
 , Valentina La Sorsa 2, Iole Macchia 3, Sara Giancristofaro 4, Michela Pagnotta 4, Donatella Pietraforte 1, Massimo Sanchez 1, Maurizio Zini 1, Giada Cairo 4, Monica Fabiani 4, Monica Monaco 4, Maria Del Grosso 4, Romina Camilli 4, Giulia Errico 4, Sergio Pandolfi 5, Luigi Valdenassi 5, Marianno Franzini 5, Alessandra Cenci 1,*
, Valentina La Sorsa 2, Iole Macchia 3, Sara Giancristofaro 4, Michela Pagnotta 4, Donatella Pietraforte 1, Massimo Sanchez 1, Maurizio Zini 1, Giada Cairo 4, Monica Fabiani 4, Monica Monaco 4, Maria Del Grosso 4, Romina Camilli 4, Giulia Errico 4, Sergio Pandolfi 5, Luigi Valdenassi 5, Marianno Franzini 5, Alessandra Cenci 1,*
1 Core Facilities, Italian National Institute of Health, 00161 Rome, Italy
2 Research Coordination and Support Service, Italian National Institute of Health, 00161 Rome, Italy
3 Department of Oncology and Molecular Medicine, Italian National Institute of Health, 00161 Rome, Italy
4 Department of Infectious Diseases, Italian National Institute of Health, 00161 Rome, Italy
5 Italian Scientific Society of Oxygen Ozone Therapy (SIOOT), Bergamo and High School Master of Oxygen Ozone Therapy, University of Pavia, 27100 Pavia, Italy
Abstract
Methicillin-resistant Staphylococcus aureus (MRSA) poses a major challenge in clinical settings due to limited susceptibility to conventional antibiotics. Ozone (O3), a naturally occurring molecule composed of three oxygen atoms, exhibits strong antimicrobial properties against a wide spectrum of microorganisms, including both Gram-positive and Gram-negative bacteria. The antibacterial efficacy of O3 against pathogenic bacteria has been demonstrated both in vivo and in vitro, although the mechanisms of action remain poorly understood. Notably, O3 has proven effective against MRSA, underscoring the potential of O3 as a valuable adjunctive therapy for managing resistant bacterial infections.
This study aimed to confirm the antibacterial effects of oxygen–O3 (O2–O3) therapy on MRSA strains and to investigate the associated immunomodulatory and antioxidant properties in human peripheral blood mononuclear cells (PBMCs) treated in vitro with MRSA.
PBMCs from healthy donors were initially exposed to heat-killed or O3-treated MRSA strains at 37 °C for 2 hours, then treated with 40 μg/mL O3 for an additional 24 hours to simulate O3 therapy. Subsequently, cytokine release, antioxidant and pro-oxidant gene expression, reactive oxygen species production, and redox-related biomarker levels were assessed.
Our findings demonstrated that O3, at various concentrations (20–60 μg/mL) within the therapeutic range (20–80 μg/mL), exerts a potent antibacterial effect against MRSA, resulting in complete loss of bacterial colonies, in the absence of antioxidants in the medium. Using a multidisciplinary approach encompassing biochemical, molecular, and cell biology techniques, we observed that O3 exerts immunomodulatory and antioxidant effects on PBMCs exposed to O3 and stimulated with MRSA as early as 1 hour after stimulation, with these effects persisting for up to 48–72 hours, regardless of whether the bacteria were inactivated by O3 or heat.
We successfully demonstrated that O3 exerts an early antibacterial effect, followed by antioxidant activity and immunomodulatory properties, as confirmed by multiple integrated analyses. These findings highlight the potential of O3 as a promising strategy to combat antibiotic resistance and support therapeutic protocols.
Keywords
- ozone
- methicillin-resistant Staphylococcus aureus
- cytokines
- immune response
- antioxidants, anti-inflammatory agents
Antimicrobial resistance (AMR) is a growing global health concern underscoring the urgent need for innovative therapeutic approaches and robust antimicrobial stewardship (WHO 2023, https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance) [1, 2].
Among resistant pathogens, methicillin-resistant Staphylococcus aureus (MRSA) remains a major clinical challenge causing infections ranging from minor skin conditions [3] to severe systemic diseases such as sepsis, endocarditis, and pneumonia [4, 5].
Ozone (O3), a triatomic oxygen molecule, has historically been used for water disinfection and the topical treatment of wound infections [6].
When properly administered, oxygen–O3 (O2–O3) therapy is
considered safe and non-toxic, as shown by in vivo studies [7]. At low
concentrations (
Moreover, O3 has been shown to potentiate the antimicrobial efficacy of antibiotics such as vancomycin against MRSA by increasing cell wall permeability and inhibiting biofilm formation [12, 13, 14]. In addition to the antibacterial properties of O3, antiviral efficacy has been demonstrated against Herpes zoster [15], Herpes simplex [16], and SARS-CoV-2, primarily through viral load reduction and immunomodulation via the regulation of cytokine signaling [17, 18, 19, 20, 21, 22, 23, 24].
Moreover, O3 therapy within the therapeutic range of 20–80 µg/mL has been shown to exert immunomodulatory and antioxidant effects that contribute to the overall associated antimicrobial potential [25, 26, 27].
Meanwhile, O3 exhibits antibacterial activity through both direct and indirect mechanisms. The direct antibacterial effects of O3, relevant primarily to in vitro applications and surface disinfection, occur through three main pathways: (a) induction of oxidative stress in bacterial cells leading to lipid peroxidation and membrane damage, culminating in cell lysis [26, 27, 28, 29]; (b) oxidation and inactivation of essential bacterial enzymes, thereby disrupting key metabolic functions [30, 31]; (c) oxidative damage to bacterial DNA, impairing replication and repair processes, and inhibiting bacterial proliferation [32].
In addition, O3 may exert indirect antibacterial activity in vivo through immunomodulatory mechanisms, by stimulating cytokine release and immune cell activation [28, 29], enhancing host defense mechanisms against microbial infections [31, 32, 33, 34].
Upon administration, medical O3 rapidly reacts with lipids, generating
reactive oxygen species (ROS) including hydrogen peroxide (H2O2) and
lipid peroxides (LOPs). This reaction leads to the formation of 4-hydroxynonenal
(4-HNE), a key lipid peroxidation product of
In both in vivo and in vitro models, exposure to low O3
concentrations induces mild intracellular and mitochondrial oxidative
stress–triggering hormetic and mitohormetic signaling through the
NRF2–Keap1–ARE pathway [36, 37, 38, 39]. ROS generated by O3, also activate the
NRF2, which translocates to the nucleus and binds antioxidant response elements
(AREs) in complex with MAF transcription factors, inducing the expression of
antioxidant and anti-inflammatory genes, including superoxide dismutase
(SOD), catalase (CAT), glutathione peroxidase (GPx),
glutathione reductase (GR), glutathione-S-transferase (GST),
NAD(P)H quinone dehydrogenase 1 (NQO1), heme oxygenase 1 (HO-1)
and the thioredoxin/thioredoxin reductase system (TXN/TXNRD) [37, 40].
Simultaneously, NRF2 activation suppresses the nuclear factor
kappa-light-chain-enhancer of activated B cells (NF-
Additionally, O3 may reduce inflammation through two complementary
mechanisms: first, it prevents the activation of the NLR family pyrin
domain-containing 3 (NLRP3) inflammasome, a key driver of interleukin-1
Thus, this study aimed to explore the mechanisms underlying the in vitro action of O3 against MRSA. We first examined the effect of O3 on MRSA
viability, and then, using an integrated biochemical, cellular, and molecular
approach, investigated the immunomodulatory and antioxidant effects of O3 on
PBMCs as a human-relevant model. Specifically, we assessed cytokine production by
PBMCs stimulated with O3 or heat-inactivated MRSA and subsequently treated
with O3 to evaluate ozone anti-inflammatory and redox homeostasis–restoring
properties. These effects were studied, in a time-dependent manner, by assessing
the: (i) production of pro-inflammatory cytokines using a Cytokine Bead Array
(CBA) to detect TNF-
Our findings demonstrated that O3 exerts both direct antibacterial effects against MRSA and potent immunomodulatory and antioxidant actions on human PBMCs. By activating the NRF2 pathway and suppressing key inflammatory mediators, O3 promotes redox homeostasis and an anti-inflammatory cellular phenotype, supporting the potential use of O3 as an adjunctive therapeutic strategy for antimicrobial-resistant infections.
The experiments were conducted employing the MEDICAL 99 IR Multiossigen device (Gorle, Bergamo, Lombardy, Italy), provided by SIOOT. The device is certified by the Notified Body of the Ministry of Health as Class IIa under Directive 93/42/EEC.
For the experiments, we used the MRSA USA300 reference strain from our collection of isolates derived from clinical specimens. The growth and inactivation of MRSA were conducted in Tryptic Soy Broth (TSB, OXOID) overnight (O/N) at 37 °C. The O/N bacterial culture was diluted 1:100 in fresh TSB broth and incubated until the exponential growth phase was reached (OD600 = 0.6). The bacterial culture was then processed using two protocols, namely one-step and multi-step, prior to O3-induced bacterial killing. In the one-step approach, 5 mL of the bacterial culture (OD600 of 0.6) was washed and resuspended in phosphate-buffered saline (PBS) (v/v) before proceeding with O3 killing. The bacterial culture was treated, in independent experiments, with three O3 concentrations (20 µg/mL, 40 µg/mL and 60 µg/mL, respectively) for 5 minutes (MRSA O3I). To verify whether bacterial killing was successful, 10-fold serial dilutions of the bacterial suspension from each treatment condition, were prepared, plated on Tryptic Soy Agar (TSA; Oxoid Ltd., Basingstoke, Hampshire, UK) and incubated at 37 °C O/N. The absence of bacterial growth was taken as evidence of the efficacy of the killing method.
In the multi-step protocol, 5 mL of the bacterial culture (OD600 of 0.6), was washed twice in RPMI 1640 and resuspended in ozonated RPMI 1640 (v/v) obtained by a prior O3 treatment at 60 µg/mL for 20 minutes. RPMI 1640 ozonation was necessary to reduce the antioxidant effect of the medium. The bacterial suspension in ozonated RPMI 1640 was subsequently treated with an O3 concentration of 60 µg/mL at 20-minute intervals for at least 1 hour, depending on the initial bacterial concentration. After O3 treatment, 200 µL of the bacterial suspension was plated at serial dilutions either on TSA or on Chromid MRSA SMART plates (bioMérieux SA, Marcy-l’Étoile, France) to assess killing activity.
In addition, two further bacterial-killing experiments were performed with a bacterial cell density (107 colony-forming units (CFU)/mL) using heat inactivation or gamma-ray irradiation. In the first experiment, the bacterial culture was washed in PBS and heat-inactivated at 100 °C for 10 minutes. In the second experiment, bacterial culture was treated with O3 for up to five hours according to the multi-step protocol, and the surviving rare colonies were subsequently subjected to gamma-ray irradiation over the weekend at a dose of 2700 Gy [46] as described in more detail in the Results.
Gamma irradiations from a 137Cs source were performed at the Istituto Superiore di Sanità (ISS, Rome, Italy) using the Gammacell Exactor 40 (Nordion) [47].
The MRSA bacterial culture (107 CFU/mL) was heat-inactivated at 100 °C, to prepare the control (MRSA HI) [48]. To confirm the lack of viable bacteria following heat inactivation and gamma-ray irradiation, 200 µL of the bacterial suspension was plated on TSA and incubated at 37 °C O/N.
The study was approved by the Ethics Committee of the Italian National Institute of Health (Istituto Superiore di Sanità, ISS), Rome, Italy (Protocol PRE BIO CE n. 0041072, November 24, 2021). Peripheral blood mononuclear cells (PBMCs) were isolated from healthy adult volunteers who provided written informed consent, in accordance with the Agreement between the Umberto I University Hospital (AOU Policlinico Umberto I, Rome) and the ISS regarding the “transfer of blood and its derivatives for laboratory use”, established under Resolution No. 0000853 of August 18, 2020. Only primary PBMCs were employed to ensure a physiologically relevant model; no immortalized or transformed cell lines were used.
PBMCs were isolated from buffy coats obtained from healthy donors (n = 3/5) released by the Transfusion Center of Rome Policlinico Umberto I, as previously described [49]. PBMCs were cultured in RPMI 1640 with 10% fetal bovine serum (FBS) and stimulated with MRSA at a multiplicity of infection (MOI) of 1, using bacteria inactivated by the methods that yielded the most effective bactericidal results. After two hours of stimulation, PBMCs were treated with O3 at 40 µg/mL to investigate, in vitro, the immunomodulatory and anti-inflammatory actions observed in vivo. Appropriate negative controls and Staphylococcal enterotoxin B (SEB) from Sigma-Aldrich (St. Louis, Missouri, USA; catalog no. S4881) positive controls were included in all experiments, conducted with both MRSA -O3-inactivated (MRSA O3I) and MRSA HI bacteria.
Our experiments were conducted using PBMCs isolated from freshly obtained buffy coats, ensuring that blood donations were processed within 24 hours of collection. This short interval between blood withdrawal and PBMC isolation is unlikely to allow the emergence of detectable changes in cellular morphology, surface marker expression, or mycoplasma contamination.
Cytokine production (TNF-
For intracellular cytokine analysis, we used a dried commercial panel (Duraclone IF T Helper Cell kit #C04666, Beckman Coulter, Brea, CA, USA) as the backbone, supplemented with liquid-format antibodies against CD8, IL-10, and a dead-cell exclusion marker, see Table 1 for details of the antibodies. Briefly, PBMCs from healthy donor buffy coats were cultured under three conditions: unstimulated (NT), stimulated with SEB at 2 µg/mL as a positive control, or stimulated with MRSA HI or MRSA O3I (at a 1:1 MRSA:PBMC ratio). After 2 hours, samples were treated with O3 (40 µg/mL or 20 µg/mL) and then incubated for 24 hours with Brefeldin A and Monensin (Becton Dickinson), which were added ~1-hour post-O3 treatment.
| Fluorescence channel | Fluorochrome | Marker | Catalog | Company | Formulation |
| FL-1 | FITC | IFN- |
C04666 | Beckman Coulter | Dried |
| FL-2 | PE | IL-10 | 501404 | Biolegend | Liquid |
| FL-5 | PC7 | IL-4 | C04666 | Beckman Coulter | Dried |
| FL-6 | APC | CD4 | C04666 | Beckman Coulter | Dried |
| FL-7 | AF700 | CD8 | 344724 | Biolegend | Liquid |
| FL-8 | AF750 | CD3 | C04666 | Beckman Coulter | Dried |
| FL-9 | PB | IL-17 | C04666 | Beckman Coulter | Dried |
| FL-10 | Aqua | LIVE/DEAD™ | L34966 | Invitrogen | Liquid |
Table 1 presents the integrated ICS T helper panel, in which antibodies
conjugated to different fluorochromes and directed against specific surface and
intracellular markers are listed. FL (Fluorescence channel), fluorochrome, and
antibody markers used in the integrated ICS T helper panel. Cells were stained
with antibodies targeting surface markers (CD3, CD4, CD8) and intracellular
cytokines (IFN-
The next day, cells were stained for dead cell exclusion, followed by staining with the Duraclone antibody cocktail, which incorporated anti-CD8 and anti-IL-10 monoclonal antibodies. Samples were acquired on a FACS Gallios instrument (Beckman Coulter, Brea, CA, USA), and data were analyzed using the Kaluza Analysis Software v.1.3 (Beckman Coulter Brea, CA, USA).
The gating strategy for multiparameter intracellular cytokine analysis within
the CD3+, CD4+, and CD8+ T gates is shown in Supplementary Fig. 1,
beginning with singlet and live-cell selection followed by T cell subset
identification and cytokine detection (IFN-
Total RNA was isolated from 3
Among the various methods available to assess ROS production or oxidative stress
in cells, we used the probe 2′,7′-Dichloro-dihydro-fluorescein diacetate (DCFH-DA). This is
a lipophilic, cell-permeable, non-fluorescent probe that, upon hydrolysis by
intracellular esterases, yields DCFH, which fluoresces upon oxidation by
intracellular ROS. Oxidation of the probe results from the action of several
types of peroxynitrite, hydroxyl radicals, nitric oxide, metal-derived oxidants
and other peroxides, and can be detected by flow cytometry. PBMCs were seeded at
1
After ozonation, the samples were centrifuged at 1200 rpm at room temperature for 10 minutes and the pellets were then suspended in PBS containing the vital dye Sytox Blue (Invitrogen™ Cat. n. S34857) at 1:1000 for flow cytometric analysis of ROS using the CytoFLEX LX (Beckman Coulter) with excitation and detection wavelengths of 488 nm and 535 nm, respectively. As negative controls, we used PBMCs and PBMCs plus DCF, with and without O3 insufflation, to estimate the number of unstained cells with autofluorescence in the green emission range and to quantify basal ROS levels in our sample. As a positive control, we incubated cells with 0.1 mM H2O2, with and without O3 insufflation.
ROS production was assessed every 20 minutes for 2 hours following O3 administration, with a final measurement taken at 24 hours. The gating strategy for ROS measurement with DCF is described in Supplementary Fig. 2.
PBMCs (2
We computed the between-samples Pearson’s correlation coefficients to check the
consistency among different subjects (samples) across all the measured variables.
The results indicated strong consistency among independent subjects, providing
confidence in the reliability of the data (Table 2). Therefore, given the
similarity among profiles, we considered the pattern reproducible despite the
small sample size (n = 3–5) used. A two-tailed paired Student’s
t-test was then performed according to the experimental design, with
p
| Techniques | Samples | ||||||
| DCF | Exp 5 | Exp 3 | Exp 4 | Exp 2 | Exp 1 | | |
| Exp 5 | 1 | 0.905 | 0.903 | 0.753 | 0.840 | ||
| Exp 3 | 0.905 | 1 | 0.882 | 0.680 | 1 | ||
| Exp 4 | 0.903 | 0.882 | 1 | 0.746 | 0.703 | ||
| Exp 2 | 0.753 | 0.680 | 0.746 | 1 | 0.691 | ||
| Exp 1 | 0.840 | 1 | 0.703 | 0.691 | 1 | ||
| CBA | Exp 5 | Exp 3 | Exp 4 | Exp 2 | |||
| Exp 5 | 1 | 0.857 | 0.857 | 0.857 | |||
| Exp 3 | 0.857 | 1 | 0.936 | 0.936 | |||
| Exp 4 | 0.936 | 0.924 | 1 | 0.786 | |||
| Exp 2 | 0.786 | 0.909 | 0.856 | 1 | |||
| ICS | Exp 1 | Exp 2 | Exp 4 | Exp 3 | Exp 5 | ||
| Exp 1 MRSA O3I | 1 | 0.985 | 0.985 | 0.985 | 0.985 | ||
| Exp 2 MRSA O3I | 0.985 | 1 | 0.933 | 0.933 | 0.933 | ||
| Exp 4 MRSA O3I | 0.933 | 0.949 | 1 | 0.873 | 0.873 | ||
| Exp 3 MRSA HI | 0.873 | 0.911 | 0.863 | 1 | 0.741 | ||
| Exp 5 MRSA HI | 0.741 | 0.757 | 0.725 | 0.786 | 1 | ||
| Redox status | Exp 3 | Exp 4 | Exp 5 | Exp 3 | Exp 4 | Exp 5 | |
| Exp 3 MRSA HI | 1 | 0.937 | 0.937 | 0.937 | 0.937 | 0.937 | |
| Exp 4 MRSA HI | 0.937 | 1 | 0.920 | 0.920 | 0.920 | 0.920 | |
| Exp 5 MRSA HI | 0.920 | 0.990 | 1 | 0.980 | 0.980 | 0.980 | |
| Exp 3 MRSA O3I | 0.980 | 0.994 | 0.984 | 1 | 0.955 | 0.955 | |
| Exp 4 MRSA O3I | 0.955 | 0.979 | 0.962 | 0.992 | 1 | 0.955 | |
| Exp 5 MRSA O3I | 0.955 | 0.979 | 0.962 | 0.958 | 0.958 | 1 | |
| qRT-PCR (24–72 h) | Exp 3 | Exp 5 | Exp 4 | ||||
| Exp 3 | 1 | 0.788 | 0.788 | ||||
| Exp 5 | 0.788 | 1 | 0.422 | ||||
| Exp 4 | 0.422 | –0.360 | 1 | ||||
| qRT-PCR (48–72h) | Exp 3 | Exp 4 | |||||
| Exp 3 | 1 | 0.843 | |||||
| Exp 4 | 0.843 | 1 | |||||
Table 2 presents Pearson correlation coefficients is a statistical value to assess consistency across subjects (samples) for all measured variables. The range of values goes from 0, which indicates no correlation, to +1 or –1. +1 means that the correlation is maximally positive, so the two variables grow together, while –1 indicates that the correlation is maximally negative, so as one variable grows, the other decreases. MRSA O3I and MRSA HI are always evaluated, even when not indicated. The coefficient value, close to 1, indicated a strong similarity among independent subjects, providing confidence in the reliability of the data. MRSA O3I, MRSA O3-inactivated; MRSA HI, heat-inactivated.
The relatively small sample size in this study is justified by the preliminary nature of the analysis, aimed at exploring specific in vitro effects of O3 exposure. This initial investigation provides valuable insights that can inform the design of future, larger-scale studies to achieve a more comprehensive understanding. While the in vitro model does not fully replicate the complexity of living organisms, a useful platform is provided to examine the associated cellular and molecular responses to ozone.
Our choice to use inactivated bacteria was a deliberate methodological decision guided by the primary objective of our study. The aim of the study was not to replicate an active infection, but to isolate and investigate the direct effect of ozone on the immunomodulatory and antioxidant responses of host cells to bacterial components without the additional complexities and unpredictable variables introduced by live bacterial replication and metabolism. Thus using inactivated bacteria allowed us to isolate specific response mechanisms and ensure reproducibility, which is essential for reliable comparative analysis across experimental groups.
The immunomodulatory effects of O3 were assessed in PBMCs using functional assays to measure cytokine production, ROS, and oxidative stress. In addition, we performed qRT-PCR analysis on selected pathway regulators including key genes such as NFKB1, NFKB2, and NLRP3 (involved in immune activation) at early stages of O3 treatment, and NRF2 (from gene NFE2L2) (associated with antioxidant and anti-inflammatory responses) at a later stage. This approach reflects the hypothesis-driven nature of our study, which focuses on the involvement of well-established genes in immune response, rather than on a broad, data-driven exploration. Meanwhile, qRT-PCR was selected for the associated high sensitivity and accuracy of the assay in quantifying expression changes in the genes of interest, thereby supporting our hypotheses. Although RNA-sequencing could aid broader preliminary analyses, this method was deemed unnecessary for the specific goals of this study.
Together, these functional and molecular evaluations provide robust evidence of the biological impact of O3. Nevertheless, further studies employing more complex models, larger sample sizes, and in vivo validation will be necessary to substantiate and expand upon these findings.
The bactericidal activity of O3 against MRSA was first assessed using a single-step exposure protocol. MRSA suspensions (106–107 CFU/mL) in PBS were exposed to O3 for 5 minutes, which was sufficient to achieve maximal bacterial killing (data not shown). For subsequent PBMC stimulation experiments, heat-inactivated MRSA (MRSA HI) was used as a control, providing a non-replicating bacterial stimulus while preserving antigenic properties (Table 3).
| OD | CFU/mL | |||
| PBS single-step (A) | – | – | ||
| MRSA O3I untreated | 0.70 | 7 | ||
| MRSA O3I treated with 20 µg/mL O3 for 5 min | – | 0 | ||
| MRSA O3I untreated | 0.73 | 8.7 | ||
| MRSA O3I treated with 40 µg/mL O3 for 5 min | – | 0 | ||
| MRSA O3I untreated | 0.70 | 6.4 | ||
| MRSA O3I treated with 60 µg/mL O3 for 5 min | – | 0 | ||
| HEAT-inactivated (B) | ||||
| MRSA HI untreated | 0.70 | 3.7 | ||
| MRSA HI treated for 15 min | – | 0 | ||
| RPMI multi-step (C) | – | – | ||
| MRSA O3I untreated | 0.60 | 5.4 | ||
| MRSA O3I treated with 60 µg/mL O3 | – | – | ||
| 20 min | Not done | 7.1 | ||
| 40 min | Not done | 1.1 | ||
| 60 min | – | 0 | ||
Table 3 summarizes MRSA inactivation under three experimental conditions: (A) single-step O3 treatment in PBS, (B) heat inactivation as a control, and (C) multi-step O3 exposure in RPMI.
When using RPMI medium, a multi-step protocol was implemented to provide a closer clinical replication of relevant exposure conditions. This protocol involves repeated O3 treatments at 60 µg/mL every 20 minutes, consistent with clinical protocols for managing patients with antibiotic-resistant bacterial infections, where O3 is administered twice weekly for MRSA treatment [11]. As shown in Table 3C, MRSA suspensions (106 CFU/mL) exposed to three sequential O3 applications over 1 hour exhibited a complete inhibition of bacterial growth.
To further evaluate the efficacy of O3 in the presence of antioxidant components within RPMI, additional experiments were performed using high-titer MRSA suspensions (107–109 CFU/mL) and the multi-step exposure scheme. Table 4, summarizes three experiments in which bacterial suspensions were treated every 20 minutes for 3–5 hours. A time-dependent and concentration-dependent logarithmic decline in CFU counts was observed in all cases. After approximately 3 hours of repeated O3 exposure, a five-log reduction in viable bacteria was achieved, confirming the potent bactericidal effect of O3 even under conditions that partially mimic physiological environments.
| Conditions | CFU/mL | ||
| Experiment 1: In 180 minutes from 109 CFU/mL | |||
| MRSA untreated | 4.3 | ||
| MRSA treated with 60 µg/mL O3 | |||
| After 20 min | 1.7 | ||
| After 180 min | 7.6 | ||
| Experiment 2: In 300 minutes from 109 CFU/mL | |||
| MRSA untreated | 6.8 | ||
| MRSA treated with 60 µg/mL O3 | |||
| After 100 min | 1.2 | ||
| After 300 min | 1.4 | ||
| Experiment 3: In 180 minutes from 107 CFU/mL | |||
| MRSA untreated | 1.9 | ||
| MRSA treated with 60 µg/mL O3 | |||
| After 60 min | 1.3 | ||
| After 180 min | 1.6 | ||
Table 4 presents the results of high-titer MRSA (107 to 109 CFU/mL) treated with O3 using a multi-step protocol in RPMI. A time-dependent logarithmic decrease in CFUs was observed in all experiments.
However, at very high bacterial densities, S. aureus tends to form aggregates, which can transiently reduce O3 efficacy due to the protective effect of outer-layer killed cells, meanwhile those within the clusters remain protected, a phenomenon also reported for Pseudomonas aeruginosa exposed to conventional antibiotics [50].
To ensure complete sterility before use in stimulation assays with PBMCs, residual bacterial colonies were eliminated by gamma irradiation at 2700 Gy for 3 days. No bacterial growth was observed in any of the preparations after 24 hours. This approach is well documented for decontaminating S. aureus in food matrices and has the advantage of preserving protein epitopes, thereby maintaining antigenic integrity [51, 52].
Both MRSA-O3I and MRSA HI preparations were subsequently used to stimulate PBMCs isolated from 3/5 healthy donors.
Cruciani et al. [48] showed that S. aureus induced a specific
panel of regulatory cytokines (IL-23, IL-12, and IL-10) and pro-inflammatory
mediators (TNF-
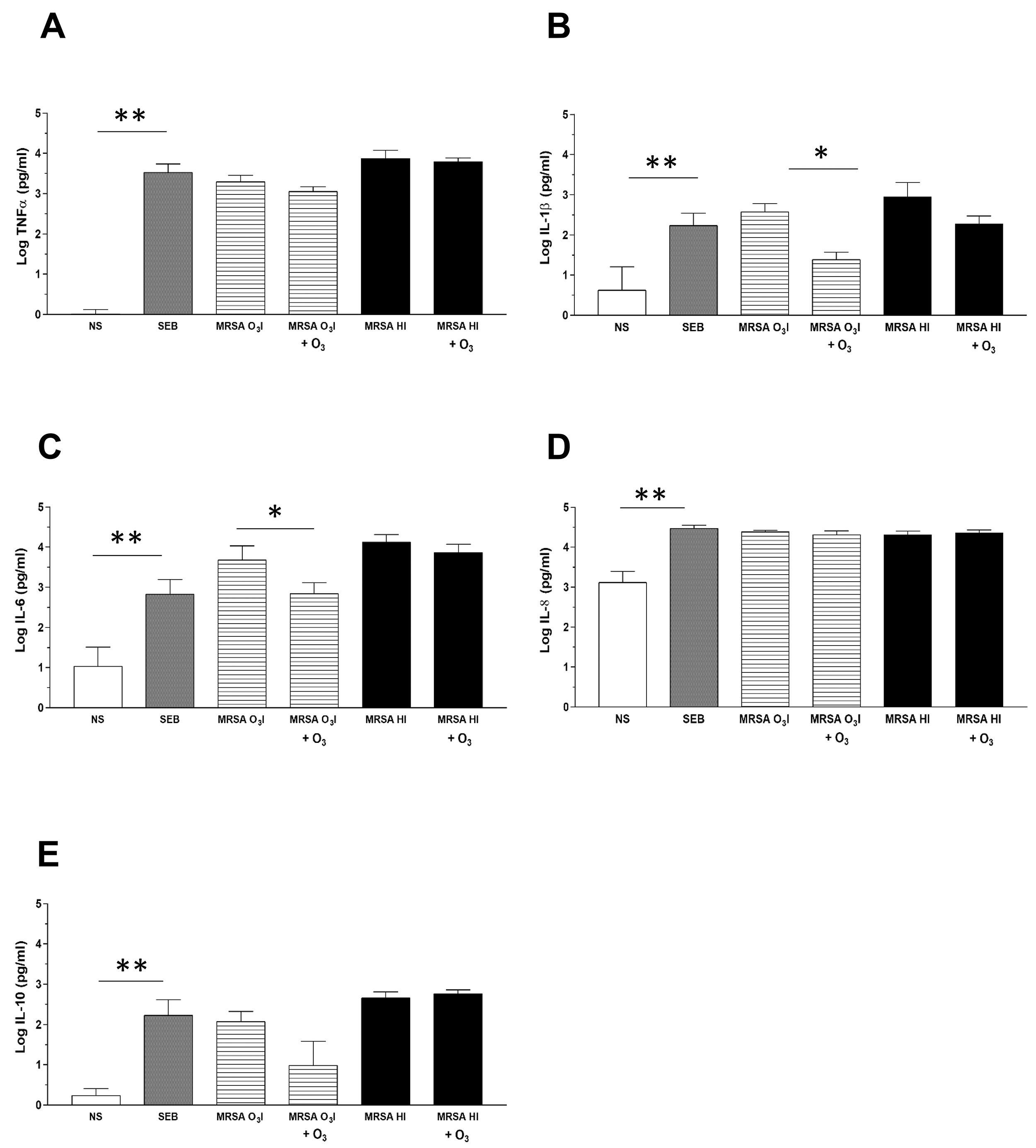
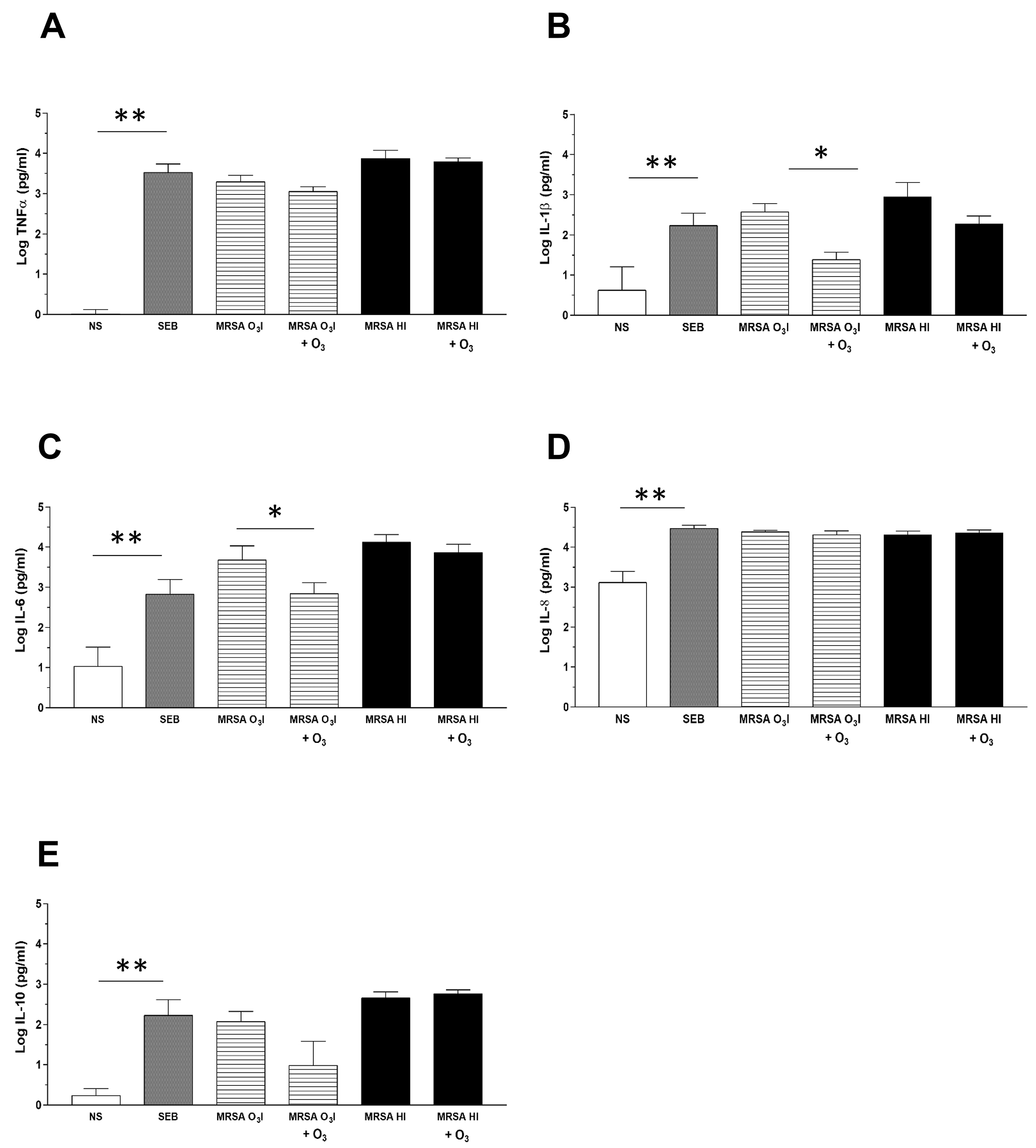 Fig. 1.
Fig. 1.
Cytokine detection in culture supernatants by CBA. The
bar graphs depict cytokine levels in culture supernatants from PBMCs
collected 24 hours after treatment, with or without O3 added after 2 hours
of stimulation with MRSA O3I or MRSA HI. Cytokines are expressed in pg/mL on
a logarithmic scale. Cytokine production of TNF
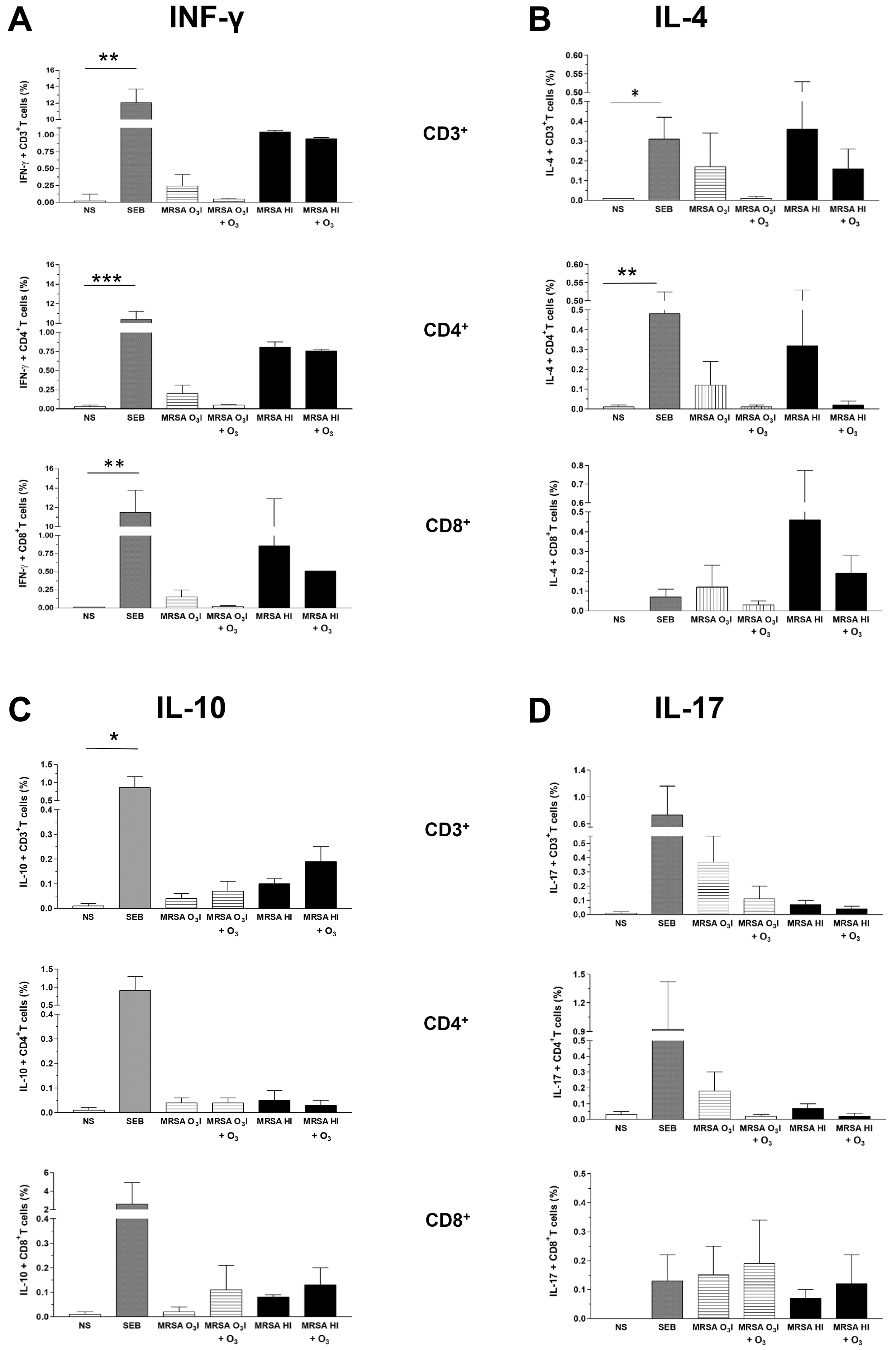
The intracellular production of cytokines associated with the activation of T
lymphocytes was assessed by intracellular cytokine staining in PBMCs stimulated
with MRSA O3I and MRSA HI. The gating strategy is shown in
Supplementary Fig. 1. A reduction in IFN-
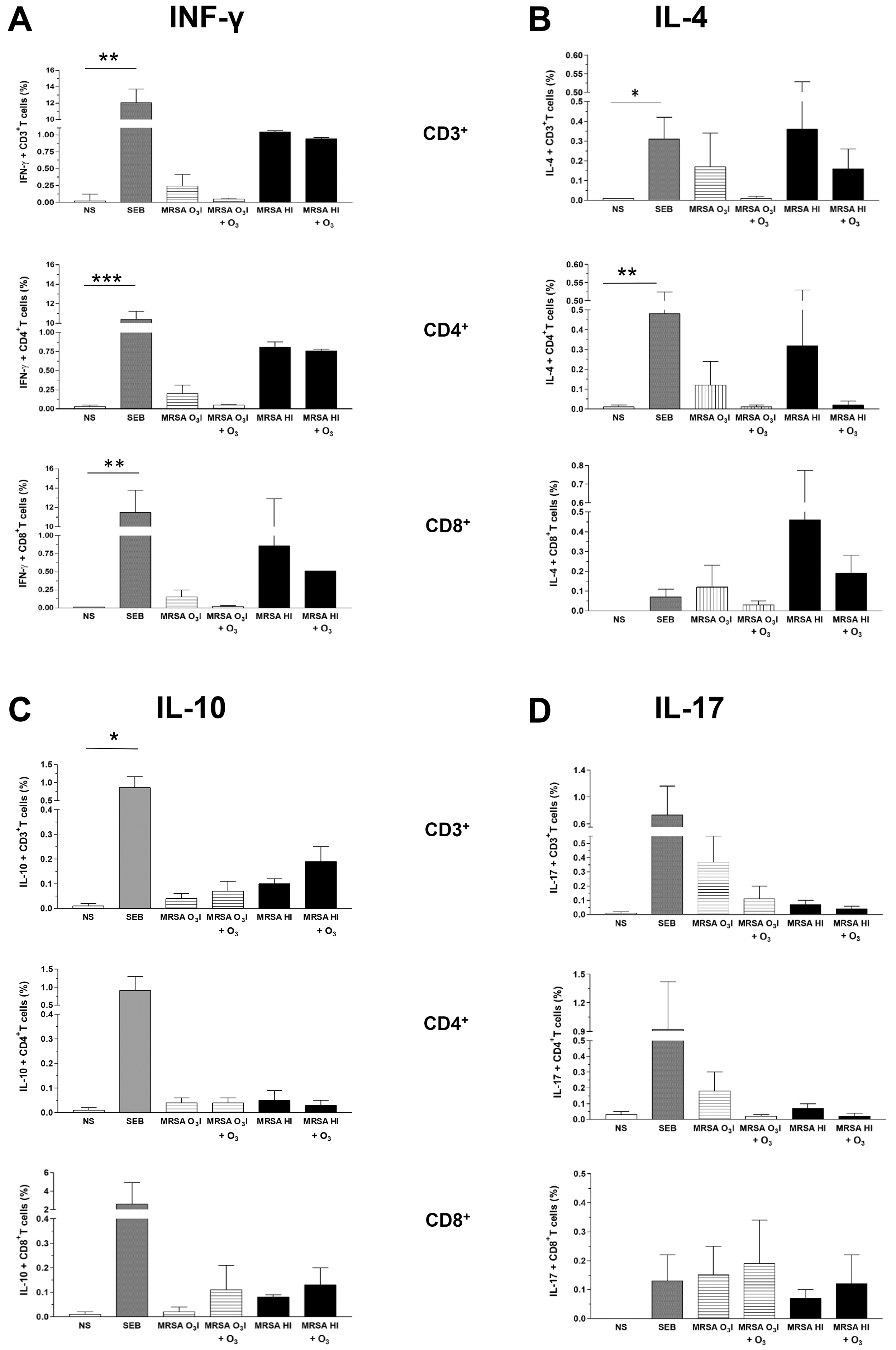 Fig. 2.
Fig. 2.
Intracellular cytokines in CD3, CD4, and CD8 lymphocyte gates.
Bar graphs of the percentage of intracellular cytokines IFN-
The O3-induced reduction in IL-4 levels was comparable across all T cell subsets, regardless of the bacterial inactivation method (Fig. 2B).
Conversely, IL-10 was increased in CD3+ and CD8+ T cells following O3 treatment of cells stimulated with either MRSA O3I or MRSA HI. In contrast, in lymphocytes in the CD4+ gated population, IL-10 remained unchanged in MRSA O3I-treated samples, while the IL-10 levels were slightly decreased in the MRSA HI-stimulated samples (Fig. 2C).
An O3-induced decrease in IL-17 levels was detected exclusively within the CD3+ and CD4+ T cell populations, whereas a slight increase of this cytokine was observed in CD8+ T cells. This trend was consistent across both MRSA strains (Fig. 2D). The increase in IL-17 in CD8+ T cells may be partially correlated with the rise in IL-10 within the same gate in cells exposed to both bacterial strains. Although not statistically significant, these data suggest a trend toward differential expression of intracellular cytokines, except for SEB, which induces a significant increase in all tested cytokines, except IL-17, compared with untreated control samples.
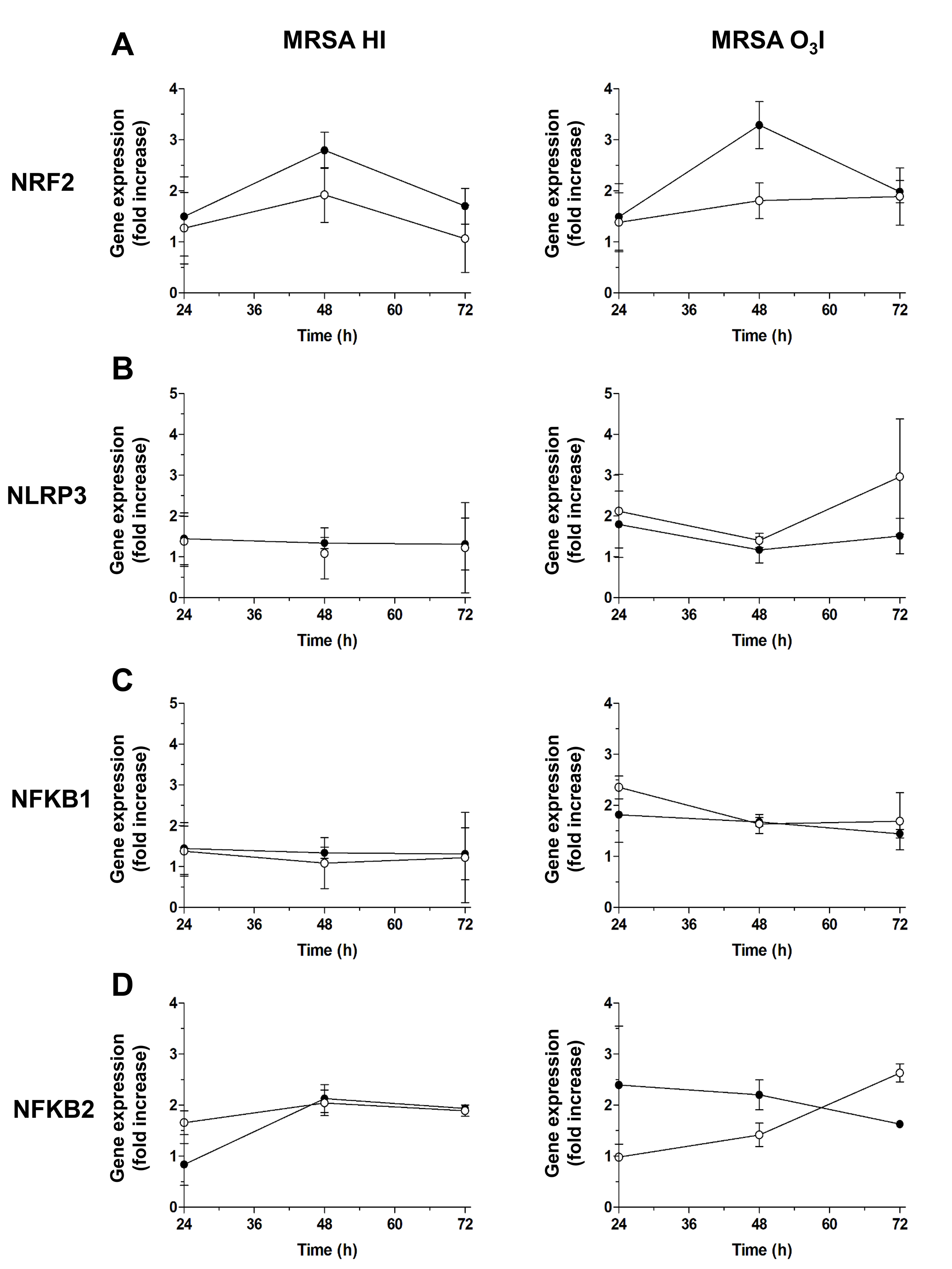
To assess the effects of O3 on key regulators of inflammation, we used
qRT-PCR to measure the effect of O3 on prooxidant- (NLRP3,
NF-
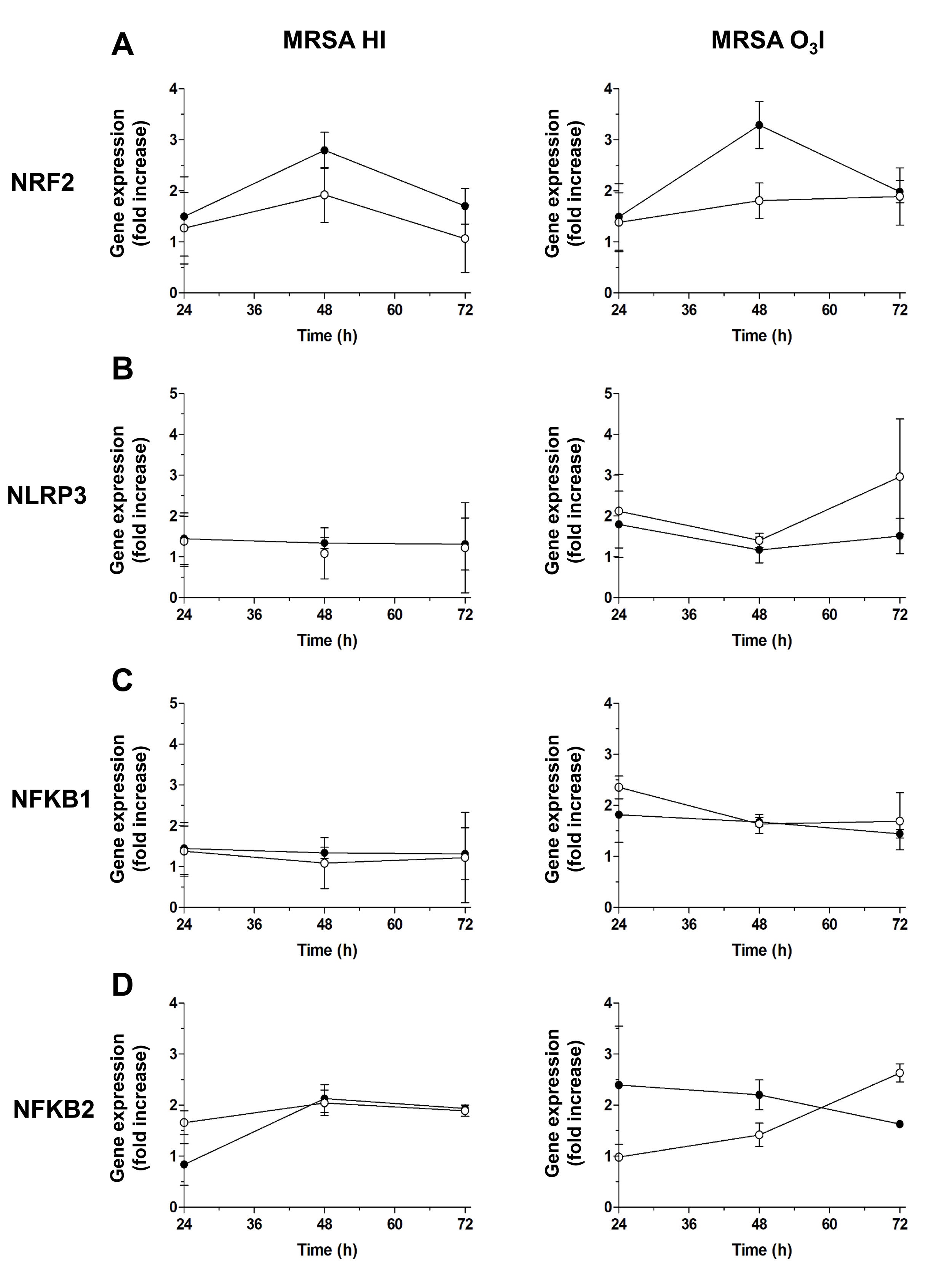 Fig. 3.
Fig. 3.
Gene expression of key O3 regulators. Graphs of the gene
expression time course for key O3 regulators assessed by RT-PCR from 24 to
72 hours: (A) antioxidant NRF2, (B) pro-oxidant NLRP3, (C)
NF-
NRF2 expression, a key regulator of the antioxidant response, increased following O3 treatment, with the most pronounced upregulation observed after 48 hours in PBMCs exposed to O3-killed bacteria in PBS and treated with O3. A more modest increase was noted in cells treated with heat-killed MRSA HI and subsequently exposed to O3 (Fig. 3A), indicating that O3 enhances antioxidant defenses, particularly in the context of O3I bacterial stimulation.
In contrast, NLRP3, a proinflammatory gene, was strongly induced by MRSA O3I at 24 and 72 hours. O3 treatment attenuated this induction, reducing NLRP3 expression is reduced to approximately 1.8-fold after 24 hours and decreases further at 48 hours, and by approximately 50% at 72 hours, while a similar reduction was observed in unstimulated cells at 48 hours. PBMCs stimulated with MRSA HI showed only minimal changes in NLRP3 expression (Fig. 3B).
Similarly, NFKB1 expression induced by MRSA O3I was affected by O3 addition at 24 hours and 72 hours while no changes were observed at 48 hours with NFKB1 below the threshold (Fig. 3C) Likewise, but to a lesser extent NFKB1 induced by MRSA HI at 24 hours was downregulated by O3, while no substantial variations in the expression of NFKB1 were observed in MRSA HI stimulated PBMCs at 48 and 72 hours before and following O3 treatment. For NFKB2, an increase was observed at 24 and 48 hours only in MRSA O3I stimulated cultures after O3 treatment, whereas expression decreased at 72 hours. A slight reduction was also measured in MRSA HI-treated PBMCs following O3 stimulation (Fig. 3C).
NFKB1 expression followed a similar pattern: NFKB1 was induced by MRSA O3I at 24 and 72 hours and downregulated by O3 treatment, whereas at 48 hours expression remained below the threshold. In PBMCs stimulated with MRSA HI, NFKB1 expression showed only a slight reduction at 24 hours following O3 exposure, with negligible changes at 48 and 72 hours (Fig. 3C).
Correlation analyses revealed minimal changes in NFKB1 and NFKB2 expression at 24 hours, while at 48 and 72 hours, strong correlations were observed among NRF2, NLRP3, NFKB1, and NFKB2 (Table 2). These findings indicate that O3 maintains the associated antioxidant and modulatory effects over time, supporting sustained regulation of both pro- and anti-inflammatory pathways for up to 72 hours.
As expected, NRF2 was not induced by SEB at any time point (24–72 hours), consistent with the role of NRF2 in anti-inflammatory signaling. Similarly, NLRP3, a key component of the inflammasome, was not upregulated at 48–72 hours, suggesting that additional inflammatory stimuli are likely required to induce NLRP3. In contrast, the proinflammatory transcription factors NFKB1 and NFKB2 were both induced by SEB at 24 and 48 hours. However, by 72 hours, only NFKB2 remained upregulated, while NFKB1 returned to baseline levels (Supplementary Fig. 3).
Data are presented as the means
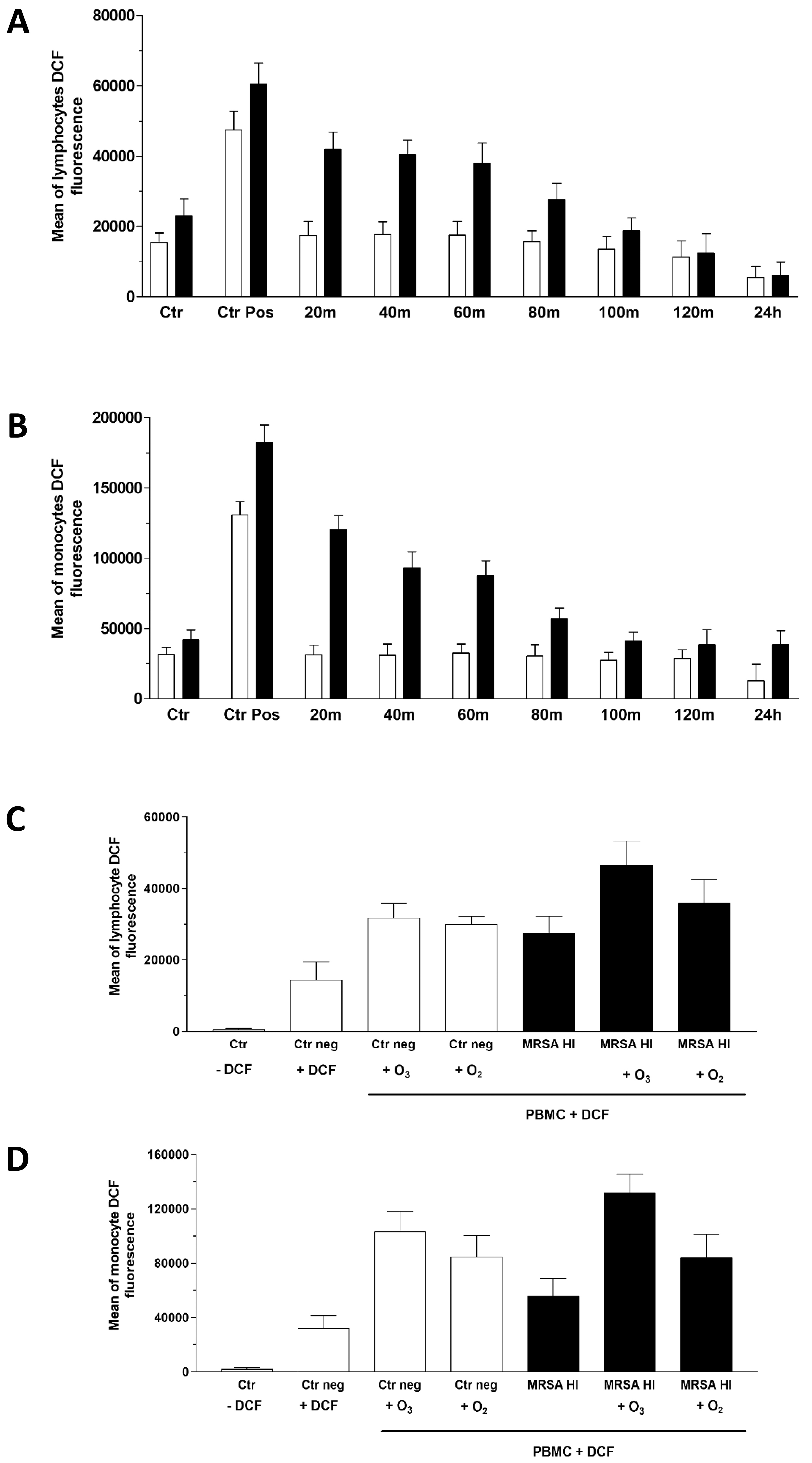
ROS comprise a wide range of chemical species derived from O2 that are more reactive than solely O2, exhibiting distinct properties, reactivities and interactions.
When ROS are generated in vivo, numerous antioxidants are activated, delaying, preventing or removing oxidative damage to target molecules [53].
Thus, we evaluated DCF detection in lymphocytes and monocytes separately across the total PBMC population and observed similar kinetics in both cell types within 2 hours and after 24 hours.
As shown in Fig. 4, the presence of the inactivated MRSA in the culture slightly increased the amount of intracellular ROS released by lymphocytes (Fig. 4A) and monocytes (Fig. 4B). Nevertheless, the O3 treatment initially increased ROS levels, showing an initial oxidant effect of the highly reactive gas. The highest ROS levels were observed within the first 20–40 minutes after ozonation and gradually decreased reaching an antioxidant status comparable to that of the corresponding untreated samples within 2 hours, and even falling below control levels after 24 hours in lymphocytes and monocytes stimulated with MRSA HI, where significant p values were found (Supplementary Table 2). Meanwhile, a similar trend to MRSA HI was observed for those treated with MRSA O3I and exposed to O3 (data not shown). This confirms that initial exposure to low doses of O3 produced only a partial, reversible oxidizing effect, resulting in a more stable antioxidant outcome. Because O3 is a highly reactive gas composed of three oxygen atoms, we compared the effects of the two gases using the same protocol described previously, applying pure oxygen (O2) and O3 treatments. The experiment was performed using MRSA HI and DCF fluorescence intensity was measured after 20 minutes. As expected, since Staphylococcus aureus is an aerobic bacterium, O2 MRSA HI had an oxidizing effect. As shown in Fig. 4, enhanced O3 derived ROS production in both lymphoid (Fig. 4C) and monocytic (Fig. 4D) subpopulations is higher in the presence of O3. Furthermore, it must be considered that O2 does not inactivate bacteria unlike O3.
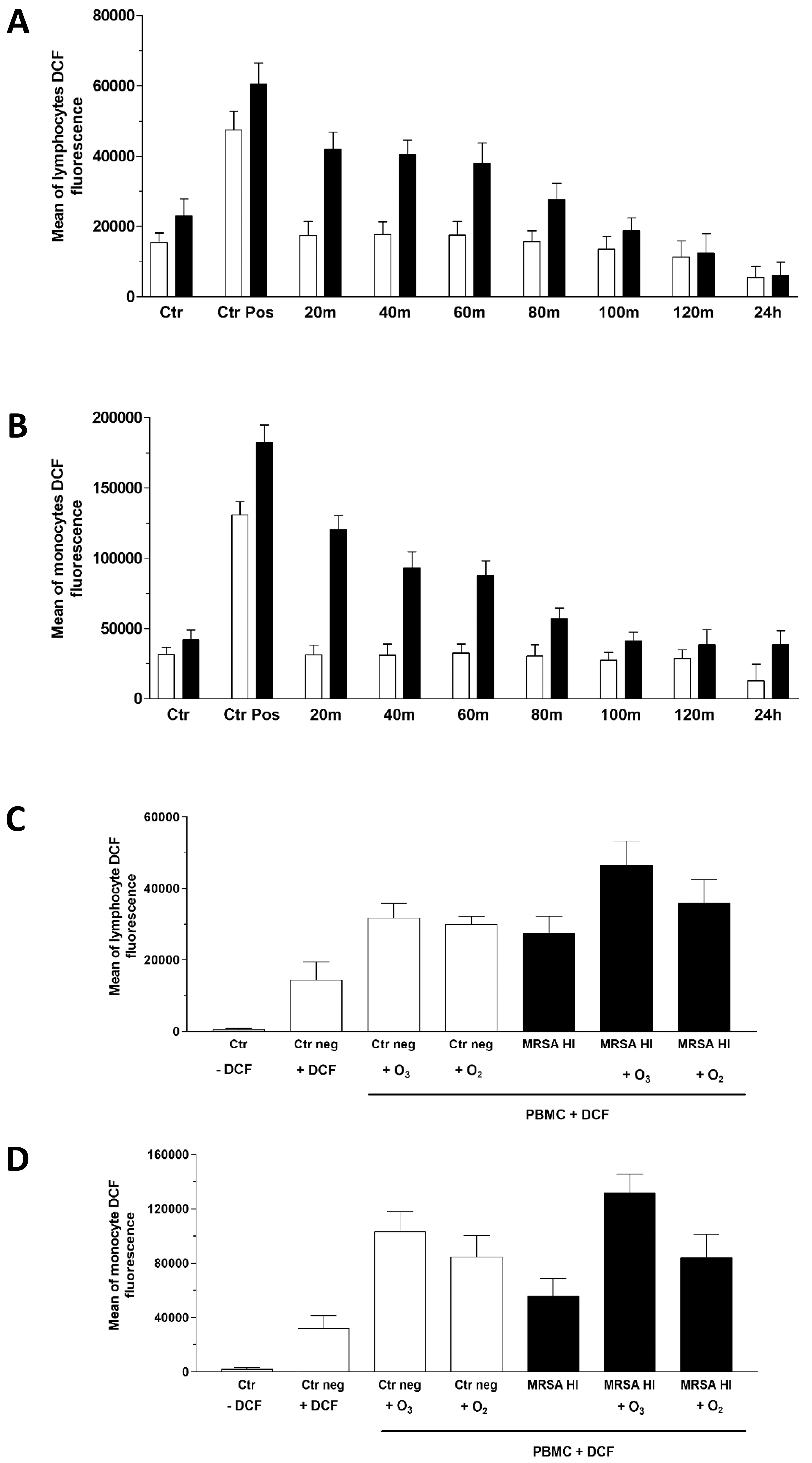 Fig. 4.
Fig. 4.
Measurement of reactive oxygen species (ROS) via 2′,7′-Dichlorofluorescein (DCF) assay. Flow cytometry histograms of the ROS levels in PBMCs from MRSA-infected and non-infected samples treated with O3. ROS production is measured at multiple time points (0–2 hours and 24 hours post-treatment) using the DCF assay. H2O2 was used as the positive control. Data are shown as the fluorescence intensity profiles for lymphocyte (A) and monocyte (B) populations. In panels (C) and (D) the comparison of ROS levels in MRSA-infected and non-infected PBMCs treated with O3 or oxygen (O2) was reported. ROS production has been assessed over time with the DCF assay. Statistical analysis was performed to compare the effects of O3 and O2 on ROS generation in lymphocytes (C) and monocytes (D), respectively. The experiments were conducted with five healthy subjects (see Raw data), but the figure presents descriptive statistics for a representative sample. Statistical analysis was performed to compare the effects of O3 on ROS generation. The p-values for three experiments were significant in MRSA HI in both lymphocytes and monocytes. Data are described in (Supplementary Table 2). The p-value for three independent experiments, conducted by three different donors, was 0.075 (two-tailed) for paired samples comparing MRSA O3I and DCF at 20 minutes, compared with MRSA O3I and DCF at 24 hours (data not shown).
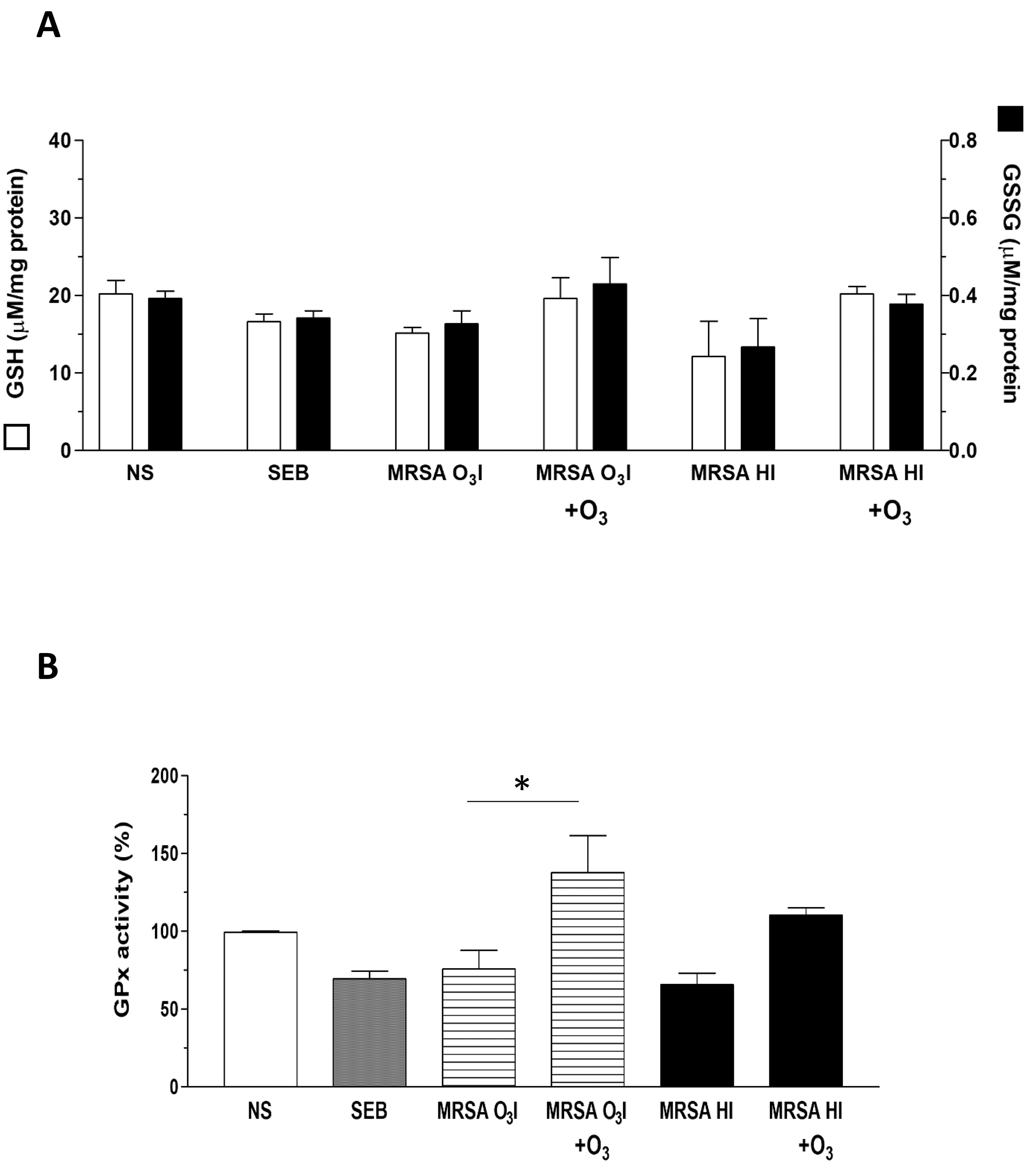
GSH plays a key role in maintaining the cellular redox balance by directly neutralizing ROS and serving as a cofactor for various enzymes. During this process, GSH is oxidized to oxidized glutathione (GSSG), which is then reduced back to GSH by specific enzymes. GSH is also essential for GPx activity, which protects cells from oxidative damage by reducing peroxides using GSH as an electron donor [54].
The increase in ROS formation in MRSA-treated PBMCs and the protective effects of O3 prompted us to investigate the status of intracellular antioxidant defenses. It is well known that increased oxidative stress alters intracellular redox status due to the depletion of antioxidant defenses. Thus, we investigated the ability of O3 to modulate the intracellular concentration of glutathione and the activity of the antioxidant enzyme GPx in unstimulated PBMCs or those activated with SEB as a positive control, MRSA O3I, and MRSA HI (Fig. 5A). The GSH concentration was significantly decreased by SEB and by both MRSA O3I and MRSA HI (Fig. 5A), indicating that the thiol was consumed in counteracting oxidant species formed in activated PBMCs. Interestingly, the O3 treatment restored GSH levels to levels comparable to controls when PBMCs were activated with MRSA O3I and MRSA HI, whereas GSH levels were even higher than controls when activated with SEB.
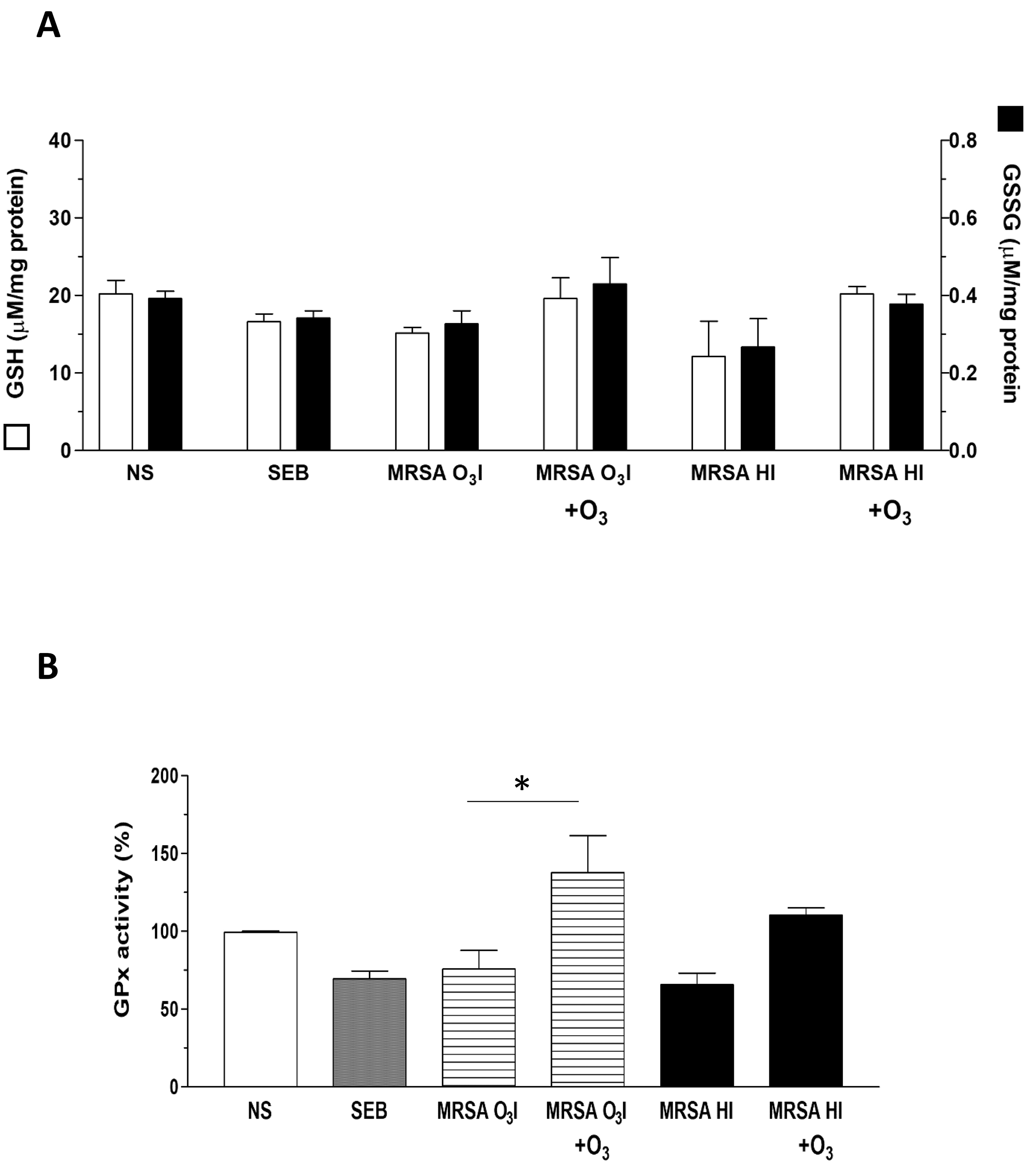 Fig. 5.
Fig. 5.
Restoration of redox status in MRSA-treated PBMCs by ozone. (A)
Graphs of the GSH and GSSG concentrations in PBMCs treated with SEB, MRSA
O3I, and MRSA HI. O3 treatment is shown to restore redox balance to
control-like levels. (B) GPx activity levels in PBMCs under various treatments,
with data on the restorative effect of O3 treatment. The results are
presented as the mean
The GSSG intracellular concentration was unaffected by the exposure of PBMCs to SEB or to MRSA O3I; meanwhile, the GSSG intracellular concentration was significantly decreased by the MRSA HI treatment (Fig. 5A). The 40 µg/mL O3 treatment did not significantly modify the GSSG concentration in any experimental condition tested with respect to the unstimulated cells or the corresponding treatment group.
The activity of GPx was significantly reduced in PBMCs activated with SEB or with MRSA O3I and MRSA HI by 31, 25, and 35%, respectively, with respect to the unstimulated cells (Fig. 5B). In MRSA O3I + O3 versus MRSA O3I, the difference was significant with a p-value of 0.026 for the two-tailed paired sample analysis. Interestingly, the 40 µg/mL O3 treatment, not only restored but rather increased the enzyme activity in PBMCs activated with SEB or with MRSA O3I and MRSA HI by 24, 37 and 10%, respectively, compared to the unstimulated cells (Fig. 5B). Overall, these results show that O3, can protect cells from MRSA-dependent oxidative stress, decreasing the levels of ROS formed and restoring the intracellular redox equilibrium.
This in vitro study highlights the potential of O3 therapy as a complementary strategy for managing Staphylococcus aureus infections, particularly MRSA, which poses a serious challenge due to the associated resistance to conventional antibiotics. Notably, O3 demonstrated a dual effect: direct bactericidal activity against MRSA and significant immunomodulatory and antioxidant effects on human PBMCs.
MRSA, when inactivated by a single-step O3 treatment in PBS, induced a pro-inflammatory response in PBMCs, as evidenced by cytokine production within 24 hours. However, subsequent addition of O3 to the cultures led to a marked reduction in both intracellular and secreted pro-inflammatory cytokines, as shown by the ICS and CBA assays, respectively, indicating an anti-inflammatory effect. This trend was also observed with heat-inactivated bacteria, suggesting that the immunomodulatory response is influenced more by bacterial viability than by the inactivation method [48]. An intriguing discrepancy emerged in IL-10 levels between ICS and CBA measurements. Indeed, while IL-10 levels consistently decreased in the CBA assay, these levels increased in the ICS assay. This pattern was evident under both O3-inactivated and heat-inactivated bacterial conditions. This effect was particularly evident in the CD3+ and CD8+ T cell subsets, whereas IL-10 levels remained stable in CD4+ lymphocytes. Given that IL-10 is an O3-induced anti-inflammatory mediator, this finding warrants further investigation [55]. One possible explanation is that the early time point (24 hours), may be sufficient to detect intracellular IL-10 synthesis but not secretion. Additionally, differential responses between CD4+ and CD8+ cells may indicate an effect of O3 on other immune subsets such as CD3– NK cells (CD56+), double-negative T cells (CD3+CD4–CD8–), or Tregs, which warrants further investigation. These cells, which can be either CD4+ or CD8+, play a role in maintaining homeostasis, preventing autoimmunity [56] and reducing exacerbation of CD8+ lymphocyte responses to pathogens, thereby helping to restore the integrity of damaged tissues [57].
A general decrease in IFN-
The slight increase in IL-8 secretion, despite the general reduction in
TNF-
At the molecular level, qRT-PCR data demonstrated activation of the NRF2-KEAP1
antioxidant pathway by 48 hours, accompanied by downregulation of NLRP3 and
NFKB1—key mediators of the inflammatory response, thereby confirming that
O3 may exert antioxidant and immunomodulatory effects. Interestingly, NFKB2
exhibited a divergent pattern in response to MRSA HI stimulation, being
upregulated 48 hours after O3 treatment and subsequently downregulated at 72
hours. This pattern suggests differential regulation of NF-
With respect to oxidative stress, our data confirmed that MRSA induces significant ROS generation, which is rapidly reduced by O3 treatment within 24 hours, as previously described both in vitro [62, 63] and in vivo [64, 65, 66] models. This was accompanied by a restoration of the intracellular redox balance, likely mediated by activation of the AMPK/Keap1/Nrf2 axis. This pathway promotes transcription of antioxidant response element (ARE)-regulated genes such as GCLC and GPx, thereby enhancing GSH synthesis and antioxidant enzyme activity [67]. Overall, this study demonstrates, using biochemical, immunological, and molecular approaches, that O3 exerts rapid and sustained anti-inflammatory and antioxidant effects, with maximal impact observed between 48–72 hours post-treatment. These findings underscore the potential of O3 as a supportive therapy for multidrug-resistant infections such as MRSA.
This pilot study suggests that O3 therapy exerts antibacterial, antioxidant, and immunomodulatory effects on MRSA-stimulated PBMCs. These findings support the potential use of O3 therapy as an adjunct strategy to combat antimicrobial resistance and warrant further validation in more advanced models.
Although the limited number of PBMC donors presents a challenge, the consistent and reproducible patterns observed across all experimental methods, except for qRT-PCR at 24 hours, indicate a robust and non-random response. Future research should include a larger donor pool and examine additional cell subsets, such as Tregs, NK cells, and double-negative T cells, to further elucidate the cellular targets potentially affected by O3. Additionally, extending time-course analyses beyond 24 hours will be crucial for clarifying the kinetics of cytokine secretion, particularly IL-10, IL-17A, and IL-8.
The analyzed data sets generated during the study are available from the corresponding author upon request.
AC: Conceptualization and Project design, supervision, funding acquisition, data curation and writing—original draft. CC: Development of techniques for cell ozonation, investigation, data analysis, writing—review and editing. VLS: Investigation, validation, writing—review and editing. IM: Investigation, validation, data analysis, writing—original draft, and writing—review and editing. SG: Development of techniques for bacterial cells ozonation, investigation and techniques validation; data analysis, writing and editing. MP: Development of techniques for bacterial cells ozonation, investigation and techniques validation; data analysis, writing and editing. DP: Investigation, data curation, writing—review and editing. MS: Supervision (cellular group), methodology, validation, writing—review and editing, data curation. MZ: Investigation, methodology, and validation. GC: Investigation, methodology, and data curation. MFab: Investigation, methodology, and data curation. MM: Conceptualization, supervision, methodology, validation, data curation, writing, and editing. MDG: Conceptualization, supervision, methodology, validation, data curation, writing, and editing. RC: Conceptualization, supervision, methodology, validation, data curation, writing, and editing. GE: Conceptualization, supervision, methodology, validation, data curation, writing, and editing. SP: Conceptualization support, data interpretation, and visualization. LV and MFra: Literature search, provision of the MultiOssigen 99 IR device, funding acquisition, and project administration. All authors contributed to editorial changes in the manuscript. All authors read and approved the final version of manuscript and have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
This study was approved by the Ethics Committee of the Italian National Institute of Health with protocol PRE BIO CE n.0041072 approved on 24/11/2021, and has been conducted in accordance with the Declaration of Helsinki. Human PBMCs were isolated from freshly collected buffy coats derived from healthy male and female volunteers (Blood Bank of University “La Sapienza”, Rome, Italy) that gave written informed consent.
We thank all our colleagues of the Italian National Institute of Health, Rome, Italy. In particular, Eliana Coccia and Marilena Etna for their valuable assistance and helpful discussion; Alessandro Giuliani for his important support in statistical analysis; Simone Martinelli and Giuseppe Esposito for their advice and assistance in use of the gamma-ray technique; Fabio D’Ambrosio for his aid in bacterial collection. We thank Massimo Delle Femmine for improving the figures quality.
This work was supported by SIOOT project funding number BB08.
The authors declare no conflicts of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBE44093.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.