

 , Nardis Nkoudou Ze 1,*
, Nardis Nkoudou Ze 1,* , Marielle Neisha Belinga Atangana 1, Eliane Flore Eyenga 2, Jean Justin Essia Ngang 1
, Marielle Neisha Belinga Atangana 1, Eliane Flore Eyenga 2, Jean Justin Essia Ngang 11 Department of Microbiology, Faculty of Science, University of Yaoundé 1, Yaounde, Cameroon
2 Department of Biochemistry, Faculty of Science, University of Yaoundé 1, Yaounde, Cameroon
Abstract
This study aimed to improve the cassava retting efficiency of starter Sta_96, a fermented cassava flour obtained after four days of fermentation, by incorporating bacteria of the genus Streptomyces.
Streptomyces isolates were obtained from Sta_96, farm soils, and a cassava peel dump. The subsequent pectinolytic and cellulolytic activities, along with the associated ability to soften cassava roots, were evaluated. The most effective isolates were incorporated into Sta_96 at a concentration of 106 CFU/g, and the associated retting and detoxification efficiencies were assessed.
Of the 33 Streptomyces isolates recovered, 24 produced pectinases, with hydrolysis halos ranging from 11.0 ± 1.4 mm to 42.5 ± 3.5 mm. Sixteen of these displayed cellulase activity, with halos ranging from 16.0 ± 0.1 mm to 40.0 ± 0.1 mm. STF 16 showed the highest softening efficiency, achieving a retting time of 38.6 ± 0.4 h, and significantly improved the performance of Sta_96 by reducing the retting duration from 44.5 ± 0.6 h to 24.3 ± 0.4 h, representing a 45.39% reduction. STF 16 also demonstrated substantial detoxification efficiency (~85%).
This is the first report demonstrating that Streptomyces species can act as efficient cassava retting agents. Incorporation of STF 16 into the Sta_96 starter markedly accelerated root softening to about 24 hours while ensuring complete elimination of cyanide. This approach represents a promising strategy for improving fermentation efficiency and product safety in cassava-processing industries.
Keywords
- cassava
- fermentation
- Streptomyces sp.
- pectinase
- cellulase
- cyanides
Cassava is one of the most important tuberous roots cultivated in tropical regions, serving as the staple food for nearly 800 million people worldwide [1]. In Cameroon, this tuberous root is the primary source of calories [2], representing a valuable alternative to imported cereals such as rice, sorghum, and wheat [3]. Its high carbohydrate content and adaptability to marginal soils make cassava a critical crop for food security and rural livelihoods. Despite these benefits, roots from high-yielding varieties contain elevated levels of cyanogenic glycosides, mainly linamarin and lotaustralin [4]. Their enzymatic hydrolysis releases cyanide, which is associated with severe health disorders, including goiter, cretinism, ataxic neuropathy, and xerophthalmia [5]. Effective detoxification during processing is therefore essential, highlighting the critical role of cassava retting. Traditional retting practices used by smallholder farmers in Central and West Africa reduce cyanide levels while contributing to the characteristic sensory attributes of fermented cassava products [6, 7]. However, fermentation remains slow, typically lasting 3–7 days, depending on environmental conditions, cassava variety, root maturity, and the composition of microbial communities [8]. Prolonged retting increases postharvest losses due to spoilage, limits production capacity, and constrains the revenue of processors—most of whom are women working in small-scale industries. Additionally, the lack of standardized processing conditions leads to variability in product safety and quality. To shorten fermentation time and improve product consistency, the use of defined microbial starters has been proposed [9]. Starters based on lactic acid bacteria, yeasts, and fungi have shown promise under controlled laboratory settings [10, 11], yet often perform inconsistently in field conditions due to microbial competition and substrate heterogeneity. In response, the traditional starter Sta_96—comprising a naturally selected microbial consortium derived from pre-fermented cassava chips—has gained traction as a more robust option [12]. Sta_96 reduces retting time by approximately 50% while maintaining residual cyanide below recommended safety thresholds (20 mg/kg) [12]. Nevertheless, further improvement is necessary to meet increasing consumer demand and industrial requirements for rapid and reliable processing cycles. Enzymatic degradation of cassava root cell walls is essential for efficient retting, suggesting that enrichment of starter cultures with microbes capable of producing strong hydrolytic enzymes could further enhance fermentation performance. Streptomyces species are well-recognized soil bacteria known for secreting diverse extracellular enzymes, including cellulases, xylanases, proteases, and pectinases [13], which contribute to plant tissue softening and have wide applications in agriculture and food biotechnology [14]. Despite this enzymatic potential, the use of Streptomyces as direct retting agents for cassava has not yet been investigated, representing a significant knowledge gap and an untapped biotechnological opportunity. Therefore, the objective of this study is to evaluate whether incorporating pectinolytic and cellulolytic Streptomyces strains into the Sta_96 starter can enhance its retting efficiency. By improving cell wall degradation and potentially accelerating detoxification, this approach aims to standardize fermentation duration, ensure product safety, and promote industrial-scale cassava processing. Ultimately, such advancements could strengthen food security, reduce postharvest losses, and support the economic empowerment of local producers by increasing both the quality and availability of fermented cassava products in Central and West African markets.
Streptomyces spp. were isolated following the protocol described by
[15] from three distinct sources: the traditional starter Sta_96 produced in the
laboratory according to [9], farm soils, and a cassava peel dump soil. For soil
samples collection, 100 g of soil was collected from three different farms
located in Yaoundé, Cameroon (3.848° N and 11.502° E). Soil
samples were taken at a depth of 10–15 cm below the surface using sterile
trowels to minimize contamination and ensure consistency. Sampling was conducted
during the rainy season (April–May), when microbial activity is typically
highest due to increased moisture and nutrient availability. At the time of
collection, environmental conditions were recorded: average temperature ranged
from 24–28 °C, relative humidity was between 75–85%, and the soil was
moderately moist with visible organic matter. Samples were immediately placed in
sterile plastic bags, labeled, and transported to the laboratory for processing
within 24 hours to preserve microbial viability. The cassava peel dump sample was
collected from a site with visible microbial colonization and was processed
similarly to the soil samples. Once in the laboratory, samples were mixed with
calcium carbonate (ACS reagent grade,
Cellulolytic and pectinolytic activities were assessed as described by [16] using mineral media supplemented with either Sodium-carboxymethyl cellulose (CMC 10 g/L, Sigma Aldrich CAS Number: 9004-32-4, Saint-Quentin-Fallavier, Isère, France) or pectin (10 g/L, Sigma Aldrich CAS Number: 9000-69-5, Saint-Quentin-Fallavier, Isère, France) as the sole carbon source. Isolates were first grown on ISP4 medium to obtain a confluent microbial lawn. Cylindrical plugs (6 mm diameter) were aseptically removed and placed onto CMC agar or pectin agar. Plates were incubated at 37 °C for 96 h, then flooded with 1% (w/v) potassium iodide solution and stained for 5 minutes. Clear halos surrounding the inoculum discs indicated extracellular cellulase or pectinase production. Hydrolysis halo diameters were measured using a caliper following the standard method described by [16]. All enzymatic activity tests were performed in triplicate for each isolate to ensure reproducibility. Only isolates exhibiting at least one of the enzymatic activities were retained for subsequent experiments. It is important to emphasize that, this study evaluates whole microbial retting performance rather than purified enzyme kinetics, reflecting real fermentation conditions where enzymatic cocktails, microbial succession, and matrix-substrate interactions determine efficacy.
Cassava roots from a 9-month-old bitter local variety (“Six-mois”) [17] selected for its prevalence in Cameroonian fermentation processes, highly cyanogenic glycoside content (400~655 mg HCN/kg fresh weight), and high starch content (70–74% dry matter basis), were peeled, washed, and cut into 1-cm-thick cylinders. Batches of 100 g were inoculated with Streptomyces isolates at 106 CFU/g of root tissue, submerged in 100 mL of tap water, and incubated for natural fermentation. A non-inoculated batch was included as a negative control. Retting progression was monitored over 4 days. Every 3 h, six randomly selected cylinders from each batch were tested for firmness using a penetrometer. The softening level was expressed as the mean penetration depth (six measurements per cylinder) [14]. The penetration measurements were performed using the Penetrometer RPN10 Berlin model; the pressure was applied at a controlled rate of 10 mm/s to insure reproducibility and accuracy of the measurements. Penetration data obtained from each time point were compiled in Microsoft Excel 2016 and fitted using DM-fit software (https://browser.combase.cc/DMFit.aspx). The resulting sigmoid kinetics were modeled according to the equation described by [18], thereby, enabling calculation of the retting time as follows:
Here, Dr, Lag, L and µ are respectively the retting time (h), the time required to initiate fermentation (h), the critical penetrometry index (cm/h) and the softening speed (cm/h).
The percentage reduction of the retting time of each strain was calculated as follows:
Isolates demonstrating the shortest retting times were selected for further
testing. They were incorporated into cassava roots at a concentration of 106
CFU/g either individually or in combination with the traditional Sta_96 starter.
The effectiveness of the enriched starters in accelerating retting was evaluated
by comparing their retting duration with that of the non-enriched Sta_96
starter. Cassava roots were peeled, washed with tap water, and cut into
1-cm-thick cylinders. Batches of 100 g were prepared, and each selected isolate
was mixed with Sta_96 starter at a load corresponding to 10⁶ CFU/g of starter.
The mixtures were then applied to each batch, except for the controls
(spontaneous fermentation and Sta_96-only control), and submerged in 100 mL of
tap water. Where relevant, isolates showing retting-enhancing properties were
also tested in combination with the best-performing isolate to further improve
the efficiency of the Sta_96 starter. All treatments were incubated under
laboratory conditions (28 °C
The effect of Streptomyces enrichment on the detoxification activity of
Sta_96 was assessed by quantifying the total cyanide content of the retted
cassava paste using the method of [11]. Briefly, 0.1 g of fermented cassava paste
was placed into screw-cap vials containing 1 mL of distilled water and a strip of
picrate paper. Vials were sealed and incubated at 30 °C for 18 h to allow
cyanide interaction with the picrate paper. Thereafter, the picrate papers were
transferred into test tubes containing 5 mL of distilled water and boiled for 5
min. Following boiling, the strips were removed and the solution was allowed to
cool to room temperature (28 °C
All statistical analyses were performed using the IBM SPSS Statistics version
26.0 (IBM Corp., Chicago, IL, USA). The Duncan range test was applied to compare
differences in pectinolytic and cellulolytic activities among
Streptomyces isolates, retting times of Sta_96–Streptomyces combinations, and cyanide detoxification among treatments. Significant
differences are represented by distinct letters in figures and tables. All data
are expressed as arithmetic means
To investigate the phenotypic characterization of all 33 Streptomyces isolates, the macroscopic and microscopic characteristics of strains from different sources were investigated. The summary reported in Table 1 shows that, among isolates originating from the starter Sta_96, 66.66% produced orange to brown pigments. In contrast, 58.82% of isolates from cassava field soils exhibited pigment production, predominantly pink. Additionally, 53.84% of isolates obtained from cassava peel soil produced pigments ranging from orange to brown.
| Sources | Isolates | Mycelium color | Gram strain | Shape | Pigment coloration |
| Sta_96 | STF 16 | Cream | + | Filamentous | Orange |
| STF 28 | Wheat | + | Filamentous | Brown | |
| STF 33 | Pink | + | Filamentous | None | |
| Cassava cultivated soil | CCS 4 | White | + | Filamentous | None |
| CCS 8 | Brown | + | Filamentous | None | |
| CCS 17 | Wheat | + | Filamentous | None | |
| CCS 18 | Cream | + | Filamentous | None | |
| CCS 20 | Gray | + | Filamentous | None | |
| CCS 21 | Brown | + | Filamentous | White | |
| CCS 6 | Gray | + | Filamentous | Green | |
| CCS 9 | White | + | Filamentous | None | |
| CCS 11 | Cream | + | Filamentous | Pink | |
| CCS 22 | Wheat | + | Filamentous | Wheat | |
| CCS 29 | Brown | + | Filamentous | pink | |
| CCS 30 | Brown | + | Filamentous | Brown | |
| CCS 13 | Gray | + | Filamentous | Gris | |
| CCS 19 | Pink | + | Filamentous | Pink | |
| CCS 23 | Pink | + | Filamentous | None | |
| CCS 26 | Green | + | Filamentous | Brown | |
| CCSl27 | Gray | + | Filamentous | Pink | |
| Cassava dumping soil | CDS 2 | White | + | Filamentous | Orange |
| CDS 7 | Light-gray | + | Filamentous | None | |
| CDS 14 | Gray | + | Filamentous | White | |
| CDS 15 | Gray | + | Filamentous | None | |
| CDS 24 | Blue | + | Filamentous | None | |
| CDS 25 | Blue | + | Filamentous | Brown | |
| CDS 31 | Gray | + | Filamentous | Orange | |
| CDS 32 | Brown | + | Filamentous | Brown | |
| CDS 1 | White | + | Filamentous | None | |
| CDS 3 | Wheat | + | Filamentous | Orange | |
| CDS 5 | White | + | Filamentous | None | |
| CDS 10 | Orange | + | Filamentous | None | |
| CDS 12 | Pink | + | Filamentous | Pink |
STF, Strain from Sta_96 Flour; CCS, Strain from Cassava Soil; CDS, Strains from Cassava Dumping Soil.
Table 2 summarizes the pectinolytic and cellulolytic activities of the strains.
A total of 24 out of 33 (72.7%) demonstrated pectinolytic activity, indicated by
clear zones of pectin hydrolysis varying from 11.0
| Sources | Strains | Pectinolytic activity | cellulolytic activity |
| Sta_96 | STF 16 | 30.0 |
29.5 |
| STF 33 | 31.0 |
30.0 | |
| STF 28 | 27.5 |
||
| Cassava cultivated soil | CCS 4 | 39.0 | |
| CCS 20 | 37.5 |
33.0 | |
| CCS 21 | 41.5 |
21.5 | |
| CCS 6 | 30.0 |
22.0 | |
| CCS 22 | 42.5 |
25.0 | |
| CCS 8 | 32.0 |
20 | |
| CCS 17 | 38.5 |
||
| CCS 18 | |||
| CCS 9 | |||
| CCS 11 | 30.0 |
||
| CCS 29 | 34.5 |
||
| CCS 30 | 11.0 |
||
| CCS 13 | 37.5 |
||
| CCS 19 | 30.0 |
||
| CCS 23 | |||
| CCS 26 | |||
| CCS 27 | |||
| Cassava dumping soil | CDS 2 | 42.5 |
27.5 |
| CDS 15 | 41 |
28.5 | |
| CDS 24 | 36.0 |
35.0 | |
| CDS 31 | 34.0 |
34.5 | |
| CDS 32 | 38.0 |
25.0 | |
| CDS 12 | 27.5 |
17.5 | |
| CDS 1 | 40.5 |
16.0 | |
| CDS 7 | 40 | ||
| CDS 14 | 43 |
35 | |
| CDS 25 | 23 |
21.5 | |
| CDS 3 | 19.5 |
||
| CDS 5 | 36.5 |
32 | |
| CDS 10 | 27.5 |
The results are presented as the means
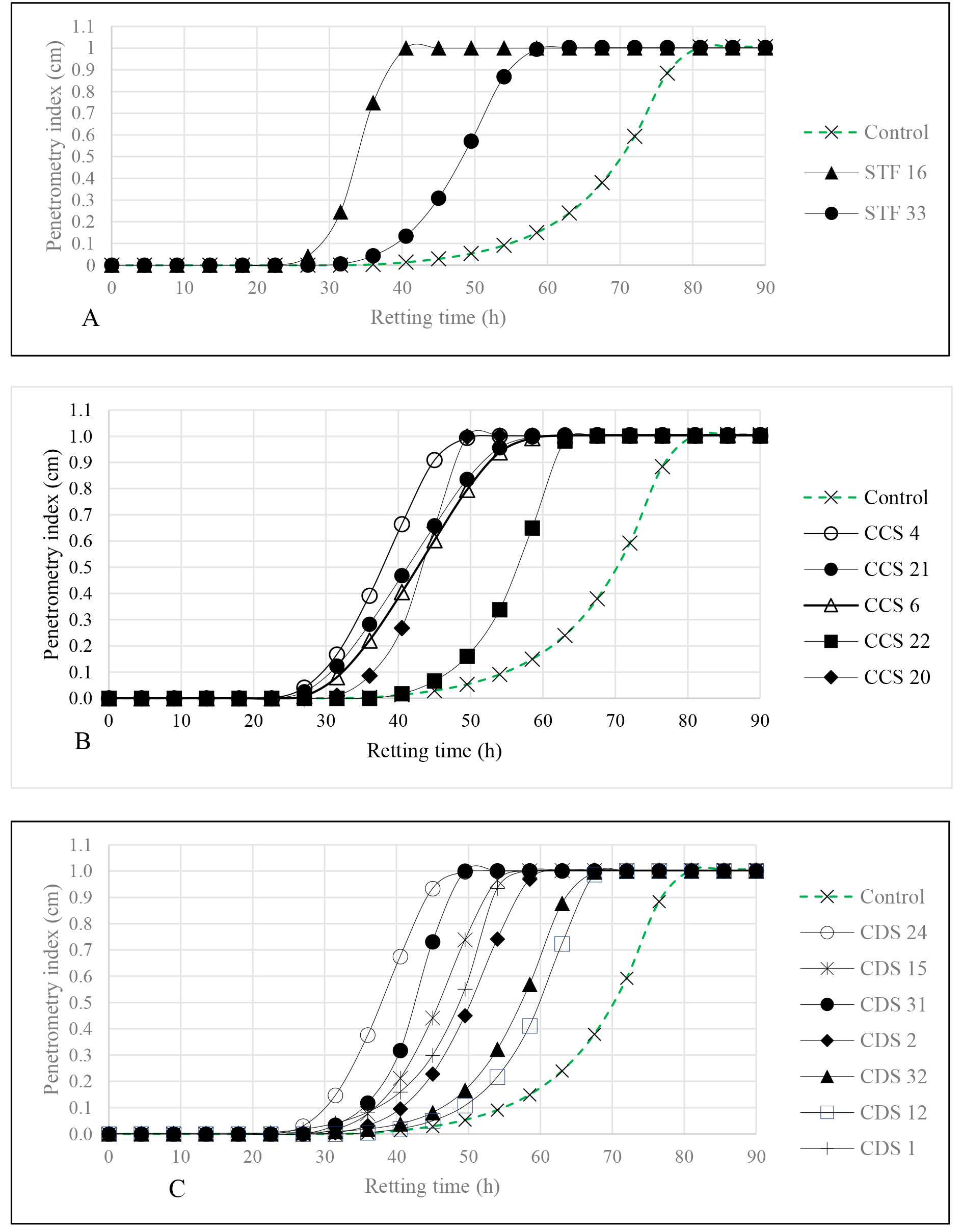
The ability of the isolates to soften cassava roots presented in Fig. 1, was
compared with that of the control, using rating time as evaluating evaluation
metric. Retting time, ranged from 38.6
 Fig. 1.
Fig. 1.
Cassava roots: (left) non-retted cassava root and (right) retted-softened.
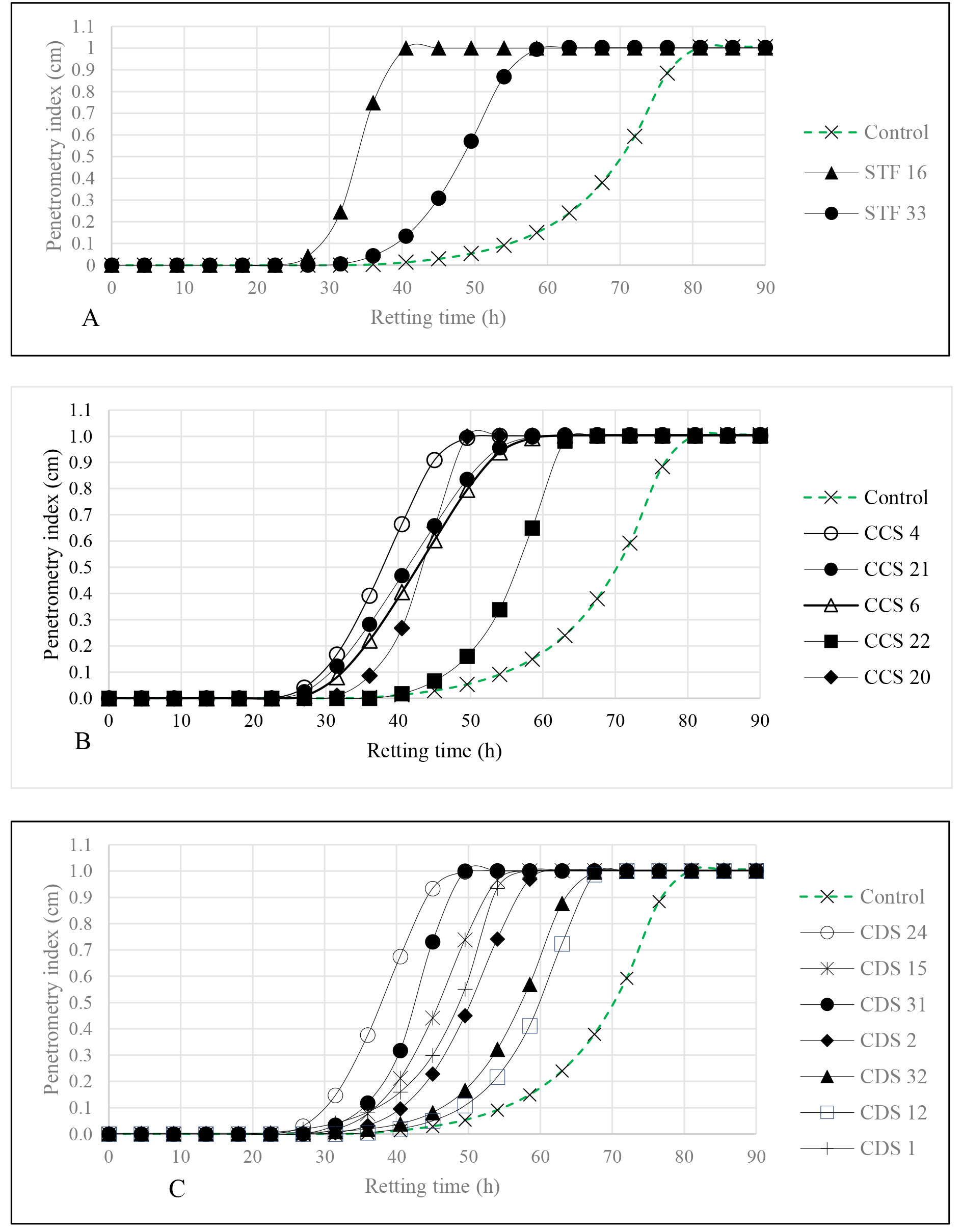 Fig. 2.
Fig. 2.
Kinetics of cassava roots softening by: starter isolates (A), soil from a cassava field (B) and soil from cassava peels (C) Control defined as sterile water-inoculated group.
| Sources | Strains | Retting time (h) | Retting time percentage Compared to control | Retting time of control (h) |
| Sta_96 | STF 16 | 38.6 |
57.4% | 90.5 |
| STF 33 | 55.3 |
38.9% | ||
| STF 28 | 0.0% | |||
| Cassava cultivated soil | CCS 4 | 45.2 |
50.1% | |
| CCS 20 | 44.7 |
50.6% | ||
| CCS 21 | 52.9 |
41.6% | ||
| CCS 6 | 53.1 |
41.3% | ||
| CCS 22 | 67.3 |
25.6% | ||
| CCS 8 | 0.0% | |||
| CCS 17 | 0.0% | |||
| CCS 18 | 0.0% | |||
| CCS 9 | 0.0% | |||
| CCS 11 | 0.0% | |||
| CCS 29 | 0.0% | |||
| CCS 30 | 0.0% | |||
| CCS 13 | 0.0% | |||
| CCS 19 | 0.0% | |||
| CCS 23 | 0.0% | |||
| CCS 26 | 0.0% | |||
| CCS 27 | 0.0% | |||
| Cassava dumping soil | CDS 2 | 57.7 |
36.3% | |
| CDS 15 | 52.2 |
42.% | ||
| CDS 24 | 44.7 |
50.8% | ||
| CDS 31 | 47.9 |
47.1% | ||
| CDS 32 | 65.0 |
28.2% | ||
| CDS 12 | 69.2 |
23.6% | ||
| CDS 1 | 82.0 |
9.4% | ||
| CDS 7 | 0.0% | |||
| CDS 14 | 0.0% | |||
| CDS 25 | 0.0% | |||
| CDS 3 | 0.0% | |||
| CDS 5 | 0.0% | |||
| CDS 10 | 0.0% |
The results are presented as the means
Retting trials using Sta_96 enriched with the five most effective isolates (STF
16, CCS 4, CCS 20, CDS 24, and CDS 31), either individually or in combination,
are presented in Table 4. Enrichment significantly reduced retting times
(p
| Combination | RT (h) | Percentage of improvement compared to Sta_96 | Percentage of improvement compared to Control |
| Control | 90.5 |
0 | 0 |
| Sta_96 | 44.5 |
0 | 50.8% |
| Sta_96 + CCS 4 | 26.9 |
39.6% | 70.3% |
| Sta_96 + CDS 31 | 26.5 |
40.5% | 70.7% |
| Sta_96 + CCS 20 | 27.6 |
38.1% | 69.5% |
| Sta_96 + CDS 24 | 30.3 |
31.9% | 66.5% |
| Sta_96 + STF 16 | 24.3 |
45.5% | 73.2% |
| Sta_96 + CCS 4 + STF 16 | 30.2 |
32.2% | 66.6% |
| Sta_96 + CDS 31 + STF 16 | 26.2 |
41.2% | 71.1% |
| Sta_96 + CCS 20 + STF 16 | 24.5 |
45.1% | 72.9% |
| Sta_96 + CDS 24 + STF 16 | 26.8 |
39.7% | 70.4% |
The results are presented as the means
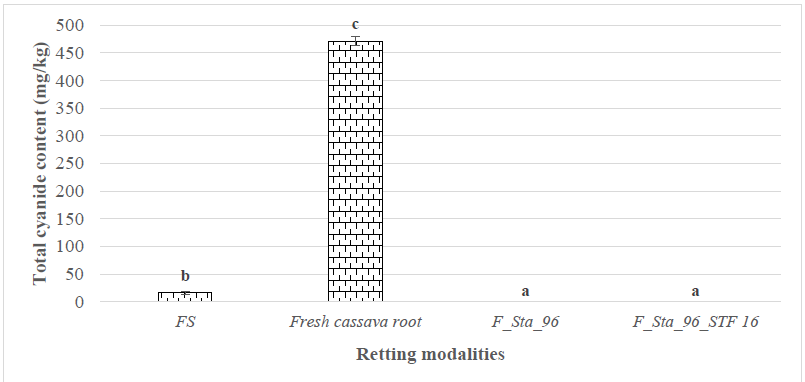
A study of the detoxification efficiency of cassava roots retted with the best
isolate yielded the data shown in Fig. 3. Fresh cassava roots contained 471.5
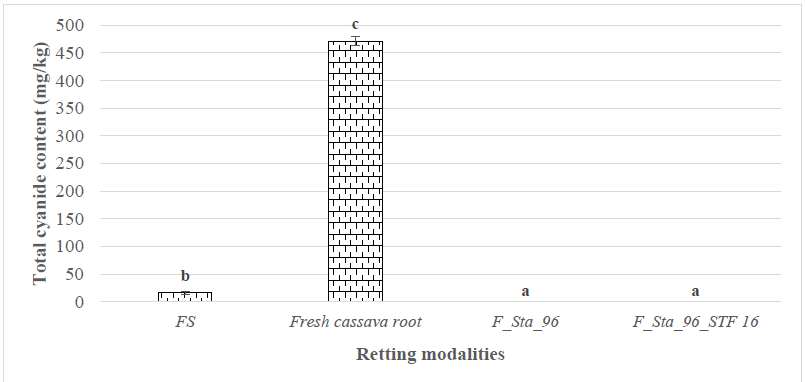 Fig. 3.
Fig. 3.
Residual total cyanide content of fermented cassava. (FS) in the absence
of the traditional starter Sta_96; (F_Sta_96) in the presence of the
traditional starter Sta_96; (F_Sta_96_STF16) in the presence of Sta_96 with
added Streptomyces. Sample size for each group is n = 3. Histograms represent the
mean
Despite the socioeconomic importance of cassava and its role as a staple food in many tropical regions, processing cassava roots into edible and marketable products remains constrained by the long duration of retting, a key unit operation in fermentation. Although various strategies have been explored to shorten retting times, current solutions remain insufficient to support growing consumer and industrial demands. In this context, this study aimed to enhance the retting efficiency of a traditional starter (Sta_96), previously reported to reduce retting duration [9], through supplementation with selected Streptomyces strains.
A total of thirty-three Streptomyces isolates displaying diverse morpho-cultural and microscopic characteristics were collected from Sta_96 flour samples, field soil and a cassava peel dump soil. This diversity reinforces the well-documented ecological variability of the genus and its capacity to produce a broad spectrum of bioactive molecules, including hydrolytic enzymes [20, 21, 22]. Of the isolates obtained, three originated from the traditional starter, seventeen from cultivated soils, and thirteen from cassava peel disposal sites.
The unequal distribution likely reflects differences in nutrient availability and environmental selective pressures across sampling habitats. Functional screening showed that fourteen isolates demonstrated the ability to soften cassava roots, although to varying extents. This variability may be attributed to differences in microbial physiology, including enzyme synthesis potential, extracellular enzyme concentrations, and environmental tolerance factors such as temperature and pH [23]. Because each microorganism possesses specific optimal growth conditions, cultivating all isolates under identical physicochemical conditions may have favored those better adapted to the assay environment. Additionally, the initial microbial load influences the lag phase duration and, consequently, the overall fermentation kinetics. This may explain why addition of STF 16 to Sta_96 markedly accelerated softening, reducing retting to about 24 hours. One plausible explanation of the notably outperformance of strain STF compared to other isolates in accelerating root softening lies its enhanced enzyme production profile. STF 16 may synthesize higher quantities or more efficient forms of key enzymes—such as pectinases, cellulases, and hemicellulases—that directly degrade plant cell wall components integral to cassava tissue structure. Moreover, STF 16 might possess enzymes with greater catalytic efficiency or broader activity ranges under the assay conditions, enabling more rapid and effective hydrolysis of the matrix.
Evaluation of enzymatic activity revealed that all retting-efficient isolates produced both pectinases and cellulases to varying degrees, except isolate CCS 4, which exhibited only cellulolytic activity. The ability of CDS 4 to soften cassava roots despite lacking detectable pectinolytic activity suggests that softening is not exclusively mediated by pectinases, contrary to earlier assumptions [6, 13]. Cellulose hydrolysis also contributes significantly to tissue disintegration, as cellulose is a key structural component involved in maintaining cell wall rigidity [24, 25]. Furthermore, specific enzymes such as pectate lyases and endoglucanases play pivotal roles in plant cell wall degradation by depolymerizing complex pectin chains and hydrolyzing cellulose microfibrils, respectively [26]. The hypothesis of synergistic enzymatic yield is supported by studies demonstrating that microbial co-fermentation enhances substrate accessibility through sequential release of cell wall polymers, thereby optimizing overall retting performance [27, 28]. Therefore, synergistic action of cellulases and pectinases is likely necessary for optimal retting performance.
Furthermore, the reduction of total cyanide content to non-detectable levels following fermentation with Sta_96 alone or in combination with STF 16 may be explained by both the sensitivity of the quantification assay and the metabolic activity of microorganisms in the retting environment. Enzymatic degradation of structural polysaccharides during fermentation promotes cell wall breakdown, enabling contact between linamarin and endogenous linamarase, thus facilitating cyanogenic glycoside hydrolysis and detoxification of cassava tissues [17]. While this study primarily targeted high-cyanide, bitter cassava varieties—predominant in fermentation processes in Cameroon and across Africa—the detoxification mechanism achieved through enhanced microbial retting is expected to be transferable to sweet varieties, which, despite their lower cyanogenic potential and higher sugar content, may require re-validation of microbial consortium compatibility and process optimization. Industrial parameters for the composite inoculum (Sta_96 supplemented with STF16) are under optimization, and its good storage performance—e.g., 30 days at 4–8 °C—has been preliminarily observed in practical use.
This study demonstrates the potential of pectinolytic and cellulolytic Streptomyces sp. to enhance cassava retting efficiency via the traditional starter Sta_96. Nevertheless, three key limitations temper these conclusions. First, it ignores microbial interactions within Sta_96 and lacks profiling of fermentation end-products. This could potentially overattribute the 85% detoxification to Streptomyces alone. Second, screening only 33 Streptomyces isolates from narrow sources risks selection bias and overlooks superior strains. Finally, the absence of long-term stability, sensory data, and economic comparisons weakens claims of a promising strategy for cassava processing. However, the results of this work remain highly worthy of interest.
In this study, we demonstrated that among the Streptomyces isolates
tested, 24 exhibited pectinolytic activity, while 16 showed both cellulolytic and
pectinolytic activities. Notably, the isolate STF 16 achieved the shortest
retting time (38.6
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
BBDO and BAN performed the research and analyzed the data, NZN designed the research study and curated, analyzed and validated the data. EEF participated in experiments, provided help and advice on biochemistry analysis. BBDO, BAN and NZN wrote the manuscript. JJEN conceptualized and supervised the study. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the manuscript.
Not applicable.
The authors gratefully acknowledge the University of Yaounde 1 for providing the equipment and facilities necessary to carry out this study.
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sector.
The authors declare no conflicts of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBE45577.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.