



 , Natalia Belkova 1,†, Anna Gorkavenko 1, Nadezhda Smurova 1, Raisa Zugeeva 1, Antonina Morozova 2, Natalya Golovnyova 2, Natalya Semenova 3, Lyubov Rychkova 4
, Natalia Belkova 1,†, Anna Gorkavenko 1, Nadezhda Smurova 1, Raisa Zugeeva 1, Antonina Morozova 2, Natalya Golovnyova 2, Natalya Semenova 3, Lyubov Rychkova 41 Institute of Epidemiology and Microbiology, Federal State Public Scientific Institution “Scientific Center for Family Health and Human Reproduction Problems”, 664003 Irkutsk, Russia
2 Laboratory of Lactic Acid and Bifidobacteria, State Scientific Institution “Institute of Microbiology of the National Academy of Sciences of Belarus”, 220084 Minsk, Belarus
3 Personalized and Preventive Medicine Department, Federal State Public Scientific Institution “Scientific Center for Family Health and Human Reproduction Problems”, 664003 Irkutsk, Russia
4 Pediatrics Department, Federal State Public Scientific Institution “Scientific Center for Family Health and Human Reproduction Problems”, 664003 Irkutsk, Russia
†These authors contributed equally.
Abstract
Lactobacillus paragasseri strains are widely used as probiotics. According to modern requirements, it is necessary to characterize their morphological, cultural, physiological-biochemical, and antagonistic properties, as well as to determine their virulence, toxigenicity, toxicity, and secondary metabolite production, to study their potential and safety. The main aim of the study was to determine the probiotic potential of the L. paragasseri lg94 strain based on its biochemical features and genomic landscape.
L. paragasseri lg94 was isolated from human feces. The genus and species were determined by mass spectrometry using the MALDI Biotyper. The main cultural, biochemical, and genomic characteristics of L. paragasseri lg94 were established. The biochemical profile and L-lactic acid content of the strain were determined. Additionally, the strain’s resistance to bile and low pH stress, as well as its antibiotic susceptibility, were assessed. The genomic sequence of strain lg94 was obtained using Illumina NextSeq550 sequencing.
In this study, the strain lg94 isolated from the feces of a healthy child was identified as L. paragasseri. After 24 h of cultivation in API 50 CH/CHL medium, strain lg94 exhibited positive results in the fermentation of 25 carbohydrates. Strain lg94 exhibited intense L-lactic acid production, reaching a concentration of 6.41 g/L in 24 h of cultivation, indicating high acid-forming capacity. The results showed that L. paragasseri lg94 demonstrated low resistance to acid stress (pH 2.0) but high resistance to 0.3% bile, with no changes in colony morphology observed. The disk diffusion assay revealed that L. paragasseri lg94 was phenotypically susceptible to several antibiotics, including ampicillin, amoxicillin, penicillin G, tetracycline, erythromycin, and chloramphenicol, with intermediate resistance to oxacillin and resistance to fosfomycin, gentamicin, and clindamycin. The whole-genome of L. paragasseri lg94 was sequenced. The absence of antibiotic resistance, pathogenicity, and virulence determinants in the genome was confirmed. Strain lg94 has genes for the production of gassericin T, enterolysin A, and helveticin J.
The results of the studies showed that L. paragasseri lg94 has probiotic potential, and it is recommended to continue studying its antagonistic activity and secondary metabolite production.
Keywords
- Lactobacillus paragasseri
- probiotic
- genome
- lactic acid
- bioinformatics
Recent and ongoing advances in research on the gut microbiome have shown that gut microbiota compounds are a robust, yet understudied, source of probiotic research [1, 2]. There is growing awareness and broader acceptance of probiotic supplements, especially in light of these recent findings. Therefore, academic and industrial efforts are focused on identifying and characterizing new microbial strains to develop effective probiotics [3].
We have accumulated extensive experience in studying the human gut microbiota and its relationship with various health disorders [4, 5]. Hence, our focus is on treatments to address dysbiotic disorders, primarily through research on probiotics [6]. The isolation of probiotic strains from healthy people facilitates the creation of innovative multicomponent probiotics via improved methods. Previous research has shown that the biocompatibility of candidate strains and a potential synergistic effect, marked by increased antagonistic activity from the consortium compared to single strains, are crucial for the effective formation of a probiotic consortium [7, 8].
Lactobacillus paragasseri strains are widely used in probiotic selection and formulation. The positive effects of L. paragasseri are strain-specific and can be used to correct various diseases. For example, the probiotic strain L. paragasseri K7, isolated from the gut of an infant, maintains intestinal homeostasis, participates in the regulation of the immune system, reduces allergic symptoms, prevents bacterial and viral infections, and helps alleviate infectious diseases [9]. It has been experimentally proven that taking L. paragasseri OLL2809 helps increase the content of commensal Akkermansia muciniphila, Bifidobacterium, and Lactobacillus in the intestinal microbiota and reduces depressive behavior in mice [10]. A placebo-controlled study found that long-term use of a drug based on the L. paragasseri OLL2809 strain had a positive effect on menstrual-related symptoms [11]. The L. paragasseri LPG-9 strain has gastroprotective properties and the ability to inhibit the growth of Helicobacter pylori in the stomach of mice [12]. Another probiotic effect of L. paragasseri was demonstrated by Kim et al. [13], who noted a decrease in white adipose tissue and normalization of the blood lipid profile after adding L. paragasseri LG2055 lyophilisate to the diet of obese mice. In addition, the mechanisms for increasing lipid excretion by the L. paragasseri LG2055 strain were determined due to the regulation of mRNA of proteins involved in lipid metabolism [13].
Nowadays, no safety group has been established for L. paragasseri. However, according to the classification of the Canadian Food Inspection Agency, L. gasseri is a sister taxon of L. paragasseri [14] and belongs to risk group 1, meaning that this species is not a pathogen for terrestrial animals, including humans. This suggests that the safety information for L. gasseri may be applicable to L. paragasseri. However, in 2021, the first case of cavernous abscess caused by L. paragasseri was described [15]. Therefore, the safety of strains of this species still needs to be confirmed.
According to modern requirements, pre-registration preclinical safety studies of potential probiotic strains of microorganisms require characterization of physiological, biochemical, and antagonistic properties, determination of virulence, toxigenicity, and toxicity [16]. The taxonomic affiliation of strains must be established using molecular methods, and the presence of antibiotic resistance genes, plasmid material, and prophages had to be studied [17].
The complete 16S ribosomal RNA gene sequence of L. gasseri and L. paragasseri differs only in two single-nucleotide polymorphisms [18]. Whole-genome sequence analysis can reliably distinguish between two species [18, 19, 20]. Moreover, distinguishing between L. paragasseri and L. gasseri species using matrix-assisted laser desorption ionization (MALDI) is difficult, whereas biochemical identification allows for the accurate differentiation of these two related species [14, 15].
Currently whole-genome sequencing allows to distinguish taxonomy between closed species and establish of the absence of the required gene determinants (plasmid material and prophages). The metabolic potential of strains can also be predicted, possible mechanisms of interaction with the host organism can be determined, and probiotic effects can be substantiated using this approach.
Studying the biochemical properties of microbial strains is important for identification and evaluation of their probiotic and biotechnological potential. Despite the published data on the phenotypic properties of L. paragasseri strains [14], the ability to use different substrates may vary. Therefore, it is necessary to establish metabolic features for candidate probiotics.
Therefore, this study aimed to determine the probiotic potential of the L. paragasseri lg94 strain based on its biochemical features and genomic landscape.
L. paragasseri lg94 strain was isolated from the human feces. Cultivation was performed in MRS lactobacillus broth (HiMedia, Mumbai, Maharashtra, India) at 37 °C for 48 h. Identification was performed using a bacteriological method that assessed the morphological, cultural, tinctorial, and biochemical properties of the isolates, with confirmation provided via MALDI-TOF direct protein profiling [21]. The strain genus and species were determined by mass spectrometry using the MALDI Biotyper system. In brief, isolates were grown at 37 °C for 24 h after isolation of pure cultures. Protein extraction was performed according to the manufacturer’s recommended standard operating protocol. A Microflex instrument (Bruker, Billerica, MA, USA) was used for mass spectrometric analysis. Spectra were collected manually or automatically. The server interface (version 4.3.1) and server module (version 4.3.18) were used for identification.
The biochemical profile of the L. paragasseri lg94 strain was studied using API 50 CH/CHL kits (Biomerieux, Marcy-l’Étoile, France) according to the manufacturer’s instructions, and the plates were incubated at 37 °C. The results were visually assessed by the change in well color after 24, 48 and 72 h of incubation.
A 24-h culture was used for the inoculum, and the inoculum cell titer was 1.8
| Component of the bacterial medium | Concentration, per 1 L |
| Glucose/lactose | 10 g |
| Yeast extract | 5 g |
| Peptone | 10 g |
| Potassium phosphate trihydrate | 4 g |
| Magnesium sulfate heptahydrate | 0.2 g |
| Ammonium citrate | 5 g |
| Manganese sulfate | 0.05 g |
| Meat-peptone broth | 100 mL |
| Distilled water | Up to 1000 mL |
We determined the concentration of L-lactic acid in the culture fluid by photometric method using the commercial Lactate-Vital kit (Vital Development Corporation, Saint Petersburg, Russia), according to the manufacturer’s instructions. To ensure the linearity and accuracy of measurements, serial dilutions of the culture fluid were made with sterile distilled water in the following ratios: 1:50, 1:100, and 1:200. A solution of L-lactic acid with a concentration of 2 mmol/L, included in the commercial Lactate-Vital kit, was used as a positive standard. We determined the concentration of L-lactic acid by measuring the optical density at a wavelength of 540 nm using an F300TP photometer (JSC Vityaz, Vitebsk, Republic of Belarus).
The concentration of L-lactic acid (C, mmol/L) was calculated using the following formula:
• EL—the value of the optical density of the culture liquid.
• EP—the value of the optical density of the positive standard.
To ensure the accuracy of the calculations, the optical density of the working solution of Lactate-Vital was measured, and it value was subtracted from the optical density values of all other samples resulting the EL value. The obtained concentration of L-lactic acid in mmol/L was converted to g/L according to the formula:
• D—dilution factor of culture fluid (50, 100, 200 or 1).
• ML-lactic acid = 90 g/mol.
All calculations were performed for each dilution, and the average value between all dilutions was calculated.
The L. paragasseri lg94 strain was grown in liquid MRS medium with lactose/glucose and sterile milk at 37 °C for 24 h. Table 1 shows the medium composition. Sterile milk was obtained from sterilization of ultra-pasteurized commercial milk with 3.2% fat content. Titratable acidity was determined by titration of the culture liquid with 0.1 N NaOH solution in the presence of 1% phenolphthalein solution as an indicator.
A second-generation L. paragasseri lg94 culture was used for inoculum preparation. 5 mL of the L. paragasseri lg94 culture were inoculated into 100 mL of liquid MRS medium, resulting in a 5% (v/v) inoculum. Cultivation was performed at 37 °C for 24 h under thermostatically controlled conditions. Following incubation, the culture was centrifuged at 8000 rpm for 10 min at 10 °C to collect the bacterial cells. The supernatant was discarded, and the cell pellet was washed twice with 100 mL of sterile 0.9% (w/v) NaCl solution. Finally, the pellet was resuspended in 10 mL of sterile 0.9% (w/v) NaCl solution. Stress exposure was tested under the following conditions:
• Control: MRS liquid medium.
• Low pH: Britton-Robinson buffer, pH 2.0.
• Bile: Britton-Robinson buffer containing 0.3% bile, pH 7.0.
In the experiment, 1.5 mL of concentrated L. paragasseri lg94 biomass was added to 8.5 mL of the corresponding solution, thoroughly mixed, and incubated at 37 °C for 2 h. The viable cell count (CFU/mL) was determined by serial dilution in tubes containing semi-solid MRS medium, followed by incubation at 37 °C for 48 h. CFU/mL values were recorded before and after exposure to the respective stress factors (low pH and bile).
The in vitro susceptibility of the strain to antibiotics was determined
using the disk diffusion method [22]. A 24-h culture was grown on MRS agar and
suspended in 0.9% NaCl until a turbidity of 0.5 according to McFarland (1.5
Antagonist activity was assayed using two methods: the perpendicular streak method and the well method, each in triplicate.
Test cultures included the reference strains Escherichia coli ATCC 25922 and Pseudomonas aeruginosa ATCC 27853, as well as clinical isolates with confirmed multidrug resistance (MDR) obtained from the collection of Laboratory of Microbiome and Microecology (Scientific Center for Family Health and Human Reproduction Problems, Irkutsk, Russia). They were E. coli 235-LMM, P. aeruginosa ODKB-58, Klebsiella pneumoniae ODKB-81 and K. pneumoniae ODKB-82.
For the well method, the strain was grown in MRS broth (37 °C, 24 h)
under anaerobically maintained conditions. Anaerobiosis was achieved using an
Anoxomat atmospheric generation system (Advanced Instruments, Inc., Norwood, MA,
USA): sealed anaerostats containing culture tubes were filled with a gas mixture
(80% N2, 10% H2, 10% CO2). The optical density of the culture
at 600 nm (OD600) was measured in triplicate using a Nano-500 spectrophotometer
(Allsheng Instruments Co., Ltd., Hangzhou, China) with sterile MRS broth as the
reference medium. The average OD600 value was 2.42
For the perpendicular streak method, the strain lg94 was streaked onto Bifidum agar medium (State Research Center of Applied Microbiology and Biotechnology, Obolensk, Russia) and cultured in an anaerobic jar at 37 °C for 48 h. After incubation, an exponential culture of the test strains was streaked perpendicular to the L. paragasseri lg94 growth line. Further incubation was carried out under aerobic conditions (37 °C, 24 h) to ensure the growth of the test strains.
The sizes of the inhibition zones were measured in millimeters (mean
The 24 h cell culture of L. paragasseri lg94 was used for DNA
extraction. Genomic DNA was isolated using the Quick-DNA Fungal/Bacterial
Miniprep Kit (Zymo Research, Irvine, CA, USA). DNA quantity was estimated using a
Qubit 4 fluorimeter (Thermo Fisher Scientific, Waltham, MA, USA) with the Qubit
ds-DNA HS Assay Kit (Invitrogen, Waltham, MA, USA). DNA concentration
The nucleotide sequence of the L. paragasseri lg94 genome was obtained by sequencing on the Illumina NextSeq550 platform (Illumina, San Diego, CA, USA) using the Illumina® DNA Prep Tagmentation, IDT® for Illumina® DNA/RNA UD Indexes Set Tagmentation, and NextSeq 500/550 High Output Kit v2.5 (300 Cycles) library preparation reagents according to the manufacturer’s recommendations (Illumina, San Diego, CA, USA).
The genome sequence was obtained by aligning the obtained reads to the reference genome of Lactobacillus paragasseri JCM 5343 (NCBI RefSeq assembly GCF_003584685.1) using Bowtie2 v.2.5.4 [23] (Langmead Lab, Baltimore, MD, USA). MOB-suite v3.0.3 [24] (Public Health Agency of Canada, Guelph, ON, Canada) was used to search and identify plasmid sequences. Annotation was performed using Prokka v1.14.6 [25] (Victorian Bioinformatics Consortium, Melbourne, Victoria, Australia) and Prodigal v2.6.3 [26] (Oak Ridge National Laboratory, Oak Ridge, TN, USA). The search for prophages and CRISPR-Cas loci was performed using PHASTEST v1.0.1 [27] (University of Alberta, Edmonton, Alberta, Canada) and CRISPRCasFinder v4.2.20 [28] (Institut Pasteur, Paris, France). The search for gene functions was performed using BlastKOALA v3.0 [28] (Kanehisa Laboratories, Tokyo, Japan). KEGG Mapper v5.0 [29, 30] (Kanehisa Laboratories, Tokyo, Japan) was used to analyze potentially implemented metabolic pathways. The search for antibiotic resistance and disinfectant resistance genes was performed using the ResFinder v2.4.0 and DisinFinder v2.0.1 databases [31, 32] (Technical University of Denmark, Copenhagen, Denmark). The online tool MiGA Web v2.0 (Georgia Institute of Technology, Atlanta, GA, USA) was used to confirm the identification of the strain under study [33]. antiSMASH v8.0 (Technical University of Denmark, Copenhagen, Denmark) was used to search for secondary metabolism genes [34]. The sequencing results are presented in the NCBI database in PRJNA1248600 as sample SAMN47864912.
Strain lg94 was isolated from the feces of a healthy child in the Laboratory of Biomedical Microecology of the Scientific Center for Family Health and Human Reproduction Problems (Irkutsk, Russian Federation) on MRS medium. The strain was a facultative anaerobe; it grew well under conditions with low oxygen content both in test tubes on liquid nutrient media and in conditions of complete absence of oxygen: on agar media in an anaerobic jar with gas-generating bags.
Optimum cultivation conditions were determined in the temperature range of
25–41 °C with a step of 4 °C and pH range of 5.5–7.0 with a
step of 0.5. The optimal temperature for the growth of strain lg94 was determined
to be 37 °C and pH 6.5, at which the strain reached a concentration of
live cells of 2
L. paragasseri lg94 is gram-positive, non-motile, and does not form spores. The cells are rod-shaped, short with rounded ends, located singly, in pairs, or in short chains. When grown on MRS medium, growth was observed in the form of broth turbidity and precipitation of a white-gray sediment at the bottom of the test tube. L. paragasseri lg94 cells formed small, mucous, and rounded white colonies on MRS agar.
For strain lg94, biochemical profile tests were conducted, as well as the quantitative determination of L-lactic acid content and full-genome sequencing, followed by genome assembly and gene landscape analysis.
The study of carbohydrate metabolism of the L. paragasseri lg94 strain using the API 50 CH/CHL test revealed pronounced saccharolytic activity, which was characterized by different carbohydrate fermentation rates depending on the incubation duration. Data on the biochemical profile of the strain are presented in the Supplementary Table 1.
After 24 h of cultivation in API 50 CH/CHL medium, L. paragasseri lg94
showed positive results regarding the fermentation of 25 carbohydrates
(Supplementary Table 1). In particular, the ability to utilize
monosaccharides (D-ribose, D-galactose, D-glucose, D-fructose, D-mannose, and
L-sorbose), sugar alcohols (D-mannitol, D-sorbitol),
The subsequent increase in incubation time to 48 h resulted in an expansion of the spectrum of fermentable carbohydrates. L. paragasseri lg94 demonstrated the ability to utilize L-arabinose, D-melibiose, and gentiobiose. Further incubation for up to 72 h revealed inositol, D-lyxose, and potassium gluconate fermentation.
L. paragasseri lg94 exhibited intense L-lactic acid production, reaching a concentration of 6.41 g/L after 24 h of cultivation, indicating the high acid-forming capacity of this strain (Table 2).
| Sample variant | Eraw | EL | C, mmol/L | C, g/L | C, g/L |
| 1:50 | 0.27 | 0.18 | 1.241 | 5.584 | 6.41 |
| 1:100 | 0.19 | 0.10 | 0.69 | 6.21 | |
| 1:200 | 0.15 | 0.06 | 0.41 | 7.452 |
Based on their acid-producing capacity, the strains were classified into two
groups. Strains producing 1–2 g/L of acid in the culture medium were defined as
moderate acid producers, whereas strains producing
The optical density of the culture fluid was calculated as follows:
Analysis of the titratable acidity of L. paragasseri lg94 grown on various substrates (glucose, lactose, and sterile milk) revealed the dependence of titratable acidity on the nature of the substrate used: on a medium with lactose, titratable acidity was 142 °T, which exceeded the value obtained during cultivation on a medium with glucose (100 °T); however, the level of titratable acidity on lactose was lower than on sterile milk (176 °T). A dense, uniform clot was observed during L. paragasseri lg94 cultivation on sterile milk. The formation of a characteristic aroma combining sweet and sour-milk notes was noted. The present results indicate the influence of substrate type on the metabolic activity and acid-forming capacity of the studied lactobacilli strains, as well as the formation of the product’s organoleptic properties. Milk fermentation is a direct consequence of the strain’s active acid formation. Lactic acid accumulation in milk leads to reduced pH, destabilizing casein micelles, and subsequent protein coagulation.
The results indicated that exposure to a universal buffer (pH 7.0) did not
significantly affect the viability of the L. paragasseri lg94 strain.
However, the strain demonstrated low resistance to acid stress; when exposed to a
medium with pH 2.0, the CFU count was 0.5
According to the disk diffusion assay, strain lg94 exhibited phenotypic susceptibility to ampicillin, amoxicillin, penicillin G, tetracycline, erythromycin, and chloramphenicol, as well as intermediate resistance to oxacillin. Phenotypic resistance to fosfomycin, gentamicin, and clindamycin was also observed.
Using the perpendicular streak method, high antagonist activity of L.
paragasseri lg94 was detected against all six test cultures: the width of the
inhibition zones ranging from 15.0 to 19.5 mm (Table 3). The largest zone (19.5
| Test strain | Growth inhibition zone, mm (wells) | Activity level (wells)* | Growth inhibition zone, mm (streaks) | Activity level (streaks)** |
| E. coli ATCC 25922 | 13.6 |
Weak | 17.5 |
High |
| E. coli 235 LMM | absent | – | 16.5 |
High |
| P. aeruginosa ATCC 27853 | 13.0 |
Weak | 18.5 |
High |
| P. aeruginosa ODKB-58 | absent | – | 19.5 |
High |
| K. pneumoniae ODKB-81 | absent | – | 15.0 |
High |
| K. pneumoniae ODKB-82 | absent | – | 16.5 |
High |
*Well method: 10–15 mm—weak, 15–20 mm—moderate,
However, the well method showed significantly weaker or no activity. Inhibition
zones were detected only against the reference strains E. coli ATCC
25922 (13.6
The resulting antagonism profile—strong activity in the perpendicular streak method with little or no effect in the well method—is typical of an acid-dependent inhibition mechanism and suggests that lactic acid is the primary antagonist of L. paragasseri lg94 under these culture conditions, which creates localized acid stress at the pathogen contact zone.
Assembly genome analysis yielded one chromosomal sequence and two plasmid sequences. The chromosome length was 1,894,585 bp, and the GC nucleotide percentage was 35.24. In the process of coding sequence labeling, 4001 sequences that potentially encode proteins, 85 tRNA sequences, 8 rRNA, 66 small miscRNA, and 3 tmRNA were identified.
The BlastKOALA annotation revealed that the genome contains 2105 functional units (Fig. 1). The largest number of genes was classified as “Genetic information processing”. Using the KEGG Mapper application, it was found that L. paragasseri lg94 had the necessary pentose phosphate pathway enzyme complex and enzymes that ensure the transport and metabolism of pentasaccharides and hexasaccharides.
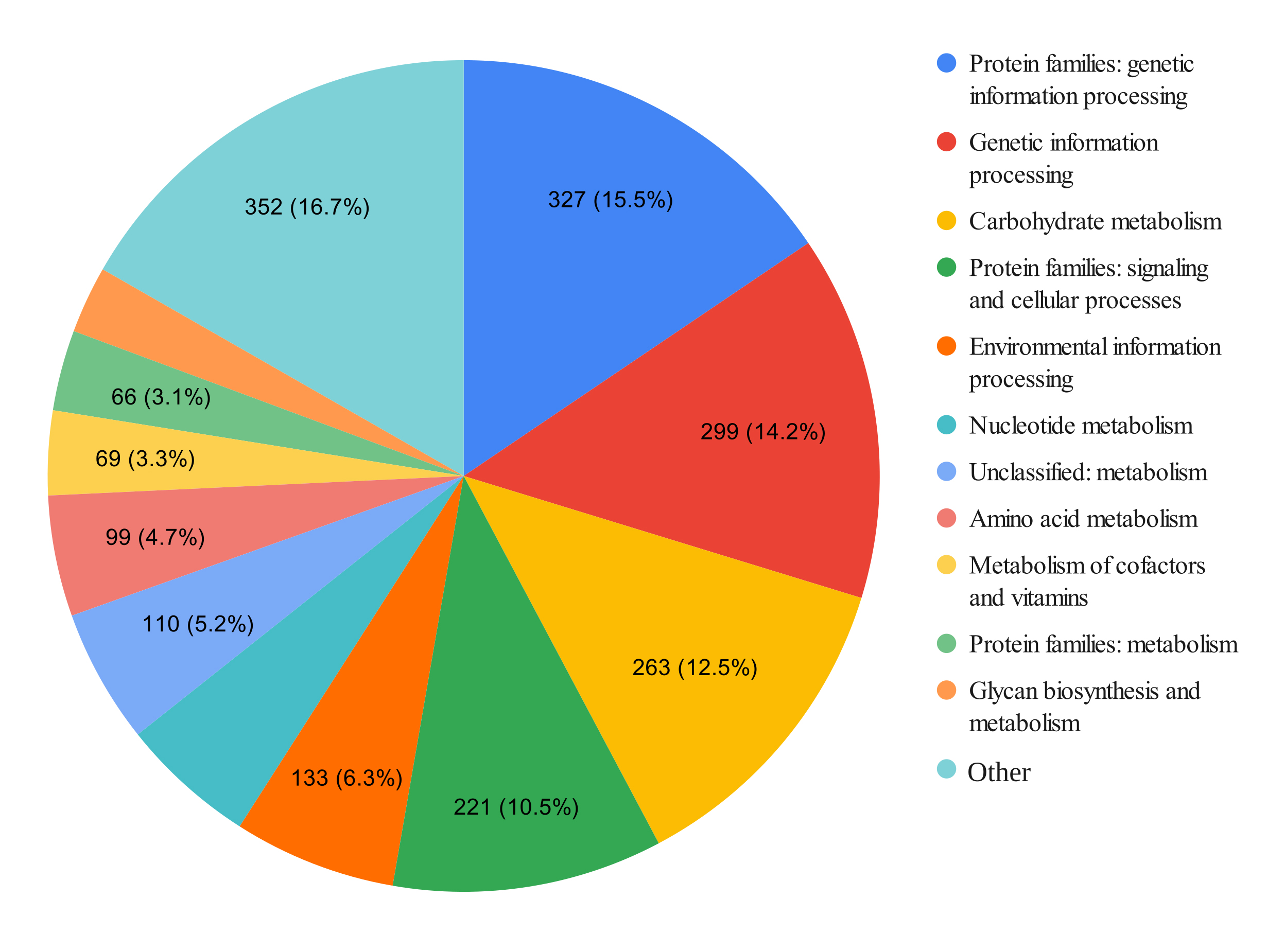 Fig. 1.
Fig. 1.
Distribution of the number of genes in the genome of L. paragasseri lg94 by functions of metabolic pathways. Different colors represent different groups of metabolic pathways, grouped according to the KEGG classification. KEGG, Kyoto Encyclopedia of Genes and Genomes.
Biochemical testing of L. paragasseri lg94 revealed its ability to
ferment various carbohydrates, including mono-, di-, and trisaccharides, sugar
alcohols,
Genes potentially responsible for the putative probiotic properties were identified in the genome. Genes or proteins involved in acid and bile tolerance, intestinal mucosal adhesion, temperature tolerance, host immunomodulation, antimicrobial activity, and internal defense were identified.
The genes NhaC (Na+/H+ antiporter NhaC) and F0F1 ATP synthase are involved in pH and gastric tolerance. Moreover, bile acid tolerance is mediated by cbh choloylglycine hydrolase and ycaD, an MFS transporter. The adhesion genes triose phosphate isomerase tpiA and elongation factor Tu tuf are responsible for the strain’s adhesive properties. The molecular chaperone genes DnaJ and DnaK and the cold shock protein cspA mediate temperature tolerance. Proteins encoded by the LTA synthase family protein ltaS1, D-alanyl-lipoteichoic acid biosynthesis protein (DltD), glycosiletransferases (mshA, epsF, ftsW, rodA, glyE), and glucosaminefructose-6-phosphate aminotransferase (GlmS) genes participate in host immunomodulation processes.
The antimicrobial oxidase genes pyruvate oxidase pox5, which confer resistance to other bacteria, were also found in the genome.
The online tool MiGA was used to confirm the identification of the strain under study. The closest relatives of the strain lg94 found by MiGA in the database were L. paragasseri GCF 003584685 T (98.74% AAI) and L. gasseri GCF 000014425 T (96.64% AAI). The dataset most likely belongs to the same root GCF 003584685 1 (p-value: 0), probably belongs to the species Lactobacillus paragasseri (p-value: 0.012), and may even belong to the same dataset as GCF 003584685 (p-value: 0.37).
No antibiotic resistance, pathogenicity, or disinfectant resistance genes were found in the studied strain’s genome. The key virulence genes (gelE, hyl, asa1, cylA, esp, cytK, nhe, hbl, and sprE [35]) were also absent.
Screening of genes responsible for the production of biogenic amines was conducted. Lactic acid bacteria produce biogenic amines in fermented and spoiled food products [36]. Increased levels of biogenic amines can negatively affect the human body, causing hypertension, headache, increased heart rate, vomiting, or diarrhea [37, 38]. No genes associated with the production of histamine, putrescine, and tyramine (hdcA, tyrRS, odc, aguA, potE) were found in the genome of the L. paragasseri lg94 strain, but the hisS gene (histidyl-tRNA synthetase) was found. However, this gene is a housekeeping gene present in many bacteria.
Two plasmids were found in the L. paragasseri lg94: AC196 and AE118 (Table 4). Plasmids did not carry pathogenicity or resistance determinants.
| Parameters | lg94-AE118 | lg94-AC196 |
| Contigs | 7 | 1 |
| Size | 19,433 | 3239 |
| GC content % | 31.5 | 37.4 |
| Replicone type | - | rep_cluster_707 |
| Relaxase type | - | MOBQ |
| Predictable mobility | Non-mobile | Mobile |
| Cluster | AE118 | AC196 |
| Hypothetical host carrier | Lactobacillus | Lactobacillaceae |
Plasmid AC196 contained only one contig with a length of 3239 bp. Six coding sequences were identified and annotated as proteins with unknown functions. Plasmid AE118 contained seven contigs with a length of 19,433 bp. A total of 22 coding sequences were identified, 14 of which were annotated as proteins with unknown functions. The remaining genes were represented by transposase, sporulation inhibitors, and various transport proteins.
No prophage sequences or CRISPR-Cas loci were detected in the studied genome.
Clusters of genes producing enterolysin A, helveticin J and gassericin T were found for the strain lg94. A cluster of genes responsible for the production of gassericin T is shown in Fig. 2. We also identified the ribB, ribT, and cobC genes responsible for riboflavin and cobalamin synthesis.
 Fig. 2.
Fig. 2.
Gassericin T production cluster in the genome of L. paragasseri lg94. Different genes of the gassericin production cluster are represented in different colors.
Despite the growing interest in this species, information on its genetics, and particularly on the genetic basis of its probiotic potential, remains limited. At the same time, bacteriocins have been identified as one of the key factors underlying probiotic activity; however, their detection using conventional in vitro methods is complex and labor-intensive. In this context, in silico genome analysis for the presence of bacteriocin operons is considered a promising and more efficient approach for bacteriocin screening. According to the literature, genomes of L. paragasseri harbor operons responsible for the production of various bacteriocins, including pediocin, gassericins A, T, and K7B, acidocin B, bacteriocin LS2 (chains a and b), bacteriolysin, helveticin-J, and enterolysin A, with genes encoding gassericin T, acidocin B, and gassericin A being the most prevalent (Table 5, Ref. [39, 40, 41]).
| Strain | Reference | Bacteriocin gene cluster |
| lg94 | this study | enterolysin A, helveticin J, gassericin T |
| UMB3776 | Jackson et al., 2024 [39] | gassericin T |
| UMB4347 | Jackson et al., 2024 [39] | gassericin T |
| UMB1634 | Jackson et al., 2024 [39] | gassericin T, A, S |
| UBLG-36 | Mehra et al., 2021 [40] | gassericin T, S, acidocin B |
| 79 genomes | Zhou et al., 2020 [41] | gassericin T, acidocin B, bacteriocin-LS2chain, helveticin J |
The UBLG-36 genome contains genes encoding proteins that may determine its putative probiotic properties. Genes or proteins involved in acid and bile tolerance, intestinal mucosal adhesion, temperature tolerance, host immunomodulation, antimicrobial activity, and endocrine defense have been identified [39]. Strain lg94 has the same spectrum of genes that provide probiotic potential, except for some adhesion genes (LPXTG cell wall anchor domain-containing protein, Membrane lipoprotein lipid attachment site-containing protein, CpsD/CapB family tyrosine-protein kinase and MucBP domain-containing protein) and resistance to temperature shock (Co-chaperone GroES, Chaperonin GroEL).
CRISPR-Cas adaptive immune systems have previously been described in a number of L. paragasseri and L. gasseri genomes and are considered an important factor in resistance to invasive DNA, including bacteriophages and plasmids, as well as a potential taxonomic marker for distinguishing closely related species. A published study by Zhou et al. [41] demonstrated the widespread distribution of the CRISPR-Cas subtype II-A in both species, along with species-specific differences in Cas protein composition and locus organization, as supported by phylogenetic analyses of Cas1, Cas2, and Cas9. However, in the present study, no components of the CRISPR-Cas system, including the cas1, cas2, and cas9 genes, were detected in the analyzed L. paragasseri genomes. This discrepancy with previously reported data indicates pronounced intraspecies variability in L. paragasseri with respect to the presence of CRISPR-Cas systems and may reflect differences in the evolutionary histories of strains as well as adaptation to environments with reduced pressure from mobile genetic elements.
Genomic analysis of L. paragasseri lg94 allowed us to identify the
genetic determinants of its metabolism and use potential. The strain has a
complete set of genes encoding enzymes necessary for glycolysis and pentose
phosphate pathways, ensuring efficient hexose and pentose metabolism. The genome
lacks the gene encoding phosphoketolase, which allows us to classify L.
paragasseri lg94 as a homofermentative strain that mainly produces lactic acid
[41]. The presence of multiple bgl genes encoding
In addition to key metabolic characteristics, L. paragasseri lg94 possesses genes that determine additional functional properties, making it an attractive candidate for improving fermented products and enhancing probiotic action. The discovery of a gene cluster associated with riboflavin (vitamin B2) synthesis and a gene involved in cobalamin (vitamin B12) suggests that L. paragasseri lg94 synthesizes these essential vitamins. The ability to synthesize B vitamins in situ is a significant advantage of using the strain as a probiotic because it allows the intestinal microbiota to be enriched with these important nutrients. This property can be used to enrich fermented products (e.g., yogurt or kefir) with vitamins, thereby increasing their nutritional value.
The presence of genes associated with the synthesis of exopolysaccharides indicates the potential ability of L. paragasseri lg94 to produce these polysaccharides. Exopolysaccharides are important components of the bacterial cell wall and are secreted into the environment. Exopolysaccharides produced by lactic acid bacteria have valuable properties that improve the texture and rheological characteristics of fermented products [43].
Lactobacilli inhabiting the intestinal biotope are gassericin producers [44]. According to Ene et al. [18], reported that all L. paragasseri strains with publicly available genomes had gassericin genes (T, S, or both). This suggests that the properties once attributed to L. gasseri may be more represented by L. paragasseri species [18]. The biologically active properties of gassericins have been described in the literature. Liang et al. [45] showed that a modified gassericin could effectively inhibit the growth of S. mutans in vitro and the development of dental caries in vivo. These results reveal a new peptide-based agent as a topical treatment for dental caries, paving the way for clinical trials to explore its potential in caries prevention. Mahdavi et al. [46] demonstrated the effect of the bacteriocin gassericin A, which is produced by some intestinal bacteria, on an in vivo obesity model. Although gassericin A did not cause weight loss, it may alleviate obesity-related complications. There is evidence for the suitability of gassericin as a food preservative [47] and in veterinary medicine [48].
Several limitations of this study should be considered. First, the investigation was based on a single strain, L. paragasseri lg94, which limits the generalizability of the findings at the species level. Given the known strain-specific nature of probiotic properties, further comparative analyses involving multiple L. paragasseri strains are required.
Second, the functional properties of the strain were primarily evaluated using in vitro assays and in silico genome analysis. Although these approaches provide valuable preliminary insights into the potential of probiotics, they do not fully reflect the gastrointestinal environment’s complex conditions. The observed low resistance to highly acidic conditions (pH 2.0) may have different implications in vivo, where buffering effects of food and host factors are present.
Third, the antagonistic activity results suggest an acid-dependent inhibition mechanism; however, the contribution of specific antimicrobial compounds (e.g., bacteriocins such as gassericin T, enterolysin A, and helveticin J) was not experimentally validated. Additional targeted studies are needed to confirm their expression and biological activity.
Fourth, although whole-genome sequencing did not reveal genes associated with virulence, pathogenicity, or antibiotic resistance, genomic predictions alone cannot fully exclude potential safety risks. A comprehensive safety assessment, including in vivo toxicity and long-term colonization studies, is required.
Finally, this study did not include in vivo experiments or clinical validation, which are essential to confirm the strain’s probiotic efficacy, safety, and functional effects in the host organism.
The main characteristics of the L. paragasseri lg94 strain were established. The biochemical profile results indicate that L. paragasseri lg94 has several saccharolytic activities. The diversity of fermentable carbohydrates may be an important factor determining the adaptation of the strain to various ecological niches and its potential use in the food industry or as a probiotic. L. paragasseri lg94 has high L-lactic acid production and the ability to ferment milk, making it potentially useful for the production of fermented products (e.g., yogurt, cheese, sauerkraut, etc.) or as a biopreservative. The high acid-forming activity of L. paragasseri lg94 can be used to suppress the growth of unwanted microorganisms and increase the shelf life of food products. The absence of antibiotic resistance, pathogenicity, and virulence determinants, as well as prophage sequences and CRISPR-Cas loci in the genome, was confirmed. The ability of L. paragasseri lg94 to synthesize exopolysaccharides can be used to improve the quality of fermented dairy products. L. paragasseri lg94, which encodes genes for acetaldehyde metabolism, has the potential to produce this aromatic compound. The cultivation conditions must be optimized to control the acetaldehyde concentration to ensure a pleasant aroma. This strain is capable of gassericin, riboflavin, and cobalamin biosynthesis. Thus, whole-genome sequencing confirmed the safety and probiotic potential of L. paragasseri lg94. The next stage of the work is recommended to study its antagonistic activity and metabolite production.
AAI, average amino acid identity; CFU, colony-forming units; Cas, CRISPR-associated; CRISPR, clustered regularly interspaced short palindromic repeats; DNA, deoxyribonucleic acid; KEGG, Kyoto Encyclopedia of Genes and Genomes; MALDI, matrix-assisted laser desorption ionization; OD, optical density; RNA, ribonucleic acid.
The genome sequencing results are submitted to the NCBI database under PRJNA1248600 (https://www.ncbi.nlm.nih.gov/bioproject/?term=PRJNA1248600) as accession SAMN47864912 (https://www.ncbi.nlm.nih.gov/sra?LinkName=biosample_sra&from_uid=47864912). The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
NB, NSe and LR designed the research study. EK, AG, NSm, RZ, AM and NG performed the research. EK, AG and NSm analyzed the data. EK, NB, AG, and NSm wrote the manuscript, EK wrote the first draft. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
When working with the study participants, the ethical principles set forth in the Helsinki Declaration of the WMA (2013 edition) and the Order of the Russian Ministry of Health dated April 1, 2016, No. 200n “On Approval of the Rules of Good Clinical Practice” were observed. The study was approved by the Biomedical Ethics Committee of the Federal State Budgetary Scientific Institution ‘Scientific Center for Family Health and Human Reproduction Problems’ (Protocol No. 5 dated May 16, 2016) and was conducted with the informed consent of adolescents over 15 years of age or parents/legal guardians of children under 15 years of age.
The study was conducted using equipment from the Center for the Development of Progressive Personalized Health Technologies and the Irkutsk Region Human Microbiota Collection (Irkutsk). We thank the reviewers of this publication for their opinions and suggestions.
The study was carried out within the framework of state topic “Metabolic potential of microbial consortia as a basis for a technological approach to the development of new probiotic preparations” (No. 125011600372-4).
The authors declare no conflicts of interest.
During the preparation of this work the authors used TrinkaAI in order to check spell and grammar. After using this tool, the authors reviewed and edited the content as needed and takes full responsibility for the content of the publication.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBE47162.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.