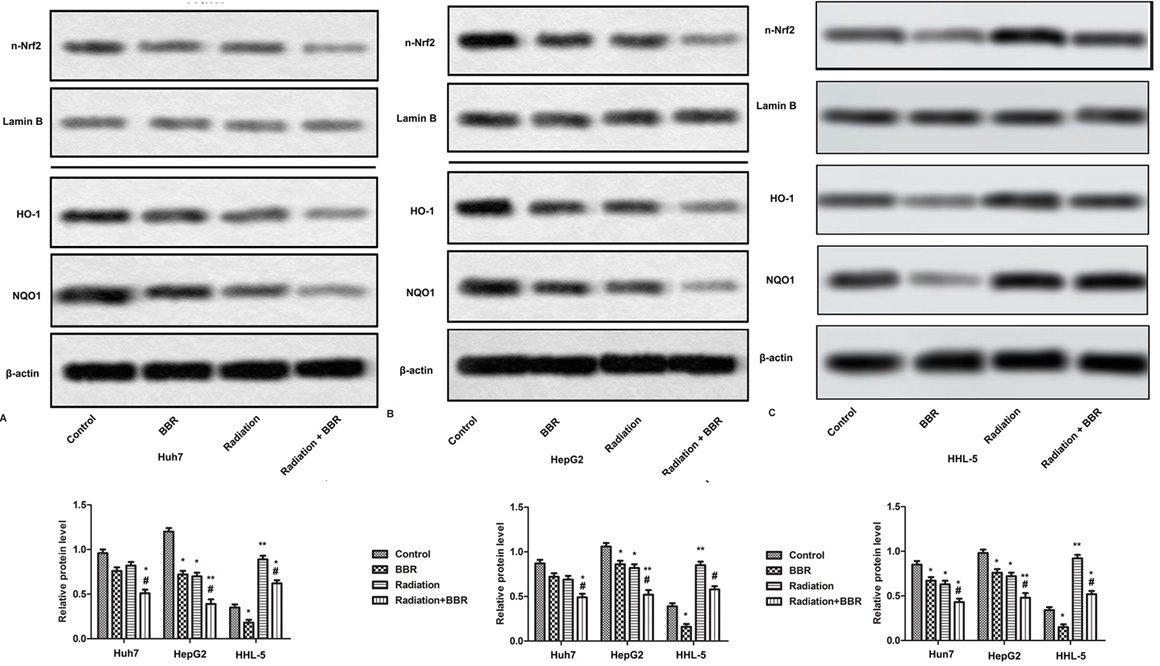 Figure 5.
Figure 5.BBR strengthens radiosensitivity of HCC cells via suppressing Nrf2 signaling pathway. Huh7 and HepG2 cells are treated with BBR (20 μM) for 24 h or X-ray radiation (2 Gy) for 2 h and in combination with X-ray radiation (2 Gy). (A-B) Expressions of nuclear Nrf2, HO-1, NQO1 are measured by Western blot. (C-E) Quantification of Figure 5A-B. All the experiments are repeated at least three times. β-actin and Lamin B are used as loading control. *P < 0.05, **P < 0.01 versus control, #P < 0.05 versus radiation.
To further demonstrate that the Nrf2 signaling pathway is involved in BBR-mediated radiosensitivity, specific Nrf2 siRNA is synthesized and transfected into Huh7 cells. The effect of BBR on Nrf2-deficient cell growth, apoptosis and ROS production is confirmed by Western blotting. As shown in Figures 6A and B, Nrf2 specific siRNA significantly reduces the level of Nrf2 (Figures 6A and B, **P < 0.01). Furthermore, as shown in Figure 6C, in Nrf2-deficient cells, BBR failed to enhance the promotion of ROS production by radiation, and had no effect on cell survival and apoptosis (Figure 6D-E, *P < 0.05, ** P < 0.01, #P < 0.05). These results strongly suggest that Nrf2 is essential for BBR-mediated radiosensitivity of HCC cells.
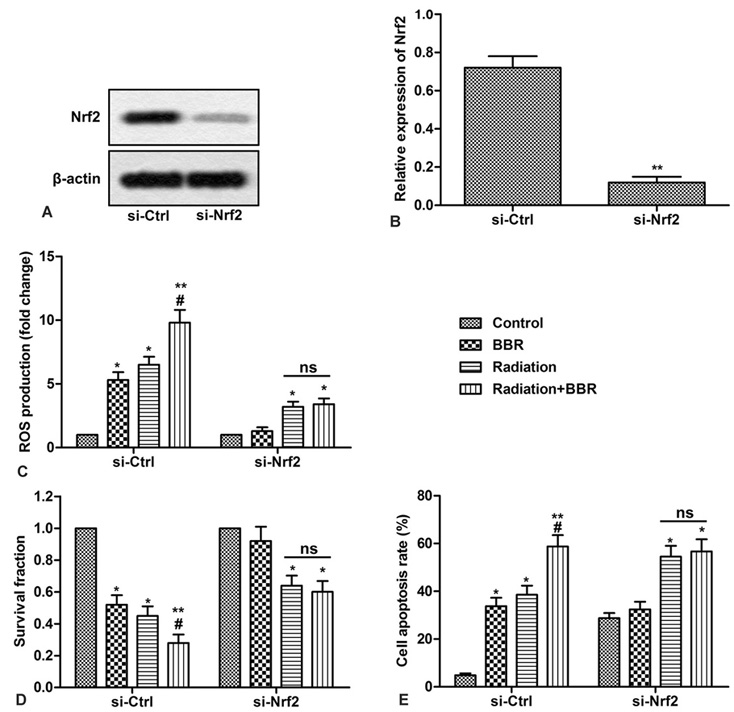 Figure 6.
Figure 6.Nrf2 is indispensable in BBR-mediated radiosensitivity of HCC cells. Huh7 cells are transfected with si-Nrf2 or si-Ctrl. (A) Expression of Nrf2 is detected by Western blot in Huh7 cells. (B) Quantification of Figure 6A. (C) ROS production is measured with DCFH-DA in Huh7 cells. (D) Cell survival fractions are measured by colony score in Huh7 cells. (E) Cell apoptosis is analyzed by Annexin V flow cytometry in Huh7 cells. All the experiments are repeated at least three times. β-actin is used as loading control. *P < 0.05 versus control, #P < 0.05 versus radiation.
In order to study the inhibitory effect of BBR and radiation on the growth of HCC cells, Nhf2 gene-deficient Huh7 cells are used and a Nrf2 gene-deficient liver cancer xenograft model is established. As shown in Figure 7A, BBR significantly reduces the tumor weight in the si-Ctrl group as compared with the single irradiation group, but no significant effect is found in the si-Nrf2 group mice. (* P < 0.05, **P < 0.01, #P < 0.05) Ki67 and PCNA play an important role in cell proliferation. The results of immunohistochemistry show that the expression of Ki67 and PCNA is further decreased after BBR combined irradiation, but there is no significant change in the si-Nrf2 group. (Figure 7B-D, * P < 0.05, ** P < 0.01, #P < 0.05) These results indicate that BBR Nrf2-dependently enhances growth inhibition of radiation in vivo.
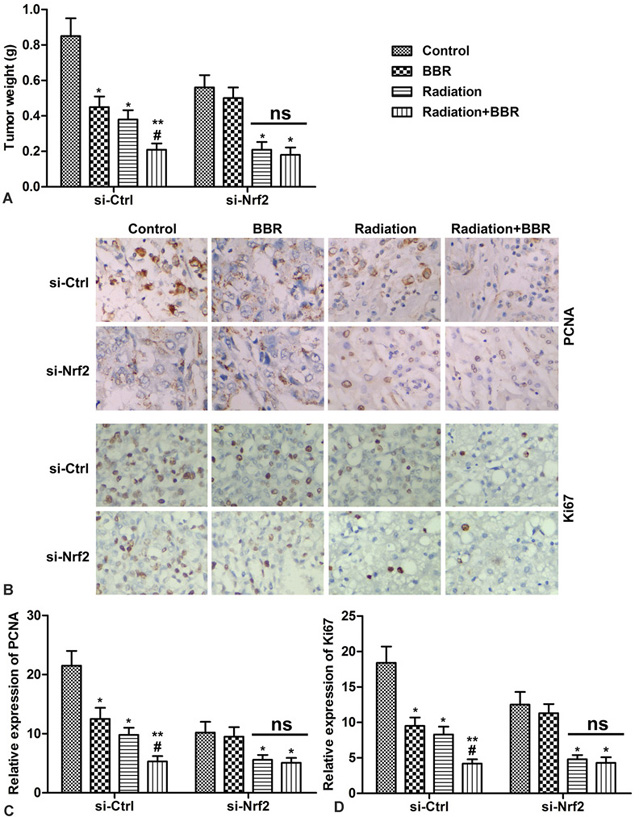 Figure 7.
Figure 7.BBR Nrf2-dependently enhances the growth inhibitory effect of radiation in vivo. Huh7 cells transfected with si-Nrf2 or si-Ctrl are subcutaneously injected into mice to form tumors. After successful modeling, mice are divided into four groups (n=6): Control (mice are treated with vehicle), BBR (mice are treated with 5 mg/kg/day BBR), Radiation (mice are radiated with 8 Gy X-ray), Radiation plus BBR (mice are treated with 5 mg/kg/day BBR and radiated with 8 Gy X-ray). (A) Tumor weight at 25 days after injection. (B) Expressions of Ki67 and PCNA in tumor tissues are detected by immunohistochemistry. (C-D) Quantification of Figure 7B. *P < 0.05, **P < 0.01 versus control, #P < 0.05 versus radiation.
BBR is a widely used antibacterial and anti-inflammatory drug that exhibits anti-tumor activity in a variety of cancers (19, 20). More importantly, recent studies have reported that BBR enhances the radiosensitivity of nasopharyngeal carcinoma, esophageal squamous cell carcinoma and prostate cancer by inhibiting the expression of hypoxia-inducible factor-1a (30-32). In this study, we have demonstrated that BBR can reduce the survival rate of HCC cells and provide evidence that BBR enhances the radiosensitivity of HCC cells in vitro and in vivo. These results are consistent with previous reports (25).
Radiation therapy is one of the main methods of treating various cancers. However, radiation resistance is a major obstacle to radiation therapy applications. Fortunately, numerous studies have shown that oxidation plays an important role in radiation resistance (37, 38). Radiation therapy and chemotherapy can induce intracellular DNA damage and repair, cell cycle arrest, apoptosis, signal transduction and oxidative stress by producing reactive oxygen species (Ros), thereby effectively killing tumor cells (39, 40). However, some cancer cells with strong antioxidant capacity can escape radiation damage through overexpression of antioxidant enzymes and clearance of ROS, leading to radiation resistance (41). Studies have shown that blocking these oxidative defense systems increases the sensitivity of cancer cells to radiation and chemotherapy drugs (42). In this study, we found that low doses of BBR inhibited HCC cell survival without a significant effect on HHL-5 cell viability. Different doses of X-rays (0, 2, 4, 6 Gy) reduced the survival of Huh7, HepG2 and HHL-5 cells in a dose-dependent manner. However, low-dose BBR enhanced X-ray inhibition of Huh7 and HepG2 cell survival in a dose-dependent manner while protecting HHL-5 cells from radiation damage. Generally, this will greatly improve the radiosensitivity of hepatoma cells.
Nrf2 is a key regulator of redox homeostasis and encodes a variety of cytoprotective enzymes (43). Under alkaline conditions, Nrf2 is separated in the cytoplasm by the inhibitor Kelch-like ECH-related protein 1 (Keap1) and is degraded by the ubiquitin-proteasome pathway. When the cells are exposed to oxidative stress or other deleterious attack, Nrf2 is activated and enters the nucleus and binds to the antioxidant response element (ARE) (44). Nrf2 is abnormally expressed in pancreatic cancer, ovarian cancer and other tumors (44, 45). Therefore, the Nrf2 signaling pathway plays an important role in tumor progression and anti-radiation. Previous studies have reported that down-regulation of Nrf2 expression by shRNA increases the radiosensitivity of cancer cells (46). Furthermore, the inhibition of Nrf2 overcomes the radiation resistance of nasopharyngeal carcinoma cells (47). As reported by Matsuoka et al, IL-6 inhibits oxidative stress in oral squamous cell carcinoma (48) via the Nrf2-antioxidation pathway, thereby reducing radiation resistance (48). In addition, IM3829 increases the radiosensitivity of NSCLC by blocking Nrf2-dependent antioxidant responses (20). In the liver, it is found that isoglycyrrhizin (ISL) combined with X-ray irradiation significantly increased the apoptosis of HepG2 cells and inhibited the growth of HepG2 cells by down-regulating the expression of Nrf2 (49). In addition, it has been reported that Wingless / int-3A (Wnt3a) inhibitor (LGK-974) enhances the inhibition of HepG2 cell apoptosis and growth by modulating the Nrf2 signaling pathway (35). Consistent with these results, current studies indicate that Nrf2 exhibits high expression levels in Huh7 and HepG2 cells, and high Nrf2 levels are inhibited by radiation. BBR treatment significantly enhanced the inhibition of Nrf2 expression by radiation. In addition, the reduction of Nrf2 further reduced the levels of HO-1 and NQO-1, blocked the antioxidant pathway, and accelerated apoptosis. However, in HHL-5 cells, Nrf2 showed low levels of expression, and low doses of BBR further reduced levels of Nrf2. Instead, the radiation promoted the expression of Nrf2. Interestingly, BBR, on the other hand, inhibited the promotion of Nrf2 expression by radiation. It appears that in HHL-5 cells, Nrf2 is fixed by Keap1 or degraded by ubiquitin-proteasome, whereas in cancer cells (Huh7 and HepG2), ROS stimulates Nrf2 to dissociate from Keap1, transfer to the nucleus, interact with ARE, and promotes the expression of antioxidant proteins (HO-1 and NQO-1). In summary, the Nrf2 pathway is involved in the role of BBR and radiation in HCC.
In conclusion, this study reports for the first time that BBR enhances the radiosensitivity of HCC cells by regulating Nrf2 signaling pathway and ROS production. However, attention should be paid to some of the limitations of this study. First, as a positive control, we did not provide a good adjuvant for HCC radiation therapy. Therefore, it is necessary to further confirm the relative effectiveness of BBR radiation against liver cancer. Second, the mechanism by which BBR inhibits the Nrf2 signaling pathway in HCC cells is unclear. However, our research still provides a new perspective for the application of BBR in radiation therapy, and describes the new mechanism of BBR radiosensitization in HCC.
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Abbreviations: HCC, Hepatocellular carcinoma; BBR, Berberine; ROS, Reactive oxygen species; Nrf2, Nuclear factor E2-related factor-2; HO-1, Heme oxygenase-1; NQO1, NAD(P)H quinine oxidoreductase 1; MDA, Malonaldehyde; SOD, Superoxide dismutase; GPx, Glutathione peroxidase; Bcl-2, B-cell lymphoma-2; Bax, Bcl-2-associated X protein; Keap1, Kelch-like ECH-associated protein1; ARE, Anti-oxidant responsive elements; PCNA, Proliferating cell nuclear antigen.