

 , Tohru Suzuki 1,‡, Takumi Morioka 2, Mayuko Abe 2, Yassien Badr 3,4
, Tohru Suzuki 1,‡, Takumi Morioka 2, Mayuko Abe 2, Yassien Badr 3,41 The United Graduate School of Agricultural Science, Gifu University, 501-1193 Gifu, Japan
2 Graduate School of Natural Science and Technology, Gifu University, 501-1193 Gifu, Japan
3 Department of Infectious Diseases and Epidemics, Faculty of Veterinary Medicine, Damanhour University, 22511 El-Beheira, Egypt
4 Faculty of Applied Biological Sciences, Gifu University, 501-1193 Gifu, Japan
‡Deceased.
Abstract
Gamma-aminobutyric acid (GABA) is the principal inhibitory neurotransmitter in the central nervous system and exerts multiple health-promoting effects, including antidepressant, hypotensive, immunomodulatory, and antidiabetic actions. Previous studies have reported a positive correlation between the abundance of Bifidobacterium species in the gut and fecal GABA levels. However, the direct contribution of specific GABA-producing Bifidobacterium strains and the associated interaction with prebiotics remain insufficiently characterized.
This study aimed to elucidate the direct effects of a GABA-producing strain, Bifidobacterium adolescentis 4-2, in combination with the prebiotic mannooligosaccharides (MOS), on fecal GABA concentrations using the Kobe University Human Intestinal Microbiota Model (KUHIMM), an in vitro simulation of the human colonic microbiota. GABA levels were quantified via high-performance liquid chromatography, and changes in microbial composition following oligosaccharide supplementation were assessed by next-generation sequencing of 16S rRNA genes. The presence and activity of the β-mannosidase producing genes, essential for MOS degradation and present in the B. adolescentis 4-2 genome, were evaluated using enzymatic assays.
B. adolescentis 4-2 significantly increased fecal GABA levels within the KUHIMM. MOS supplementation elevated GABA levels, reduced fecal culture pH, and increased the relative abundance of Actinobacteria, particularly the Bifidobacterium species. Notably, the combined application of MOS and B. adolescentis 4-2 produced a synergistic increase in GABA production compared with the individual application of each component. β-mannosidase activity assays confirmed the effective utilization of MOS by B. adolescentis 4-2, supporting its role in enhancing GABA biosynthesis.
These findings demonstrate a synergistic interaction between MOS and B. adolescentis 4-2 in promoting microbial GABA production in the KUHIMM in vitro. This synbiotic combination shows promise for modulating gut-derived GABA levels in vitro and warrants further investigation in animal models and human clinical studies.
Graphical Abstract
Keywords
- Bifidobacterium adolescentis
- mannooligosaccharides
- GABA
- synergistic interactions
- prebiotic
- probiotic
- gut microbiota
Gamma-aminobutyric acid (GABA) is the principal inhibitory neurotransmitter in the central nervous system and plays a pivotal role in maintaining neurological homeostasis. Beyond its function in the brain, GABA exhibits a wide range of health-promoting effects, including antidepressant, hypotensive, immunomodulatory, and antidiabetic activities [1]. Emerging evidence suggests that the gut microbiota plays a significant role in GABA biosynthesis, with specific bacterial taxa demonstrating the ability to produce this bioactive compound [2, 3]. Among these, Bifidobacterium has been identified as a key genus involved in intestinal GABA production.
Bifidobacteria are beneficial symbionts that predominantly colonize the gastrointestinal tract of mammals, including humans. However, their relative abundance declines with age, often accompanied by an increase in genera such as Escherichia coli and Clostridium [4]. This age-related shift in microbiota composition can adversely affect host health by altering the gut metabolite landscape [5, 6]. These findings underscore the significance of Bifidobacterium supplementation in maintaining microbial balance and promoting host health and well-being. In addition to their gastrointestinal benefits, Bifidobacteria exert systemic physiological and psychological effects [7]. One of their notable metabolites is GABA, categorized as a “postbiotic”, a non-viable bacterial product or metabolic byproduct that confers health benefits to the host [8, 9]. Microbially derived GABA may influence the host via systemic circulation or through gut-brain neural pathways [10].
Prebiotics, particularly oligosaccharides, are known to enhance the growth and metabolic activity of beneficial gut microbes such as Bifidobacterium [11]. Certain strains possess the enzymatic machinery necessary to metabolize oligosaccharides, improving their colonization potential and functional outputs [12, 13]. As indigestible dietary components, prebiotics are fermented by the gut microbiota into short-chain fatty acids and other metabolites with systemic effects [14]. Acting as selective substrates, prebiotics modulate the composition and function of the gut microbiota [11, 12, 13, 15]. For instance, seaweed-derived polysaccharides can enrich Bacteroides species [16], while galactooligosaccharides were reported to increase particular Bifidobacterium species [17]. These microbial shifts contribute to broader changes in the gut metabolome, including altered production of postbiotics [18]. Given its diverse physiological functions, GABA is considered one of the most valuable postbiotics [19].
Our previous work demonstrated a positive correlation between Bifidobacterium abundance and fecal GABA levels in the human gut [20]. Building on these findings, the present study aimed to investigate the direct impact of a GABA-producing Bifidobacterium strain, in combination with oligosaccharide-based prebiotics, on fecal GABA production using an in vitro fecal fermentation model. Experiments were conducted using the Kobe University Human Intestinal Microbiota Model (KUHIMM), a standardized in vitro colonic fermentation system designed to simulate the human gut microbiota under controlled and reproducible conditions [21]. KUHIMM enables longitudinal assessments of microbiota dynamics and metabolite production in response to experimental interventions.
Several Bifidobacterium adolescentis strains were isolated from the gastrointestinal tracts (GIT) of healthy individuals residing in Japan. These isolates are part of the microbial strain collection maintained at the Laboratory of Genome Microbiology, Gifu University, Japan. Additional commercial strains were obtained from the Japan Collection of Microorganisms (JCM).
All strains were cultivated in de Man, Rogosa, and Sharpe (MRS) broth (BD, Cat#
MD21152, USA) at 37 °C under anaerobic conditions using a BUG Box
anaerobic chamber (Ruskinn Technology Ltd., Bridgend, Wales, UK) and a gas
mixture composed of 80% N2, 10% CO2, and 10% H2. To support
GABA production, 1% (v/v) monosodium glutamate (MSG; Sigma-Aldrich, Cat#
G1626, St. Louis, MO, USA) was added to the MRS broth. For standard cultivation,
30–50 µL of frozen stock (stored at –80 °C) was inoculated into
12 mL of MRS broth and incubated at 37 °C for 24 hours. The culture was
subsequently sub-cultured in fresh MRS broth containing 1% MSG and incubated for
an additional 48 hours. Cells were then harvested by centrifugation at 6000
Fecal samples were obtained from healthy adult volunteers with no history of systemic or psychiatric disorders and no antibiotic usage for at least three months prior to sample collection. Donors (n = 8) were healthy adults aged 21–36 years, including both males and females, with no known dietary restrictions. All reported following a mixed diet typical of the local population. Samples were collected using BD BBL CultureSwab Plus (BD Co., New Jersey, US, Cat# 212550), immediately stored at 4 °C, and processed within 12 hours of collection. Written informed consent was obtained from all participants. The study protocol was approved by the institutional ethics review board of Gifu University (Certificate No. 2019-283).
Batch fermentation was conducted using a pH-controlled, multi-channel fermentation system (Kobe University Human Intestinal Microbiota Model, KUHIMM; Kobe, Hyogo, Japan) [22]. The simulator consisted of six parallel and independent vessels. The working volume per vessel was 100 mL of Gifu Anaerobic Medium (GAM broth; Nissui Pharmaceutical Co., Ltd., Code: 05422, Tokyo, Japan) with an initial pH adjusted to 6.5 using manual acid/base titration. KUHIMM is equipped with a pH sensor that continuously monitors and records pH values throughout the entire cultivation period. No fixed pH set point was applied, as the system was designed to record the natural pH changes resulting from microbial fermentation. Anaerobic conditions were maintained by continuous flushing with an N2/CO2 (80:20) gas mixture at a rate of 10 mL min-1 through a 0.2 µm polytetrafluoroethylene membrane (Pall Corporation, Port Washington, NY, USA) for 30 minutes at 37 °C prior to fermentation. The pH was continuously monitored for each vessel, and continuous stirring was maintained at 300 rpm to ensure homogeneous microbial distribution.
For inoculation, fecal samples were suspended in 2 mL of physiological saline and introduced through the side port of each vessel. Based on the experimental design, three additives were prepared: mannooligosaccharides (MOS; van wankum ingredients BV, Maarssen, Netherlands; powder form), fructooligosaccharides (FOS; SigmaAldrich, Cat. F8052, St. Louis, MO, USA) and dextrin (Dex; Shandong Bailong Chuangyuan BioTech Co., Ltd., Dezhou, Shandong, China), each dissolved in 10 mL of sterilized water at a final concentration of 0.5%. These additives were then added to the test vessels. Additionally, sterile water was added to the control vessels. Fully grown B. adolescentis 4-2 was added to designated vessels at 0.1% (v/v) of the total volume.
Cell-free supernatants were obtained by membrane filtration (0.45 µm) from
both bacterial and fecal cultures. GABA concentrations were quantified using HPLC
(1100 Series, Agilent Technologies, Santa Clara, CA, USA) equipped with a fluorescence detector
(excitation at 350 nm and emission at 450 nm) and a COSMOSIL packed column
(5C18-MS-II, 3.0 mm ID
DNA extraction and next-generation sequencing (NGS) were performed as previously described [20]. Raw NGS data were analyzed using QIIME 2 (version 2020.2; QIIME 2 Development Team) [24]. Additionally, bar plots displaying microbial composition were generated at both the phylum and species levels based on identified operational taxonomic units (OTUs).
Apart from the KUHIMM culture, liquid MRS culture (1 mL) was disrupted by
sonication. Sonication was performed using a Fisherbrand™
Sonicator with probe (Fisher Scientific, Waltham, MA, USA) for cell disruption
prior to enzyme assay. The disrupted cells were then centrifuged at 13,000 rpm
for 15 min at 4 °C, and the supernatant (cell-free extract) was used for
the
Enzymatic activity was quantified based on the concentration of p-nitrophenol released, using a standard curve prepared from known concentrations of p-nitrophenol (Sigma-Aldrich, Cat# N7660, St. Louis, MO, USA), under identical assay conditions.
All statistical analyses were performed using GraphPad Prism (version 9;
GraphPad Software, San Diego, CA, USA). Data were presented as mean
To identify strains with strong GABA-producing capabilities, a collection of
Bifidobacterium isolates, comprising both fecal-derived and commercially
available strains (Table 1), was screened. Consistent with previous findings
[25], GABA production was found to be markedly strain-specific rather than
species-dependent. Among the B. adolescentis isolates, strain 4-2
demonstrated the highest GABA yield, achieving 1.4
| Bacterial strain | GABA (g/L) |
Source | Origin |
| B. dentium JCM 1195 | 2.00 |
JCM company | Human |
| B. adolescentis 4-2* | 1.40 |
Kobe, Japan | Adult human |
| B. adolescentis* | 0.60 |
Kobe, Japan | Adult human |
| B. adolescentis 4-16* | 0.30 |
Kobe, Japan | Adult human |
| B. adolescentis 3-117* | 0.20 |
Kobe, Japan | Adult human |
| B. adolescentis JCM 7042 | 0.10 |
JCM company | Human |
| B. adolescentis 12-111* | 0.10 |
Kobe, Japan | Adult human |
| B. ruminantium JCM 8222 | 0.18 |
JCM company | Animal |
| B. catenulatum* | 0.20 |
Kobe, Japan | Human |
| B. adolescentis JCM 1275 | ND | Kobe, Japan | Human |
| B. animalis subsp. animalis JCM 1190 | 0.00 |
JCM company | Animal |
| B. animalis subsp. lactis JCM 10602 | 0.00 |
JCM company | Animal |
| B. indicum JCM 1302 | 0.00 |
JCM company | Human |
| B. longum 105A | 0.00 |
JCM company | Human |
| B. longum subsp. infantis JCM 1222 | 0.00 |
JCM company | Human |
| B. breve JCM 1192 | 0.00 |
JCM company | Human |
| B. minimum JCM 5821 | 0.00 |
JCM company | Human |
Note: Strains marked with an asterisk (*) are fecal isolates, ND refers to non-detected.
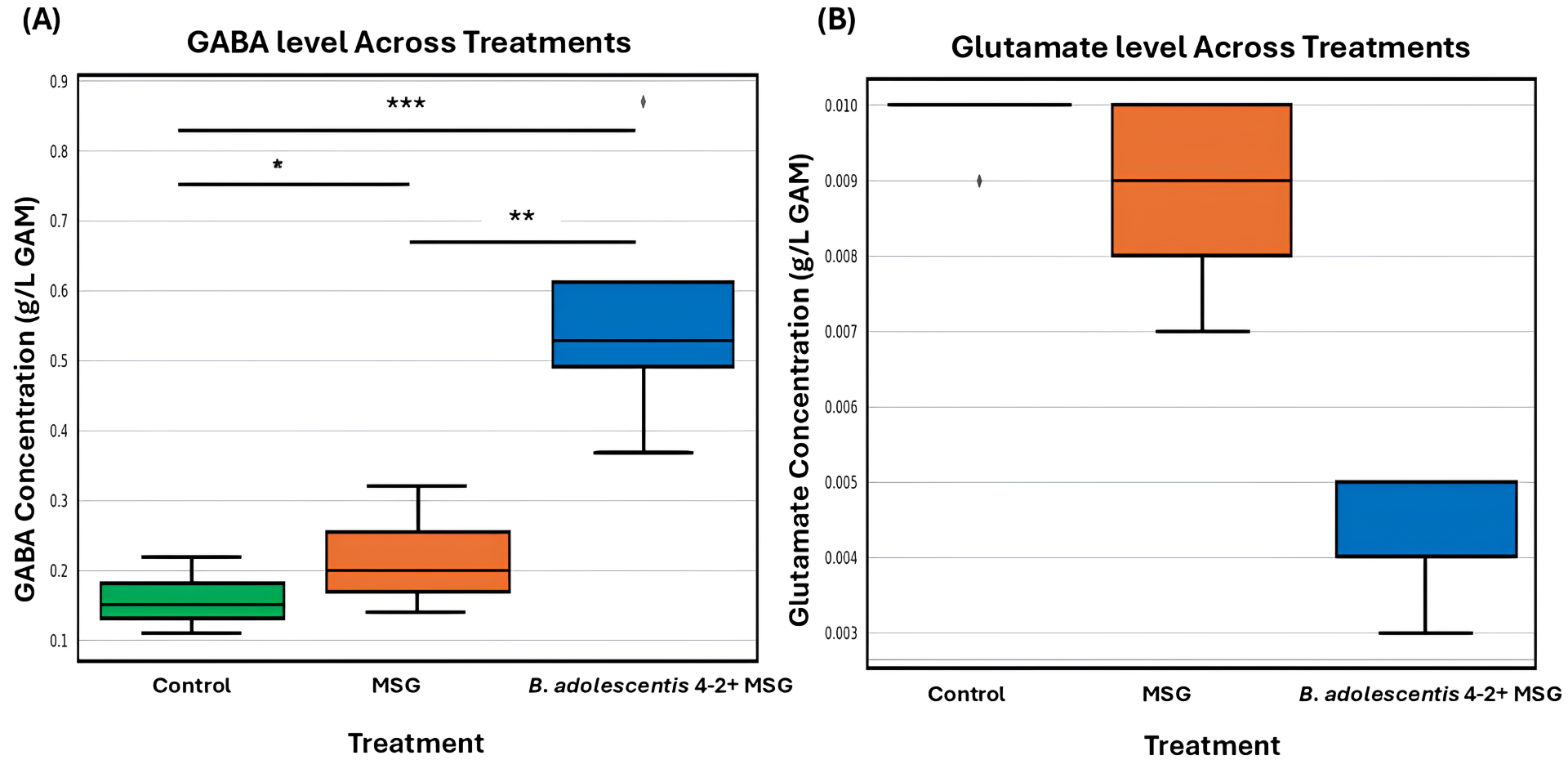
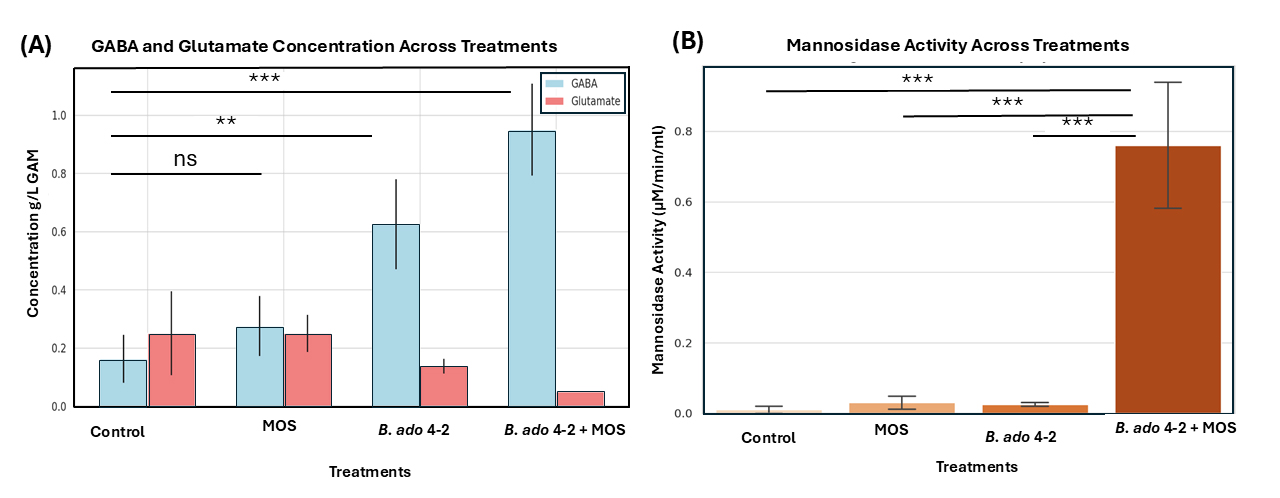
To further assess its functional potential, B. adolescentis 4-2 was introduced into the KUHIMM, an in vitro system inoculated with human fecal microbiota from donors exhibiting low baseline GABA levels. Monosodium glutamate (MSG) was provided as the precursor substrate. As depicted in Fig. 1A,B, the presence of B. adolescentis 4-2 resulted in a significant increase in fecal GABA concentration compared with both the control and MSG-only conditions. Concurrently, Fig. 1B illustrates a marked reduction in glutamate levels in the B. adolescentis 4-2 + MSG group, indicating efficient microbial conversion of glutamate to GABA. These results underscore the strain’s capacity to modulate key gut neurotransmitter levels by elevating GABA while reducing glutamate, thereby supporting its application in microbiome-based interventions aimed at the gut-brain axis.
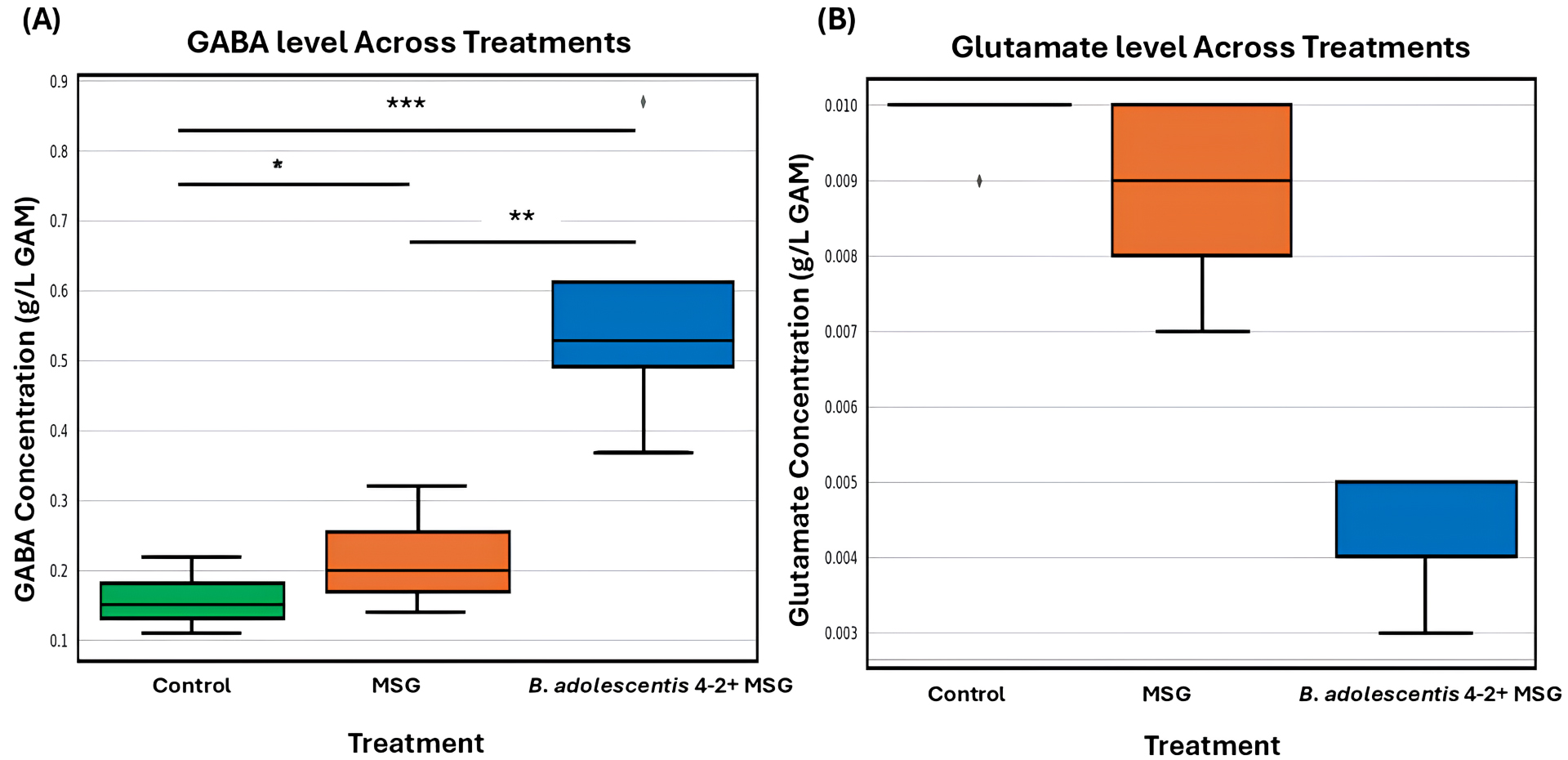 Fig. 1.
Fig. 1.
Gamma-aminobutyric acid (GABA) and glutamate concentrations across different fecal culture conditions. GABA (A) and glutamate
(B) concentrations were measured in fecal culture samples obtained from five
individual volunteers under three conditions: Control (green), MSG
supplementation (orange), and Bifidobacterium adolescentis 4-2 with MSG
(blue). Boxplots depict the data distribution across volunteers, where the horizontal line represents the median, the box represents the interquartile range (IQR), and whiskers extend to
1.5
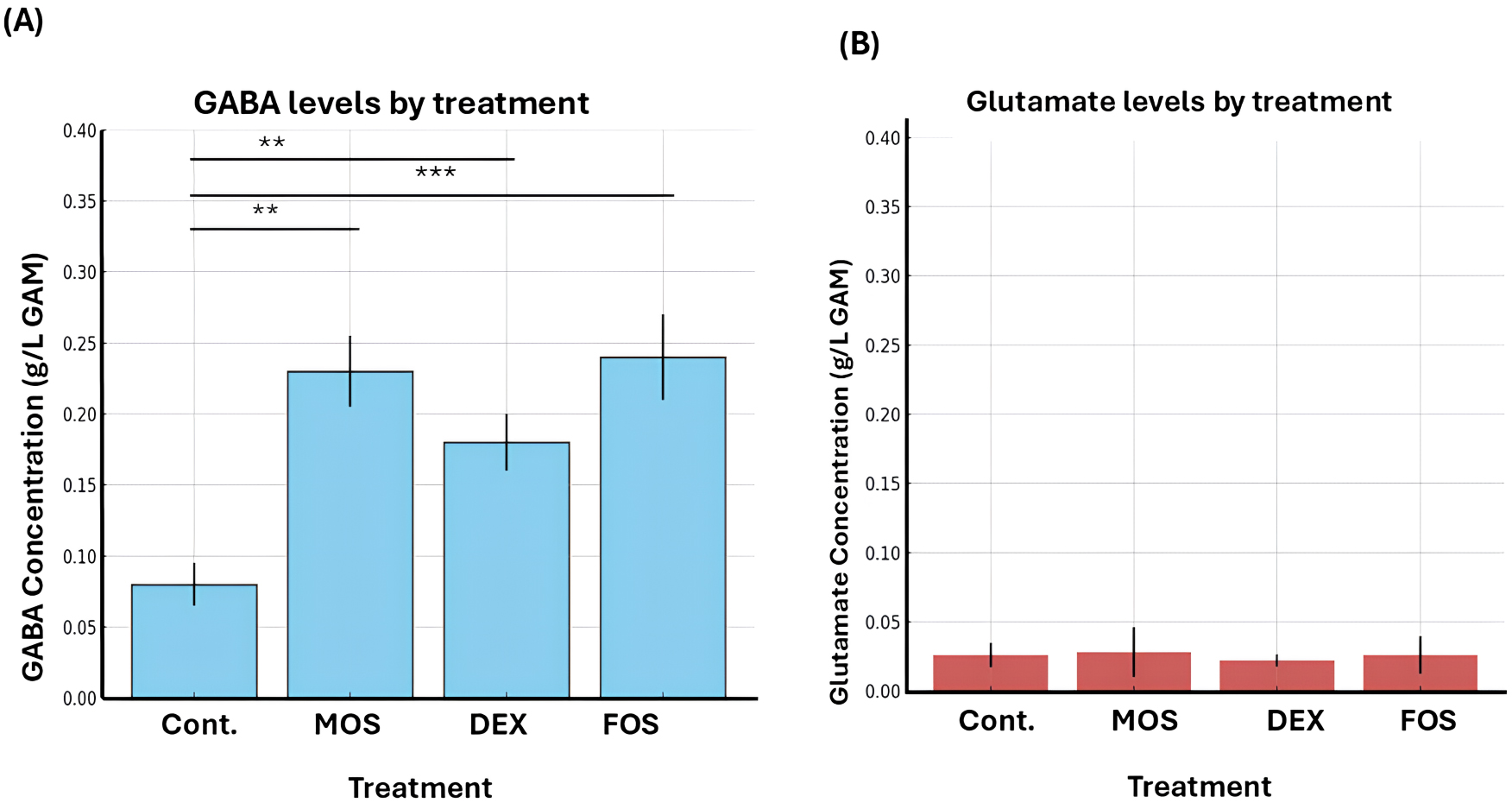
To further investigate the effect of dietary components on microbial activity,
prebiotics including MOS, DEX, and FOS were introduced. Supplementation with
these oligosaccharides resulted in a significant increase in GABA levels across
all treatment groups compared with the control (untreated) group. Among the
evaluated prebiotics, MOS and FOS produced the highest GABA concentrations, with
statistically significant differences (p
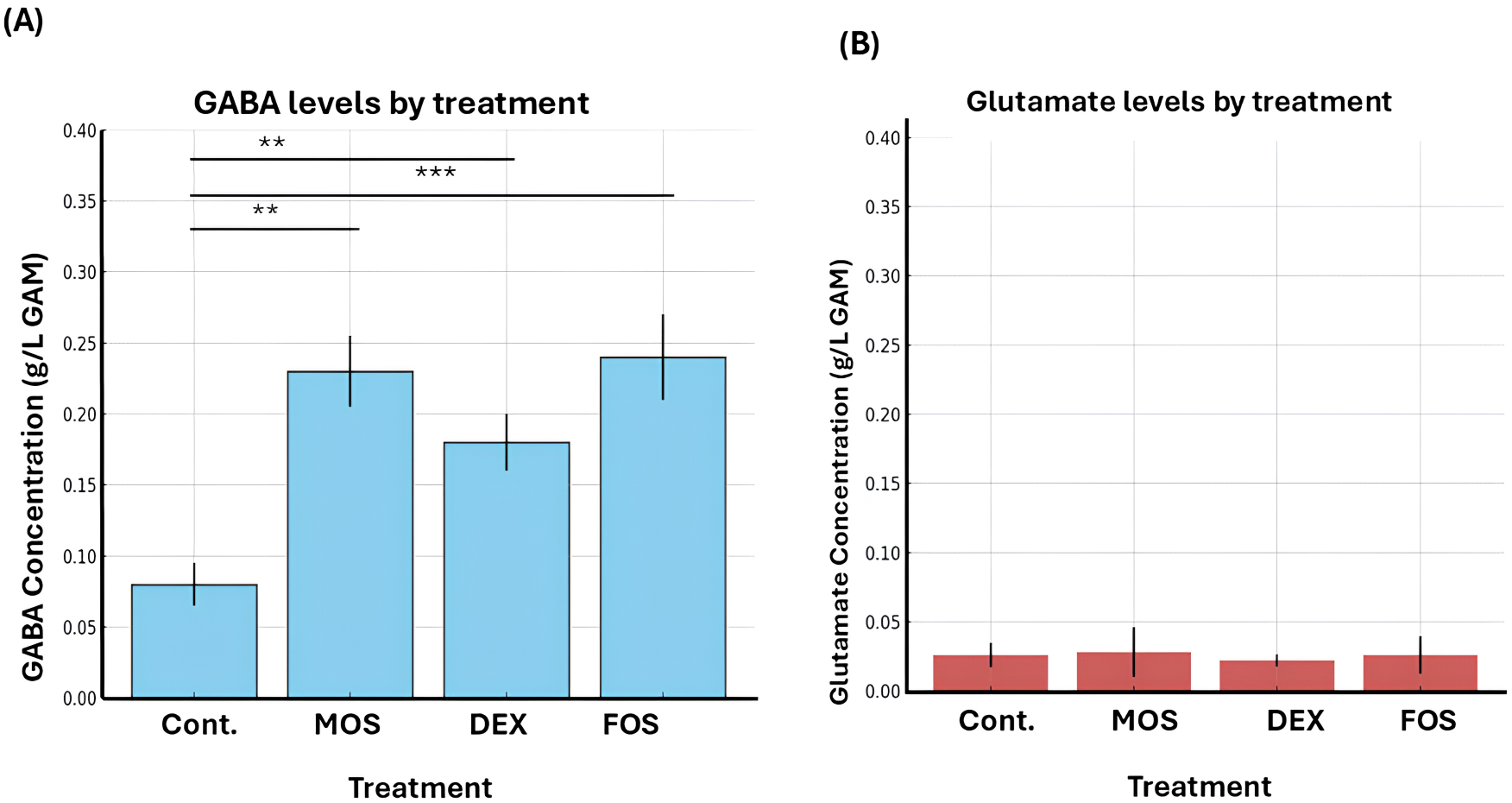 Fig. 2.
Fig. 2.
Effect of different treatments on gamma amino butyric acid
(GABA) and glutamate concentrations in the gut microbiota culture. Bar graphs
show the mean concentrations (
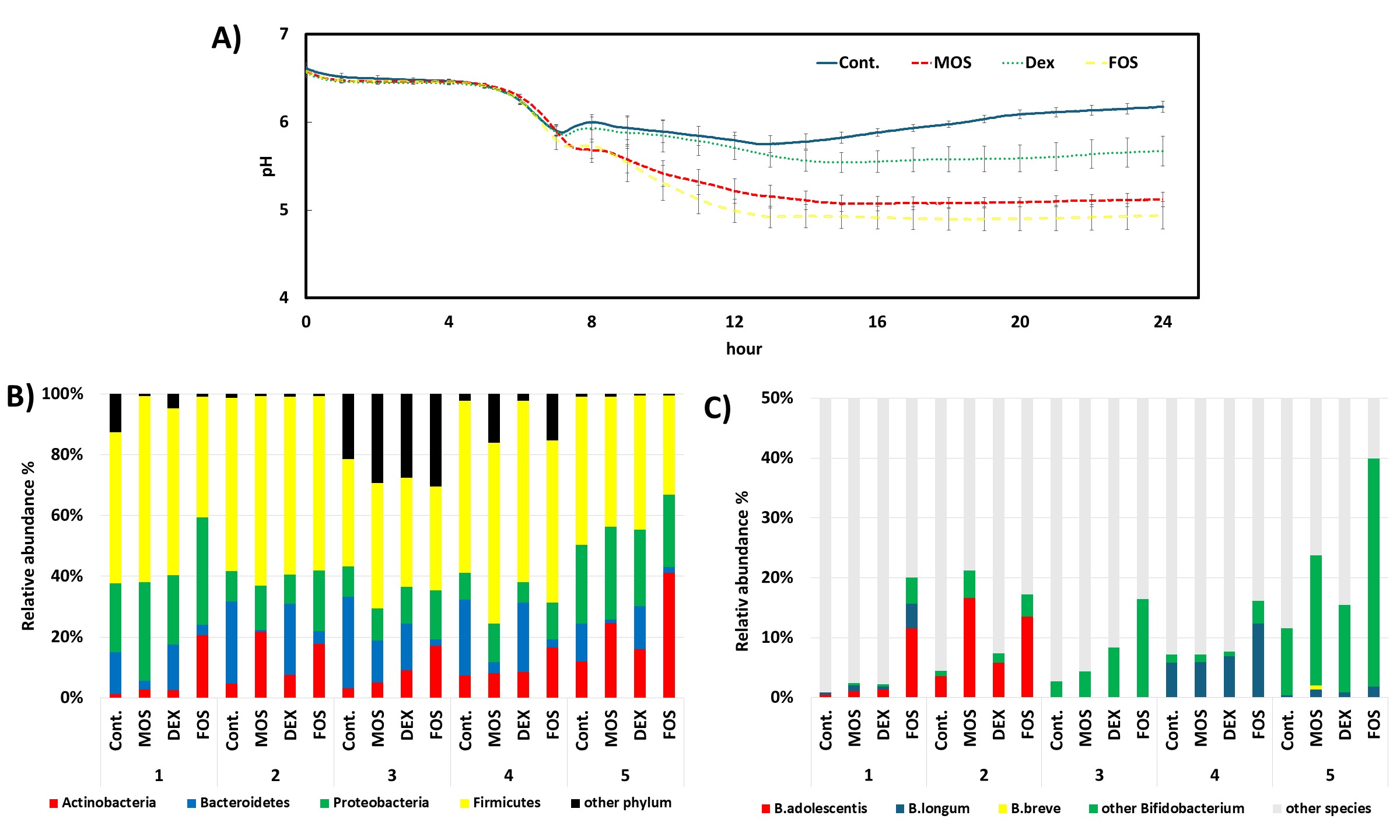
During the 24-hour fermentation, pH values decreased more markedly and remained lower in cultures treated with oligosaccharides, particularly MOS and FOS, compared with the control, which showed only a slight drop followed by partial recovery (Fig. 3A). This sustained acidification indicates enhanced microbial fermentation activity.
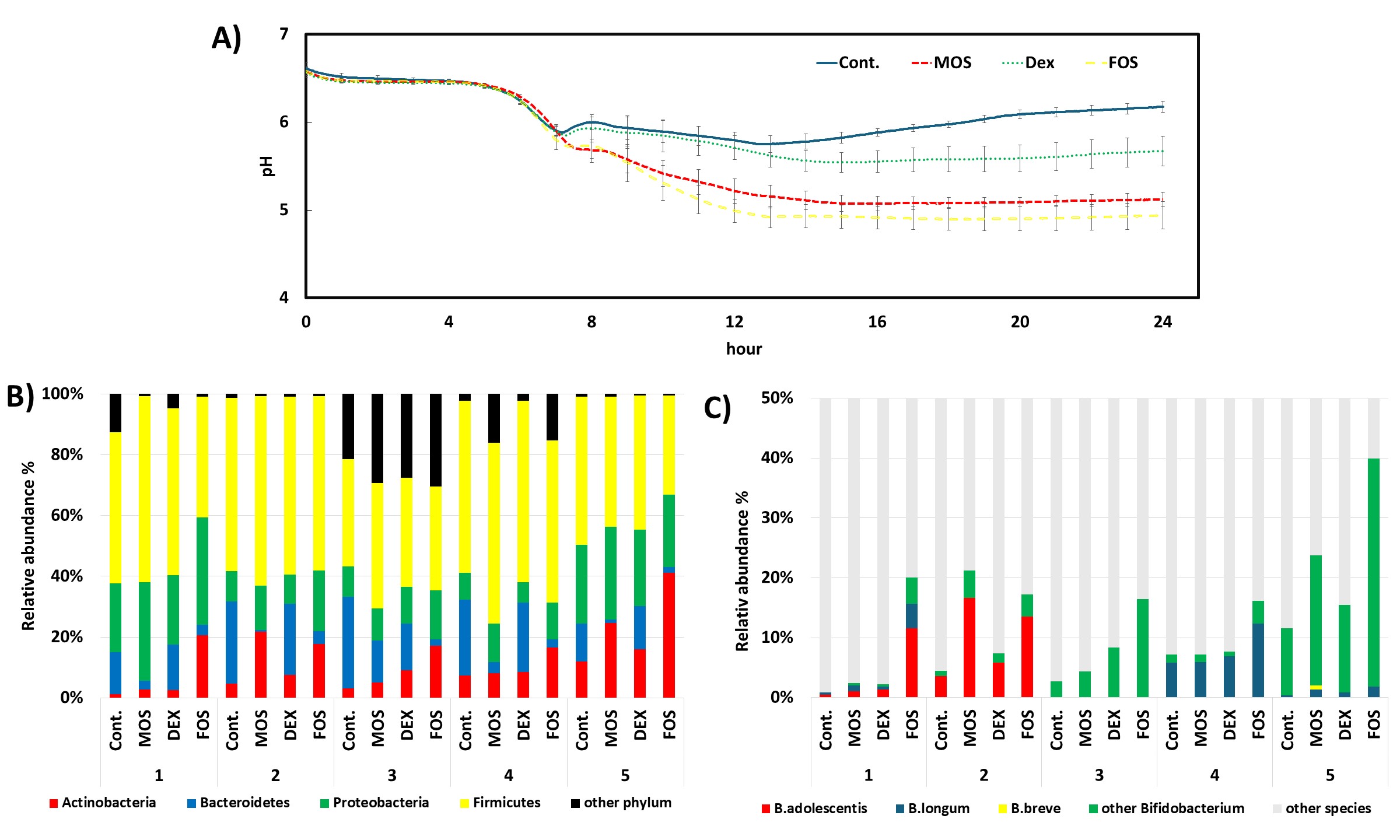 Fig. 3.
Fig. 3.
Impact of different treatments on gut fermentation and microbial composition. (A) pH dynamics over 24 hours of fermentation under different treatments: Control (Cont.), mannooligosaccharides (MOS), dextrin (DEX), and fructooligosaccharides (FOS). A rapid decline in pH occurred in all groups within the first 8 hours, followed by partial recovery in the control and DEX groups, whereas the pH remained low in the MOS and FOS treatments, indicating enhanced fermentation activity. (B) Phylum-level relative abundance of gut microbiota across five biological replicates per treatment. Firmicutes and Proteobacteria were dominant across most treatments. In most donners, MOS and FOS treatments exhibited increased proportions of Actinobacteria and Proteobacteria compared to the control. (C) Relative abundance of Bifidobacterium species. B. adolescentis and other Bifidobacterium species were particularly enriched in MOS and FOS treatments, with noticeable inter-individual variation. B. longum and B. breve were detected in lower but variable proportions.
Microbiota analysis indicated an enrichment of Actinobacteria across all treatment groups, with the greatest increase observed under MOS and FOS conditions (Fig. 3B). Within this phylum, Bifidobacterium species, particularly B. adolescentis, became more predominant, especially in donor samples 1 and 2 (Fig. 3C). This taxonomic shift aligns with previous reports demonstrating B. adolescentis’s preference for oligosaccharides and its ability to synthesize GABA [25, 26].
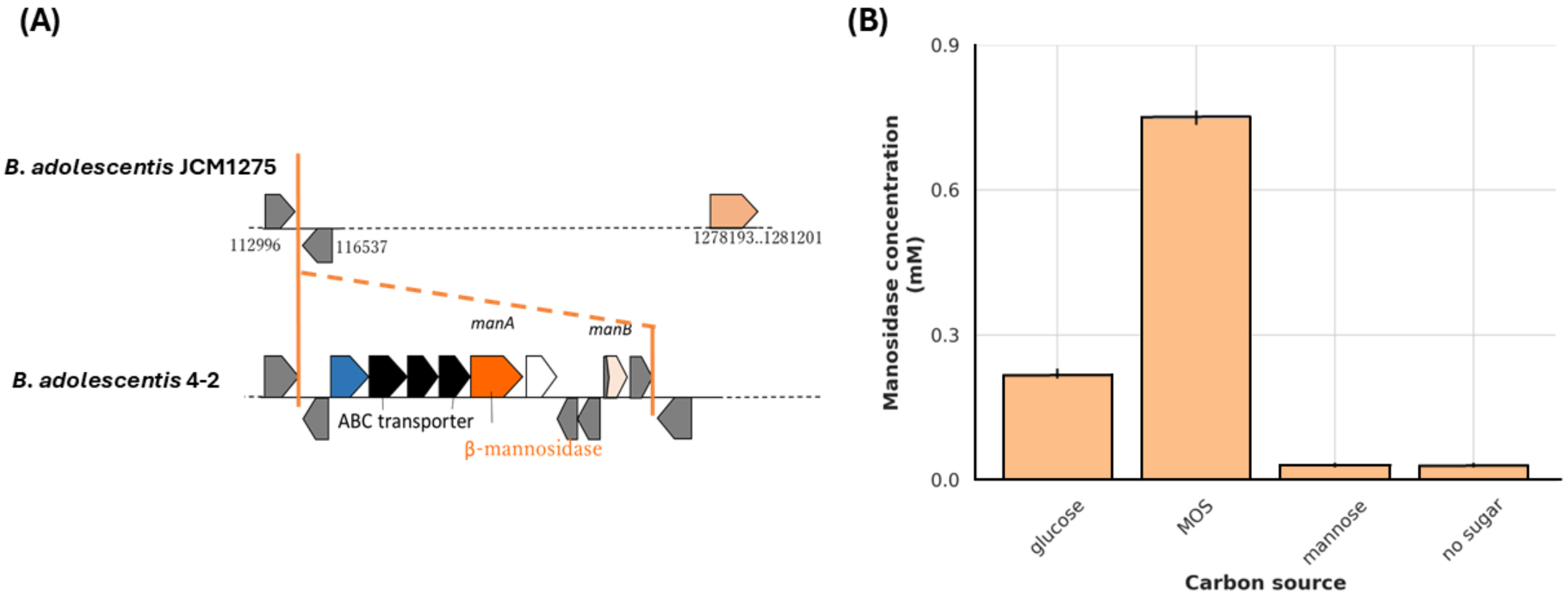
Mannan hydrolysis requires multiple synergistic glycoside hydrolases. Genomic
analysis revealed that B. adolescentis 4-2 possesses two key genes
involved in this process: manB, encoding a
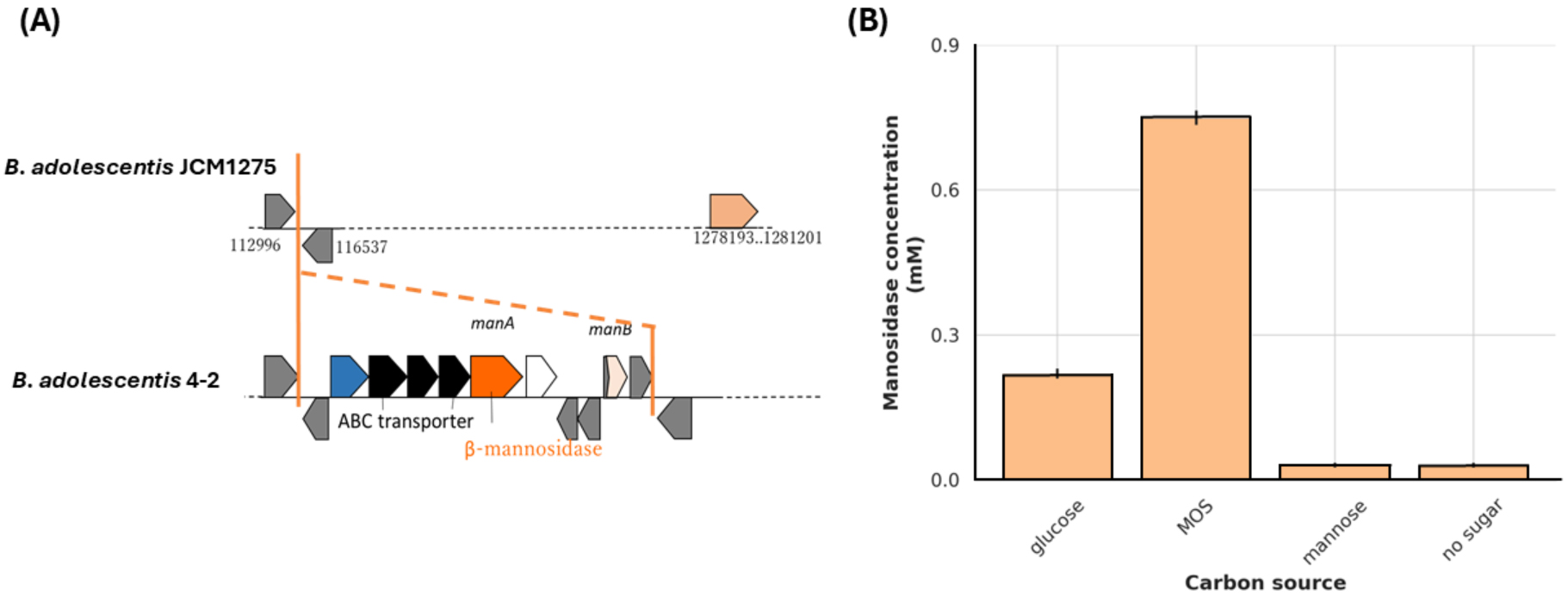 Fig. 4.
Fig. 4.
To assess the potential symbiotic effect between B. adolescentis 4-2
and oligosaccharides, a gut microbiota model was employed using fecal samples
characterized by low baseline GABA levels. Four experimental setups were tested:
control (no additives), 0.5% MOS alone, B. adolescentis 4-2 alone, and
a combination of 0.5% MOS with B. adolescentis 4-2. GABA concentrations
increased progressively across the groups in the following order: MOS, B.
adolescentis 4-2, and MOS + B. adolescentis 4-2, while glutamate levels were similar to those in the control group (Fig. 5A). The combined treatment produced the highest
GABA yield, demonstrating a synergistic interaction between the probiotic strain
and MOS under gut-simulating conditions. To confirm this symbiosis,
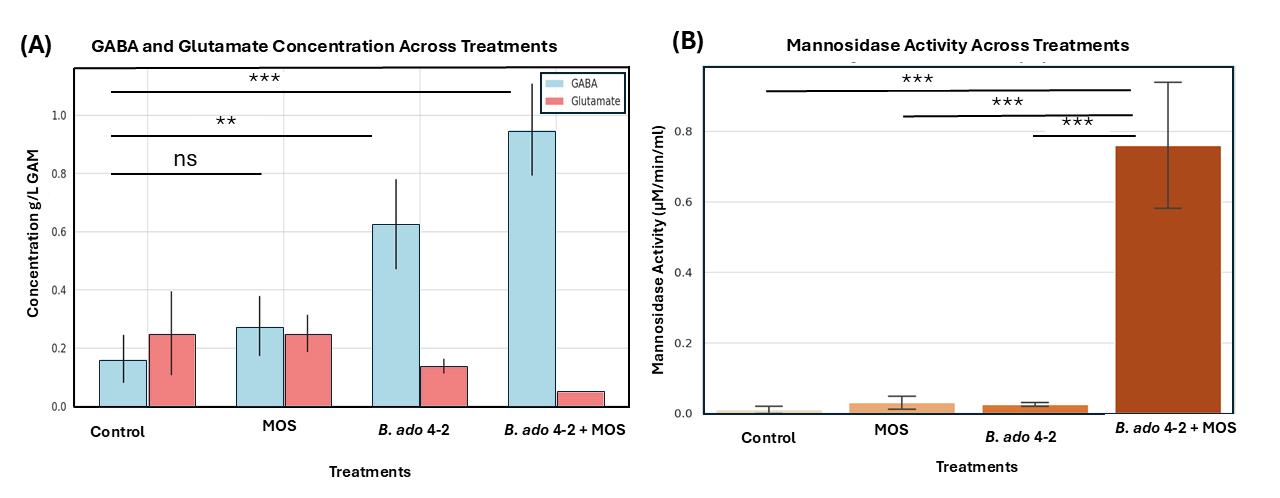 Fig. 5.
Fig. 5.
Effects of treatments on gamma amino butyric acid (GABA) and
glutamate concentrations and mannosidase activity. (A) Bar graph displaying the
concentrations of GABA (blue bars) and glutamate (red bars) across four treatment
groups: Control, MOS (mannan oligosaccharides), B. adolescentis 4-2, and
B. adolescentis 4-2 + MOS. GABA concentrations increased significantly
in both the B. adolescentis 4-2 and B. adolescentis 4-2 + MOS
treatments, accompanied by corresponding reductions in glutamate levels. (B) Bar
graph showing mannosidase activity (measured in µM/min/mL) in the
same groups. The highest enzymatic activity was recorded in the B.
adolescentis 4-2 + MOS group, indicating a synergistic effect. Statistical
significance was determined using one-way ANOVA followed by Tukey’s multiple
comparison test, with comparisons noted above the bars (ns = not significant; **
= p
This study elucidates the synergistic interaction between the GABA-producing
strain B. adolescentis 4-2 and the prebiotic MOS in enhancing GABA
production within an in vitro model of the colonic microbiota. Our
findings align with and extend previous reports that identified
Bifidobacterium species as key contributors to gut-derived GABA [2, 3, 27]. In particular, the strain specificity of GABA synthesis observed in our
screening is consistent with earlier studies demonstrating heterogeneity in GABA
production among strains of B. adolescentis and B. breve [27, 28, 29]. B. adolescentis strain 4-2 not only exhibited superior GABA
biosynthesis in monoculture but also retained its functional capacity within the
complex fecal microbial community simulated by KUHIMM. Although B.
dentium JCM 1195 showed higher GABA production in monoculture, B.
adolescentis 4-2 was prioritized for synbiotic development because it was
isolated from healthy adults, performed stably in the KUHIMM, and possesses
Certain gut microbes, such as Bacteroides species, possess GABA transaminase activity, which can degrade GABA and thereby influence net concentrations observed in fecal cultures [32]. The introduction of MSG as a precursor facilitated robust microbial conversion of GABA. Corroborating studies have shown that exogenous glutamate enhances GABA production in various lactobacilli and Bifidobacteria [33]. Notably, the concurrent depletion of glutamate and the rise in GABA levels in the B. adolescentis 4-2 treatment group highlight the enzymatic activity of glutamate decarboxylase (GAD), a key enzyme responsible for this metabolic conversion, which has been previously characterized in other probiotic strains [34].
Beyond monoculture effects, our study demonstrated that MOS supplementation significantly boosts fecal GABA levels and supports the proliferation of Actinobacteria, particularly Bifidobacterium spp., which aligns with prior research showing selective bifidogenic effects of MOS and similar oligosaccharides [12, 15]. Notably, MOS-driven acidification of the culture medium is indicative of active microbial fermentation, a hallmark of oligosaccharide metabolism [13, 14]. The enrichment of B. adolescentis in MOS-treated cultures, especially in low-GABA fecal donors, highlights the prebiotic potential of MOS to selectively enhance strains capable of psychobiotic function. While these findings emphasize the selective bifidogenic effects of MOS, it is important to note that other gut commensals may utilize MOS. In particular, members of the genera Bacteroides and Firmicutes harbor diverse glycoside hydrolases and transport systems that enable the degradation and assimilation of mannan-derived oligosaccharides [22]. Such taxa often act as primary degraders within the gut ecosystem, breaking down complex polysaccharides into smaller metabolites that can be cross-fed to other microorganisms [16]. Consequently, competitive utilization of MOS by these microbes could reduce the relative availability of the substrate for Bifidobacteria, potentially influencing the magnitude of GABA production [22, 35]. On the other hand, cross-feeding interactions may generate complementary metabolic outputs, such as short-chain fatty acids, which could synergize with GABA to modulate host physiology [15, 36]. Previous reports have demonstrated that the balance between competition and cooperation for prebiotic substrates strongly shapes microbial community structure and functional outcomes both in vitro and in vivo [15, 16, 35, 37].
The synergistic increase in GABA production observed with the combined
administration of B. adolescentis 4-2 and MOS marks a pivotal
contribution of this study. While earlier works have proposed the benefit of
combining prebiotics with probiotics to create synbiotics [38], our study is
among the first to mechanistically link this synergy to enhanced GABA
biosynthesis in a fecal microbiota model. This finding is further supported by
the elevated
Importantly, the application of the KUHIMM allowed for controlled evaluation of microbial and metabolic shifts over time. This model mimics key aspects of the human colon environment, providing a reproducible platform to test gut-derived bioactivity and overcoming limitations associated with in vivo variability [21, 22]. While KUHIMM provides a reproducible platform to assess microbial and metabolic responses, it does not capture host-microbe interactions, absorption, signaling, or immune responses, which should be investigated in subsequent animal and human intervention studies.
In conclusion, the synergistic increase in GABA production observed with B. adolescentis 4-2 and MOS illustrates the potential of targeted synbiotic strategies to modulate gut neurochemistry under in vitro conditions. These findings demonstrate the potential of MOS and B. adolescentis 4-2 to synergistically enhance GABA production in the KUHIMM. Further studies in animal and human systems are required to establish whether these effects translate to clinical outcomes.
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
HA and TS designed the research study. TM, YB and HA performed the research. YB, HA, and MA conducted experiments and analyzed the data. HA wrote the first draft of the manuscript. All authors contributed to editorial revisions of the manuscript. All authors read and approved the final version of the manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work. The late Professor TS contributed to the conception and execution of this study. His scientific insight and dedication were instrumental, and this work is respectfully dedicated to his memory.
The study was conducted in accordance with the guidelines of the Declaration of Helsinki. The protocol was approved by the institutional ethics review board of Gifu University (Certificate No. 2019-283). Written informed consent was obtained from all participants.
Not applicable.
This research received no external funding.
The authors declare no conflict of interest.
During the preparation of this work, the authors used AI assistance to check spelling and grammar. After that, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.