


1 School of Nutritional Sciences and Wellness, University of Arizona, Tucson, AZ 85721, USA
Abstract
Bacteria, plants, and animals all express a conserved group of proteins known as the ZIP family, which mediate the transport of divalent cations, including zinc (Zn) and manganese (Mn). These proteins belong to a larger group of metal transporters named after the zinc-regulated transporter (Zrt) and iron-regulated transporter (Irt) proteins. In humans, the ZIP family comprises 14 members encoded by the SLC39A1–SLC39A14 genes. The roles of ZIP proteins in Zn metabolism have been extensively characterized, particularly in regulating cellular Zn uptake and distribution. First, this review introduces the ZIP family and the established functions of these transporters in Zn homeostasis, with a focus on cellular Zn transport, to provide a foundation for understanding the associated broader metal transport activities. We then place particular emphasis on those ZIP proteins with emerging roles in Mn homeostasis, an area that remains less well understood.
Keywords
- SLC39A
- zinc
- manganese
- micronutrient homeostasis
- ZIP
ZIP proteins transport divalent metals such as Zn and Mn [1]. In human erythrocyte-like cells and prostate cancer cells, ZIP1 is localized to the plasma membrane and functions as a Zn importer [2, 3]. Despite transcriptomic data indicating ubiquitous ZIP1 mRNA expression across mammalian tissues, reports describing ZIP1 protein subcellular localization in specific tissues and cell types are limited [4, 5]. Overexpression of ZIP1 in prostate cancer cells significantly increased Zn uptake, whereas overexpression of other ZIP family members did not, indicating that ZIP1 is primarily responsible for Zn uptake into the prostate [6]. In PC-3 cells, a prostate cancer cell line, site-directed mutagenesis of a histidine residue within the intracellular loop between transmembrane domains III and IV of ZIP1 resulted in decreased Zn uptake without affecting protein expression or change in localization, as confirmed by confocal images [3]. These findings suggest that this intracellular histidine motif is critical for ZIP1’s Zn transport activity. Additionally, in a study of 6-week-old rats fed a low-Zn diet, ZIP1 protein levels were decreased in liver tissue compared to controls, consistent with systemic Zn regulation of ZIP1 expression [7]. However, the mechanism controlling ZIP1 protein expression is not clear.
The role of ZIP2 in Zn transport was first inferred from its homology to known Zn transporters in yeast and plants [8]. After confirming that the transporter was responsible for Zn uptake in K562 cells, further experiments suggested that ZIP2 may also bind Mn, copper (Cu) and cadmium (Cd). In HEK293 cells expressing ZIP2, transport activity was shown to be voltage- and pH-dependent, but independent of ATP or ion gradients, indicating that ZIP2 mediates passive transport of Zn from the extracellular space into the cytoplasm [9]. Localization studies revealed that ZIP2 traffics to the plasma membrane in transfected K562 cells [8], and immunohistochemistry of human prostate tissue demonstrated ZIP2 expression on the apical membrane of glandular epithelial cells, suggesting a role in Zn uptake from prostate fluid [10]. ZIP2 protein is expressed at moderate levels across various human tissues, with higher levels observed in the stomach, prostate, and seminal vesicles. However, RNA expression data show low ZIP2 transcript levels in most tissues, except for relatively higher expression in the skin, prostate, and seminal vesicles [4, 5].
The initial functional characterization of ZIP3 involved transfecting HEK cells with mouse Zip1 (mZip1), Zip2 (mZip2), and Zip3 (mZip3) genes to express each transporter on the plasma membrane. This study found that ZIP3 mediated Zn uptake at levels comparable to ZIP1 and ZIP2, suggesting a conserved role in Zn import among these family members [11]. Given the conservation of these genes between mice and humans, these findings informed the understanding of human ZIP3 function. Further evidence supporting ZIP3’s role in Zn uptake comes from studies of pancreatic adenocarcinoma, where cancerous cells exhibited reduced Zn accumulation and markedly lower ZIP3 expression. In vitro knockdown of the Ras-responsive element-binding protein 1 (RREB-1) resulted in decreased ZIP3 expression and intracellular Zn levels, indicating that ZIP3-mediated Zn import is regulated by RREB-1 [8]. Additional reports confirm ZIP3 expression in pancreatic tissues, with localization to the basolateral membrane of epithelial cells [12].
ZIP4 plays a critical role in Zn transport, with clinical relevance demonstrated by mutations that cause acrodermatitis enteropathica, a genetic disorder resulting in severe Zn deficiency [13, 14]. Under Zn-deficient conditions, ZIP4 is trafficked to the apical membrane of intestinal epithelial cells to enhance Zn absorption [15]. Conversely, in cell-based studies, re-exposure to Zn in previously Zn-deficient cells triggered rapid endocytosis of ZIP4, indicating that Zn levels regulate ZIP4 activity post-translationally to prevent excess uptake [16]. Tissue expression studies have shown that ZIP4 is predominantly expressed in the intestine and kidney, with minimal expression in the liver, heart, and brain [13]. In mice, intestinal epithelial-specific knockout of ZIP4, including in Paneth cells, disrupted Paneth cell gene expression and impaired intestinal mucosal function, in addition to causing severe Zn deficiency. These findings suggest that the rapid deterioration observed in patients with ZIP4 mutations results not only from Zn deficiency but also from broader nutritional and intestinal dysfunction due to the loss of ZIP4-mediated transport [17].
ZIP5 is closely related to ZIP4 in both protein structure and expression profile, but differs notably in its regulation during Zn deficiency. When Zn is removed from the diet, ZIP5 appears to internalize and is no longer found on the cell surface. With normal Zn intake, ZIP5 is expressed on the basolateral side of the intestinal enterocytes and pancreatic acinar cells [18]. The physiological role of ZIP5 in Zn homeostasis was further elucidated in mouse models with global or tissue-specific ZIP5 deletion. Loss of intestinal ZIP5 led to increased Zn accumulation in the pancreas, suggesting that ZIP5 facilitates Zn clearance by transporting Zn from the bloodstream into enterocytes for excretion [19]. Like other ZIP family members, ZIP5 can transport multiple divalent metals. While early studies suggested that ZIP5 preferentially transports Zn over other metals [20], more recent work by Polesel et al. [21] using radiolabeled metal uptake assays in HEK cells revealed that ZIP5 transports Cu more efficiently than Zn, and elevated Cu levels can inhibit Zn uptake. Despite this finding, the role of ZIP5 in Cu homeostasis remains unclear, as no in vivo studies have examined Cu regulation in ZIP5-deficient animals.
ZIP6 is a Zn transporter closely tied to breast cancer. Gene expression is associated with increased epithelial-to-mesenchymal transition (EMT) and cancer metastasis via downregulation of E-cadherin [22, 23]. Mechanistically, ZIP6 mediates Zn influx into cancer cells, which leads to the phosphorylation of enzyme GSK-3Beta and in turn keeps transcription factor SNAI1 in the nucleus to repress E-Cadherin expression and promote EMT [22]. Expression profiling across tissues shows that ZIP6 is most highly expressed in the breast, prostate, placenta, and kidney, with minimal expression in the intestine and heart [24]. Functional studies in Chinese hamster ovary (CHO) cells overexpressing ZIP6 demonstrated increased Zn uptake, consistent with its role as a plasma membrane Zn importer [24]. In the nervous system, ZIP6 is localized to neuronal membranes in rat pups and to the apical surface of choroid plexus epithelial cells, suggesting a role in Zn uptake into neurons and across the blood–cerebrospinal fluid barrier via the choroid plexus [25].
Unlike other members of the ZIP family, ZIP7 is not localized to the plasma membrane but instead functions within intracellular compartments to regulate Zn distribution. Initial characterization of ZIP7 (also referred to as HKE4) in mammalian cells revealed its localization to the endoplasmic reticulum (ER) and ubiquitous gene expression, with slightly elevated levels in the placenta, liver, and pituitary gland [26]. Subsequent studies confirmed ZIP7’s intracellular localization and its role in mobilizing Zn from internal stores. Specifically, ZIP7 was found on the Golgi apparatus membrane, and its protein expression was downregulated in response to Zn treatment, suggesting feedback regulation by Zn level [27]. In human MG-63 osteosarcoma cells, ZIP7 deletion led to increased Zn retention in the ER, reduced cytosolic Zn, induction of ER stress, and impaired cell proliferation [28]. These findings establish ZIP7 as a key intracellular Zn transporter, crucial for maintaining cytosolic Zn availability and cellular homeostasis.
ZIP8 was first identified in 2002 as a multi-pass membrane zinc transporter, initially named BIGM103, in cultured monocytes exposed to inflammatory stimuli [29]. Its broader significance emerged with the discovery of a gene locus on mouse chromosome 3, known as Cdm, which was linked to increased susceptibility to Cd-induced testicular necrosis. Mutations in this gene led to decreased Cd uptake, providing a protective effect in mutants [30]. Within this region, the Zip8 gene was identified as the key mediator of Cd uptake [31]. In fetal fibroblast cells, manganese (Mn) was the most effective competitor for Cd transport via ZIP8, followed by mercury (Hg) and lead (Pb) [32]. Further studies confirmed that ZIP8 also facilitates the uptake of essential metals. In HEK293 cells overexpressing human ZIP8 (hZIP8), iron (Fe) uptake increased by 200% and Zn uptake by 40% upon metal exposure, supporting its role in transporting multiple divalent cations [33]. Additionally, ZIP8 was found to be expressed primarily on the plasma membrane of HEK293 cells, with some expression on endosomes [33]. ZIP8 is now recognized as a key regulator of Mn homeostasis, a topic discussed in detail later in this review. Briefly, several studies highlight ZIP8’s high affinity for Mn. In fetal fibroblast cells, Mn outcompeted Cd in transport assays, indicating a stronger affinity [32]. In Xenopus oocytes overexpressing ZIP8, Mn inhibited Zn uptake, supporting the view that ZIP8 preferentially transports Mn [34]. Moreover, siRNA-mediated knockdown of ZIP8 in A549 cells—a model of type II alveolar epithelial cells—led to a significant reduction in Mn uptake, further confirming its central role in Mn transport [35].
ZIP9 was first shown to be expressed in the trans-Golgi network, with minimal presence on the plasma membrane. In conditions of either Zn abundance or scarcity, ZIP9 expression level or localization did not appear to change [36]. However, a later study revealed the expression of ZIP9 on the cell membrane, where it functions as a membrane androgen receptor by binding testosterone [37]. In prostate cancer cells, ZIP9 was shown to associate with G-proteins on intracellular membranes, and testosterone treatment enhanced Zn import into the cytoplasm via ZIP9. This increase in intracellular Zn triggered G-protein-mediated signaling, upregulation of pro-apoptotic genes, and ultimately, programmed cell death [38, 39, 40]. Notably, ZIP9 is the only known ZIP family member that binds androgens, suggesting potential sex-dependent differences in its activity. A 2021 study demonstrated that ZIP9 expression influenced melanoma cell proliferation in the presence of testosterone, with reduced growth observed in females and ZIP9-deficient cells [37]. Outside of cancer models, ZIP9 is broadly expressed in normal tissues, with the highest levels found in the skin, bone marrow, liver, brain, and reproductive organs [4, 5].
ZIP10 has a high affinity for Cd but generally acts as a Zn transporter. It appears to localize to the nucleoplasm, cytosol, and plasma membrane, and ZIP10 mRNA increases as extracellular Zn increases [4, 5, 41]. Gene expression of ZIP10 is found throughout the major organs responsible for Zn storage and metabolism, with the highest expression in the small intestine [41]. Though ZIP10 has largely been studied as a Zn transporter, its affinity for Cd suggests that it would readily transport Cd, even in the presence of Zn. A study in renal brush border cells confirmed that the presence of Cd reduced Zn uptake [41]. Similar to ZIP6, ZIP10 has been implicated in breast cancer progression. A 2007 study identified ZIP10 as a marker of metastatic breast cancer, showing that ZIP10 knockdown significantly reduced metastasis. Likewise, Zn chelation also impaired metastatic potential, suggesting that ZIP10-mediated Zn import supports cancer cell migration during metastasis [42].
ZIP11 is highly expressed in the brain, intestinal tract, and testes, and has also been found to be upregulated in kidney cancer [43]. It localizes to the plasma membrane, nucleus, and Golgi apparatus, where it functions as a Zn importer to increase cytosolic Zn availability. In HEK cells overexpressing mouse ZIP11 (mZIP11), intracellular Zn levels were significantly elevated compared to cells transfected with an empty vector, confirming its role in Zn uptake. Notably, mZIP11 overexpression also led to increased intracellular Cu, suggesting broader metal transport capabilities [43]. In mice subjected to Zn restriction, ZIP11 expression was upregulated, likely as a compensatory mechanism to enhance Zn uptake under low-Zn conditions [43, 44]. More recently, a 2024 study linked ZIP11 to Mn homeostasis [45], a topic that will be explored in the next section.
Early studies identified high ZIP12 expression in retinal pigment epithelial cells, the choroid plexus, and glial cells of the brain, suggesting a role in neural and ocular Zn transport [46]. Subsequent research showed that ZIP12 mRNA and protein levels are upregulated in endothelial, interstitial, and vascular smooth muscle cells following hypoxic injury in rats [47]. In cultured human vascular smooth muscle cells, hypoxia induced significant Zn accumulation, a response that was markedly reduced when ZIP12 expression was silenced by siRNA. While the impact of reduced Zn uptake on hypoxia recovery remains unclear, these findings suggest a role for ZIP12 in cellular adaptation to low oxygen conditions. ZIP12 has also been implicated in neuronal development. In mouse neurons, ZIP12 localizes to the plasma membrane, and Zn chelation increases its presence at the membrane and in the cytosol, with reduced localization in the perinuclear region [48]. In Neuro-2a (N2a) cells, a mouse neuroblastoma line that differentiates into neuron-like cells, ZIP12 expression was strongly upregulated during differentiation, accompanied by increased Zn accumulation. Knockdown of ZIP12 impaired this Zn uptake and led to a reduction in both the number and length of neurites, indicating a critical role for ZIP12 in neuronal morphology and development [48].
Mutations in ZIP13 have been linked to a subtype of Ehlers-Danlos syndrome (EDS), a connective tissue disorder characterized by joint hypermobility and impaired collagen formation. In 2008, researchers predicted—based on sequence homology with ZIP7—that ZIP13 localizes to the endoplasmic reticulum (ER) and plasma membrane, where it regulates Zn transport into the cytosol from both the ER and the extracellular space [49]. EDS symptoms in ZIP13-deficient individuals are thought to arise from disrupted collagen hydroxylation, as many collagen-modifying enzymes require Fe. It has been proposed that altered Zn homeostasis interferes with Fe availability in key organelles. Supporting this, a recent study showed that loss of ZIP13 causes Fe deficiency in the ER, Golgi, and mitochondria, accompanied by increased Fe accumulation in the cytosol [50], suggesting that ZIP13 plays a role in coordinating intracellular Zn and Fe distribution essential for connective tissue integrity.
ZIP14 is ubiquitously expressed throughout the body, with the highest protein expression in the liver, intestine, kidney, and female reproductive tissue [5]. Initially characterized as a divalent metal transporter involved in Fe uptake, ZIP14 was shown to localize to the plasma membrane and mediate the import of both Zn and Fe in early studies [51]. In mouse hepatocytes, ZIP14 knockdown led to decreased intracellular Zn and Fe levels, supporting its role as a key metal importer in the liver. This finding also suggests that ZIP14 may help regulate Zn clearance, as Zn would first need to be imported by hepatocytes before being excreted into the bile. However, manganese (Mn) transport was not assessed in this early work. Subsequent studies expanded ZIP14’s metal transport profile. In proximal kidney tubule cells, siRNA-mediated knockdown of ZIP14 significantly reduced both Cd and Mn uptake, confirming its function as a Mn transporter and its importance in renal metal handling [52]. Later animal studies and human genetic analyses further established ZIP14 as a critical regulator of Mn homeostasis—an aspect that will be discussed in detail in the following section.
While the majority of ZIP family proteins have been investigated primarily for their roles in Zn metabolism, two members—ZIP8 and ZIP14—stand out for their critical involvement in Mn homeostasis. These two transporters have been shown to mediate the cellular uptake of Mn and are essential for maintaining Mn balance in various tissues. A recent review has compared the structures of these two transporters, finding biochemical similarities that allow ZIP8 and ZIP14 to preferentially transport Mn in addition to Zn [53]. Unlike most other ZIP proteins whose functions are largely limited to Zn transport, ZIP8 and ZIP14 exhibit broader substrate specificity and play key roles in regulating Mn levels under both physiological and pathological conditions. Despite the similar structures, these proteins exhibit differences in localization and tissue-specific expression. Their unique ability to transport Mn highlights the functional diversity within the ZIP family and underscores the need to further explore their roles beyond Zn metabolism.
As an essential trace metal, Mn must be tightly regulated to maintain a balance within the body. Disruptions in Mn homeostasis, such as mutations in ZIP8 and ZIP14, can result in severe Mn deficiency or overload, respectively, both of which lead to human diseases characterized by significant neurological and systemic symptoms. Normal Mn concentrations in human blood range from 7–12 µg/L, and 0.6–4.3 µg/L in serum [54, 55]—substantially lower than Zn, which is typically present at 800–1200 µg/L in serum [56].
Mn is widely distributed in the environment and plays critical roles in both plant and animal biology. It is found abundantly in soil and accumulates in many plants and shellfish. Dietary sources rich in Mn include grains, rice, nuts, leafy greens, mussels, and oysters [57, 58]. In areas with heavy soil contamination of Mn, higher Mn levels can also be detected in drinking water. Mn-dependent enzymes are vital for numerous biological processes, including immune function, energy metabolism, digestion, blood sugar regulation, and protection against oxidative stress [58]. One of the most well-characterized Mn-dependent enzymes is manganese superoxide dismutase (MnSOD), which catalyzes the conversion of harmful superoxide radicals into hydrogen peroxide [59, 60]. The activity of MnSOD is directly influenced by Mn availability; for example, the yeast homolog of MnSOD shows increased enzymatic activity when exposed to excess Mn [61]. Conversely, Mn deficiency may impair antioxidant defense by reducing MnSOD activity. Other Mn-dependent enzymes include glutamine synthetase, which is essential for neurotransmitter production and normal brain function [62]; arginase, which facilitates the conversion of L-arginine into urea and ornithine, promoting polyamine synthesis and cell proliferation [63]; and glycosyltransferases, which are necessary for proteoglycan synthesis during bone formation [64].
Because both Mn deficiency and overload can have serious physiological consequences—especially for the brain—tight regulation of Mn uptake and distribution is critical. ZIP8 and ZIP14 are key transporters responsible for maintaining systemic Mn balance. In addition, a recent study has implicated ZIP11 in Mn homeostasis for the first time, and its role will also be briefly discussed here.
ZIP8 deficiency was first described in a cohort of six children from an ethno-religious community with a high incidence of consanguineous marriages. All affected individuals carried the same homozygous mutation in the ZIP8 gene, resulting in a non-functional protein. Although ZIP8 was expressed at normal levels and correctly localized, it lacked divalent metal transport activity. The children exhibited severe to profound intellectual and physical disabilities, cerebellar atrophy, strabismus, and recurrent infections. While blood levels of Mn and Zn were low in some individuals and within normal range in others, most showed markedly elevated urinary excretion of both metals, indicating excessive clearance and impaired reabsorption or retention [65]. A separate clinical case reported a child with ZIP8 mutation presenting with skull malformations, limb shortening, and strabismus. Biochemical testing revealed abnormal serum transferrin glycosylation, consistent with a congenital disorder of glycosylation (CDG). Notably, Mn levels in the blood were undetectable, whereas Fe and Zn remained within normal ranges. This case further established the role of ZIP8 in Mn homeostasis and linked its deficiency to defects in glycosylation pathways dependent on Mn availability [66].
Animal studies further underscore the essential role of ZIP8 in Mn homeostasis. In the liver, ZIP8 is localized to the canalicular membrane, where it mediates Mn uptake from bile, thus regulating hepatic Mn storage (Fig. 1). In mice with liver-specific Zip8 knockout, Mn levels in both the liver and kidney were significantly reduced. Restoration of ZIP8 expression via adenovirus-associated vector delivery of the wild-type gene successfully normalized Mn levels in these tissues, confirming ZIP8’s role in hepatic Mn uptake [67]. ZIP8 also plays a critical role in intestinal Mn absorption (Fig. 1). A recent study using an intestine-specific Zip8 knockout mouse model examined whether systemic Mn homeostasis could be maintained when ZIP8 was absent only in the gut. Despite normal ZIP8 expression in other tissues, these mice exhibited reduced Mn absorption and significantly lower blood Mn levels compared to controls. Supporting these findings, intestinal organoid cultures revealed that ZIP8 localizes to the apical membrane of enterocytes, consistent with a role in Mn uptake from the intestinal lumen [68].
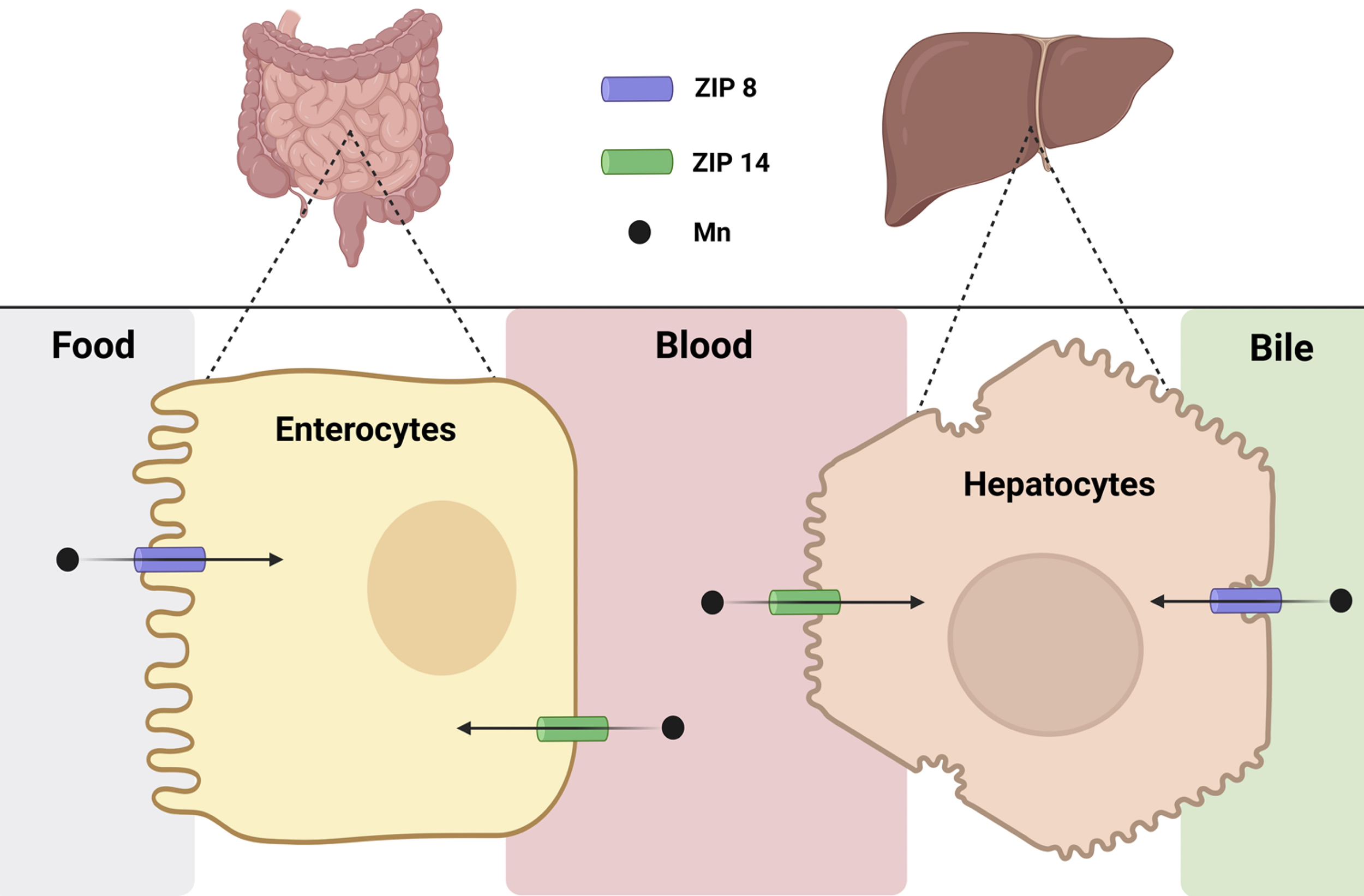
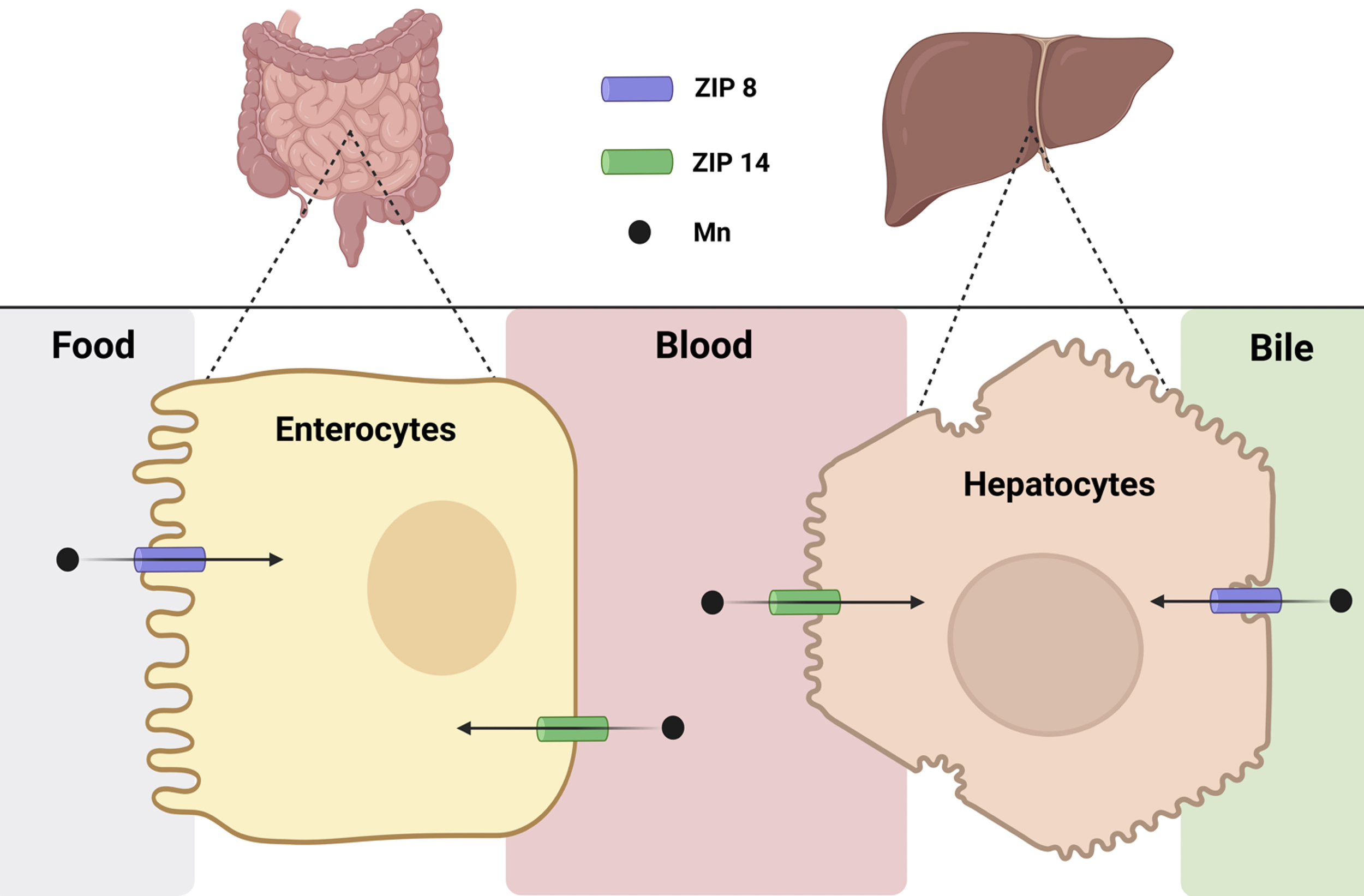 Fig. 1.
Fig. 1.
ZIP8 and ZIP14 control systemic Mn homeostasis. ZIP8 and ZIP14 contribute to systemic Mn homeostasis by mediating Mn transport at key sites of absorption and clearance, namely, the intestine and the liver. In the intestine, ZIP8 imports Mn from the intestinal lumen into enterocytes, where it is eventually absorbed into the bloodstream by an unknown exporter. ZIP14 limits Mn absorption by importing the freshly absorbed Mn from the blood back into enterocytes, thereby keeping blood Mn levels low. Within the liver, ZIP14 imports Mn from the blood into the hepatocytes, where it can be exported by another transporter (ZnT10) into the bile for later excretion. ZIP8 expressed on the bile-facing apical side of the hepatocyte reclaims Mn from the bile and imports the metal back into the hepatocytes. This allows the liver to maintain adequate Mn stores. Together, ZIP8 and ZIP14 play complementary and tissue-specific roles in maintaining Mn balance by regulating its absorption, distribution, and excretion, thereby protecting against both Mn deficiency and toxicity. This figure is created in BioRender. Favre, D. (2025) https://BioRender.com/s8u7hjq.
ZIP8 likely plays important roles in Mn metabolism at additional physiological barriers, including the lung alveoli [35, 69] and brain endothelium [70, 71], where it is highly expressed. The essential nature of ZIP8 is further demonstrated in mouse models, where whole-body Zip8 deletion in utero resulted in embryonic or perinatal lethality, with all homozygous mutant pups dying before or shortly after birth [72]. While ZIP8 is clearly required for Mn homeostasis, evidence suggests it may also regulate the expression of genes or proteins unrelated to metal transport, indicating broader functional roles. Additionally, increased Mn intake downregulates the level of hepatic ZIP8 expression in a study including juvenile mice [69]. As research into ZIP8 advances, its relevance to human health continues to grow. Recent studies have linked ZIP8 mutations to a range of diseases, including Crohn’s disease and inflammatory bowel disease [73, 74], Schizophrenia [75], and idiopathic scoliosis [76], highlighting the expanding scope of ZIP8 beyond Mn metabolism.
ZIP14 was first identified in 2004 as a plasma membrane transporter that increases cytosolic Zn concentrations, based on studies in Chinese hamster ovary (CHO) cells. Notably, stronger ZIP14 staining was observed near cell–cell junctions, suggesting a possible role in intercellular signaling or junctional integrity [77]. Although initially studied for its involvement in Zn and Fe transport [51, 78], ZIP14 is now more closely associated with Mn homeostasis. This was confirmed by reports of ZIP14 mutations in children, which caused severe disease. ZIP14 dysfunction causes severe Mn accumulation in the blood and brain, which leads to cognitive and motor symptoms that cannot be cured, even when high blood Mn is alleviated with chelators [79]. ZIP14 protein is most abundant in the liver, followed by the small intestine [80]. In addition to these peripheral sites, ZIP14 is also enriched in the choroid plexus epithelium [70, 71], where it may contribute to Mn regulation in the cerebrospinal fluid and central nervous system.
ZIP14 is a critical regulator of Mn in the body, and the excretion of Mn from the blood can prevent Mn accumulation in the brain, thus avoiding the severe neurological symptoms associated with whole-body Mn overload. ZIP14 can also be regulated by Mn concentration—in a study of liver cells exposed to excess Mn concentrations, it was found that higher Mn availability led to decreased ZIP14 expression [81]. Localization studies in mouse tissue and human cells have revealed that ZIP14 protein is specifically located on the basolateral side of the intestinal epithelium [82, 83] and enterocytes [84]. In the liver, ZIP14 is expressed on the basolateral side of polarized WIF-B hepatocytes [85] and Hepa-RG cells [81]. Early radiotracer studies indicated that Mn was excreted through the liver and intestines [86] and clinical studies showed that patients with cirrhosis would have impaired Mn clearance which resulted in neurotoxicity [87, 88]. Mouse studies using organ-specific deletions of ZIP14 have revealed that a lack of ZIP14 in the liver alone does not cause an accumulation of brain or blood Mn [84, 89], but the combined loss of both liver and intestinal ZIP14 causes an increased Mn load in the body [90]. Additionally, deletion of ZIP14 in the intestine alone causes blood and brain Mn accumulation and leads to neuroinflammation and motor dysfunction [91]. It appears that intestinal ZIP14 can compensate for the loss of hepatic ZIP14, therefore allowing normal Mn excretion and preventing the onset of brain Mn hyperaccumulation and associated symptoms [90, 92].
To summarize, ZIP14 is present in the intestine, liver, brain, choroid plexus, and kidneys. Mn homeostasis is largely controlled by intestinal ZIP14 which is expressed on the basolateral side of intestinal epithelial cells. Here, ZIP14 is an importer that removes Mn from blood to eventually be excreted through the intestinal lumen (Fig. 1). In the liver, ZIP14 is expressed in hepatocytes and localizes to the basolateral side of the cell where it takes up Mn from blood and aids in the excretion of Mn via the bile duct (Fig. 1).
Although less extensively studied than ZIP8 and ZIP14, ZIP11 has recently emerged as a potential player in Mn homeostasis. While its role in Mn regulation is not yet fully defined, growing evidence suggests that ZIP11 may contribute to Mn transport at the subcellular level, particularly in maintaining intracellular Mn distribution.
In mice, ZIP11 is highly expressed in the stomach and intestinal mucosa; however, it is not found at the plasma membrane. Using established markers for cell surface and nuclear localization, one study demonstrated that ZIP11 colocalizes with a nuclear marker, suggesting that the transporter is enriched either within or on the nuclear membrane [44]. This subcellular positioning hints at a possible role in regulating Mn (and potentially other metal) availability within the nucleus, although the precise function remains to be determined.
Further interest in ZIP11 arose from a genome-wide association study (GWAS) conducted by Zeng et al. [93] in a population of Chinese centenarians. The study identified a single nucleotide polymorphism (SNP) in the SLC39A11 gene that was significantly enriched in male participants, leading the authors to propose a potential link between ZIP11 and increased longevity [93]. This discovery prompted a focused investigation into ZIP11 using multiple experimental models, including cell culture, zebrafish, and mouse models [45].
In zebrafish, zip11 mutant males displayed structural abnormalities in muscle fibers and reduced swimming speed, whereas females did not exhibit similar motor deficits. However, following injury, both male and female mutants showed impaired muscle regeneration, with males being more severely affected. Additionally, both sexes of zip11 mutants experienced shortened lifespans, suggesting that ZIP11 may play a sex-dependent role in tissue maintenance and longevity. In embryonic zebrafish exposed to solutions containing Fe, Mn, or Zn, slc39a11 expression was significantly downregulated by Mn exposure and modestly upregulated by Zn, indicating that ZIP11 is responsive to metal availability and particularly sensitive to Mn.
In mouse models, tissue-specific Zip11 knockouts were generated to assess the role of ZIP11 in systemic Mn homeostasis. Liver-specific Zip11 knockout mice exhibited decreased serum Mn levels but no significant changes in Mn content within the liver or other major organs, including the intestine, brain, spleen, or kidney. In contrast, intestinal-specific Zip11 knockout mice displayed elevated Mn levels in the serum, liver, and small intestine—an effect observed only in female mice. These findings suggest a tissue- and sex-specific role for ZIP11 in regulating Mn absorption and distribution, particularly through the intestinal route.
Collectively, these studies point to a previously unrecognized role for ZIP11 in Mn homeostasis. The gene’s sensitivity to Mn exposure, impact on lifespan, and sex-specific effects on metal regulation and tissue function suggest that ZIP11 may contribute to protecting against Mn toxicity. Furthermore, its association with human longevity highlights the broader physiological significance of this transporter, opening new avenues for research into Mn metabolism and age-related health outcomes.
Additional evidence from human fibroblast cell studies supports a functional role for ZIP11 in protecting cells from Mn-induced toxicity. In this model, ZIP11 knockdown followed by Mn exposure led to premature cellular senescence, an effect that was not observed when cells were treated with Zn. This selective response further underscores the transporter’s relevance in Mn regulation [45].
Several transporters in the ZIP family are noted for their abilities to control Zn and Mn levels intracellularly and contribute to homeostasis of micronutrients throughout the body. Studies of the primarily-Zn transporters are critical to understand disorders of Zn deficiency. Additionally, ZIP transporters could be used as markers for some cancers, as their upregulation is associated with increased Zn and metastasis. Such is the case for ZIP10, which is upregulated in metastatic breast cancer, but inhibition of the transporter reduces mitotic activity. The recommended dietary intake of Mn is just a fraction compared to that of Zn, but this metal must be tightly controlled to maintain homeostasis. The study of ZIP transporters has allowed us to understand the mechanism by which Mn is absorbed, stored, and excreted in the body. ZIP8 and ZIP14 are necessary for Mn homeostasis by importing Mn either from the bile or as it is digested in food (ZIP8), or importing Mn from the blood to reduce the concentration of Mn circulating in the body (ZIP14).
SM and NZ designed the research. SM and DF made the figure. SM, DF, and NZ wrote the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Thanks to all the peer reviewers for their opinions and suggestions.
This work was supported by the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK) and the Office of Dietary Supplements (ODS) of the National Institutes of Health (NIH) (R01DK123113).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.