






 , Vitaly A. Khanadeev 1, Lev A. Dykman 1
, Vitaly A. Khanadeev 1, Lev A. Dykman 11 Institute of Biochemistry and Physiology of Plants and Microorganisms – Subdivision of the Federal State Budgetary Research Institution Saratov Federal Scientific Centre of the Russian Academy of Sciences (IBPPM RAS), 410049 Saratov, Russia
Abstract
The need for rapid and cheap synthesis of large numbers of chemical compounds has contributed to the emergence of combinatorial chemistry (simultaneous synthesis of different compounds, in contrast to traditional synthesis, in which each substance is produced individually). Combinatorial library methods were initially applied only to peptides and oligonucleotides. By now, the scope of these libraries has expanded considerably to include proteins, synthetic oligomers, small molecules, and oligosaccharides. The enormous variety of antibodies (Abs) makes it possible to detect clones able to interact highly specifically with almost any natural or synthetic antigen (Ag). Phage Abs are an excellent alternative to mono- and polyclonal Abs, because they are highly stable, have no disulfide bonds, and are much cheaper to make. Monitoring of various substances, including proteins, in a living organism is much in demand. Despite the vast amount of literature available on Ab phage display, the use of phage display to determine diagnostically important Ags has not been sufficiently covered. Many studies have confirmed that unlike other types of Abs, phage Abs ensure highly sensitive Ag detection. Therefore, this review focuses on the use of phage display to prepare Abs specific to diagnostically important Ags (allergens, disease and cancer biomarkers, toxins) and on their application in analytical systems, including biosensors. The use of phage Abs in Ag diagnostics is compared with the use of classical Abs, and the prospects are shown for the use of phage Abs as biosensor sensing elements. This review analyzes the recent advances in the detection of diagnostically important Ags by using phage display–based biosensors. Systematic information is presented about allergens, disease and cancer biomarkers, and toxins detected by using phage Abs. Phage display Abs for sensor-based Ag detection are presented as an affordable alternative to classic tests.
Keywords
- phage antibodies
- biosensors
- antigen
- detection
The need for rapid and cheap synthesis of large numbers of chemical compounds has contributed to the emergence of combinatorial chemistry (simultaneous synthesis of different compounds, in contrast to traditional synthesis, in which each substance is produced individually). Combinatorial library methods were initially applied only to peptides and oligonucleotides. By now, the scope of these libraries has expanded considerably to include proteins, synthetic oligomers, small molecules, and oligosaccharides. Although types of libraries differ, the processes are similar and include three main steps:
-Preparing a library.
-Screening the library’s components.
-Determining the chemical composition of the active components.
The enormous variety of antibodies (Abs) makes it possible to detect clones able to interact highly specifically with almost any natural or synthetic antigen (Ag). Modern medicine needs Abs with specific properties, and a common method to prepare them is to screen combinatorial Ab libraries.
Phage display is a molecular technique that allows expression of foreign proteins on the surface of phage particles. The phages transformed in this way become capable not only of expressing and carrying foreign proteins but also of further replicating foreign DNA. Phage display greatly expands the number of nucleotide sequences that can be transformed into populations of different peptides and proteins, allowing selection of those with properties of research interest. In the earliest experiments, only one peptide was expressed on the surface of a phage particle against the background of the entire population of wild-type phage proteins (peptide libraries). Currently, the scope and capabilities of the method have increased considerably. Natural and synthetic peptides, proteins and protein domains, and synthetic Abs all can be expressed on the phage particle surface.
The strong points of phage display are as follows: (1) the possibility to select clones producing mini-Abs in vitro and bypass animal immunization, (2) the possibility of producing Abs to auto-Ags, toxins, and weakly immunogenic compounds, (3) the absence of need to use laboratory animals and long-term cultures of eukaryotic cells, (4) the shortened time needed to generate individual clones (10–14 days, as compared with several months in the case of hybridoma technology), (5) the relative simplicity of preparation of mini-Abs and their low cost, and (6) the possibility of designing hybrid Ab molecules with marker proteins.
Broadly speaking, both phage Abs and those produced by hybridoma technology can be used in the same range of applications in immunoassays and biosensorics. However, phage display has some advantages over hybridoma technology. One limitation of hybridoma technology is that hybridoma Abs must be used at parameters close to physiological, which may be inconvenient for some test objects. Phage display technology allows isolation of Abs with sufficient binding capacity under any physiological conditions.
Phage Abs as biological macromolecules are an excellent alternative to mono- and polyclonal Abs, because they are highly stable, have no disulfide bonds, and are much cheaper to make. They have established themselves as specific bioreceptors in the detection of bacteria [1, 2, 3, 4, 5], bacterial spores [6], and viruses [1, 7, 8, 9].
Monitoring of various substances, including proteins, in a living organism is much in demand. Despite the vast amount of literature available on Ab phage display [10, 11, 12], the use of phage display to determine diagnostically important Ags has not been sufficiently covered. Therefore, this review focuses on the use of phage display to prepare Abs specific to diagnostically important Ags (allergens, disease and cancer biomarkers, toxins) and on their application in analytical systems, including biosensors.
The property of affinity presents unique possibilities for the study of Ag–Ab interactions. An affinity interaction is the ability of two or more molecules to form a chemical bond. The Ag–Ab reaction results in the specific binding of Ag molecules to Abs, giving rise to immune complexes. In vitro this reaction, as a rule, proceeds in two stages, which differ in mechanism and speed.
The first, specific stage proceeds rapidly and without visible effects even at low temperature. It obeys the general laws of physical chemistry and thermodynamics. At this stage, variable Ab sites interact reversibly and specifically with antigenic determinants.
The second, nonspecific stage proceeds slowly. It can be observed visually, for example, when immune complexes precipitate during agglutination.
The second stage may be absent if Abs interact with haptens (nonimmunogenic low-molecular-weight substances). The rate and possibility of this stage depends on a number of conditions, including temperature and the presence of salts [13].
The Ab site that binds to an Ag is called a paratope, and the Ag site that is recognized by an Ab is called an epitope. The strength of the Ag–Ab binding, also called affinity, can be measured by determining the association constant [14]. The paratope and epitope are highly complementary.
Complementarity-determining regions (CDRs, hypervariable regions of the Ab molecule) are important in the Ag–Ab interaction. The three-dimensional structure of Abs, consisting of six CDR loops, is involved in the recognition of its complementary epitope on the Ag surface [15].
The amino acid residues that make up the CDRs and interact with the epitope are called specificity-determining regions. The shape of the contacting surface of Abs depends largely on the Ag size. With high-molecular-weight Ags, it is almost flat, whereas with low-molecular-weight ones, it is more concave. In addition to amino acid residues that are in direct contact with the epitope, the key part in Ag recognition is played by those amino acid residues that have a structural function and ensure the necessary conformation of CDR loops [16]. The Ag–Ab binding is reversible and is brought about by hydrogen bonds, electrostatic and hydrophobic interactions, and van der Waals forces. Although these bonds are weaker than covalent ones, together they ensure a high-affinity interaction between epitope and paratope [17].
Because Abs can bind specifically to complementary Ag sites, they are commonly used in biotechnology, including in Ag detection [18, 19]. Successful use of Abs depends not only on their specificity but also on the method of their preparation.
The simplest and easiest method to develop Abs specific for a particular Ag is animal immunization, which produces high-affinity, high-selectivity Abs. These Abs or their genes can then be isolated, sequenced, and modified to impart the desired properties to them. The serum of an immunized animal contains polyclonal Abs specific for the introduced Ag, which differ in their structure, epitope specificity, and affinity. The weak point of polyclonal Abs is that they are specific for many Ag determinants [15].
A more complex method is hybridoma technology (once obtained and characterized, hybridomas represent an unlimited source of monoclonal Abs) [20]. Although this method affords high-quality Abs, hybridomas grow relatively slowly, do not reach high densities, and require complex and expensive media.
An alternative way to obtain specific Abs is provided by display methods, which are based on combinatorial library synthesis [the variable regions of light and heavy immunoglobulin chains are connected at random and are presented on the surface of a filamentous bacteriophage (M13)] [21, 22, 23, 24]. This method displays foreign peptides or proteins on the phage particle surface as part of a chimeric coat protein [24, 25]. Matching of genotype to phenotype requires screening and amplification of protein libraries in in vitro selection, a process similar to natural selection [26, 27, 28].
Generally, combinatorial phage libraries are made as follows (Fig. 1) [29]:
1. The scFv, Fab, or other genes are cloned from the mRNA isolated from donor immune or intact B cells (human, mouse, rabbit, chicken, pig, dog, monkey, sheep, cow, etc.).
2. The gene coding for the capsid protein (normally pIII) is introduced into a single translation frame of the phagemid containing these genes.
3. After the phagemid genes are expressed and virions are formed, the resultant phagemids are used to infect Escherichia coli. Foreign Ab fragments will be displayed as components of the virions’ capsid proteins [29].
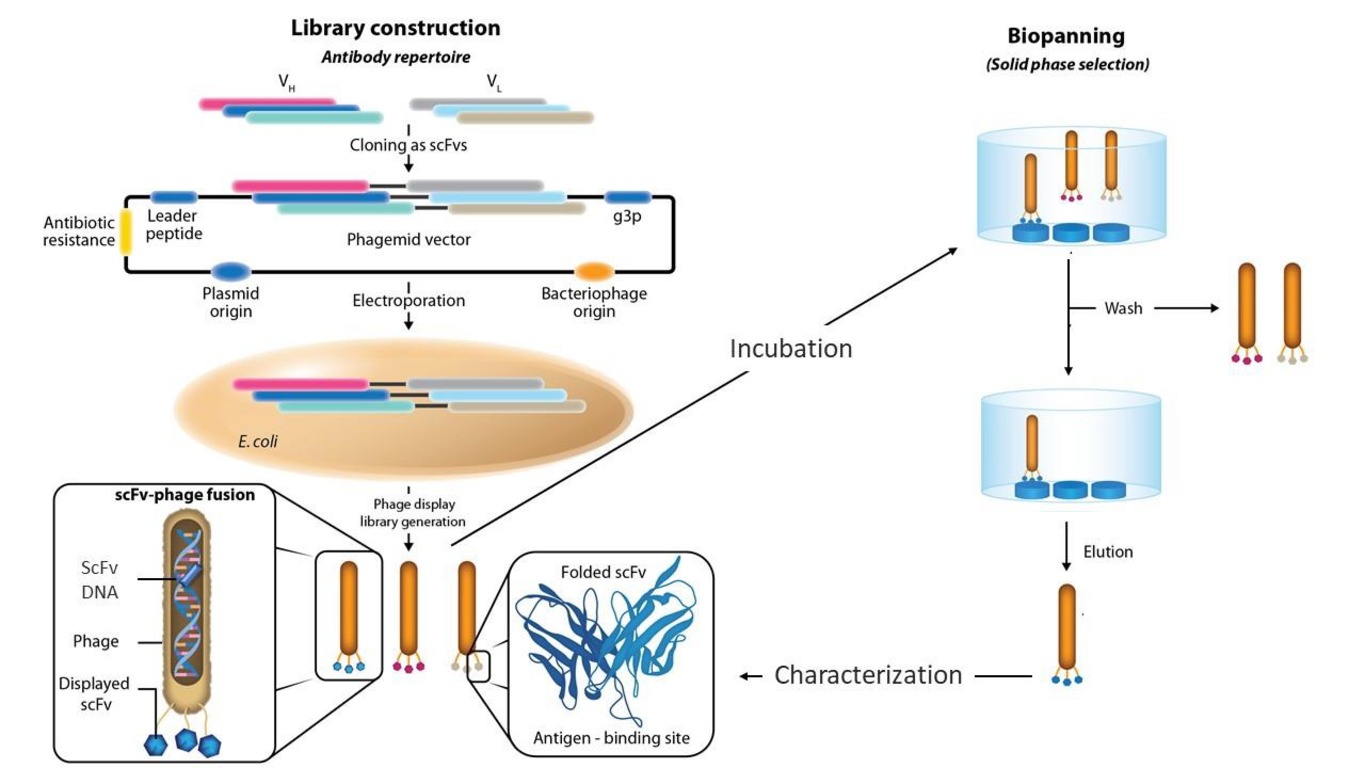
This generates a population of bacteriophages, each of which has a specific Ag-binding domain on its surface. Several rounds of selection are done to increase phage Ab sensitivity.
Filamentous phages are used in phage display because of their unique shape, the ways in which the bacterial cell secretes them, and the potential for significant phage genome modification without compromising viability. On the basis of these phages, gene fragment libraries have been made such as those of Mycobacterium tuberculosis (causative agent of tuberculosis), Plasmodium falciparum (causative agent of malaria), Salmonella typhimurium (causative agent of salmonellosis), Streptococcus pneumoniae (causative agent of pneumonia), human cytomegalovirus, and human immunodeficiency virus [30].
Although the first phage display was based on filamentous bacteriophages (phage
M13), the use of such phages has its limitations. For example, some peptides and
cytoplasmic proteins, owing to their structure, cannot pass through the secretion
systems of a bacterial cell and, consequently, cannot be displayed on the surface
of filamentous phages. In this case, systems based on lytic phages have clear
advantages, because the phage progeny is formed in the cytoplasm of the bacterial
cell and is released by its lysis. The display systems based on three lytic
phages, viz.
-They enable the display of larger proteins, because they can carry larger foreign DNA.
-They increase the likelihood of successfully displaying soluble proteins, because they are fully assembled in the cytoplasm and exit the host cell through lysis, negating the need for the displayed soluble proteins to cross the cell membrane.
-The dilemma of which phages (virulent or temperate) are better to use is raised
by the variety of phage display tools available. Using T7 and
Nonetheless, filamentous phage-based systems are simpler and cheaper to use and
can be adapted to different tasks, as compared with lysogenic phage-based ones.
They do not require specialized packaging systems to produce phage progeny.
Likewise, they do not require homologous or site-specific recombination systems
needed for the introduction of foreign genetic material, which are characteristic
of the
A crucial step after library construction or selection from the available libraries is the enrichment of the initial repertoire of library Abs with molecules specifically directed to the target Ag (Fig. 2) [34, 35].
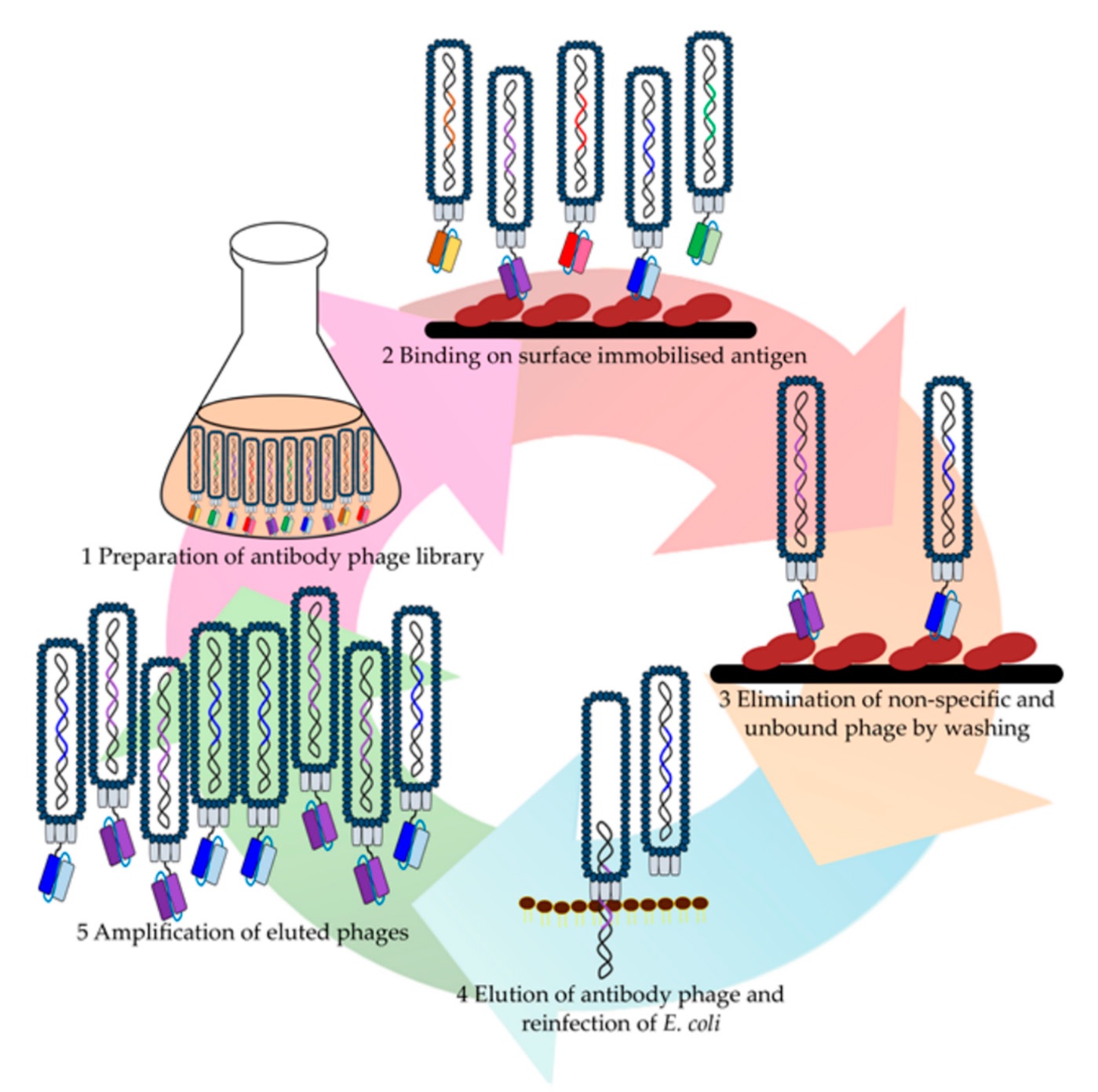
The quality of the resulting fragment library depends on the ability to obtain a variety of Abs. Two library-generating approaches exist, viz. natural (naïve) and synthetic.
Natural libraries of Ab variable domains are obtained by reverse-transcription polymerase chain reaction (RT-PCR) of natural genes from human or animal lymphoid tissues or peripheral blood. The strong point of this method is that the functional Abs encoded by the resultant genes are in the right conformation. The weak point is that the coverage of the natural immune system limits sequence diversity. Furthermore, the quality and composition of fragments from natural libraries vary widely and many fragments may be unstable or may be unacceptable for other reasons [36].
Synthetic libraries are generated by inserting artificially synthesized DNA into sequences encoding variable domains. DNA is synthesized to introduce completely random mutations into the resulting Ab fragments and thereby increase the diversity of the complementarity-determining CDR site. Synthetic libraries allow the use of a specific core structure of the variable domain, known to be the most represented one in each individual, nonimmunogenic, and stable. On the other hand, the addition of completely synthetic CDR sites may cause protein misfolding and aggregation.
An important advantage of phage display over hybridoma methods is its ability to select only those Abs that do not bind to unwanted proteins, such as those that are close to the target but do not need to be blocked to avoid causing toxicity. For this purpose, it is enough to immobilize the now unwanted protein and select those phages that have not bound to it.
A library for selecting Fab fragments from a vaccinated human was also described, which can be considered a human immune library. Libraries derived from naturally immunized humans, such as vaccinated people, are difficult to find, but even libraries from nonimmunized humans have been used to successfully obtain high-affinity Abs against auto- or non-self-Ags. The main strategies for constructing phage libraries are presented in [37].
Other types of display, such as ribosome and yeast display [38], are based on the same principle. They provide a link between the gene encoding the Ab fragment and its affinity for the target (i.e., phenotype), and they allow selection for a given property from a huge number of variants in the library. Here we focus on phage Abs and their use to detect diagnostically important Ags.
Biosensors are very promising for rapid Ag analysis. Biosensor-based methods have a wide range of applications and are gradually becoming integral in clinical diagnostics [39], environmental monitoring [40], cellular physiology [41], and studies of space effects on astronauts [42]. Biosensors are designed according to the substance to be analyzed and the mechanism of the transducer used. Biosensors have revolutionized the field of diagnostics by combining biological sensing elements with transducers to convert the recognition of a specific target molecule into a measurable signal [43, 44].
Antibodies, microbial cells, aptamers, enzymes, and other objects can be used as sensing elements [44]. Antibodies are a classical recognition tool, and the specific Ag-Ab interaction is used widely in Ag-detecting sensors. Phage display enables preparation of Abs to low-molecular-weight Ags (haptens), and phage Abs are very promising as bioreceptors.
Several studies have used phage Abs to detect various target analytes. Phage Abs have several advantages over their natural counterparts:
-The small size of Ab fragments is usually accompanied by a reduction in nonspecific binding.
-Phage Abs can be immobilized more densely.
-Phage Abs can reproduce in bacteria (E. coli), which strongly reduces production costs [45].
These advantages determine the prospects for phage Ab application in biosensors [46]. Success in the application of phage Abs as sensing elements depends on the preliminary refinement of the method used for their production and testing. In what follows we consider the prospects for the use of phage Abs in sensing systems to detect diagnostically important Ags.
The number of patients with allergic reactions increases constantly, already exceeding 150 million. It is predicted that in Europe in the next decade allergy will become a public health problem on the scale of a pandemic [47].
Allergy is the body’s immune response to an Ag, which is characterized by damage to cell structure and function. If this reaction is excessive, the damage can extend to tissues and organs. On entering the body, Ags provoke protective immune phenomena: Ag recognition by the immune system, synthesis of Abs (immunoglobulins), and activation of T (killer) cells. Foreign organic substances, microorganisms, and products of microbial vital activity can all act as Ags. Fig. 3 shows the main types of allergens.
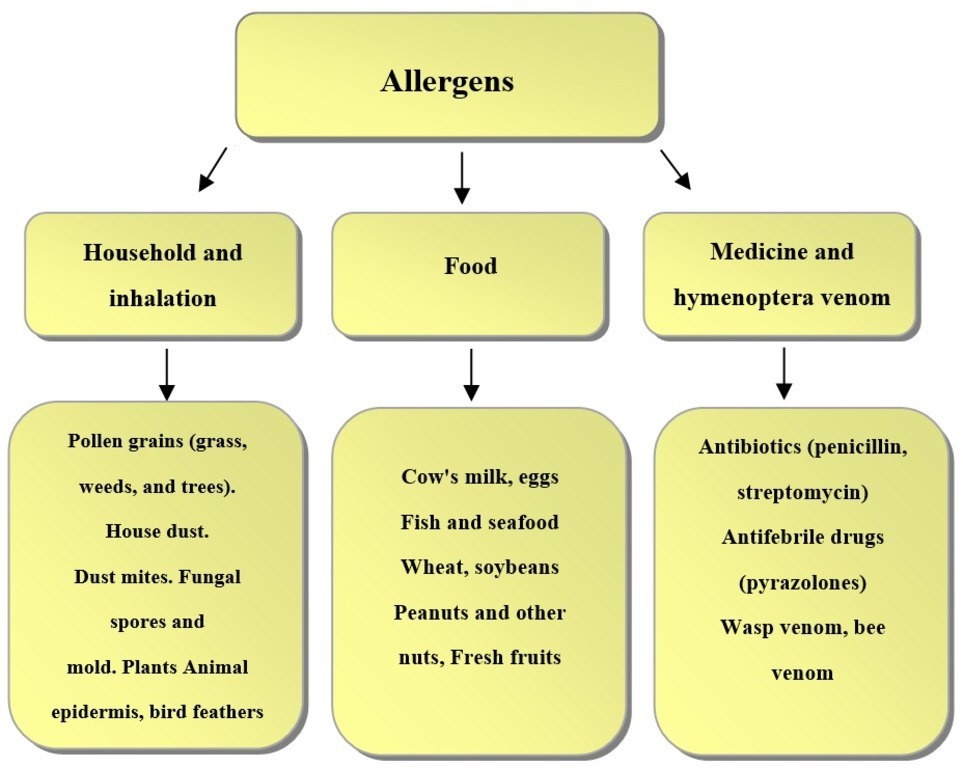 Fig. 3.
Fig. 3.Main types of allergens.
On the basis of their structure and properties, allergens are divided into complete and incomplete, and they differ in the nature of their interaction with Abs. Most often, complete allergens are protein compounds that trigger immune responses on their own. Such allergens include food additives, pet hair, pollen, and others. Incomplete allergens, also called haptens, are low-molecular-weight substances that cannot trigger immune responses on their own. They become complete when they are combined with body proteins (carriers) and form a high-molecular-weight complex. This complex provokes an allergic reaction. Usually, drugs, paints, metals, plastics, latex, and other chemical products act as incomplete allergens.
A hapten can change the structure of its carrier, and then the body recognizes foreignness not in the allergen but in its own metabolites (products of cellular activity). This phenomenon, called an autoallergic reaction, can catalyze autoimmune diseases – chronic pathologies in which Abs are produced against tissues of self, eventually destroying them. Some examples of autoimmune diseases are systemic lupus erythematosus, rheumatoid arthritis, and multiple sclerosis.
Allergies are characterized by specific inflammations associated with the interaction between allergen and allergen-specific Immunoglobulin E (IgE) [48]. Major allergens contain B-cell (conformational and linear) epitopes, responsible for allergic reactions. The most reliable way to determine an allergy is through a blood test, which determines specific Abs to various allergens and identifies groups of substances responsible for allergic reactions [49].
Food allergy is a hypersensitivity reaction that affects about 4–10% of the world’s population, and its occurrence may be increasing [50, 51]. Phage Abs are a powerful tool for food allergen detection. Single-chain variable Abs (scFv) for determining allergenic nut proteins in foodstuffs were described in [52].
The use of TomLinson I+J artificial libraries (MRC Laboratory of Molecular Biology and MRC Centre for Protein Engineering) resulted in phage-ELISA systems for the successful detection of almond and walnut proteins in commercial food products.
Cross-linking of IgE molecules with the appropriate food Ags results in allergic reactions. The AllerScan programmable phage display device was designed to use this principle to probe the binding specificity of antiallergenic IgG and IgE Abs in serum. By using AllerScan, the persistence of the anti-wheat IgE reaction was shown in people with a wheat allergy but not with wheat sensitivity [50].
Finding allergen epitopes is essential to understanding the pathophysiology of type I allergies and the existence of cross-reactivity.
Mario Geysen first used the term “mimotope” in 1986 [53] to refer to peptides that mimic epitopes. These peptides can compete with native proteins for Ab binding by mimicking protein-binding sites. The peptides displayed on the phage and recognized by antiallergic Abs mimic the physicochemical characteristics of amino acids and, therefore, are mimotopes. In addition to being used for the competitive immunoscreening of peptide libraries, phage particles are helpful as immunogenic carriers of allergen mimotopes [54]. Fig. 4 (Ref. [47]) shows the identification of allergen mimotopes by screening peptide libraries displayed on bacteriophages.
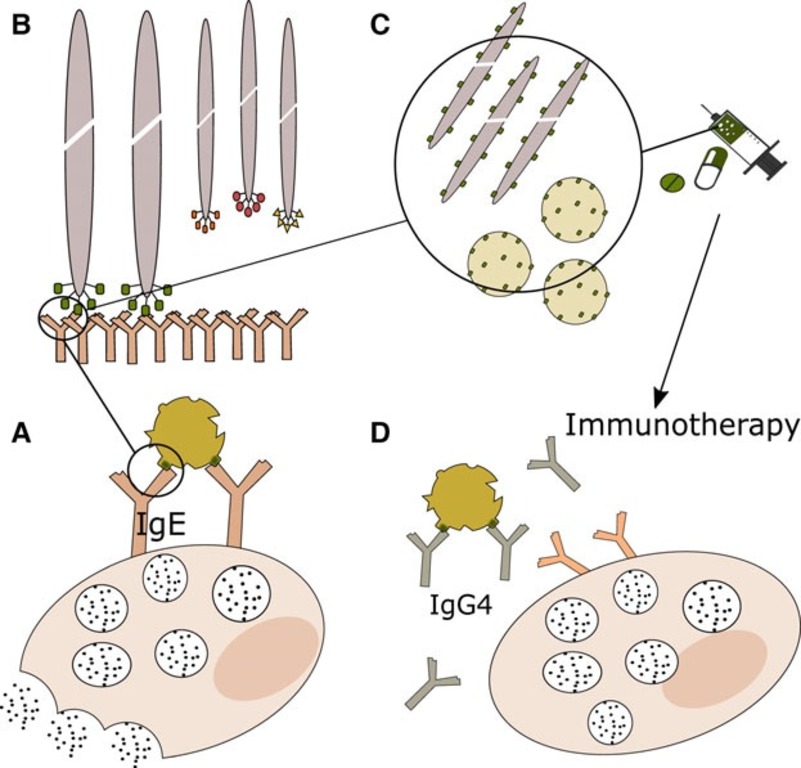 Fig. 4.
Fig. 4.Phage particles as immunogenic carriers of allergen mimotopes. Phage particles displaying oligopeptide libraries enable identification of allergen mimotopes (A), peptide mimetics of epitopes’ physicochemical properties through competitive immunoscreening (B). They are suitable immunogenic carriers (C) and function in mimotope immunotherapy (D) [47].
Phage display facilitates the identification of dominant epitopes of allergens and can subsequently be used to develop mimotopic immunotherapy [47]. By identifying IgE epitope profiles in patients, it is possible not only to recognize cross-reactivity but also to improve diagnosis and immunotherapy. The prospects for the use of phage peptide display to identify IgE-binding mimotopes (simulators of natural epitopes) were presented in [51]. This approach offers a promising alternative to crystallography for the identification of allergen epitopes that contribute to the formation of allergen-Ab complexes. When the interaction between serum IgE and specific allergens is fully characterized, it will be possible to develop new therapeutics based on high-affinity competing Abs and/or peptides.
Ig sequences produced by allergen-specific B lymphocytes were characterized by Hoh et al. [48] in a 3-year study of patients with aeroallergies who received targeted immunotherapy. Allergen-specific IgE-expressing clones were identified by using combinatorial libraries of scFvs in nasal biopsy specimens over a 3-year period with Ab gene repertoire sequencing. The characteristics of private IgE-expressing clones were compared with those of stereotyped, or “public”, IgE responses to grass pollen.
Petrenko et al.’s study [55] identified a panel of phage mimotopes closely associated with the cellular receptors interacting with the viral spike protein. Their interaction was shown by ELISA and confirmed by molecular modeling. The phage probes that compete with viruses for binding to specific cellular receptors can provide important information on the mechanisms of virus infectivity. They may also be essential for vaccine or drug development or be used as interfaces in diagnostics.
In developed countries, nut allergy is a serious public health problem, and reliable allergen detection methods are needed to comply with food-labeling regulations.
Recombinant Abs against pistachio nuts were produced by phage display [56]. These Abs can be used in immunoassays to detect allergenic pistachio in foodstuffs. The authors selected a clone producing the PVF4 phage-domain Abs (dAbs) that did not cross-react with cashew despite its being evolutionarily close to pistachio. The basic component of pistachio 11S globulin (allergen Pis v 2) is linked to a distinct band of 22 kDa that this clone detects, according to the results by Western blotting and matrix-assisted laser desorption/ionization tandem mass spectrometry. The suitability of the PVF4 phage-dAbs for the examination of 77 commercial foodstuffs was evaluated by enzyme-linked immunosorbent assay.
Another study described the isolation of almond-specific scFv by using a commercial phage display library [57]. Two scFv (PD1F6 and PD2C9) were used, which had been isolated after two biopanning rounds. An indirect ELISA detected almond protein in food products. The isolated scFv were highly specific and detected raw and roasted almond protein, respectively. The indirect ELISA with phage Abs was validated by analysis of 92 commercial food products, and the results correlated well with those yielded by a previously developed real-time PCR method for the detection of almond in food. In addition, scFv can be genetically engineered with a wide variety of expression tags such as enzymes and fluorescent probes that can be used in a multitude of assay formats.
de la Cruz et al. [58] showed the prospects for obtaining fragments of recombinant Abs from a naïve library (Tomlinson I and J) that are specific to an allergenic Brazil nut protein. The selected phage scFv was further used as an affinity probe to develop an indirect phage-ELISA for nut allergen detection in experimental binary mixtures and commercial food products.
Monitoring of various substances, including proteins, in a living organism is much in demand. Membrane proteins are the most popular targets for diagnostic and therapeutic applications. The membrane protein structure usually consists of an extracellular, a transmembrane, and an intracellular domain. Extracellular domains are the main targets of drug discovery and diagnostics. About 60% of drug targets are membrane proteins [59], which fulfill the following functions:
• They are implicated in signal transduction and in cellular channels (thus, drugs targeting them can effectively manipulate cellular functions).
• They are present on the cell surface and, therefore, are more accessible than cellular proteins.
Certain membrane proteins can determine cell types. Membrane proteins, for instance, are markers of the differentiation cluster that identify different types of immune cells. More importantly, diseases typically change the expression levels of membrane proteins. One well-known example is the overexpression of the membrane protein of human epidermal growth factor receptor 2 (HER2) in 20–25% of breast cancers [59]. Presenting correctly folded targets, i.e., native structures of extracellular domains (ectodomains) in membrane proteins is essential for effective phage display.
Khondee and Piyawattanametha [59] used phage peptides for the detection and treatment of diseases, including clinical imaging. They pointed out that molecular imaging based on appropriate sensors (phage peptides) offers invaluable possibilities for studying complex disease-related bioprocesses in vivo, with early detection of cancer and metabolic disorders.
Many studies have shown that phage Abs are more sensitive than other types of Abs [60, 61]. The use of single-domain Abs (sdAbs) strongly reduces limits of detection (LODs) (by 5–10 times), as compared with the use of natural Abs or their soluble fragments. Such enhancement of the detection signal shows the promise of the use of Ab fragments in biosensing [62]. Phage Abs to Brucella melitensis are excellent reagents for ELISA because of the strong signal amplification ensured by antiphage Abs. The isolated phage Abs are highly specific for B. melitensis and do not recognize Yersinia pseudotuberculosis, in contrast to existing diagnostic monoclonal Abs [60]. Also phage Abs can independently detect Marburg virus variants without cross-reactivity with Ebola viruses, and the assay takes less than 30 min [61].
A method was described for the detection of ferritin in animal serum by using phage Abs. The prepared Abs were tested by dot immunoassay of ferritin isolated from the cow liver and ferritin present in the blood sera of cattle. Because the Abs specifically recognized ferritin, this test system opens prospects for the development of a similar biosensor [63].
In yet another study [64], phage display was used to obtain biomarkers for brain injury diagnosis. Fab fragments of Abs to rheumatoid factor were obtained by phage display, and for IgG studies in rheumatoid arthritis patients, the pComb3 vector was used to create a library of combinatorial IgG Abs [65].
One of the main causes of death is tuberculosis. Current clinical diagnostic
techniques, such as sputum culture, PCR, and smear microscopy, are not flawless.
Although Ab-based assays require particular Abs against an appropriate biomarker,
they are acceptable substitutes. Mycolic acid, which is present in patient sputum
samples and makes up a major part of the mycobacterial cell wall, is a prime
target. However, the use of mycolic acid as a diagnostic marker is limited
because of the difficulty of generating antilipid Abs by standard hybridoma
procedures. Thus, Chan et al. [66] isolated and characterized four
anti-mycolic acid Abs from a nonimmune phage display library. The Abs could
detect mycolic acids up to an LOD of 4.5 ng. The prepared Abs were specific for
methoxy mycolic acids, exhibiting weak binding to
Isocitrate lyase (ICL) is an important enzyme for the growth and survival of M. tuberculosis during latent infection, because it is involved in the mycobacterial glyoxylate and methylisocitrate cycles. ICL inhibition may disrupt the M. tuberculosis life cycle. Phage display Abs was used to identify an scFv Ab against the recombinant ICL protein from M. tuberculosis. The soluble a-ICL-C6 scFv clone showed a good binding ability and inhibited ICL activity in vitro [67].
Malaria is a disease that mainly affects the poorest populations in many countries of the world, where it is most often fatal. Therefore, early testing for malaria is one of the World Health Organization’s (WHO) core principles for the control of the disease. Worldwide, about 429,000 people die from malaria per year [68].
Five species are associated with human malaria: Plasmodium falciparum, P. vivax, P. malariae, P. ovale, and P. knowlesi [69]. P. falciparum and P. knowlesi cause human life-threatening malaria. These parasites infect red blood cells, inducing physicochemical changes in the erythrocyte plasma membrane. Currently, malaria detection tests (traditional microscopy, rapid diagnostic tests, nucleic acid amplification tests, and ELISA) have disadvantages such as high costs and need for specialized laboratories and highly trained personnel. The potential of different immunosensors for malaria diagnosis was described in [69].
The use of phage display to obtain malaria-specific biomarkers is important for the early diagnosis of the disease. Eda et al. [70] used a phage display library to identify a peptide, LVDAAAL, which is a target compound for the design of antimalarial agents.
Phage display has been used to develop Abs against a wide variety of eukaryotic pathogens. These recombinant Abs target parasites, including Taenia solium; protozoa, e.g., Cryptosporidium parvum, Plasmodium falciparum, Toxoplasma gondii; and fungi (Aspergillus fumigates) [1]. Also targeted are veterinary pathogens, including the fish pathogen Myxobolus rotundus, the dog pathogen Babesia gibsoni, and the plant pathogens Aspergillus niger, Fusarium verticillioides, and Sclerotinia sclerotiorum [1].
One method for detecting adhesins in pathogens that are challenging to culture in the laboratory or difficult to manipulate genetically is to utilize phage display in vivo. Among these pathogens is Borrelia burgdorferi, the causative agent of Lyme disease. It is spread from the site of a bite by ixodid ticks and causes systemic infection. Although B. burgdorferi molecules that facilitate contact with the endothelium in vivo have not yet been found, dissemination takes place through the circulatory system. A number of novel adhesin candidates were discovered through the in vivo selection of filamentous phages expressing B. burgdorferi protein fragments on the phage surface [71].
When pathogen-specific Abs are found in human blood, it means that the patient has previously been exposed to and infected with that specific pathogen (such as a virus or bacterium). Immunoassays such as ELISA and Western blotting are typically used to measure diagnostic Abs. A serodiagnostic method called “Domain-Scan”, based on the phage display of epitopes, was published by Hada-Neeman et al. [72]. They measured serum binding to several hundred epitopes that were concurrently produced from HIV-1 and HCV by next-generation sequencing. Machine learning was used to simulate the problem of separating healthy people from those infected with HIV-1 or HCV on the basis of values corresponding to the content of determinant-specific Abs in the sample. If enough training samples are available, it may be possible to extend the experimental computational method of Domain-Scan to different diseases.
By using phage display, a peptide interacting with the YB-1 protein was
detected. The YB-1 protein regulates the transcription and translation of many
genes involved in cell division and differentiation. On the basis of monoclonal
Abs and a peptide within the bacteriophage structural protein, tests were
developed for the measurement of the YB-1 protein in serum by using ELISA,
Western blotting, and immunocytochemistry. The LOD was 0.3 ng mL
A surface plasmon resonance–based biosensor was proposed for the measurement of carbonic anhydrase in biological fluids. For analyte detection, the authors used a Biacore T200 device and gold nanoparticle-labeled anti-carbonic anhydrase Abs obtained by phage display [74].
Rheumatoid arthritis (RA) is a very widespread chronic autoimmune disease in the world. Despite progress in the treatment of RA, many RA patients often fail to achieve remission. Semaphorin 5A (SEMA5A) plays a key part in the progression of RA, promotes pannus formation, and is a therapeutic target. Qin et al. [75] described a treatment strategy using human Abs (SYD12–12) generated against semaphorin 5A. SYD12–12 effectively inhibited angiogenesis and aggressive phenotypes of RA synoviocytes and bone destruction in mice in vitro.
The use of phage Abs for the detection of cancer biomarkers is very promising [76, 77, 78]. Abs that have been obtained by phage display and have received clinical approval have been adapted for use in cancer (36%) and immunoregulation (32%) research [37].
Adalimumab (Humira) is the first phage display–derived monoclonal Ab and the first human Ab approved for therapy. In 2002, adalimumab (Humira) became the phage display–derived Ab to receive marketing approval and is now the best-selling Ab. In 2016, six human Abs were discovered or further engineered by phage display and subsequently were approved for therapy [79].
Phage Abs have been used to detect melanoma and breast cancer [80]. Phage Abs bound to five of six tumor cell lines and showed no measurable reactivity against normal cells. This result is an important step toward effective immunotherapy for cancer treatment. Biopanning of the phage library yielded specific Ab fragments to hepatocellular carcinoma, which makes it possible to use phage Abs in tumor immunotherapy [81]. Potential therapeutic Abs against chronic lymphocytic leukemia were obtained by selecting Abs to tumor-associated proteins from immune phage libraries [82]. They were also used in the quantitative visualization of tumor cells by dark-field microscopy utilizing light-scattering nanoparticles for plasmon resonance [83].
The essential biomarker for prostate cancer screening is prostate specific antigen, or PSA. Using a sandwich pair of nanobodies against PSA, Liu et al. [84] made a sensitive and selective immunosensor for the quick measurement of serum PSA. The nanobodies were obtained from an alpaca-derived immune phage display library. The immunosensor showed high selectivity, stability, and reproducibility and can serve to determine serum PSA.
Antibodies specific to heat shock proteins isolated from MN22a mouse hepatoma cells and Sp2/0-Ag14 mouse plasmacytoma cells were affinity selected by using a naïve human scFv library. The dynamics of the heat shock protein concentration in the sera of mice with implanted MN22a cells were investigated by using the obtained phage Abs, dot immunoassay, and ELISA [85]. After implantation of tumor cells, tumor growth was accompanied by a significant increase in the serum accumulation of heat shock proteins. Mini-Abs specific to heat shock proteins were effective for determining and monitoring the accumulation of heat shock proteins in animal sera.
By using a sheep display library of scFv (Griffin.1, UK), Abs specific for heat shock proteins were prepared, and the proteins were detected in sera of diseased animals by dot immunoassay by using the biospecific interaction of phage Abs. The heat shock protein in the sera of spontaneously infected animals was detected by ELISA with specific phage Abs [86].
Several studies have shown that phage display can be successfully applied to identify novel peptides targeting different types of cancer tumors, such as bladder cancer, gliomas, and osteosarcomas [34].
Breast cancer is very common. One cause of death from breast cancer is metastasis to the lymph nodes, lungs, liver, bones, and brain. A widely used drug for the treatment of metastatic breast cancer is paclitaxel. However, the clinical use of paclitaxel is limited because of its insolubility in water and complications after use in patients. For improving the therapeutic efficacy and reducing the side effects of paclitaxel, the possibility was examined that it can be encapsulated into micelles, with subsequent targeting to tumor cells with phage Abs as the targeting agent. The anticancer effect was increased because of the improved drug delivery to the tumor and because of selective toxicity to cancer cells in vitro [87].
In another study [88], an algorithm was proven for analyzing phage populations that had evolved as a result of multistage screening of landscape phage libraries against MDA-MB-231 breast cancer cells. The suggested model for combinatorial avidity selection proposed a multistage accumulation of elementary binding units, or core motifs, in landscape phage fusion peptides. These motifs serve as evolutionary initiators for the formation of short linear motifs. Directed molecular evolution may be useful in combination with combinatorial selection to design new smart materials with a range of unique features.
The Center for Disease Control and Prevention classifies several bacterial toxins and/or toxin-producing pathogens as category A or B agents, because they pose a high risk to national security and public health [89]. Such toxins are relevant targets for development of diagnostic Abs, and Ab phage display is a powerful tool for selecting diagnostic Abs. In order to obtain Abs against poisons that cannot be utilized to immunize animals, phage display is frequently absolutely necessary. Specifically, anti-botulinum neurotoxic [90] and anti-crotoxin [91] phage Abs were obtained. These results show the potential of using various libraries of recombinant single-chain fragments in the synthesis of antidotes, in addition to their diagnostic usefulness. Methods have been proposed to detect staphylococcal enterotoxins by using phage Abs in immuno-PCR [92] and in a biosensor [93]. The limit of detection of staphylococcal enterotoxin A in milk by the phage display method was 100 pg/mL, well below the toxic–allergic dose.
Phage Abs have been obtained for immunochemical analysis of toxins, including Bordetella pertussis adenylate cyclase toxin, Clostridium perfringens alpha-toxin and epsilon-toxin, anthrax toxin, Clostridioides difficile toxin, Bacillus thuringiensis cry toxin, Bordetella pertussis CyaA-hemolysin, diphtheria toxin, E. coli enterotoxin B, Microcystis aeruginosa microcystin, Nodularia spumigena nodularin, P. aeruginosa exotoxin A, Salmonella typhi hemolysin E, shiga toxin, tetanus neurotoxin, S. aureus toxic shock syndrome toxin-1, Helicobacter pylori vacuolating cytotoxin A, Vibrio parahaemolyticus hemolysin, V. vulnificus toxin, and V. cholerae toxin subunit B [1].
Mullinax et al. [94] used a combinatorial library to obtain clones with tetanus anatoxin-binding activity and obtained fragments that had high binding affinity for tetanus anatoxin and did not cross-react with other proteins. The development of some potentially useful therapeutics may require the selection of human Abs, which are found much less frequently in the repertoire and, therefore, are difficult to obtain by the hybridoma method. This would be the case for Ags that elicit little response from the human immune system.
Enrichment with specific peptides is effective for arsenic binding [95]. A hazardous poison called arsenic is utilized in semiconductors (such as gallium arsenide). The SxHS and carboxy-terminal QxQ motifs, which may be implicated in arsenic binding, were discovered by the application of phage display and bioinformatic techniques.
For immunoassay of the MC-LR mycotoxin, Akter et al. [96] used Ab fragments selected from an immunized library together with antiimmune complex Ab fragments from a naïve phage display library. The MC-LR was detected by noncompetitive immunoassay with high sensitivity and specificity. To date, it is the fastest reported MC-LR immunoassay, with a detection limit well below that recommended by WHO.
Phage scFv pAbs to zearalenone, a mycotoxin made by Fusarium species, were developed. Zearalenone is heat stable and has estrogenic effects on mammals [97]. There was no cross-reactivity between the phage-derived Abs and other common mycotoxins. The application of these Abs in an LFIA biosensor is linked to additional optimization.
Phage sdAbs were more sensitive than soluble sdAbs and llama polyclonal Abs, as shown by ELISA. Despite the comparatively high particle size of phages, flow fluorimetry demonstrated little nonspecific binding. In addition to being extremely specific indicators, phage sdAbs produce remarkably stable signals. In terms of overall sensitivity, phage Abs outperform polyclonal Abs by 5 times [62]. They can be used in ELISA, fluorescence immunoassay, and dot blot analysis for the highly sensitive detection of ricin.
Gandhi et al. [98] described a test strip that uses gold nanoparticle–labeled scFv Abs for morphine detection (Abs were obtained by using a phage display–based antibody library). Antimorphine scFv Abs were labeled with gold nanoparticles and used as an optical immunoprobe. A competitive assay with the test strip characterized the ability of the scFv Abs to recognize free morphine (within 5 min) and could be useful for the in situ screening (actual blood, urine, and saliva samples).
Progress in phage engineering has led researchers to develop a new kind of phage display library that consists of a collection of viral surface–localized nanofiber materials with various molecular landscapes. These nanomaterials, known as “landscape phage”, are invaluable for the design of Ag detection techniques and serve as building blocks for biosensors [99]. The recognition probes produced from landscape phage are used in sensor systems to form reliable and affordable molecular recognition interfaces. These probes interact with biohazard agents and produce detectable signals. The use of landscape phage-based interfaces can largely improve the sensitivity, selectivity, reliability, and durability of these devices.
Phage display is used widely to prepare novel Abs. About 18% of all commercially approved Abs owe their origin to this technology (Fig. 5) [37].
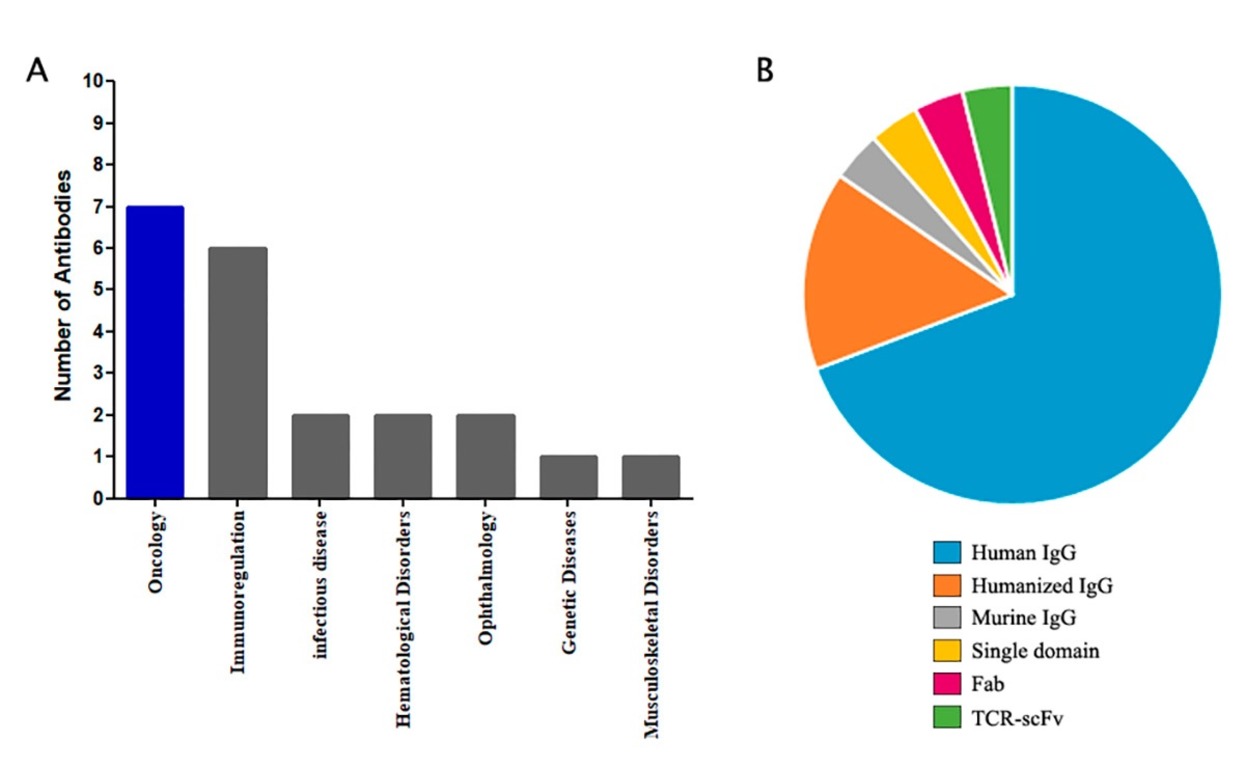 Fig. 5.
Fig. 5.Statistics of Abs derived from phage displays that are approved for clinical use (A) and proportion of each Ab type (B) [37].
An important factor in selecting a recognition element for a sensor is the shelf life and stability of the receptor. In this respect, phage Abs are characterized by high stability, improved solubility, and solvent compatibility, which makes them of particular value for use in biosensors [45].
A key quality parameter for an Ab library is its diversity (reflecting the probability of screening an Ab clone from the library against an introduced Ag with high enough affinity) [100].
High-throughput phage-selection techniques have produced a variety of cancer cell–linked phages and phage proteins with an innate ability to self-assemble into nanotransporters that target genes and medicines [99, 101]. It can be argued that Ab phage display has revolutionized the field of diagnostics and therapeutics. In the past two decades, phage display-derived Abs have been used widely in medicine and veterinary medicine as diagnostic and therapeutic agents [100]. Phage display offers new possibilities in biosensor design, as shown with electrochemical, optical, and electroacoustic sensors [8, 9, 28].
Phage display is developing rapidly as a biosensor-making tool. The combination of phage display with various biosensor platforms is attracting increasing attention. The use of phage display together with biopanning allows rapid enrichment of certain molecules in a library, thereby expanding the potential for biorecognition probes owing to the simplicity, speed, and ease of Ab generation. A potential approach to sensor improvement by using phage display is to make multimodal and multitargeted systems for the simultaneous detection of different analytes. It is expected that the interdisciplinary collaboration of engineers and scientists to develop field-applicable sensors and the advancements in phage-based sensor research will lead to considerable progress in food safety monitoring, environmental monitoring, and infectious disease diagnostics. Conditions are being established for the rapid and cost-effective high-volume production of phage Ab-based biosensors. The combination of phage Abs with analytical devices is a powerful tool for the detection of targets ranging from biological agents (proteins, bacteria, spores, viruses, toxins, etc.) to chemical agents or even explosives. It can be expected that in the near future, owing to phage display, a whole generation of new phage Ab-based biosensors will be available for use in analytical chemistry, molecular biology, and medicine [45, 102, 103].
Phage display is a robust and versatile platform for Ag detection and a powerful engineering tool to improve Ab properties. It allows the display of Abs used for the phage coat protein on the surface of filamentous phage. Although the advances in phage Ab production have been impressive, it is necessary to foster cooperation between research groups in different scientific fields to further develop phage display technology to produce specific Abs and their applications in sensor technologies. Such interdisciplinary cooperation between experts from different scientific fields would lead to increased applications of phage Abs as sensing elements for biorecognition.
OIG: Conceptualization and design, Project administration, Formal Analysis, Writing – original draft, Writing – Review & Editing. VAK: Formal Analysis, Writing – Review & Editing. LAD: Project administration, Formal Analysis and design, Writing – original draft, Writing – Review & Editing. All authors have read and agreed to the published version of the manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This research was supported by the Russian Science Foundation Project No. 24-29-00463.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.