

 , Olga A. Karavaeva 1
, Olga A. Karavaeva 11 Institute of Biochemistry and Physiology of Plants and Microorganisms – Subdivision of the Federal State Budgetary Research Institution Saratov Federal Scientific Centre of the Russian Academy of Sciences (IBPPM RAS), 410049 Saratov, Russia
Abstract
Since the discovery of penicillin in 1928, more than a thousand antibiotics have been introduced into medical practice. Initially, the introduction of antibiotics revolutionized the treatment of infectious diseases. However, as antibiotics have been used repeatedly, scientists and clinicians have observed serious adverse consequences, most notably the emergence of bacterial resistance to antimicrobial agents. Of particular concern are multidrug-resistant pathogens, which are resistant to multiple antibiotic classes. As of 2024, antibiotic-resistant bacteria are directly responsible for approximately 1.27 million deaths worldwide. Thus, in response, possible new antibacterial agents are under investigation, including bacteriophages and the associated bacteriolytic enzymes. According to the World Health Organization (WHO), phage-based drugs are leading nonclassical antibiotics currently in clinical trials. Meanwhile, endolysins, as bacteriolytic phage enzymes, have emerged as promising antibacterial agents. This study reviews the key achievements and mechanisms of action of bacteriophage endolysins against bacterial infections, including biofilms, and discusses the therapeutic potential of endolysins in combating antibiotic resistance. Finally, the combined use of endolysins with other biomolecules and the application of artificial intelligence to address antimicrobial-resistant infections are also discussed.
Keywords
- endolysins
- bacteriophages
- multidrug resistance
- infection
- therapy
Bacterial infections are a major public health problem worldwide. According to statistics for 2019, 13.7 million deaths from bacterial infections were recorded. A large percentage of mortality cases associated with bacterial infections is due to the increased bacterial antibiotic resistance [1, 2]. According to the 2024 data, the number of deaths directly caused by antibiotic-resistant bacteria was about 1.27 million [3]. Therefore, new antimicrobial drugs are being developed that are active against clinically important pathogens. One strategy to combat bacterial antibiotic resistance is to use phage therapy and bacteriolytic phage enzymes. Therapeutic strategies using bacteriophages are promising approaches to treating antibiotic-resistant bacterial infections. An effective method of using bacteriophages is the individual selection of a virus for a specific pathogen (a rather expensive and labor-intensive procedure), as well as immunogenicity [4]. In clinical practice, phage therapy is used for a narrow range of diseases (chronic otitis and rhinosinusitis, bacterial infections in cystic fibrosis, infections after burn injury) and pathogens (mainly Pseudomonas aeruginosa and Staphylococcus aureus). There have been no sufficient studies of the pharmacokinetics and pharmacodynamics of bacteriophage preparations or of their interaction with the human immune system, and no accurate data have been reported on the possible side effects of phage therapy for the human body. Usually, no information is provided on the genotype of the phage or bacterial host, levels of endotoxin contamination of the bacteriophage preparation, and the susceptibility testing results [5]. Therefore, research is actively underway on the use of phage enzymes as antimicrobial drugs. Endolysins are bacteriophage-encoded peptide-degrading enzymes necessary for the lysis of the cell wall of the host bacteria. According to WHO, in 2017, two protein drugs were developed by global pharmaceutical companies that were based on bacteriophage endolysins [6, 7, 8]. Phage enzyme preparations have not yet found widespread use in the fight against bacterial pathogens, mainly owing to their high cost. This review shows the possibilities and prospects of using endolysins in the fight against prokaryotes not only in planktonic form but also as part of biofilms, and it also discusses the limitations of endolysin use.
Owing to the lack of effective antibiotic therapy, antibiotic resistance has become a threat to humanity [9, 10, 11]. Of particular concern is the emergence of bacterial strains resistant to several antimicrobial drugs at once. In 2024, WHO updated the ranked list of bacterial pathogens on the basis of their resistance to antibiotics actively used in clinical practice and the degree of their danger [12]. More than half of all deaths from bacterial infections worldwide are caused by bacterial pathogens, including S. aureus, Escherichia coli, Streptococcus pneumoniae, Klebsiella pneumonia, Acinetobacter baumannii, P. aeruginosa, and Enterobacter spp. (ESCAPE group).
According to WHO, the incidence of strains with multiple drug resistance (MDR), including the most important ones (K. pneumoniae, P. aeruginosa, E. coli), reaches 70–80% in some regions of the world [13]. Owing to the COVID-19 pandemic and related occurrences of secondary bacterial pneumonia, the incidence of mortality from antibiotic-resistant bacteria increased strongly in 2020. Klebsiella, E. coli, Pseudomonas, and Citrobacter were the most common Gram-negative bacteria linked to coinfections in England in 2020. Staphylococci and enterococci were the most common Gram-positive bacteria. Among bacteria linked to COVID-19 pneumonia, enterococci rank among the most prominent [14, 15, 16, 17, 18].
The development of bacterial biofilms, which are difficult for antibiotics to penetrate, is frequently linked to a lack of antibiotic efficiency. Compared with their planktonic counterparts, biofilm bacteria are 10–1000 times more resistant to antibiotics [19]. Antibiotics, antibodies, and granulocyte cell populations are all slowed down by the physical barrier that biofilms provide. Antimicrobial resistance unique to biofilms can be attributed in part to low doses of antibiotics used in the long term and to varying treatment regimens, which can promote the production of biofilms [20]. The American National Institute of Health and the International Center for Disease Control and Prevention reported that at least 80% of biofilm-forming pathogens are linked to persistent infections and that bacterial biofilms are implicated in 65% of microbial diseases [21] and more than 80% of chronic infections [22, 23].
The National Institutes of Health (NIH) has suggested that approximately 80% of wound infections reported in the United States may be associated with biofilm formation [21, 24]. With account taken of these data, the priority area for counteracting and combating antibiotic resistance in bacteria is the development of innovative antibacterial agents [25], one of which is the use of bacteriophage enzymes.
The current theory views antibiotic resistance as a crucial component of the microbial population’s evolutionarily developed capacity for adaptation to outside stimuli. Antibiotic resistance results from the evolution of bacterial human pathogens owing to anthropogenic changes in the external environment [26, 27, 28, 29, 30, 31]. Primary causes of MDR include antibiotic inactivation and/or structural changes, drug binding site modifications, alterations in cell permeability, efflux pump biosynthesis, and biofilm formation [32, 33, 34, 35].
Antibiotic resistance may spread in numerous ways [35, 36]. For instance,
antibiotics can be carried outside of the bacterial cell by auxiliary pumps
(e.g., TetA proteins, which pump tetracyclines) or inactivated enzymes
(
It is obvious that bacteria solve the evolutionary problem of combating the effects of antibiotics by acquiring preexisting resistance determinants. The main antimicrobial groups, the genes responsible for antibiotic resistance, and the types of resistance mechanisms were described by Reynoso et al. [37].
Improved knowledge of the mechanisms and drivers at the individual and population levels, antimicrobial resistance surveillance, antimicrobial stewardship, enhanced infection prevention and control measures, enhanced global policy and funding, and the development of new antimicrobial strategies are all components of a comprehensive and multifaceted strategy to combat antimicrobial resistance [3].
The formation of bacterial biofilms by microbial cells contributes to the development of bacterial resistance to antimicrobial drugs; i.e., biofilm bacteria, in contrast to planktonic ones, are less sensitive to antimicrobial agents. The biofilm acts as a physical barrier and allows bacteria to resist chemotherapy and antibiotics [34]. Because biofilm-associated infections and problems with their treatment are considered a serious threat to human health, in recent years, more and more research has been carried out to investigate the formation of biofilms and develop strategies to combat them. Promising new antibacterial agents are endolysins as bacteriolytic phage enzymes, which will be discussed further.
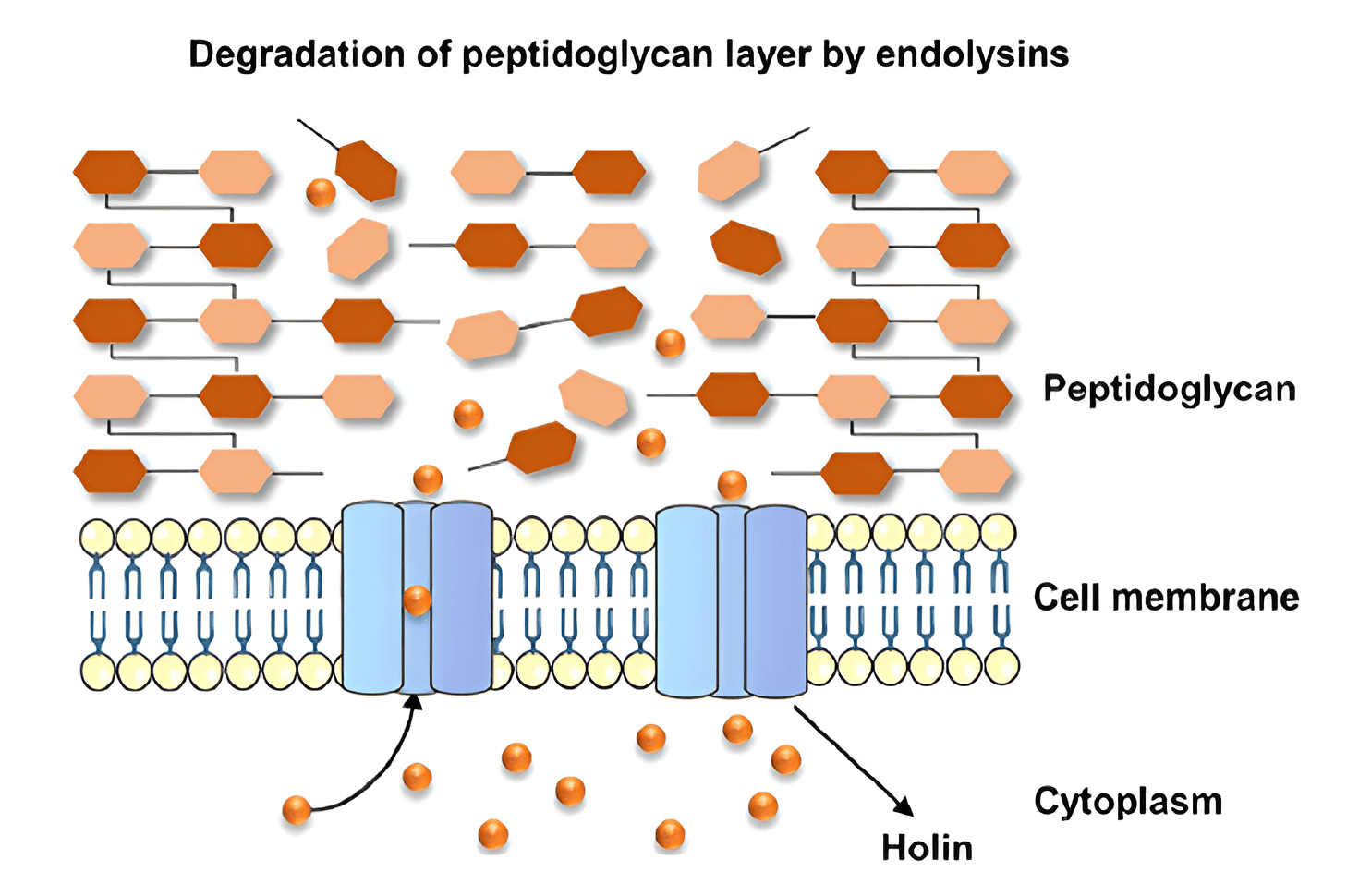
Endolysins are bacteriophage-encoded peptide-degrading enzymes required for the lysis of the host bacterial cell wall. The name was first used to describe proteins generated in phage-infected bacteria [38]. Because endolysins encourage host cell rupture, they are in charge of releasing freshly generated viral particles [39]. Before introducing its DNA into a bacterial cell, a bacteriophage must be adsorbed on its surface. This process is often complicated by structures on the surface of bacteria (mucous sheaths, capsules, exopolysaccharides, etc.). To overcome them, some bacteriophages use special enzymes that are present (most often in the case of tailed phages) on the basal plate. Such enzymes are called depolymerases, or exolysins. At the end of the lytic cycle of the phage, the choline proteins are synthesized. This form pores in the bacterial cytoplasmic membrane that enables endolysins to access and degrade the peptidoglycan. Because the cells of Gram-positive bacteria are not surrounded by outer membranes, endolysins can degrade the peptidoglycan even when outside the bacterial cell [40, 41].
The enzyme lysozyme, located at the end of the phage’s tail, participates in “piercing” of the bacterial cell wall. After piercing the cell wall, the phage DNA passes through the tail cavity and is injected into the cell cytoplasm. The remaining structural elements of the phage (capsid and tail) stay outside the cell. Under the action of phage lysozyme and intracellular osmotic pressure, the cell wall is destroyed, the phage offspring are released into the environment, and the bacteria are lysed. Cell lysis under the action of endolysins occurs either at the stage of bacteriophage adsorption on the cell surface (external lysis) or at the end of the lytic cycle of bacteriophage development owing to the destruction of the cell wall by the endolysin–choline–spanin system or by the lysis system of a single protein (internal lysis) [42, 43, 44].
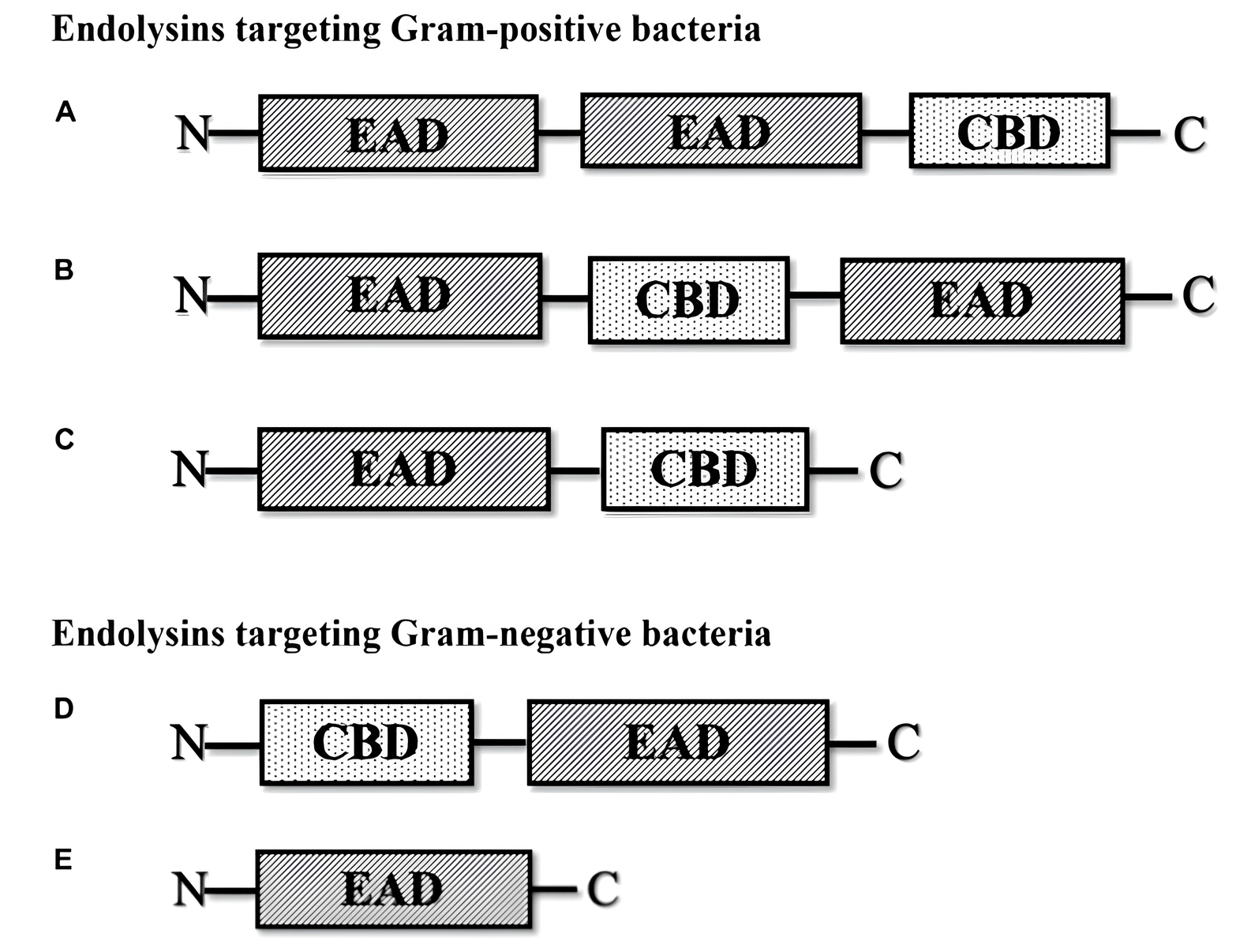
Because the cell membrane structures in Gram-positive and Gram-negative bacteria differ, endolysins have different effects on cells [45, 46, 47]. Endolysins can interact with the glycans of Gram-positive bacteria. After the endolysin makes a rupture in the peptidoglycan membrane, internal osmotic pressure causes the cell cytoplasm to flow out, which results in cell death [48, 49, 50, 51, 52]. Gram-negative bacteria have a multilayered peptidoglycan that is shielded from the environment by an outer membrane, which stops endolysins from working. Fig. 1 (Ref. [53]) shows a schematic diagram of the different types of phage-encoded peptidoglycan hydrolases (endolysins).
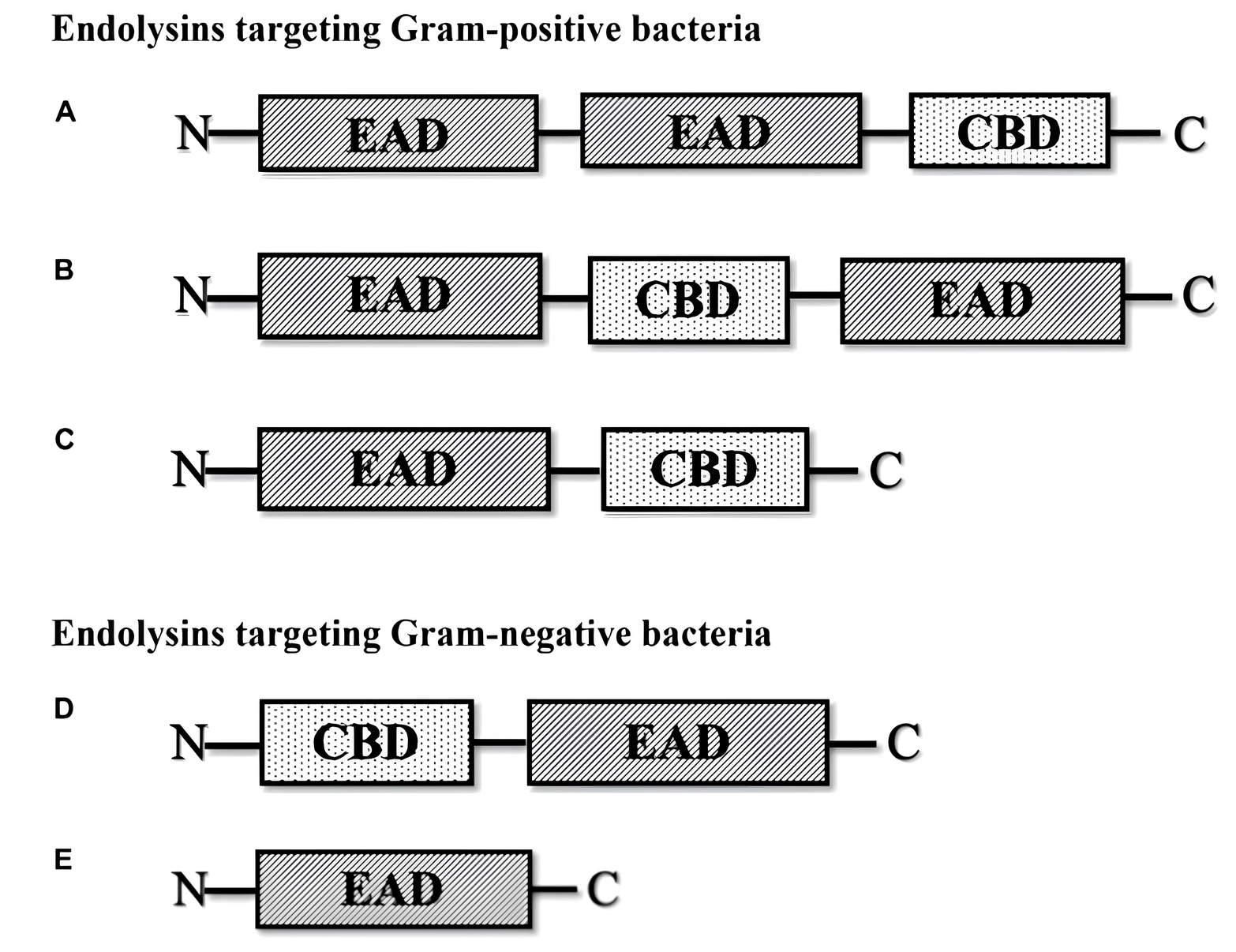 Fig. 1.
Fig. 1.
Different types of phage-encoded peptidoglycan hydrolases
(endolysins) [53]. (A) Multi-domain endolysin with more than one enzymatically
active domain (EAD) and one C-terminal cell wall-binding domain (CBD). (B)
Multi-domain endolysin with a centrally located CBD separating two EADs, found in
Streptococcus phage (
Gram-positive bacterial phage lysins typically have a C-terminal cell-wall-binding domain (CBD), which regulates the specificity of cell wall binding, and an N-terminal catalytic domain (CD), which regulates the enzyme catalytic activity. Nonetheless, the CD is frequently found at the C-terminus of Gram-negative bacteriophage lysins (such as P. aeruginosa phage lysin), whereas the CBD is found at the N-terminus [54, 55].
Most endolysins are formed from a single polypeptide chain (
Various forms of lytic endolysins exist, including acetylmuramidases, transglycosylases, glucosaminidase, amidases, and endopeptidases. Endolysins are classified into multiple functional categories of enzymes based on the types of enzymatic activity, regardless of the type of cleaved bond [49, 50, 51, 52]. Although endolysins differ in structure and mode of action, they have a common function [56]. Differences in the structure of protein domains ensure their selective specificity of interaction with the substrate, which is the bacterial cell wall [57, 58].
With the exception of lytic transglycosylases, all these enzymes are classified as hydrolases [59]. Peptidase or glucosaminidase activity is more prevalent among virion-associated lysins, whereas muramidase or amidase activity is most characteristic of endolysins. The kind of bond that the enzyme’s activity is aimed at determines its range of specificity.
Choline–endolysin and pincholine–signal-anchor-release (SAR) are two choline-mediated systems that aid endolysins. The cholines in the choline–endolysin system are phage-expressed hydrophobic membrane proteins that contribute to an increase in the permeability of the cytoplasmic membrane. This enables endolysins to enter the periplasm and target peptidoglycan [45]. Cholines facilitate the penetration of endolysins from the host bacterium’s cytoplasm to the peptidoglycan. They are generated late in infection, and when present in sufficient quantities, they oligomerize to form holes in the cell’s cytoplasmic membrane, which gives cytoplasmic endolysins access to peptidoglycan [60, 61, 62, 63].
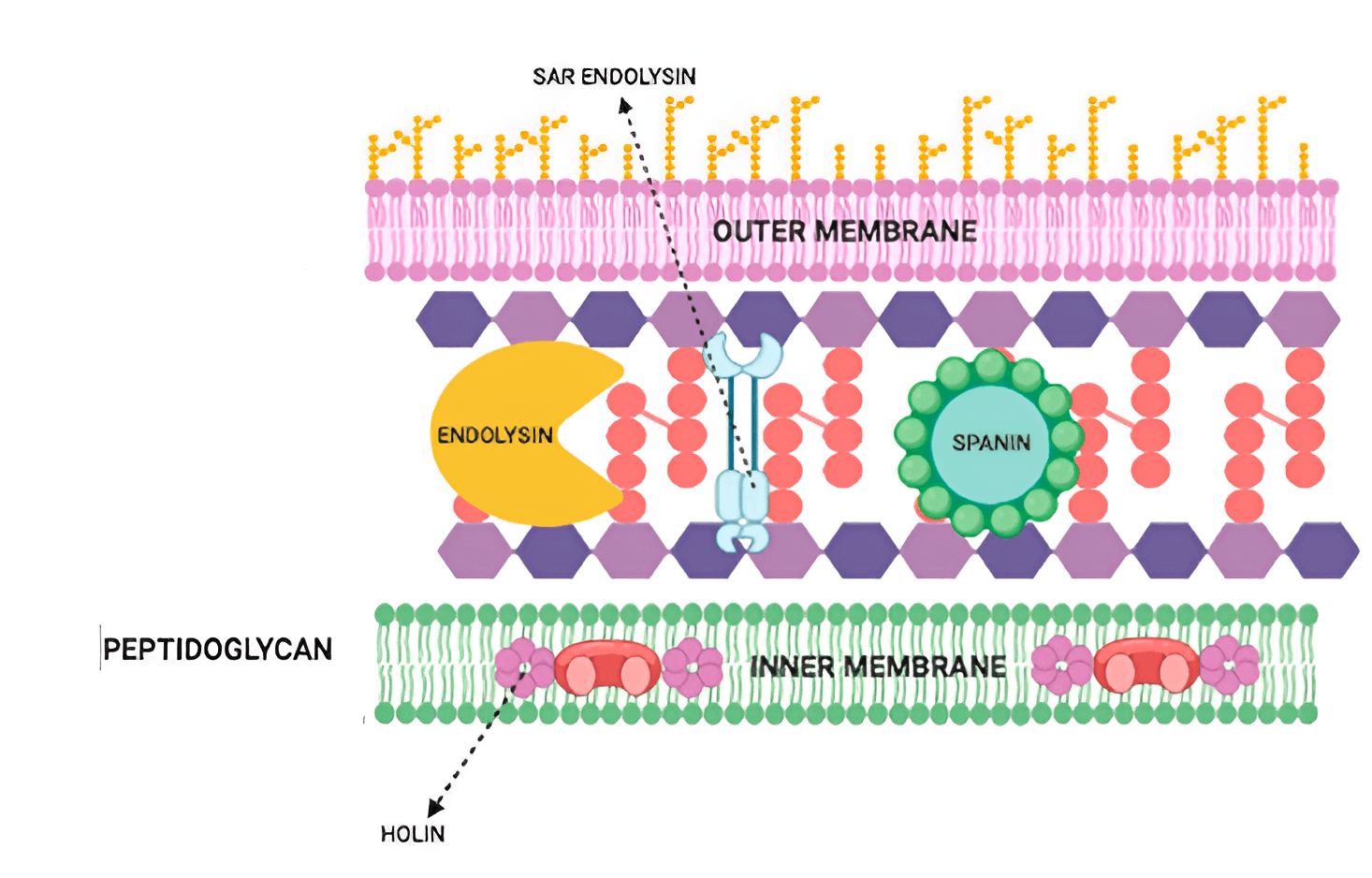
The host Sec system exports endolysins in the pinhole–SAR system to the periplasm, where they gather as membrane-anchored dormant proteins. Endolysin activation is timed by pinholins. In contrast to cell rupture in the choline–endolysin system, SAR endolysins are uniformly distributed in the periplasm before pinholin activation, which results in more uniform breakdown of peptidoglycan and progressive cell shortening before lysis [45]. Below shows the mechanism of endolysin action on Gram-negative (Fig. 2, Ref. [39]) and Gram-positive (Fig. 3, Ref. [41]) microbial cells.
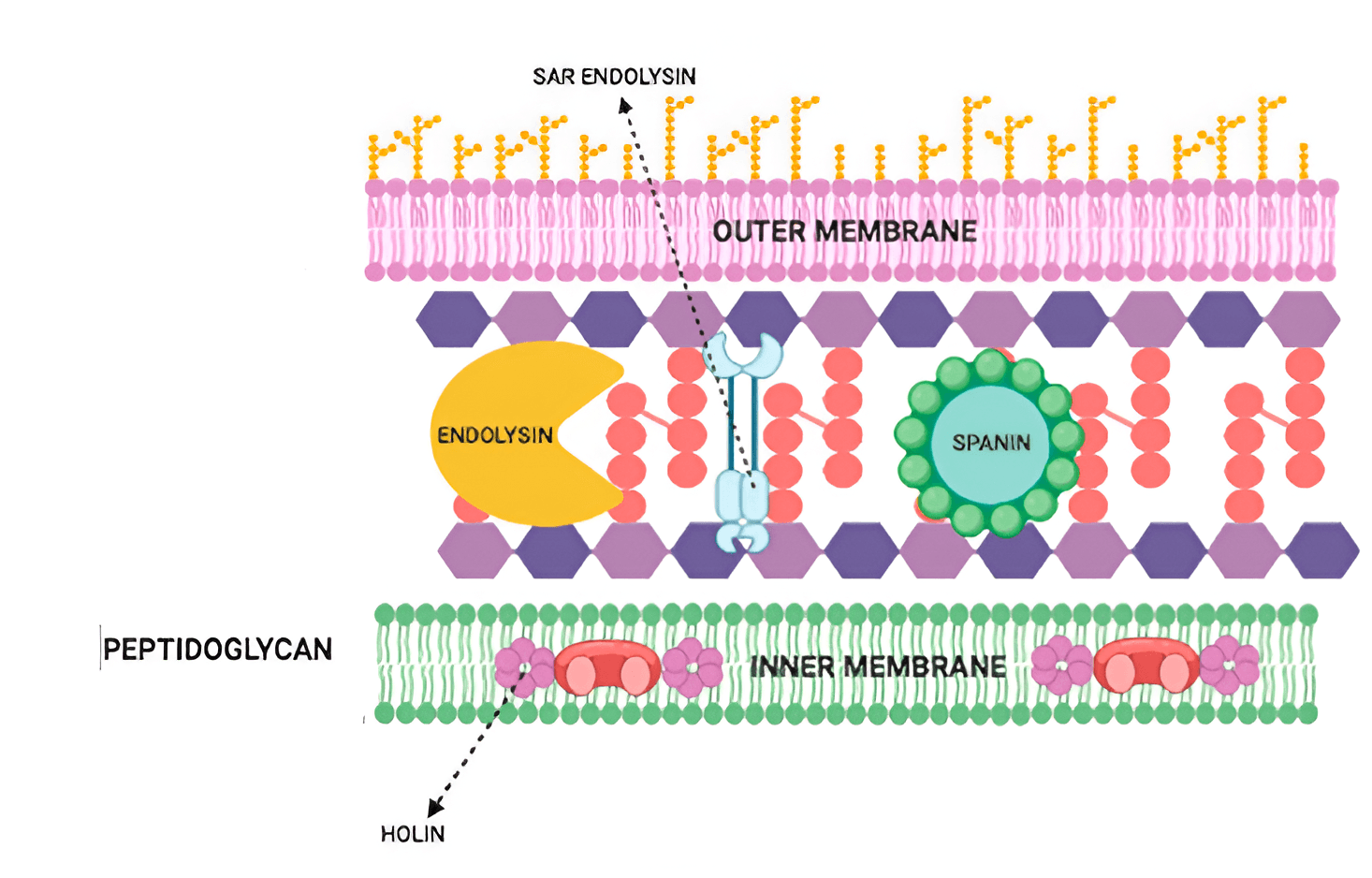 Fig. 2.
Fig. 2.
Mechanism of endolysin action on Gram-negative microbial cells [39]. Reprinted under terms of the CC-BY license. Antibiotics; published by MDPI, 2021. SAR, Signal-arrest-release.
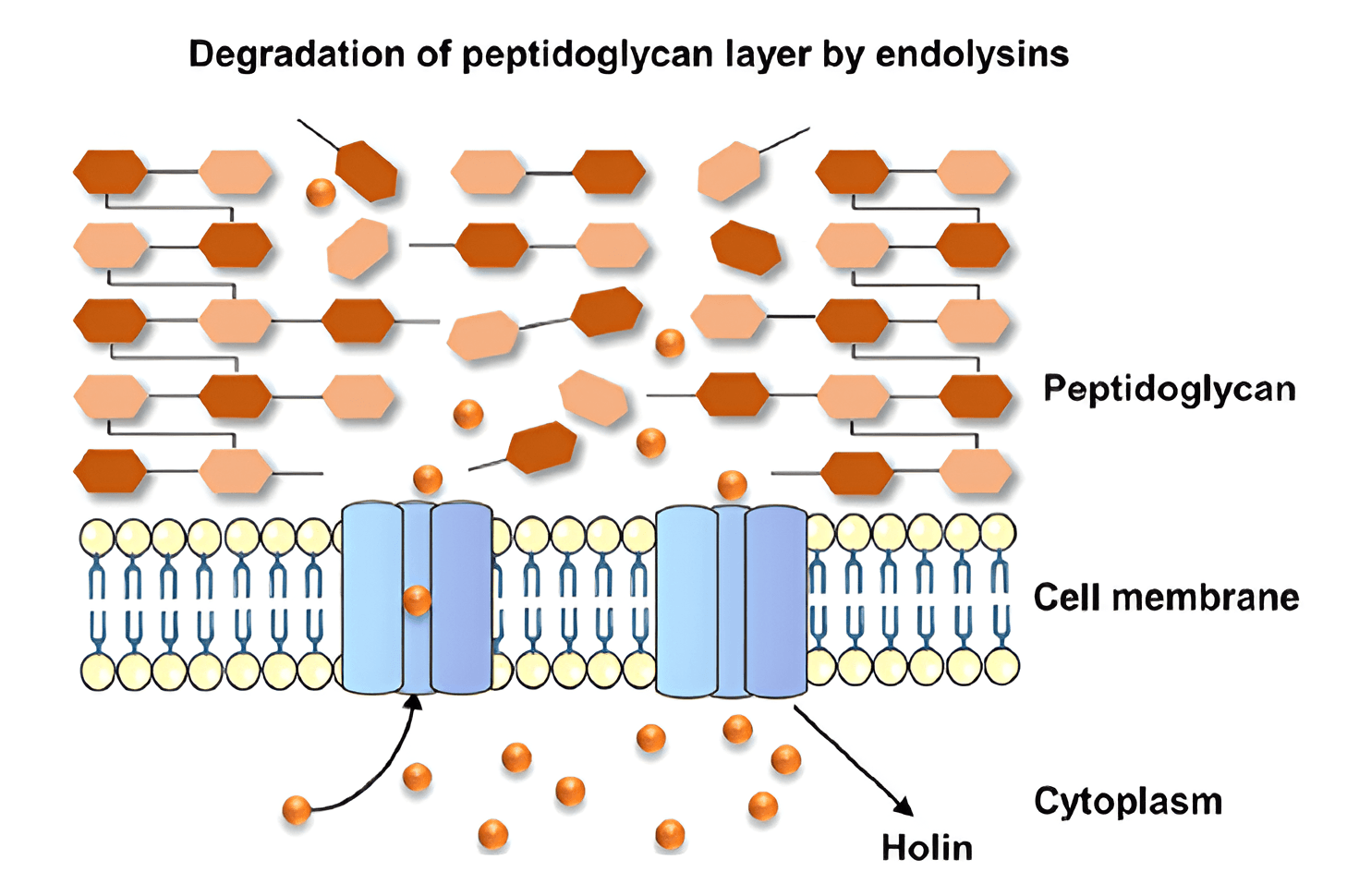 Fig. 3.
Fig. 3.
Mechanism of endolysin action on Gram-positive microbial cells [41]. Reprinted under terms of the CC-BY license. Journal of Biomedical Science; published 2023.
Because the amide bond of peptidoglycan and the
Endolysins have garnered special attention since discovery because of their potential as substitute medicinal agents [65, 66]. As a rule, endolysins have bactericidal activity against bacteria of the same genus or a group of similar species within the genus of microorganisms that the bacteriophage infects. Such a narrow specificity of action, in contrast to the action of classical antibiotics, allows selective suppression of pathogens, rather than of the entire normal flora of the macroorganism [50]. However, lysins characterized by broad-spectrum activity have been reported, such as Salmonella phage BSPM4 endolysin M4Lys, which targets many Gram-negative bacteria (various serovars of Salmonella enterica, Escherichia coli, Pseudomonas aeruginosa, and Cronobacter sakazakii) in vitro [67], and the phage endolysin LysSAP26, which has a broad-spectrum activity and can inhibit the growth of several drug-resistant species, including Acinetobacter baumanii, E. coli, Klebsiella pneumoniae, P. aeruginosa, S. aureus, and Enterococcus faecium [68].
It is crucial to treat illnesses brought on by Gram-positive bacteria. Foodborne bacteria endanger humanity by becoming more and more common. Food safety and human health are strongly affected by foodborne bacteria, which are among the main causes of food poisoning. S. aureus is one of the most prevalent and deadly foodborne bacteria, and it has a major effect on healthcare costs worldwide. It can be lethal if it enters the heart, lungs, or bloodstream and is a major cause of infections of the skin and soft tissues. S. aureus is also responsible for up to 30% of nasal infections in humans worldwide [69].
Significant morbidity and mortality are linked to infections caused by S. aureus, which frequently shows resistance to many active antibacterial drugs. These infections can produce a range of symptoms, from diarrhea and sepsis to potentially fatal endocarditis. For effective treatment of infections linked to this pathogen, new strategies must be developed, particularly those that use endolysins. For S. aureus infections, particularly those caused by methicillin-resistant strains (MRSA), new therapeutic strategies are required. The first-in-class antistaphylococcal lysin exebacase is a direct lytic agent that works in concert with antibiotics to quickly break down biofilms [70]. Table 1 (Ref. [61, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126]) summarizes the action of endolysins on microbial cells in planktonic and biofilm bacteria.
| Endolysins | Microbial cells | Inhibitory concentration | Refs. |
| P128 | S. aureus | 1–64 µg/mL | [71] |
| HY-133 | S. aureus | 0.12–0.5 µ/L | [72, 73, 74, 75] |
| LysK, phiSH2, P68, LysWMY, 80 |
S. aureus | 0.2 µM | [76] |
| PRF-119 | S. aureus | 0.024–1.563 µg/mL | [77] |
| LysSYL | S. aureus | 0.25–256 µg/mL | [78] |
| LysSA11 | S. aureus | 1.35 µМ | [79] |
| LysGH15 | S. aureus | 50 µg/mL | [80] |
| Lys84 | S. aureus (MRSA) | 10 µM | [81] |
| XZ.700 | S. aureus (MRSA) | 12.5–50 µg/mL | [82] |
| MV-L | S. aureus | - | [83] |
| SAP-2 | S. aureus | 5–20 µg/well | [84] |
| LysSA12, LysSA97 | S. aureus | 0.3 µM | [85] |
| LySMP | Str. suis | 100 IU/well | [86] |
| LySMP | Str. suis | 50 µg/mL | [87] |
| P128 | S. epidermidis, S. lugdunensis, S. haemolyticus | 15.0–31.0 µg/mL | [88] |
| S. epidermidis, S. haemolyticus | 62.5 µg/mL | ||
| CF-301 | S. aureus (MRSA) | 3.2 µg/mL | [89] |
| Lys22 | Enterococcus faecalis, | 25 µМ–100 µМ | [61] |
| S. aureus, A. baumannii | |||
| Ply113 | Enterococcus faecium, | 0.5–256 µg/mL | [90] |
| E. faecalis, | |||
| S. aureus | |||
| ClyR | E. faecalis | 50 µg/mL | [91] |
| lys08 | E. faecalis | 100 µL (50 and 100 µg) | [92] |
| LysEF-P10 | E. faecalis | 20 µg/mL | [93] |
| LysIME-EF1 | E. faecalis | 0.1 µM | [94] |
| Cpl-1 | Str. pneumoniae | 16 µg/mL | [95, 96] |
| Cpl-7 | Str. pneumoniae | 25 µg/mL | [97, 98] |
| ClyR | Streptococcus mutans MT8148, | 50 µg/mL | [99] |
| Str. sobrinus ATCC6715, | |||
| Str. sanguinis, Str. oralis, | |||
| Str. salivarius | |||
| SP-CHAP | Str. pneumoniae | 1.56 µg/mL | [100] |
| PlySs2 and PlySs9 | Str. uberis | 13.45, 27.05, 8.60, and 10.90 µg/mL | [101] |
| LyJH307 | S. bovis, E. faecalis, | 50 µg/mL | [102] |
| S. sanguinis | |||
| PlyC | Str. pyogenes | 10 ng/mL | [103] |
| PlyPy | Str. pyogenes | 0.5 U/µg | [104] |
| PlyC | Str. pyogenes | MIC 0.02–0.08 mg/L | [105] |
| MBC 0.02–1.25 mg/L | |||
| ClyV | S. agalactiae | 0, 2, 4, 8, 16, and 32 µg/mL | [106] |
| PlyGBS | Str. agalactiae | 10 U | [108] |
| PlySK1249 | Str. agalactiae | 3.3 U/mL | [107] |
| Ply5218 | Streptococcus suis | 1.25, 2.5, 5, 10, 20, and 40 µg/mL | [109] |
| LysCP28 | Clostridium perfringens | 10 µg/mL (plankton) | [110] |
| 18.7 µg/mL (biofilm) | |||
| PlyG | Bacillus anthracis | 0.5–20 U | [111] |
| LysB4 | B. anthracis | 5 µg/mL | [112] |
| PlyB | B. cereus | 1–5 g/mL (57.9 nM and 579 nM) | [113] |
| TP84_28 | Geobacillus stearothermophilus | 0.2–2 µg/mL | [114] |
| Artilysin® Engineered | P. aeruginosa | 10–2000 nM | [115] |
| Endolysin-Based | |||
| (PVP-SE1gp146) | |||
| PlyPa03, PlyPa91 | P. aeruginosa | 0.2–125 µg/mL | [116] |
| LysECD7 (E. coli phage) | K. pneumoniae | 1000 and 3000 µg/mL (62 and 186 µM) | [117] |
| LysAp22, LysECD7, LysAm24, and LysSi3 | Gram-negative pathogens, including multidrug-resistant | 100 µg/mL | [118] |
| K. pneumoniae, Salmonella sp., | |||
| P. aeruginosa, E. coli, | |||
| A. baumannii, and Enterobacter sp. | |||
| LysAB1245 | E. coli, K. pneumoniae, | MIC 4.68–9.36 µg/mL | [119] |
| P. aeruginosa, and S. aureus | MBC 4.68–18.72 µg/mL | ||
| PlyF307 | A. baumannii | 1 mg PlyF307 (250 µL) | [120] |
| LysSS | A. baumannii, | 0.063–0.25 mg/mL | [121] |
| E. coli, K. pneumoniae, | |||
| P. aeruginosa, and Salmonella sp. | |||
| Lysep3 | E. coli | 5.59 µg/µL, | [122] |
| final concentration 1.75 µg/µL | |||
| PLyE146 | E. coli, P. aeruginosa, | 400 µg/mL | [123] |
| A. baumannii | |||
| 36 endolysin candidates LytC | G. vaginalis ATCC14018 | Biofilm |
[124] |
| CCB8.1 = 25 µg/mL | |||
| PM-477 | Gardnerella | 1 to 4 mg/mL | [125] |
| CCB7.1 | Gardnerella (G. vaginalis, Fannyhessea vaginae, Prevotella bivia, and Mobiluncus curtisii) | 1–128, 256, and 512 µg/mL | [126] |
For example, by combining the lethal activity of the staphylococcal cell-wall-targeting domain of lysostaphin (SH3b) with the phage tail–associated muralytic enzyme, phage K, in hydrogel form, phage endolysin P128 (a chimeric protein) efficiently lysed clinical staphylococcal isolates extracted from human nares [71]. Rapid bactericidal and highly selective activity against S. aureus was shown in vitro for recombinant bacteriophage endolysin HY-133 [72].
HY-133 was active in vitro against the main African strains of S. aureus, both susceptible and resistant to methicillin, and against S. schweitzeri [73]. It was also highly active in vitro against a large collection of genetically diverse methicillin-resistant isolates [74]. In comparison to daptomycin and mupirocin, HY-133 was highly active against all antibiotic-resistant isolates tested, including those resistant to mupirocin. It was active at low concentrations when tested against bacteria in either stationary or exponential growth phase [75].
In another study, bacteriophage endolysins [peptide glycan hydrolases (PGHs)] were proposed as promising antimicrobial agents for S. aureus [76]. The chimeric endolysin PRF-119 was active against MRSA [77]. In comparison to its parent phage, the stable endolysin LysSYL showed a greater range of potency against staphylococcal strains (100% vs. 41.7%). It was also able to eradicate persistent S. aureus in conjunction with vancomycin. The data show substantial therapeutic potential of LysSYL for the treatment of staphylococcal infections, including S. aureus mono-and mixed biofilms [78].
The endolysin LysSA11 has an enzymatically active CHAP (cysteine, histidine-dependent amidohydrolases/peptidases) domain at its N terminus and can be used as a lytic agent in dishwashing and as an antibiotic for food [79].
Because it has a broad-spectrum lytic activity and a very effective lytic activity against MRSA without conferring resistance after repeated treatment, the lysine from the staphylococcal phage LysGH15 (GH15) is an alternative therapy option for MRSA infections [80].
In biofilm-associated infections, the effectiveness of antibiotic treatment is strongly reduced. Consequently, devising a successful treatment plan is crucial to preventing infections brought on by polymicrobial biofilms. Endolysins hold considerable promise for treating bacterial forms linked with biofilms, because they can eradicate antibiotic-resistant bacteria, as well as eliminate biofilms. Endolysins’ efficacy has been consistently shown both in vitro and in vivo. As biological agents, endolysins are promising in resolving issues related to biofilms. Some findings show remarkable potential of modified phages in the fight against complicated illnesses, superbugs, and antibiotic-resistant bacteria [127]. The endolysin Lys84 was strongly lytic against S. aureus and effectively removed about 90% of biofilms [81]. Another endolysin, PlyGRCS, showed dose-dependent efficacy against S. aureus in biofilms and planktonic forms, including MRSA [128]. In a study of the effect of the endolysin XZ.700 on biofilms of MRSA strains grown on titanium, biofilm viability decreased depending on the endolysin concentration [82]. Other variants of phage endolysins active against S. aureus have been shown elsewhere [83, 84, 85].
For instance, biofilms produced by Streptococcus and Staphylococcus species may be removed by the modified endolysins LysSMP and SAP-2 [86, 87]. The endolysin P128 is active against biofilms of S. epidermidis, S. lugdunensis, and S. haemolyticus [88].
Biofilms that develop in human synovial fluid, including drug-resistant S. aureus biofilms, can be lysed by the lysin CF-301 with remarkable efficiency. After CF-301 had been administered, all biofilms in the catheter were eliminated and the bacterial population was reduced by five orders of magnitude [89].
The application of endolysins from staphylococcal bacteriophages and their effectiveness against Staphylococcus species were reviewed by Golban et al. [129].
Enterococcus faecalis is an opportunistic pathogen that causes diseases ranging from urinary tract infections to sepsis in humans and animals. Endolysins are being actively tested to treat infections caused by these bacteria. E. faecalis is a major cause of healthcare-associated infections with high fatality rates [130] and can withstand harsh conditions [131, 132]. It is the third most common nosocomial bacterial pathogen in the world, is highly virulent, and has minimal degrees of acquired and intrinsic antibiotic resistance, especially in biofilm form [133].
An enterococcal phage-derived endolysin called Lys22 showed a wide range of host compatibility, effectively inhibiting both simple and complex biofilms in vitro and being effective against E. faecalis, many Staphylococcus species, and A. baumannii in planktonic form [61].
The endolysin Ply113 showed antibacterial and antibiofilm activities in vitro against S. aureus, E. faecalis, and E. faecium, including isolates of MRSA and vancomycin-resistant Enterococcus [90].
A principal cause for periapical endodontic lesions is E. faecalis, an opportunistic pathogen that can be present in the human intestine, the mouth cavity, and the vaginal tract. However, few therapeutic alternatives are available, because E. faecalis is strongly able to produce biofilms and is resistant to many antibiotics and therapies. ClyR, a chimeric lysin with wide-spectrum lytic action on streptococci, is effective against E. faecalis planktonic and sessile cells in a dental model both in vitro and ex vivo [91]. E. faecalis biofilms were susceptible to the pathogenic bacteriophage PHB08 and its endolysin lys08 [92]. The endolysins LysIME-EF1 and LysEF-P10 are effective against several strains of E. faecalis [93, 94].
The Streptococcaceae family includes Gram-positive, facultatively anaerobic, spherical, or ovoid asporogenous bacteria, which parasitize both humans and animals, inhabiting the digestive and respiratory systems, particularly the large intestine, the nose, and the mouth. One alternative strategy for treating such infections is the Cpl-1 lysin, a pneumococcal phagolytic muramidase, which rapidly kills streptococci [95, 96]. Another study used the endolysin Cpl-7 to combat Str. pneumoniae [97, 98].
Dental caries is a common ailment caused by oral germs. Two main cariogenic bacteria, Str. mutans and Str. sobrinus, are often found on teeth as biofilms. The efficacy of ClyR, a chimeric lysin, against Gram-positive oral bacteria and the activity of its antagonists were evaluated in rats. ClyR was highly lytic to S. mutans MT8148 and S. sobrinus ATCC6715 but had little effect on the innocuous commensal oral bacteria Str. sanguinis, Str. oralis, and Str. salivarius. ClyR may, therefore, be a helpful drug or supplement for both treating and preventing dental cavities [99]. Without affecting the various oral or nasal commensal organisms investigated, the first pneumococcal endolysin with a catalytic domain was discovered to be more active than the best studied pneumococcal endolysin (Cpl-1). It eliminated biofilms and was active against all serovars tested, including capsular and acapsular pneumococci [100].
Str. uberis is frequently responsible for mastitis in cattle. Mastitis is an infection of a cow’s mammary gland, which results in considerable financial losses for the dairy sector. The endolysins PlySs2 and PlySs9, derived from Str. suis prophage serotypes 2 and 9, have potential as an antibacterial arsenal against S. uberis [101].
An important cause of acute acidosis in ruminants, Str. bovis, is a group of Gram-positive bacteria first identified as a species linked to urinary tract infections, endocarditis, sepsis, and colorectal cancer in humans. Recombinant LyJH307 is bactericidal to S. bovis JB1 and bacteria belonging to the Lancefield D (S. bovis and E. faecalis) and H (S. sanguinis) groups [102]. The special murein hydrolase is effective at eradicating streptococci in the epithelium of the mucous membrane of the upper respiratory tract of both carriers and infected individuals. It also has a quick lethal effect on group A streptococci both in vitro and in vivo without affecting other local microorganisms examined [103]. The group A streptococcus Str. pyogenesis is prone to colonize human tissues and form biofilms. The most powerful lysin yet identified is PlyC (group C streptococcal phage lysin), which shows remarkably high specificity for a range of streptococcal species, including Str. pyogenes [104, 105].
Str. agalactiae
An important zoonotic pathogen, Str. suis serotype 2 (SS2), causes meningitis, arthritis, endocarditis, sepsis, pneumonia, and sudden death in humans and pigs. Strong lytic activity against S. suis was shown by the Ply5218 lysin [109]. The varieties of streptococcal endolysins and their potential for use as antibacterial drugs were described in detail in Wong et al. [53].
Gram-positive, spore-forming, and obligatorily anaerobic, Clostridium perfringens is frequently found in soil, wastewater, and the gastrointestinal tracts of both humans and animals. The lysin LysCP28 was effective in both biofilm removal and prevention, and it showed a wide range of lytic activity against C. perfringens types A, B, C, and D from various sources [110].
The Bacillus cereus group of bacteria (or B. cereus sensu lato) includes more than 20 species, among them B. cereus sensu stricto, which causes food poisoning; B. thuringiensis, which is used as a bioinsecticide; and B. anthracis, the causative agent of anthrax. In vitro and in vivo, isolates of B. anthracis and other members of the B. anthracis bacillus “cluster” are particularly killed by the PlyG lysin, identified from the B. anthracis phage [111].
By taking advantage of the genetic material from bacteriophages AP50 and B4, recombinant phage endolysins AP50-31 and LysB4 were obtained and used against a variety of Bacillus strains, including those of Bacillus anthracis (anthrax) [112].
B. thuringiensis, B. anthracis, and B. cereus sensu stricto were among the species in the B. cereus sensu lato group, in which the antibacterial properties of the endolysin PlyB were examined [113].
The Bacillota phylum includes the rod-shaped, Gram-positive bacterium Geobacillus stearothermophilus. This thermophilic bacterium, present in soil, hot springs, and ocean sediments, causes food to spoil. The potential ability of endolysins to inhibit the growth of Geobacillus stearothermophilus biofilm was described in Żebrowska et al. [114].
Endolysins are, therefore, promising ingredients in the fight against Gram-positive bacterial infections. Endolysins show promise as antibacterial agents and can effectively supplement the current class of antibacterial medications. This is crucial in the case of Gram-negative pathogens, dealt with in the following section.
The impermeable lipopolysaccharide coating that envelops the cell wall of Gram-negative pathogens is a primary barrier to endolysin use as antibacterial agents. To make endolysins bactericidal for Gram-negatives such as P. aeruginosa and Acinetobacter baumannii, researchers devised an optimal strategy to enable the enzymes to penetrate the bacterial outer membrane (artilysins) [115, 116, 134] (Table 1).
The endolysins PlyPa03 and PlyPa91 showed high killing kinetics against P. aeruginosa, Enterobacter, and Klebsiella at pH values ranging from 5.0 to 10.0. At high salt and urea concentrations, these lysins were active [116].
Surfaces of implanted medical devices are especially susceptible to biofilms. By
blocking antibiotic penetration and largely increasing the mechanical stability
of the colonizing community, biofilm bacteria increase morbidity and death in
clinical settings. Therefore, new tactics to counter biofilms are being developed
[33]. Surfaces of medical devices may harbor biofilms of the bacterium
Klebsiella pneumoniae [135]. For instance, a rat model was used to
assess the ability of the endolysin LysECD7 to inhibit K. pneumoniae
biofilms. The bacterial count in the biofilm that had developed on the implant
surface decreased from 1.0
The activity of the endolysins LysAm24, LysAp22, LysECD7, and LysSi3 against Gram-negative bacteria was thoroughly investigated, as were their biological safety and in vitro and in vivo action. The endolysins were effective against biofilm and planktonic bacteria, and they provided a solid foundation for the development of safe drugs for the treatment of infections caused by Gram-negative bacteria [118].
The endolysin LysAB1245 was active against key pathogens such as A. baumannii, E. coli, K. pneumoniae, P. aeruginosa, and S. aureus [119].
One of the most prevalent hospital-acquired pathogens is Acinetobacter baumannii, an MDR Gram-negative bacterium. In vitro and in vivo, the lysine PlyF307 had a strong effect on planktonic and biofilm A. baumannii [120].
MRSA was inhibited by recombinant LysSS, which also was active against Salmonella sp., A. baumannii, the MDR E. coli, K. pneumoniae, and P. aeruginosa [121]. The arsenal of endolysins used against A. baumannii was shown by Khan et al. [136].
The rod-shaped Gram-negative bacterium E. coli is frequently found in the lower intestines of warm-blooded mammals. The O157:H7 serotype of E. coli can cause serious food poisoning in both people and animals; however, most strains are safe. At the proper endolysin concentration, an endolysin treated with hydrophobic amino acids was able to kill E. coli. The lysing ability was improved by adding more hydrophobic amino acids to the endolysin’s C terminus [122].
E. coli prophage encodes the PlyE146 lysin, which was identified, characterized, and shown to be active against several strains of E. coli, P. aeruginosa, and A. baumannii [123].
Preterm delivery, an increased risk of HIV, and other sexually transmitted diseases are often associated with bacterial vaginosis, a recurrent dysbiosis. It is common for healthy women to have trace numbers of the Gram-variable facultative anaerobic bacterium G. vaginalis in their vaginal flora. Gardnerella vaginalis is a resistant biofilm that is a primary cause for this dysbiosis. Expansion of vaginal biofilms caused by the first colonization of G. vaginalis leads to recurrent symptomatic bacterial vaginosis, which is difficult to treat with conventional antibiotics. In E. coli BL21 (DE3), 36 putative endolysins were overexpressed, and 5 endolysins differing in their structure and biochemistry were comprehensively described. Every putative endolysin was highly lytic to both clinical isolates of G. vaginalis and planktonic G. vaginalis ATCC14018 [124]. Because Gardnerella species have a thin layer of peptidoglycan, endolysins have been made to target them [137, 138]. These bacteria make up the most of the biofilms linked to the emergence of bacterial vaginosis. Gardnerella-targeting endolysins were proposed as novel therapeutic agents and found to be effective against both monospecies [125] and polymicrobial biofilms (G. vaginalis, Fannyhessea vaginae, Prevotella bivia, and Mobiluncus curtisii) [126]. Therefore, phage endolysins can overcome the antibiotic resistance of Gram-negative bacteria.
New antimicrobial treatment alternatives that can be used as standalone therapeutic agents or in conjunction with other components are required owing to the increasing difficulty of treating illnesses caused by antibiotic-resistant bacteria. The pursuit of methods to boost endolysins’ antibacterial activity is a distinct field of study. Table 2 (Ref. [115, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152]) summarizes the main combinations of endolysins with other active components.
| Endolysin | Symbiotic component | Microbial cells | Refs |
| OBPgp279 and phage PVP-SE1 gp146 | Parasin I, lycotoxin I, and polycationic peptide | P. aeruginosa, A. baumannii | [115] |
| Cpl-1, pal | Cefotaxime and moxifloxacin | Str. pneumoniae | [139] |
| Cpl-1 | Gentamicin, penicillin | Str. pneumoniae | [140] |
| SAL200 | Nafcillin and vancomycin | S. aureus | [141] |
| ORF | Cefotaxime | E. faecalis | [142] |
| LysSSE1, HolSSE1 | Shigella | [143] | |
| P1y187AN-KSH3b | Gentamicin | S. aureus | [144] |
| LysAB3 | Threeantibiotics | A. baumannii | [145] |
| LysH5 | HydH5 | S. aureus | [146] |
| Ply2660 | Peptide LL-37 | E. faecalis | [147] |
| AbEndolysin | Cecropin A | A. baumannii | [148] |
| LysE | Сolistin | Aeromonas hydrophila | [149] |
| Lys68 | EDTA, citric acid, and malic acid | Pseudomonas strains (EDTA/Lys68 combination) | [150] |
| Salmonella typhimurium LT2 (combination of citric or malic acid with Lys68) | |||
| K-L | Anionic polymer matrices | S. aureus | [151] |
| Lysqdvp001 | V. parahaemolyticus | [152] |
Several endolysins in combination with antibiotics can lead to a synergistic effect and improve bactericidal effects.
Phage–antibiotic synergy (PAS) occurs when phages infect bacteria in the
presence of sublethal antibiotic concentrations. A broad variety of
bacteria–phage and bacteria–endolysin combinations with various antibiotic
classes for both Gram-positive and Gram-negative bacteria were included in a PAS
observation [153]. Progress has been made in eliminating bacterial biofilms by
combining bacteriophages with antibiotics for synergistic effects and effective
use of phage lysins. The potential for phages and lysins alone and/or in
combination with antibiotics to combat biofilms was shown by
Łusiak-Szelachowska et al. [154]. A number of studies have
demonstrated a synergistic effect of the lytic enzyme S. pneumoniae
Cpl-1 with antibiotics and anionic polymers [139, 140]. Kim et al. [141]
showed the antistaphylococcal activity of the endolysin SAL200 when used in
conjunction with vancomycin and nafcillin. The potential of cefotaxime and phage
endolysin ORF 6 for treating E. faecalis infections was investigated in
Oh et al. [142]. Phages SSE1, SGF2, and SGF3, the lytic proteins they
express (the lysozyme LysSSE1 and the holin HolSSE1), and
Mixing the HydH5 and LysH5 endolysins of S. aureus phage phiIPLA88 yielded better antistaphylococcal effects in vitro [146]. In vitro and in animal models, Str. pneumoniae or its phage can be efficiently eradicated by cell wall endolysins. The primary autolytic enzyme of Str. pneumoniae is LytA, an N-acetyl cytosolic acyl-l-alanine amidase, breaks down S. pneumoniae biofilms. Other cell wall endolysins include LytC, Pal, Cpl-1, Cpl-7, and Ejl. Additionally, Lyt-A and Cpl-1 can work together to efficiently eliminate Str. pneumoniae biofilms. These investigations showed the superior cleaning and killing activities of phage-encoded endolysins toward biofilms and the bacteria they protected. Future therapy of infectious disorders linked to biofilms may benefit from this discovery [87].
Salmonella contamination of food products continues to be a health concern. This problem has been addressed by creating a foundation for future research into strategies to fight these infections by using the phage BPS15S6 and the endolysin LyS15S6. LyS15S6 is a promising antibacterial drug that may be used to fight Enterobacteriaceae pathogens in food products [155].
The synergistic activity of endolysins was investigated to boost the therapeutic efficacy of antimicrobial peptides. For instance, the Ply2660–LL-37 combo worked in the following way: Ply2660 broke down the peptidoglycan in the cell wall, whereas LL-37 broke down the cytoplasmic membrane. Ply2660 and LL-37 worked together to prevent the growth of E. faecalis biofilms and to destroy the existing ones [147].
The combined action of the antimicrobial peptide cecropin A and the chimeric
AbEndolysin led to an increase in the antimicrobial activity of the peptide owing
to the increased cell membrane permeability and to the enhanced bactericidal
effect of the chimeric endolysin, which ultimately led to decreased bacterial
resistance to the endolysin. The chimeric eAbEndolysin had an additive effect
with meropenem and imipenem against systemic A. baumannii infection, as well as a
synergistic effect with
Recombinant LysE protein was highly effective against the Gram-negative bacterium Aeromonas hydrophila in combination with antimicrobial drugs (colistin) that alter the outer membrane permeability [149].
Depolymerases that target extracellular matrix polysaccharides have been suggested as components of a multienzyme strategy to eliminate biofilms. In static models, endolysins and depolymerases are effective against S. aureus biofilms. Both static and dynamic (flow-cell-based) models were used to examine the effectiveness of the endolysin LysK and the poly-N-acetylglucosamine depolymerase DA7 against staphylococcal biofilms. LysK showed anti-S. aureus action, and at low micromolar and nanomolar concentrations, respectively, LysK and DA7 eliminated static and dynamic biofilms from polystyrene and glass surfaces [156].
In recent years, phage endolysins have been studied in combination with membrane permeabilizers such as polymyxin B and ethylenediaminetetraacetic acid (EDTA) to penetrate the outer membrane barrier of Gram-negative bacteria [157]. P. aeruginosa phage lysin OBPgp279 and Salmonella phage PVP-SE1 gp146 were examined for their ability to produce fusion proteins with various outer-membrane-permeating agents (including hydrophobic pentapeptide, parasin I, lycotoxin I, and polycationic peptide). The “artilysins”, or outer-membrane-permeating endolysins, showed excellent lytic activity in vitro [115]. When mixed with organic acids, a Salmonella phage endolysin (Lys68) was active against Gram-negative bacteria. Research has demonstrated that the Lys68-EDTA complex lyses only some strains; however, when Lys68 is complexed with citric or malic acid, its antibacterial action is increased and extends to additional pathogens, such as Salmonella typhimurium LT2 [150].
The addition of a chimeric enzyme to anionic polymer matrices is used to reduce the immunogenicity of enzymes. For example, when the chimeric enzyme K-L is added to anionic polymer matrices, complexes with block copolymers of poly-l-glutamic acid and polyethylene glycol are formed, but the stability and activity of the enzyme are preserved. These data suggest that such a hybrid compound is effective as an antimicrobial agent [151].
The endolysin Lysqdvp001 and
The endolysin LysRODI was active against many S. aureus strains, including MRSA isolates. When encapsulated in pH-sensitive liposomes and then released from the nanocapsules, it retained 47% of its efficacy against bacteria [158].
In another study, a system for encapsulating the endolysin BSP16Lys in a cationic liposome composed of dipalmitoylphosphatidylcholine (DPPC), cholesterol, and hexadecylamine was developed and tested for its ability to penetrate the outer membrane of the Gram-negative bacteria Salmonella typhimurium and E. coli [159].
Thus, phage endolysins in combination with various biomolecules show promising results for combating pathogenic bacteria. Further prospects for the use of endolysins are aimed at clinical studies, which will be discussed below.
A quickly expanding area of the pharmaceutical industry is protein-based medications. In addition to 239 authorized medicinal proteins and peptides, the database currently includes 380 recognized variations of therapeutic proteins and peptides [160, 161].
Phage enzymes are far superior to antibiotics owing to their high selectivity [162]. WHO has recognized endolysins as new, nontraditional antimicrobial drugs. As of 2023, 15 bacteriophage-based drugs have been used in clinical trials, and they ranked first among nonclassical antibiotics [163].
Although endolysins have many advantages over antibiotics and superior competitive performance, their introduction as therapeutic drugs is associated with official approval for use in human and veterinary medicine. A weak point of endolysins is their poor water solubility owing to the strong hydrophobic and repetitive transmembrane region in their C-terminal binding domain. Natural phage generation and modification strategies are used to overcome this limitation and improve endolysins’ lytic activity and range of action [164].
With intravenous or intramuscular administration of endolysins, as with the introduction of other foreign antigens, the production of specific antibodies should occur. The presence of antibodies in the macroorganism may inactivate the bacteriophage enzymes after repeated administration. However, several in vitro experiments with endolysins against Bacillus anthracis, Staphylococcus aureus, Str. pneumoniae, and Str. pyogenesis showed that antibodies against endolysins do not block their bacteriolytic activity. In vivo experiments showed that after repeated therapy of bacterial infections with the Cpl-1 and Pal endolysins, mice did not lose their ability to recover from repeated infection and repeated administration of the same endolysins. Also, no anaphylactic shock was observed [50]. Thus, endolysin preparations can be successfully used for therapeutic purposes after they undergo the appropriate preclinical and clinical trials.
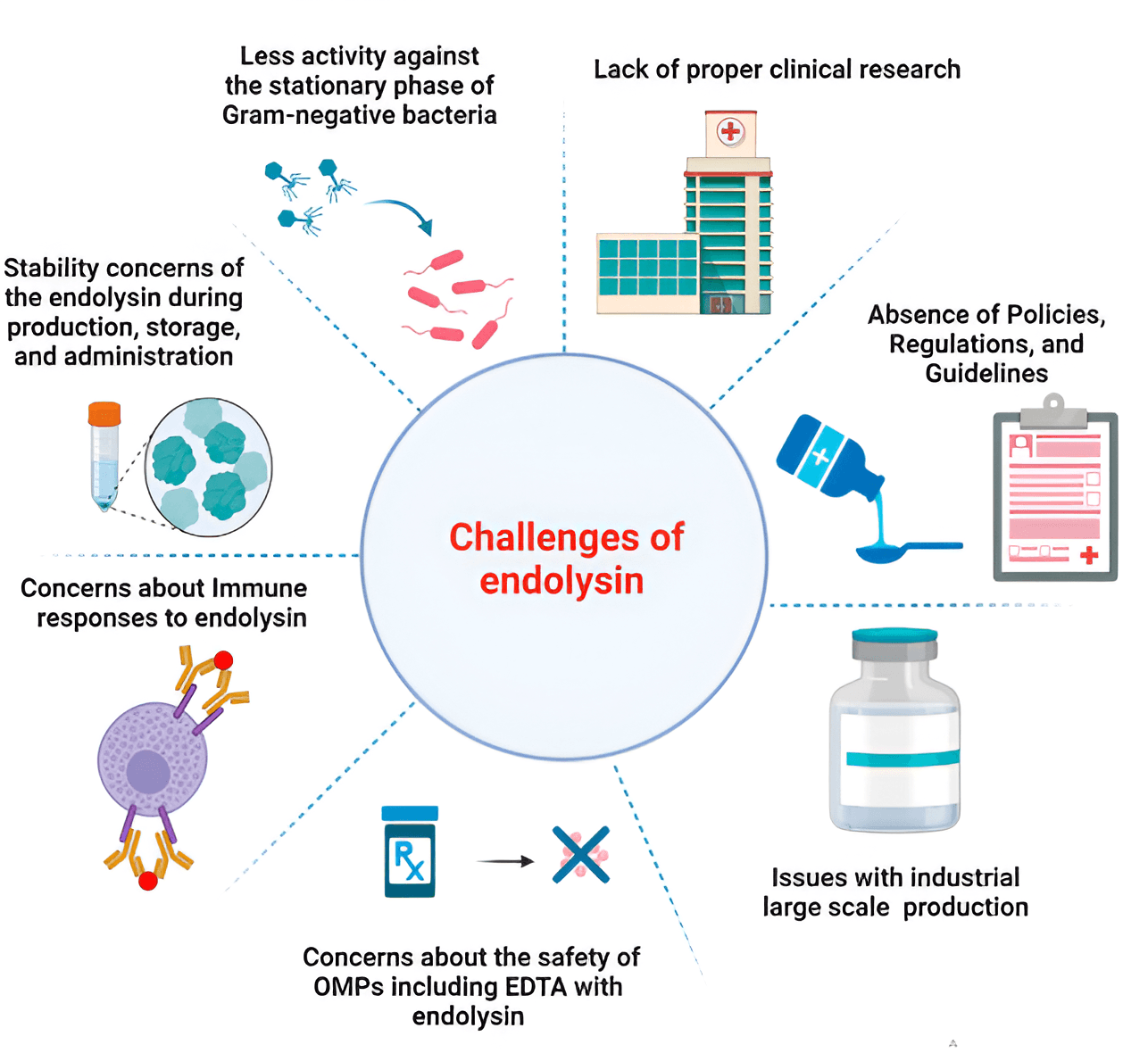
Before using endolysins as therapeutic agents in clinical practice, it is necessary to evaluate:
All these questions can be answered only after clinical trials are conducted [136].
Fig. 4 (Ref. [136]) summarizes the main problems of using endolysins in the clinic and in the healthcare market.
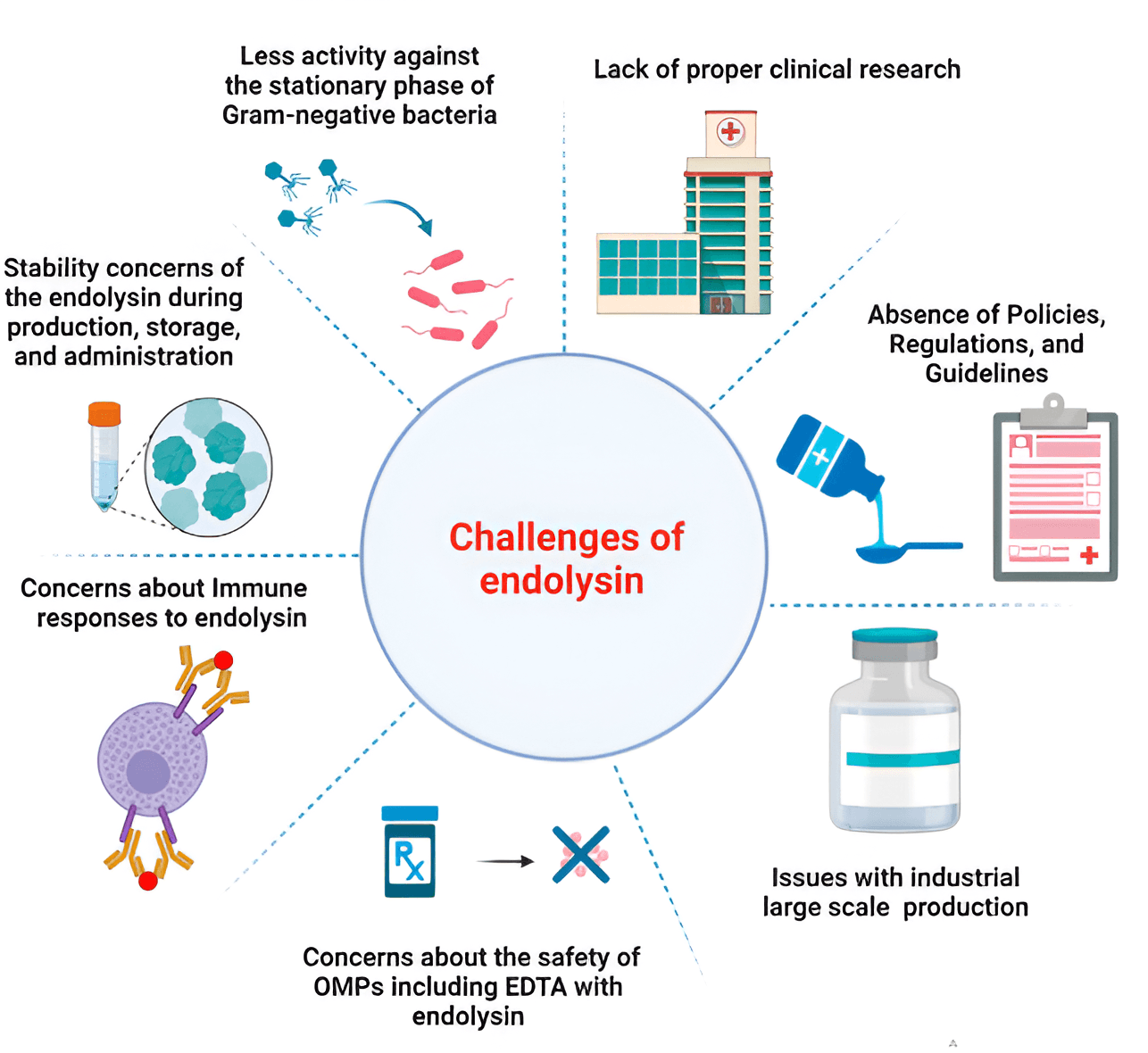 Fig. 4.
Fig. 4.
Main problems of using endolysins in the clinic and in the healthcare market [136]. Reprinted under terms of the CC-BY license. Front. Pharmacol.; published by 2024.
The phage lytic proteins P128, CF-301, and Staphefekt have been produced by the biotechnological companies GangaGen, ContraFect, and Micreos, which work with endolysins to combat S. aureus infection. Skin infections, bloodstream infections, and chronic kidney illnesses can all be effectively treated with these endolysin preparations [165, 166].
Currently, phage endolysins are being used in a number of human clinical trials. For instance, the endolysins SAL200 and CF-301 were used to combat S. aureus infection of the heart and bloodstream [167]. The endolysin SAL-1 was used to treat S. aureus infection in vitro and in vivo. SAL200 was effective against 425 clinical isolates from 421 humans and 4 animals, including 336 MRSA isolates and 1 vancomycin-intermediate S. aureus strain. When paired with calcium ions and poloxamer 188 as stabilizers and enhancers, SAL200 was very powerful against MRSA [168].
Endolysins have been used in human clinical studies by a number of biotech and pharmaceutical businesses. ContraFect (USA) is a biotech business that is leading the way in the development of therapeutic pharmaceuticals based on endolysins. The business has acquired the exclusive rights to nine phage endolysins. Endolysin No. NCT04160468 is undergoing a phase III clinical trial by ContraFect [136].
By recognizing ContraFect’s endolysin as a breakthrough therapy in phase III research, the FDA acknowledged endolysin innovation as an antibacterial biologic in February 2020. The first-in-class anti-Staphylococcus lysin, CF-301 (exebacase), proved effective against 120 MRSA strains out of 250 S. aureus strains examined. Exebacase can be used as a supplement to antibiotics and is the main ingredient in a novel family of antimicrobial drugs [169].
In a mouse with staphylococcal bacteremia, exebacase with antistaphylococcal drugs (vancomycin or daptomycin) showed synergism in vitro and increased animal survival [89]. In 2020, exebacase was used in phase III clinical studies to treat 350 patients with S. aureus–induced bacteremia. When it was combined with standard antibiotic therapy (14-day course), the number of patients with MRSA bacteremia and endocarditis increased from 30 to 70% [70]. Universities and businesses are working on the preclinical development of other endolysins [170]. To treat human infective endocarditis, ContraFect developed the exebacase Endolysin CF-301, a recombinant endolysin against a variety of Streptococcus and Staphylococcus species. In the US, CF-301 was the first of its kind to go through clinical trials on humans. When combined with antibiotics, it improved the rate of recovery from MRSA-induced infective endocarditis by 42.8% in a phase II clinical trial [171]. Rats given CF-301 with daptomycin showed superior bacterial clearance in acute MRSA osteomyelitis [172].
ContraFect’s “amurins”, a new class of phage-encoded lytic agents, target the most lethal drug-resistant Gram-negative bacteria, including biofilms (https://carb-x.org/carb-x-news/carb-x-funds-contrafect-to-develop-a-new-class-of-antibiotics-to-treat-serious-infections-caused-by-gram-negative-eskape-superbugs/).
SAL200, often referred to as N-Rephasin SAL200, is another phage-derived endolysin that has potential in therapeutic contexts. In Jun et al.’s study [167], 34 male participants in good health received intravenous injections of SAL200. When the pharmacokinetic characteristics of the endolysin and the tolerance for it were examined, no notable side effects were found.
In 2021, iNtRON Biopharma, another business, finished a phase III clinical trial of N-Rephasin Sal200 (ClinicalTrials.gov ID NCT03089697) (clinicalTrials.gov, retrieved June 21, 2022). Staphefekt SA.100, a synthetic phage endolysin designed for topical administration to the skin, has been used to treat patients with recurring and chronic S. aureus skin infections in three clinically successful cases. Based on cetomacrogol, this endolysin is marketed as a cream and a gel in pharmacies without a prescription and is registered as a medicinal product (class 1) in Europe [165]. Multidrug-resistant P. aeruginosa and A. baumannii can be effectively combated with the artilysin Art-175 (LYSANDO AG, Liechtenstein).
As seen from the presented data, endolysins have excellent prospects for replacing antibiotics in the fight against not only planktonic but also biofilm bacteria. However, further clinical trials are needed to evaluate their safety for human health.
The rapid development of AI contributes to progress in computer-aided drug discovery, which may allow overcoming of difficulties associated with traditional drug research and screening. The integration of AI-based computational methods has revolutionized lysin engineering, as it permits their rapid modification and enables functional predictions and structural optimization. Computational methods and AI are increasingly used to develop and optimize endolysins to enhance their activity against Gram-negative bacteria. Approaches such as machine learning, deep mutation scanning, and modular synthetic biology platforms (such as VersaTile) have accelerated the discovery of modified lysins with improved stability, specificity, and bactericidal activity. VersaTile is an innovative modular synthetic biology platform designed to accelerate the development and optimization of lysins for antibacterial application [173].
A comprehensive review of bioinformatic tools and databases related to the discovery, characterization, and development of lysins is presented in Bałdysz et al. [174], as are suggestions for the improvement of sequence annotation, structural modeling, and functional prediction to accelerate the development of lysin-based antimicrobials.
Some endolysins have been identified by bioinformatic analysis of the Pseudomonas and Klebsiella phage genomes in the NCBI database. Among them, the endolysin PlyKp104 was highly active, reducing the titers of K. pneumoniae, P. aeruginosa, and other Gram-negative bacteria by more than five logarithms without the need for additional modifications [4, 175]. By using a bioinformatic pipeline, 2628 new endolysins were identified from 183,298 uncultivable phage genomes, and many of these endolysins had a new domain architecture [176]. Among these endolysins, 66 new SAR endolysins were found [177].
Thus, at this stage of societal development, it is important to integrate computational and experimental approaches to better optimize endolysins for therapeutic use. The application of AI will strongly accelerate the discovery and optimization of new endolysins, thereby contributing to the design of new antimicrobial drugs.
The scientific community is searching for new and improved antimicrobial agents because of the increasing bacterial resistance, brought on by the inappropriate use of conventional antimicrobials. In this regard, bacteriophage endolysins show promise. The number of publications showing the potential of endolysins against both planktonic and biofilm bacteria has increased rapidly in the past 5 years. The active publication activity is due to the development of enzymatic drugs to treat infections caused by antibiotic-resistant Gram-positive and Gram-negative bacteria and associated biofilms. Thus far, there has been no proof of direct danger to humans, either in vitro or in vivo, and phage endolysins have shown exceptional bactericidal effectiveness and promise for the treatment and prevention of bacterial infections. Furthermore, endolysins have been genetically engineered to exert a stronger scavenging effect on biological membranes [158].
Because endolysins are synergistic with some biomolecules and antibiotics, their activity and stability can be increased and the cytotoxicity of coadministered antibiotics can be reduced. At present, a number of endolysins are successfully undergoing clinical trials. Although the inactivation of endolysins in serum remains an obstacle to their systemic use, this problem can be solved by protein engineering. However, local use of endolysins in medical institutions may enable direct application and use in agriculture or in food preservation [178, 179, 180]. Further research on endolysins will change the concept of therapy and prevention of infectious diseases including bacterial biofilms.
OIG – Conceptualization and Design, Project Administration, Software, Formal Analysis, Resources, Visualization, Writing–original draft, Writing–review & editing. OAK – Formal Analysis, Resources, Visualization. Both authors read and approved the final manuscript. Both authors contributed to editorial changes in the manuscript. Both authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This work was funded by the Ministry of Science and Higher Education of the Russian Federation as a state assignment for the Institute of Biochemistry and Physiology of Plants and Microorganisms – Subdivision of the Federal State Budgetary Research Institution Saratov Federal Scientific Centre of the Russian Academy of Sciences (IBPPM RAS).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.