


 , Coralie Silva 1,2
, Coralie Silva 1,21 Molecular and Environmental Biology Centre (CBMA), University of Minho, 4710-057 Braga, Portugal
2 Institute for Science and Innovation on Bio-Sustainability (IB-S), University of Minho, 4710-057 Braga, Portugal
3 Aquatic Research Network (ARNET), CBMA, University of Minho, 4710-057 Braga, Portugal
Abstract
Microbes are traditionally regarded as planktonic organisms, individual cells that live independently from each other. Although this is true, microbes in nature mostly live within large multi-species communities forming complex ecosystems. In these communities, microbial cells are held together and organised spatially by an extracellular matrix (ECM). Unlike the ECM from the tissues of higher eukaryotes, microbial ECM, mostly that of yeasts, is still poorly studied. However, microbial biofilms are a serious cause for concern, for being responsible for the development of nosocomial infections by pharmacological drugs-resistant strains of pathogens, or for critically threatening plant health and food security under climate change. Understanding the organization and behaviour of cells in biofilms or other communities is therefore of extreme importance. Within colonies or biofilms, extremely large numbers of individual microbial cells adhere to inert surfaces or living tissues, differentiate, die or multiply and invade adjacent space, often following a 3D architectural programme genetically determined. For all this, cells depend on the production and secretion of ECM, which might, as in higher eukaryotes, actively participate in the regulation of the group behaviour. This work presents an overview of the state-of-the-art on the composition and structure of the ECM produced by yeasts, and the inherent physicochemical properties so often undermined, as well as the available information on its production and delivery pathways.
Keywords
- extracellular matrix
- yeasts
- multicellular aggregates
- polysaccharides
- structure
- physicochemical properties
Microbial cell communities occurring in biofilms, mats, flocs or colonies, are possibly the oldest, most successful and widespread form of life on Earth [1]. Nevertheless, it is a fairly new notion that, although microorganisms are able to live as individual cells, in nature they rather live in large, organized communities, often containing multiple species from very diverse levels of organization, which help them cope with constantly changing environments, and biotic and abiotic stressors. This notion raises major questions, like how these communities are structured, how do microorganisms organize collectively, providing access to food and water to the whole community, and how they deal with competitors or predators. Ultimately, how do microorganisms communicate intra- and inter-species? Answers to these questions may provide some insight on the very origin of multicellularity, while simultaneously it requires that we look at microbial communities from an ecological point of view, as being part of a greater ecosystem, be it located in a pond or a river, in a pipe of any industrial infrastructure, or co-habiting higher order organisms as part of a plant or animal microbiota.
Biofilms in particular have driven considerable attention from researchers, mostly because of the undesirable effects associated to their development. Most often they are formed by a variety of species from one or more distinct group or kingdom, including algae and microscopic animals [2]. In specific niches, they can also be formed by a single type of microorganism, like bacteria [3], filamentous fungi [4] or yeasts [5]. Importantly, biofilms are found attached to biotic or abiotic surfaces in almost all humid or aqueous environments which include urban, agricultural or industrial facilities [6, 7, 8, 9, 10] as well as naval and sea or river-associated infrastructures [11]. Their presence causes process inefficiency, energy waste and economic losses and contributes to the spread of invasive species in many river and estuarine ecosystems with disastrous consequences for the environment [12]. Nevertheless, biofilms are most studied in regard to human health, because of their insidious development on medical devices and hospital equipment and facilities [13, 14]. In these locations, biofilms are vehicles for the development and spread of opportunistic microorganisms, which can originally be commensal, but shift into living tissue-invading virulent forms [15]. Biofilms are thus frequently the cause of nosocomial infections [16, 17]. The colonization of hospital-related surfaces also favours the contact of infectious microorganisms with low and erratic doses of pharmaceuticals, contributing to the development of multiple resistances. The most well-studied of these biofilm-forming microorganisms are the bacteria Staphylococcus aureus [3] and Pseudomonas aeruginosa [18], as well as the yeasts from Candida genus [19]. In addition, biofilms are also associated with pathologies in non-human animals [20] and plants [21], but in spite of the increased frequency and devastating consequences of these diseases [22, 23, 24, 25, 26], the associated knowledge is very disproportionate when compared to what is known about human diseases.
Biofilms provide cells with communal benefits, such as increased resistance to antibiotics, antifungals or other drugs, as well as to host defences [27, 28, 29, 30]. Also, external stimuli derived from climate or other origins, such as temperature, osmotic or oxidative stress, or nutrient deficiency, cause a much lesser impact on the survival of individual cells within biofilms when compared to planktonic cells [31, 32, 33]. Cells within a biofilm display altered plasma membrane sterol composition [34], and an increased expression of multidrug-resistance pumps [35, 36], both contributing to the higher resistance to drugs and pharmaceuticals. In these environments, cells also tend to differentiate into more invasion-prone forms, like fungal penetrating hyphae or haustoria [37, 38]. Most importantly, they secrete abundant extracellular matrix (ECM) [5]. This provides physical and life support while promoting cell-cell communication and the control of differentiation, community expansion/cell division, quorum sensing (QS) and invasion of adjacent or farther locations, all of which are former prerogatives of the ECM from animal tissues. Microbial ECM is often designated as extracellular polymeric substance (EPS), although this designation is also very often used to refer to the actual polymers that compose the ECM.
Globally taken, the knowledge on microbial ECM composition, architecture and biological roles is rather scattered and incomplete, especially in what concerns yeasts [39, 40]. Moreover, discrepancies are also found in the chemical and molecular characterization of the components of a same microorganism ECM. This has primarily to do with the different conditions in which the cells are cultivated, but it also derives from the different methodologies applied to extract and fractionate the ECM for chemical characterization [41]. For example, harvesting fungal ECM by concentrating it with ethanol, as used by Beauvais et al. [42], is good for precipitating polysaccharides, and has also been used to analyse the polysaccharide fraction from the biofilm ECM of other mucous-producing fungal species [43]. Nevertheless, this method is not appropriate to allow the detection/quantification of the glycoproteins in the same samples, which amount can thus end up very underestimated as discussed by Singh et al. [43]. On the other hand, the solubilization of the ECM deproteinized fraction in trichloroacetic acid (TCA) or other organic solvents [1, 41, 42, 43, 44] can yield different compounds. N-acetyl-glucosamine (GlcNAc) is virtually insoluble in water [45], while glucose and other common sugars or melanin cannot be quantified in TCA-solubilized samples. Moreover, some authors use very diverse methods for identification and quantification of the compounds in each fraction which differ substantially in accuracy and detection levels. These vary from classical chemical methods, and enzymatic or immunoenzymatic assays, to spectrophotometry [43] or complex chromatography (reviewed by [41]). Chemical fractionation is unavoidable, but the procedure may condition the analytical results, namely by jeopardizing the detection of all the compounds, especially those in smaller amounts, or by biasing their relative concentrations.
Yeasts’ ECM is basically not yet biochemically and molecularly understood. What molecules and what compounds form the ECM? Which could be their specific roles in the several stages of the establishment of a biofilm or a colony? What would be their contribution to the physical properties of the biofilm and in commanding the fate and the behaviour of the cells within? What pathways and molecular mechanisms are involved in their production and secretion? These questions are mostly unanswered. Yet, the knowledge from their answers is indispensable to meet the demands of the increasingly more spread and more virulent fungal diseases in plants and animals, including humans [46, 47, 48]. This work gathers the information available regarding the ECM from yeasts, from the most well-known human pathogens, like Candida albicans and Cryptococcus neoformans, to the industrially important exopolysaccharide producer Aureobasidium pullulans, and the unavoidable model yeast Saccharomyces cerevisiae.
The exopolysaccharides produced and secreted by yeasts, either as part of
biofilms or colonies ECM, or simply accumulating in the growth media of
planktonic cultures, differ in composition, structure and molecular weight (MW).
Information on their chemical structure can be found in the literature for 12
species of Ascomycetes and 27 of Basidiomycetes (Table 1, Ref. [17, 42, 43, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121]). In
total, 21 different exopolysaccharide core structures were established for
Ascomycetes, and 34 for Basidiomycetes. From these last, 95% are branched, from
which, 15% are highly branched. The available information on the structure of
these branches does not always match the information available on the core
structure (Table 1). The (1,3)/(1,6) and
| Yeast species | Designation | Core structure | Branch structure | Size | Compostion | Ref. |
|---|---|---|---|---|---|---|
| Basidiomycetes | ||||||
| Bullera alba | GXM | - | D-Manp, D-GlcpA, D-Xylp | [49] | ||
| GalXM | - | - | - | - | ||
| Bullera tsugae | Mannan | - | 20–40 kDa | D-Manp | [50] | |
| Cryptococcus (Papiliotrema) flavescens | GXM | - | Manp=Xylp (GlcA, Glc?) | [51] | ||
| Cryptococcus flavus | - | - | - | 1 MDa | Man |
[52] |
| Cryptococcus laurentii | - | - | - | 4.2 MDa | Ara |
[53] |
| Rha | ||||||
| Heteropolysacch. | - | - | - | Man |
[54] | |
| Heteropolysacch. | - | - | 8 kDa | Xyl |
[55] | |
| GalGXM | Highly branched | ca 19 kDa | D-Man |
[56] | ||
| sub-branches of |
||||||
| GXM | - | D-Man, D-Xyl, D-GlcA | [57] | |||
| GXM | Xyl-[Man]-GlcA | - | - | D-Man |
[58] | |
| Glucan | (1 |
- | - | D-Glc | ||
| GXM | - | - | D-Man, D-Xyl, D-GlcA | [59] | ||
| GM | - | D-Man |
[60] | |||
| sub-branches of |
||||||
| Amylose | - | - | - | Glc | [61] | |
| GXM | D-Man |
[62] | ||||
| GalGXM | GlcA | |||||
| GM | ||||||
| Amylose | ||||||
| Crytococcus neoformans | GXM | - | - | Man |
[63] | |
| GalXM | ||||||
| GXM and others | - | - | - | Man≈Glc≈Xyl |
[17] | |
| Fuc | ||||||
| GMX | [α-(1 |
- | Man |
[64, 65, 66, 67] | ||
| Kazachstania unispora | - | - | - | - | Gal |
[68] |
| Kuraishia capsulata | Mannan | [ |
- | 360 kDa | Man | [69] |
| Phosphomannan | 635 kDa | |||||
| Moniliella pollinis | - | - | Glc |
[70] | ||
| Pseudozima sp.NII 08165 | - | - | - | 1.7 MDa | Man |
[71] |
| Rhodosporidium babjevae | - | - | Man, Glc | [72] | ||
| Rhodosporidium paludigenum | Pullulan | [ |
no | - | Glc | [73] |
| Rhodotorula acheniorum | Mannan I | - | - | [74] | ||
| Mannan II | - | - | Man |
[75] | ||
| - | - | - | - | |||
| Rhodotorula bacarum | Pullulan | - | - | - | Glc | [76] |
| Rhodotorula glutinis | - | - | - | - | Man |
[77] |
| - | - | - | - | Man |
[78] | |
| - | - | - | - | Man | ||
| - | alternate |
- | - | Manp | [79] | |
| - | - | - | 100–380 kDa | Man |
[80] | |
| Rhodotorula minuta | - | - | - | Man, Glc | [81] | |
| - | - | - | - | Glc |
[82] | |
| Rhodotorula mucilaginosa | - | (1 |
84 kDa | Glc |
[83] | |
| Highly branched | ||||||
| RESP2-A | 7.1 MDa | Gal |
[84] | |||
| Highly branched | ||||||
| Rhodotorula rubra | Rhodexman | - | 300–500 kDa | D-Manp | [50] | |
| [85] | ||||||
| - | - | - | - | Man |
[86] | |
| Sporidiobolus pararoseus | - | - | Highly branched | 7.4 MDa | Gal |
[87, 88] |
| Sporobolomyces salmonicolor | Mannan | - | Manp |
[89] | ||
| Mannan | - | - | - | Man≈Glc | [90] | |
| Sporobolomyces singularis |
Trisacch. | [ |
- | Galp, Glcp | [91] | |
| Tetrasacch. | ||||||
| Tremella sp |
- | 162 kDa (av) | Man, Xyl, GlcA | [92] | ||
| Tremella aurantialba | TAPS | - | - | 2.9 MDa | Man |
[93] |
| Tremella fuciformis | - | - | Manp |
[92, 94] | ||
| Tremellan | - | - | - | [95] | ||
| Tremella mesenterica | Tremellan | - | Man, Xyl, GlcA | [92, 96] | ||
| Pullulan | [ |
no | ca 36 kDa | Glc | [97, 98] | |
| Trichosporon asahii | GXM | [ |
- | Man |
[99, 100] | |
| GXM | - | - | - | - | [101] | |
| Glucan | - | - | - | |||
| Ustilago maydis | Glucan | Interlinked arrangements | Glc | [102] | ||
| Ascomycetes | ||||||
| Aureobasisium pullulans | Pullulan | [ |
no | 260 kDa | Glc | [43, 98, 103, 104] |
| -Glcp-(1 |
||||||
| Aubasidian | - | Glc | Elinov et al., 1975 in [43, 105, 106] | |||
| Candida albicans | GMCx | Ara |
[107] | |||
| - | - | - | ||||
| MGCx | - | [108] | ||||
| 3-Man- |
||||||
| 6- |
||||||
| (2→6)- |
Man- |
|||||
| Man- |
||||||
| Glucan | - | - | - | [109] | ||
| - | - | 300 kDa | Glc |
[110] | ||
| Glucan | - | - | Glc |
[111] | ||
| - | - | - | - | Glc; Gal; Hexosamine | [112] | |
| Monosacch. | - | - | - | Glc | ||
| Candida famata | Glucan | - | - | Hexosamine |
[113] | |
| Candida glabrata | MGCx | [108] | ||||
| 6- |
Man- |
|||||
| (2→6)- |
||||||
| Candida parapsilosis | MGCx | [108] | ||||
| 3-Man- |
||||||
| 6- |
||||||
| (2→6)- |
Man- |
|||||
| Man- |
||||||
| Candida tropcalis | GM | 74 kDa | Glc |
[111] | ||
| MGCx | [108] | |||||
| 6- |
Man- |
|||||
| (2→6)- |
Man- |
|||||
| Candida utilis | Pullulan | - | - | D-Glc | [114, 115] | |
| GalfM | - | Man, Gal | ||||
| Knufia petricola | Mannan | [→6)- |
- | - | - | [116] |
| PM | [ |
|||||
| Kuraishia capsulata | Glucan | - | - | Glc, Man, Rha, Rib, Fuc, Xyl, | [69] | |
| Mannan | D-Gal | |||||
| Rhamnan | ||||||
| Lipomyces starkeyi | Heteropolysacch. | - | - | [117] | ||
| Nakazawaea (Hansenula, Pichia) holstii | PM | 1 MDa | Manp | [118, 119] | ||
| Saccahromyces cerevisiae | - | - | - | 35–40 kDa | Glc |
[120] and our group unpublished results |
| Monosacch. | - | - | Glc | |||
| Mannan | (1 |
- | [42] | |||
| Monosacch. | - | - | - | Glc | ||
| Schizosaccharomyces pombe | GalM | Hex |
- | - | Man |
[121] |
| Hex |
||||||
| Schizosaccharomyces japonicus | GalM | - | - | Man≠Gal |
[121] | |
All the results from Basidiomycetes were obtained in planktonic cultures, in batch or fermentors. Exceptions are (i) Cryptococcus neoformans [17] that was cultivated to form a biofilm, and (ii) Tremella fuciformis which consisted of a commercial sample of this fungus fruiting body [94]. Otherwise, the results presented for Ascomycetes vary. Some were obtained using liquid cultures and others biofilms, and yet the results from Saccharomyces cerevisiae from Beauvais et al. [42] were obtained from flocs under fermentative conditions. Some abbreviations: GM, glucomannan; Glcp, glucopyranosyl; GlcA, glucuronic acid; GalM, galactomannan; GalfM, Galactofuromannan; PM, phosphomannan; Man-N, mannosamine; GXM, glucuronoxylomannan; GalXM, galactoxylomannan; GalGXM, galactoglucoxylomannan; GMCx, glucan-mannan complex; GlcNac, N-acetylglucosamine; UAs, Uronic acids; (inc), incomplete information; - unknown; no – non-existing.
Glucose is the predominant sugar residue present in Basidiomycetes and
Ascomycetes exopolysaccharides, seconded by mannose (Fig. 1A), i.e., it
exists in most of the exopolysaccharides that were characterised. Otherwise, each
one of these sugars is present in different amounts in the different polymers.
Mannose is the sugar residue present in higher amounts in Basidiomycetes’
exopolysaccharides, while in Ascomycetes, which present a larger variety of
composing sugars, including N-acetylglucosamine (GlcNac) and unspecified
hexosamines not found in Basidiomycetes (Fig. 1A), glucose is the dominant
monomer (Table 1). Moreover, uronic acids, which are key components of mammalian
ECM polysaccharides, have also been reported to occur in some yeasts’
exopolysaccharides. The basidiomycetous Cryptococcus and
Tremella spp., Rhodotorula minuta and Trichosporum
asahii (Table 1), all produce glucuronic acid (GlcA), and the ascomycetous
C. tropicalis produces uronic acids which were not yet
identified (Table 2, Ref. [88, 90, 113, 123, 124]). Additionally, the
exopolysaccharides from both groups of yeasts also largely differ in MW (Table 1), with values globally ranging from
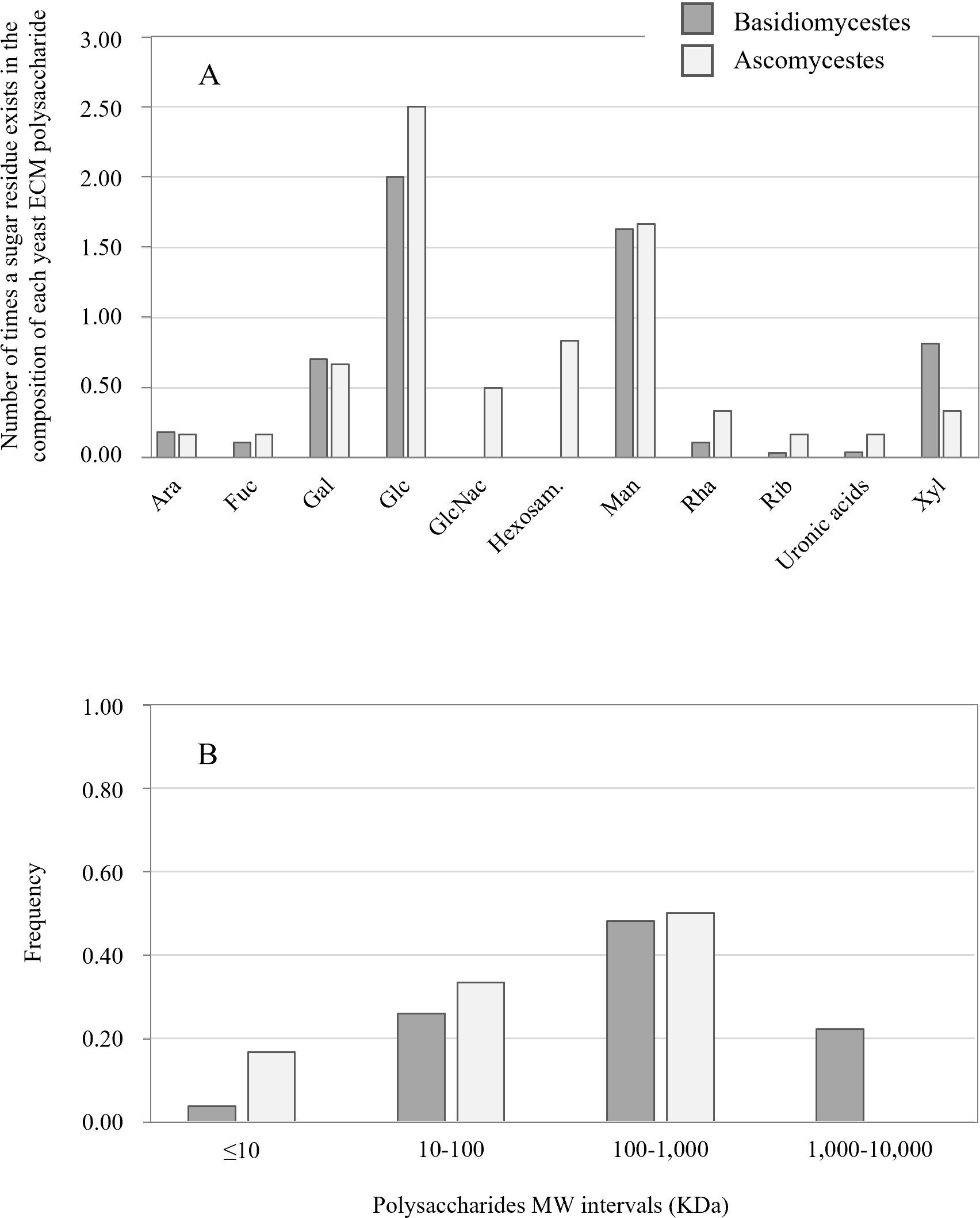 Fig. 1.
Fig. 1.Analysis of the data in Table 1 on the exopolysaccharides secreted by Basidiomycetous and Ascomycetous yeast species. (A) Number of times a given sugar is present in the polysaccharide molecule composition. (B) Distribution of the exopolysaccharides molecular mass.
| Yeast species | Carbon sources yielding exopolysaccharides maximum production | Optimal Production (g/L) | Ref. |
|---|---|---|---|
| Candida famata | Maltose | ≈2.0 | [113] |
| Candida guillermondii | ≈3.0 | ||
| Cryptococcus flavus (3 strains) | Sucrose | 2.7–3.9 | [90] |
| Debaryomyces hansenii | ≈4.0 | ||
| Pseudozyma antartica | 3.9 | ||
| Rhodotorula glutinis | 3.8 | ||
| Rhodotorula minuta (15 strains) | 1.9–2.6 | ||
| Rhodotorula mucilaginosa | 134.8 | [123] | |
| Rhodotorula sp CAH2 | 7.5 | [124] | |
| Sporobolomyces roseus (2 strains) | ≈5.0 | [90] | |
| Sporodiobolus pararoseus | Glucose | 13.1 | [88] |
Many of the compounds that were listed in Tables 1 and 2 are exopolysaccharides
produced by cells from planktonic culture conditions, in which cells are
multiplying while submersed. Exceptions are C. albicans [107, 108, 109, 111, 113], C. tropicalis [111], C. neoformans [17], Knufia
petricola [116], and S. cerevisiae [120] whose polysaccharides were
extracted from the biofilm-like cultures ECM. One single work, that of Beauvais
et al. [42], characterized the exopolysaccharides from flocs produced by
a S. cerevisiae naturally flocculating wild-type strain. Additionally,
other yeasts unmentioned in Tables 1 and 2 were reported to produce
exopolysaccharides in very low amounts (
The emerging picture from all these works suggests that Basidiomycetes tend to
produce high MW highly mannosylated and highly branched polysaccharides, while
Ascomycetes tend to produce smaller and less branched or linear highly
glycosylated polysaccharides. The branches of Basidiomycetes polysaccharides are
mostly
Two important roles of microbial ECM are those of providing a support scaffold, adherent to a living tissue or an inert surface, and contributing for efficient water/nutrients supply to all the cells in the community. The nature, concentration and solubility of the components within the ECM are under the influence of environmental factors like temperature, pH and humidity and accordingly influence these multicellular aggregates physicochemical properties. These properties include viscosity and gelation that impact directly in cells access to nutrients, and exposure to aggressors, including pharmacological drugs. Most importantly, they also impact in the ability of films and colonies to retain water and allow proper hydration to the inner cell layers. In this context, the balance between several physical features of polysaccharides commands their ability to establish intermolecular associations, which strongly translate into more or less solubility [122] and into water mobilization, and consequently to more or less stiffness of the ECM.
The solubility of exopolysaccharides is mostly commanded by their MW, as well as
their components and their molecular structure. Basic glycoside chemistry [122, 127, 128] dictate that (i) low MW polysaccharides are more soluble than high MW
ones (a polysaccharide with more than 20 oside units is supposedly insoluble in
water due to a high exclusion volume [129]); (ii) for a same MW, the more soluble
molecules are those bearing branched and irregular structure, while more
linearity or group repetitivity leads to more insolubility, (iii) the chemical
nature of monomers influences solubility. Polysaccharides containing uronic acids
are more prone to interact with water. Moreover, the existence of charged
residues, both positively or negatively, or chemical substitution, consisting of
the addition of carboxyl, sulfuric ester or other chemical groups to the sugar
monomers [122], tend to increase the interaction with water and therefore
solubility when compared to neutral molecules. Finally, (iv) the type of
glycosidic bond influences the behaviour of the polymer in solution. The most
common linkages found in ECM polysaccharides introduce diverse degrees of
flexibility to the molecules by commanding their degree of order/disorder and
their state of lower energy. Solubility is higher for (1,6), followed by (1,3)
and (1,4) glycosidic bonds. All these features are synergistically
intercorrelated, influencing each other and the ability of a certain ECM to
retain water through the solubilization of its components, in particular at a
microscale. An example of a readily water-soluble polysaccharide is that of
pullulan. In spite that it usually consists of a very large number of maltotriose
repeating units, and therefore has a large MW and repetitive structure, it is a
polydisperse molecule in which the
The retention of water at a microscale within biofilms and colonies is allowed by the ECM, therefore maintaining the hydration of the cells [131]. Together, all the physicochemical features above-mentioned concur to affect solubility, but may also contribute to the formation of hydrophobic pockets, caging water molecules, which play an important role in the ECM gelling process [132], providing lowest entropy stabilization to the ECM. Pore-scale water content is acknowledged to control the hydraulic connectivity of bacterial communities, and the retention of water in the soil at a microscale is promoted by bacterial EPS [122, 131]. Accordingly, soil bacteria contribute positively to the maintenance of moisture by producing ECM/EPS, although the actual physical influence of those polysaccharides in soil aggregation and structure is not yet fully understood [133]. Hydrophobic pockets can also be generated by the presence of rhamnose residues [127, 128], which were so far only found in a few yeasts (Table 1). Noticeably, Lipomyces starkeyi produces rhamnan [117], which is an exception in all the yeasts’ ECM so far characterized. This yeast is a probiotic, which ECM/EPS production was enhanced with the goal of prolonging effective adhesion to the gastrointestinal tract [117]. This work was based on the knowledge that the ECM/EPS from lactic acid bacteria probiotic strains, namely Lactobacillus lactis, has the physiological function of improving bacteria colonization of the mucosae, besides other important roles like those of antitumor and immunomodulating substances [134].
According to Sutherland [127, 128] the polysaccharides from microbial ECM may
adopt more or less disordered forms according to the external physical
conditions, including temperature and pH, but they tend to stabilize. The water
retention by ECM depends, namely, on the ability of the sugar molecules to
establish several types of chemical bonds with the water molecules, like London
forces established with water dipoles, or H bonds established with the more
hydrophilic sites of each osidic unit of both neutral or ionic polysaccharides
[135]. Ionic polysaccharides establish more H bonds, building hydration spheres
around each ionic group, which in turn contribute to the formation of double H
bonds with the remaining hydrophilic groups of the glucopyranosyl unit. This is
further influenced by the presence of ions in solution, like Ca
Exopolysaccharides produced by yeasts may in addition suffer a yet unknown degree of chemical substitution, e.g., sulfation. This is a common feature of the ECM exopolysaccharides from other species prompting their commercial exploitation for pharmacological applications. An example is that of ascidians which ECM provides heparan sulfate with anticoagulant and anti-cancer properties [139]. Importantly, the presence of chemical groups can easily be reverted, which transforms the ECM into a chemically and physically very dynamic environment, able to rapidly adjust to shearing forces, as well as demanding pressure, temperature or pH conditions. In biofilms formed by the filamentous fungus Aspergillus fumigatus, polysaccharides GAGs are deacetylated in order to turn them cationic, a property that was found to be required for adherence and biofilm onset [140]. Accordingly, a deacetylase complex from C. albicans that operates regularly on histones was found to mediate this yeast biofilm dispersal and drug resistance [141]. Notably, in biofilm-like mats of S. cerevisiae, the purified ECM polysaccharides induced metachromatic shift, which is suggestive of chemical substitution, corroborated by their separation by diaminopropane agarose gel electrophoresis [120]. Although the authors clearly associated these properties with the deletion of the O-acyltransferase-encoding GUP1, the subject was not further explored.
The ECM though contains large amounts of other molecules [127, 128], notably proteins that will certainly not only functionally contribute to a high microenvironmental diversity, but also contribute to shape the physicochemical properties in every niche. Different polysaccharides interacting with several amounts of different proteins, as well as lipids [107] and e-DNA [142, 143] will undoubtedly generate a dynamic environment with space-specific properties, including rigidity/strength and sheer stress resistance. The ECM mechanical properties must therefore be envisaged as multifactorial and sensibly a synergistic interaction of complex microsystems of matter and energy transfer.
Sheer stress is caused by fluid flow, which is a common feature to microbial
biofilms adhering to ship’s hulls, pipes, catheters or blood vessels, where it
translates into localized dynamic combinations of tensile and compression forces
to the microbial cells. In animal cells the transduction of mechanical stress
into chemical signals (mechanotransduction) is specifically conducted through
protein connections between the cell and the ECM (mechanotransduction foci)
[144]. Although the correspondent signalling pathways are still mostly unknown,
mechanotransduction does cause changes in gene expression [145]. A number of ion
channels have been shown to be gated by mechanical stimuli and playing putative
roles in mechanotransduction, like the degenerins or the Na
Another way that yeasts display to sense mechanical stress occurs through
osmosensors, the cell surface mucins Hkr1 and Msb2 and the transmembrane
Sln1/Sho1. These primordially sense a
One single protein has so far been clearly associated with yeast’s ECM
physicochemical properties, the S. cerevisiae O-acyltransferase
Gup1 [156]. Biofilm-like mats of
Other proteins that could be related to yeast ECM construction and remodelling
are the yeast counterparts of the human ECM metalloproteases. These enzymes have
a prominent role in human tissues ECM remodelling, which is associated with
normal physiology and pathology, as well as embryonic development, wound healing
and metastasis [160, 161]. They regulate tissue density and stiffness, by
modulating the size, structure and amounts of ECM components under the control of
major regulatory signalling pathways (namely TGF
The structure and composition of microbial exopolysaccharides as well as their production yield depend on features of the culture cultivation conditions and the medium composition. Some of the environmental requirements mentioned in the literature for the production of exopolysaccharides by yeasts regard general cultural specificities of yeasts. These include cultivation temperature, aeration and initial pH. Many yeasts species grow optimally at temperatures ranging 25–30 °C. Fewer species are known prefer lower or higher temperatures, like the human pathogens which optimal temperature for growth is 37 °C. Although many of the production conditions reported fall within that temperature range, the optimal temperature for yeast growth does not necessarily match the optimal temperature for the production of exopolysaccharides by the same yeast. For example, R. acheniorum optimal growth temperature is rather exceptionally at 20 °C but production is maximum at 26 °C [74].
Another relevant factor is that of O
The initial pH of a culture, for most of yeast common media naturally lie around
5–6. During yeast population increase it will rapidly achieve values as low as
pH 1–2. Several works mention that an initial pH of 4–5 is mandatory for
exopolysaccharide production [169, 170, 171]. Nevertheless, this is probably not a
generalizable requisite [41, 165, 170]. For example, Rhodotorula
mucilaginosa was reported to produce most at initial pH 6.0 [83]. Although
different optimal pH values have been reported [170], the production of
exopolysaccharides could be related with the very low pH achieved at the end of
culture growth. Cho et al. [80] showed that Rhodotorula glutinis
only produced exopolysaccharides if the pH of the culture was allowed to lower
beyond pH 2.0. Identical claim was made by Pavlova and Grigorova [167] for the
production of exopolysaccharides by R. acheniorum. This is concomitant
with the realization that exopolysaccharide production only occurs during
pre-stationary/stationary growth phase, i.e., when pH is lowest. In that
phase, according to several authors [75, 80, 165, 167], the nitrogen source
should be exhausted or near exhaustion, but the carbon source should still exist,
a condition that is only achieved if the C/N ratio of the culture media is very
high. This agrees with so many other authors claiming the need for a very
disproportionate C/N molar ratio like the
The possible reasons for the pH-N link are well documented in S.
cerevisiae. Although one should be cautious to generalize the information
available for Saccharomyces spp to less-conventional yeasts,
specifically because their metabolism is often respiratory and not
respiro-fermentative, the proteins involved in ammonium transport are extremely
conserved throughout plants, animals and microbes of diverse types [172, 173].
This way, in S. cerevisiae, at ammonium concentrations
In yeasts, the performance of H
In line with the above, the chemical composition of the culture media is probably the most demanding condition for optimal exopolysaccharide production. The yield is influenced not only by mineral versus organic nitrogen sources but also by carbon source. Yeasts produce exopolysaccharides while growing on many sugars, hexoses, pentoses and disaccharides, which could mean that yeasts produce exopolysaccharides in basically every sugar they can metabolize. Yet, most studies consider sucrose as the ideal carbon source promoting the best yields [113, 165, 168]. Importantly, the carbon and nitrogen sources can affect not only the yield but also the composition of the exopolysaccharide [113, 165, 183]. Grigorova et al. [75] showed that R. acheniorum MC cultivated in media with different carbon sources presented changes in yield, which was highest when cells were cultivated on sucrose, as well as in the chemical composition of the exopolysaccharides. The relative percentages of the composing sugar monomers varied, particularly those of fucose (35% variation between results obtained using glucose, galactose, sucrose or dextrose syrup, Sirodex®), and glucose (31% variation in the same media), although mannose and glucose kept being the most represented monomers. The possibility should therefore be considered that ammonium salts might also interfere with the composition of exopolysaccharides, and/or their structure, according to the fact that in yeasts, several major cross-talking signalling pathways respond to ammonium exhaustion in concert with carbon source availability.
Most of the yeast species which ECM components have been identified (Table 1)
have no information available on what enzymes/genes are involved in their
production and how they are regulated. An exception is that of pullulan
production by A. pullulans, although pullulan is mostly secreted during
planktonic growth. Pullulan is a linear
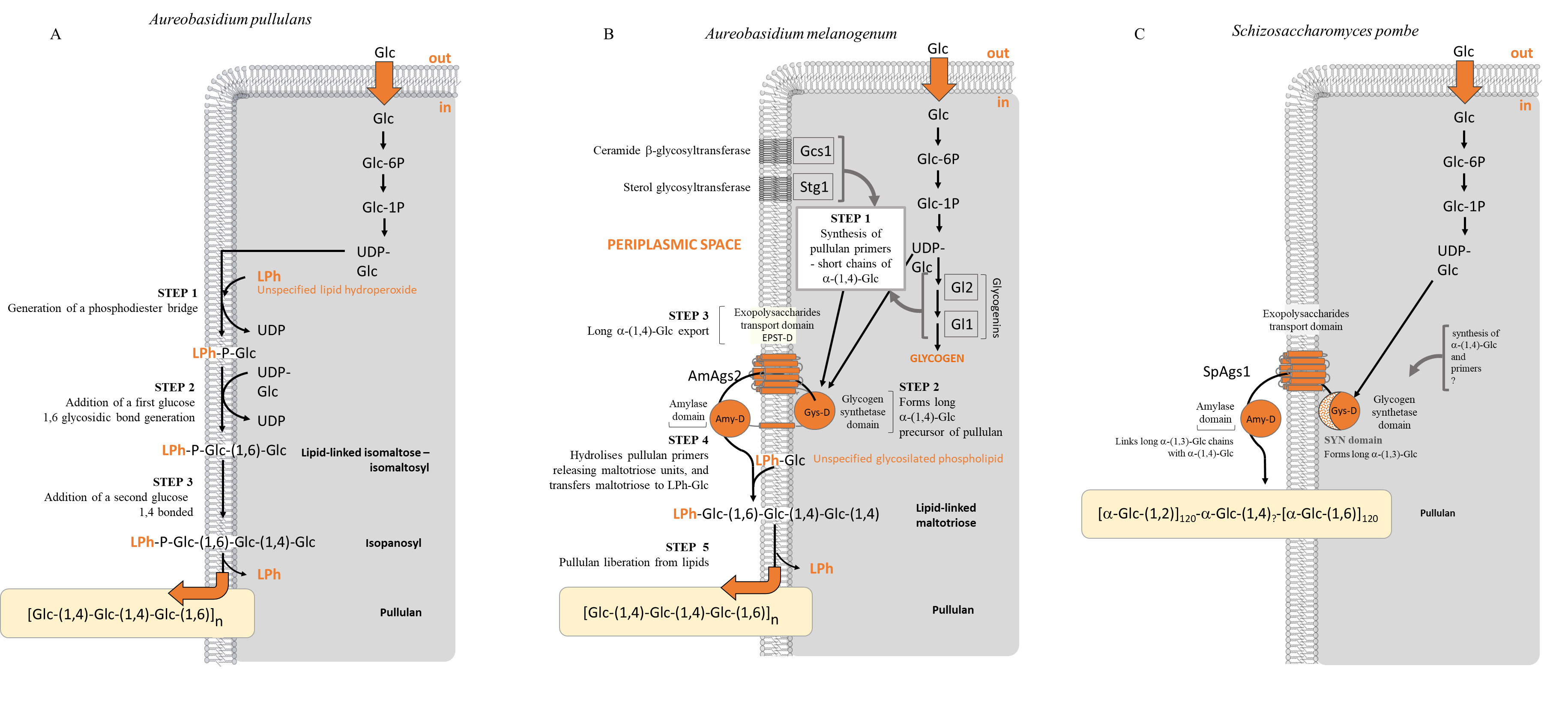 Fig. 2.
Fig. 2.Metabolic pathways proposed for the production of pullulan. (A) Aureobasidium pullulans, (B) Aureobasidium melanogenum and (C) Schizosaccharomyces pombe.
Unlike the cases above, the ECM from S. cerevisiae was described to
contain high amounts of glucose monosaccharides [42, 120] which origin or
biological purpose is still unknown. Moreover, Beauvais et al. [42] also
reported the presence of
It is possible that some interchange between
The ECM from biofilms formed by the human pathogen C. albicans also
displays
On the other hand, previous work had attempted the identification of the genetic
determinants from C. albicans genome that were associated with the
positive and negative regulation of ECM deposition in biofilms [198, 202, 203]. A
few genes, classically ascertained to the cell wall mannan and glucan pathways,
were identified in this yeast which disruption impairs both the deposition of
mannans and glucans in the ECM and appear to be active and critical for the
assembly of the ECM [204]. Some of these genes are under the negative control of
the Zap1 transcription factor. From these, only two of them, encoding the
glucoamylases Gca1 and Gca2, have actually been implicated in the formation of
the ECM [202, 203]. Although the exact mechanism through which these enzymes
might operate is unknown, they have been suggested to promote the release of
soluble
Two other proteins were suggested to govern the levels of
Pullulan production by A. pullulans has been reported as being best on sucrose, not glucose. The underlying reason comes from that pullulan production enzymes are repressed by glucose at high concentration [188]. It is though not certain how this occurs. Several authors suggest that the global glucose repressor Mig1 is involved, based on that the deletion of this gene homologue in A. melanogenum increased pullulan production. This may be less straightforward than it seems, since some strains of A. melanogenum were shown to produce pullulan in media with high amounts of fructose and glucose as it occurs in honey [205]. Authors associated this apparent constitutive derepression with the high osmotic tolerance of the strains and their concurring ability to produce and accumulate large amounts of the osmolyte glycerol, although no more data or detailed explanation are available.
Moreover, the requirement for ammonium exhaustion for highest production of exopolysaccharides strongly suggests the involvement of major N-dependent signalling pathways, which control bulk physiological responses including yeast cell wall remodelling and filamentation [206, 207]. Being exopolysaccharides a pre-requisite for the establishment of a biofilm, in which cells often differentiate into hyphae or pseudo-hyphae, it is plausible that those pathways are somehow involved.
Cells from all kingdoms of life produce and secrete extracellular vesicles as a
means to excrete proteins and other macromolecules. Additionally, all cells that
are walled require a process that allows the passage of macromolecules through
the cell wall barrier. That is the case of yeasts. Extracellular vesicles (EVs)
were found to be released by yeast human pathogens, including C.
albicans [154, 208, 209], as well as the model yeast S. cerevisiae [210, 211]. Their role has been firmly associated with the biogenesis of ECM in
C. albicans biofilms, in particular with the deposition of matrix
exopolisaccharides [154]. Mutants defective in proteins involved in late
endosome sorting complexes required for
transport (ESCRT proteins) displayed reduced matrix exopolysaccharides
and biofilm-forming ability, and consequently increased sensibility to
fungicides. These defects were all fully complemented by externally adding EVs
isolated from the wt strain biofilms [154]. These EVs are specific of
biofilms, differing from others that are released by planktonic cell cultures. In
such cultures, both S. cerevisiae [210] or C. albicans [154]
secrete two distinct sized populations of vesicles, one clearly smaller than the
other, although the sizes were different from one species to the other and from
one strain to the other within the same species. C. albicans biofilms
only present the smaller EVs. The time course of its formation and release
followed biofilm-development, but their composition is slightly different from
the biofilm ECM. Mannan/glucan proportion in EVs is approximately 5 times higher
than in the ECM [154], essentially due to a much higher presence of
The
The chromatographic analysis of the proteome of yeasts ECM revealed this is an extremely proteinaceous environment [107, 157, 216]. S. cerevisiae ECM contained 614 well-identified proteins that were not found in the control supernatant of the same strain grown in liquid medium [157], which have well-characterized roles allowing their distribution into several functional groups. The largest group aggregated enzymes from several central metabolic paths, including the full sets from glycolysis, fermentation and gluconeogenesis [157]. The presence of these two metabolically antagonistic sets of enzymes suggests that biofilms harbour two metabolically different populations of cells [217]. The second largest group includes numerous enzymes from the HSP70 family involved in the synthesis, folding and degradation of other proteins [157].
A crucial similarity is found between the proteomic analyses, that which studied the ECM extracted from S. cerevisiae biofilm-like mats [157], and which that studied this same yeast planktonic cultures EVs [210]. The most abundant functional class of EVs’ proteome contained many enzymes from metabolic processes including carbohydrate metabolism, followed by proteins involved in other proteins biosynthesis, degradation and transport [210]. This remarkable resemblance suggests that the proteome within the yeast ECM indeed might be, at least in part, delivered by EVs. Accordingly, these vesicles also contained glucanases and glucanosyl transferases which are probably involved in cell wall remodelling [210, 218]. These might also be involved in the synthesis of the ECM polysaccharides. Moreover, S. cerevisiae secretes two sizes of EVs [210], the smaller of which specifically carries enzymes from gluconeogenesis: Fbp1 (fructose 1,6-biphosphatase), Mdh1 (malate dehydrogenase), Icl1 (isocitrate lyase), Pck1 (phosphoenolpyruvate carboxykinase) as well as Tdh1/2 (glyceraldehyde 3-P dehydrogenase) [209, 211]. This specificity agrees with the existence of two populations of cells, each secreting one type of EVs.
Otherwise, the filtered supernatant of Saccharomyces planktonic cultures presented a very reduced proteome (app. 87% less proteins) [157], suggesting that in planktonic cultures most proteins are secreted to the extracellular medium via EVs and not individually, which is consistent with the fact that many of the proteins identified in association with EVs are predicted to harbour an ER and secretory pathway signal peptide instead of a secretion signal [210]. The same was observed with the proteins from EVs from C. albicans [208]. These authors showed that in this yeasts’ planktonic cultures, the proteome from EVs harboured 40% proteins that are predicted to not have a signal peptide, while in the same cultures’ vesicles-free supernatant, the number of proteins actually possessing a secretion signal was 90%. The hypothesis that the yeast ECM proteome is secreted via EVs is consistent with Thomas et al. [219] describing that most of the proteins they identified in C. albicans biofilm ECM had no secretion signal. For this reason, Gil-Bona et al. [208] suggested that C. albicans has two different secretory pathways, a classical one requiring an export signal peptide, which secretes mostly cell wall remodelling and maintenance enzymes (49%), followed by hydrolases and adhesins (21%), metabolism enzymes and virulence factors, and a second non-classical protein export system, not requiring an export signal peptide that occurs through the secretion of EVs [208]. The proteome in the biofilms ECM must derive from either or both secretion systems. Actually, the possibility that EVs are part of the yeast ECM derives from the fact that a small fraction of lipids is found in C. albicans biofilms ECM (15% according to Zarnowski et al. [107], which composition (89.2% of neutral lipids and 85.6% of the polar lipid fraction consisting of phosphatidylethanolamine (PE) [107]) is identical to that of this yeast plasma membrane (90% neutral lipids and 70.0% of the polar lipid fraction consisting of PE [220]). Although membranes’ lipid composition may vary according to the strain, yeast-hyphae differentiation and environmental cues [209, 221], this remains a prominent similarity, consistent with the EVs deriving from plasma membrane as previously suggested by Vargas et al. [209].
As in Saccharomyces cerevisiae, also the proteome from C. albicans biofilms ECM was characterized [107, 109, 219]. They identically included proteins that could be ascertained to 485 different proteins [107], whose most abundant group (351 proteins) corresponded to enzymes from carbohydrate, amino acids and energy metabolism pathways. The biggest of these groups was that from carbohydrate metabolism, with 177 enzymes from TCA cycle, pyruvate metabolism and, importantly, glycolysis/gluconeogenesis. Moreover, the ECM from C. albicans, identically to that of S. cerevisiae, also presented a set of HP70 family, as well as the enzymes above mentioned (Fks1, Bgl2, Xog1 and Phr1), involved in polysaccharides synthesis and remodelling [109, 219].
The presence of so many proteins with well-established intracellular functions, in particular the entire sets of enzymes from main metabolic pathways raises important questions. They could ensure metabolism extracellularly, which could be supported by the presence in the ECM from both S. cerevisiae and C. albicans of large amounts of monosaccharides (Table 1). Accordingly, the bacterial biofilm matrix was suggested to correspond to an external digestion system promoted by the immobilization of enzymes which may participate in general metabolic reactions as well as in the very remodelling of the ECM structural components [40]. Alternatively, those enzymes could provide other presently unknown functions as moonlighters. This terminology was applied to several yeast enzymes that perform utterly distant tasks than the one they were primarily described for. As an example, the glycolytic and/or gluconeogenic enzymes fructose 1,6-bisphosphate aldolase, glyceraldehyde-3-phosphate dehydrogenase, phosphoglycerate kinase, phosphoglycerate mutase, enolase and alcohol dehydrogenase, were found to bind fibronectin, laminin, plasminogen or to complement regulators with high specificity, and were identified as important virulence factors in C. albicans [222, 223, 224, 225]. Although S. cerevisiae does not behave identically to C. albicans, all those proteins were equally identified in that yeast biofilms ECM [157]. No study was done so far characterizing the metabolome of yeasts’ ECM. Otherwise, the biofilms of C. albicans were shown to be characterized by yeast cells displaying low rates of TCA cycle and mitochondrial activity as well as high levels of several amino acids and glycerol [226], suggesting a tendency for conservation of energy together with the accumulation of protective metabolites [227]. Identically, the cells of C. neoformans within biofilms were reported to shift from TCA cycle towards pyruvate utilization in particular through fermentation [216]. Whether the massive secretion of metabolism enzymes relates with these changes in metabolism will have to be explored in the future.
Human pathogenic biofilm-forming yeasts include many Candida species.
These include C. glabrata, C. parapsilosis, C.
dublinensis, C. tropicalis or C. auris, and are often referred
as NCAC species (non-Candida albicans Candida species) [228].
In most individuals with a healthy immune system, these yeasts are harmless
commensal populations existing in equilibrium with other members of the
microbiota and colonizing many areas of the human body, especially mucosae of the
gastrointestinal and genitourinary tracts. However, disturbances caused by shifts
in, namely, pH, O
C. albicans infections are associated with the proliferative colonization through the formation of highly structured biofilms [231]. Within these, yeast cells can differentiate into pseudo-hyphae, but mostly they differentiate into penetrative invasive true hyphae [232]. This yeast-to-hyphae transition is required for infection to develop and commands virulence [233, 234, 235, 236]. Importantly, it is also required for the formation of biofilms [5, 203]. It derives from extensive changes in gene expression [237, 238, 239, 240, 241] accompanied by high levels of horizontal gene transfer [242, 243, 244]. The genes that govern C. albicans biofilm formation are not limited by the above-mentioned PHR1, XOG1, BGL2, as well as ZAP1 and RIM101 transcription factors [109]. They cover a broad diversity of functional categories, including, namely, the yeast-to-hyphae morphological shift and filamentation growth [198, 202, 203].
Importantly, some of those genes encode drug efflux pumps like MDR1 [203], contributing to the biofilm increased resistance to antifungal drugs.
Concomitantly, C. albicans biofilms are mostly resistant to the
currently available antifungal drugs. Several mechanisms underlie this
resistance. A first consists in the increased expression of drug efflux pumps,
including CDR1, CDR2 and MDR1 [239, 245, 246]. These
genes become upregulated within the first few hours of yeast/surface contact and
remain that way throughout biofilm development, whether or not an antifungal drug
is present [239, 247, 248, 249, 250, 251] A second mechanism corresponds to the existence of
persistent cells in the biofilm. These are metabolically quiescent cells that
resist to a range of drug concentrations otherwise lethal to planktonic cells
[252, 253]. They can revert back to metabolically active cells and contribute to
the reformation of the biofilm once the antifungal drug has been removed [254]. A
third mechanism is the physical barrier to drug penetration provided by the ECM
itself [111, 112, 255, 256, 257]. The
Another human pathogenic biofilm-forming species is C. neoformans. This
is an opportunistic yeast-like fungus that causes several pathologies
(Cryptococcosis) in mammalian hosts, mainly if they are immunocompromised, from
which the most severe is meningoencephalitis [259, 260]. C. neoformans
distinguishes from Candida species namely because it is not able to form
pseudo-hyphae. This species has a very complex life cycle [261]. According to
several host and environmental cues, it may encompass yeasts or hyphae, haploid,
diploid or polyploid cells, as well as chlamydospores, basidium and
basidiospores. It may multiply endlessly as a heterothallic budding yeast, which
may belong to either of two mating types a and
C. neoformans unique morphological feature is a polysaccharide capsule involving the yeast cells [262], which is the most important virulence factor during infection [263], since non-encapsulated mutants are avirulent [264]. It is also important in the dispersal of this pathogen in the environment [265]. The capsule is composed primarily of two polysaccharides, glucuronoxylomannan (GXM) and glucuronoxylomannogalactan (GXMGal; previously called galactoxylomannan) [263, 266, 267]. The capsule confers protection to the yeast during infection as it has been shown to inhibit phagocytosis [268, 269], but also has a role in virulence, since both GXM and GXMGal are potent inducers of apoptosis [270, 271, 272]. Besides, these polysaccharides also induce an immunological unresponsiveness by, among others, inhibiting the production of antibodies [273, 274]. In addition, the capsule presents a small proportion of mannoproteins [275, 276], as well as, importantly, hyaluronic acid (HA) [277, 278]. The presence of HA apparently facilitates the adhesion of the fungal cells to the tissues [277] and its interaction with the human cells HA receptor CD44 initiates the events of invasion process in the human brain microvascular endothelial cells [278, 279]. The HA receptors CD44 and RHAMM to a lesser extent, are present in human cells plasma membrane lipid rafts, which absence, inactivation or displacement cause a very significant decrease in Cryptococcus infection ability [278]. Although there is no direct implication of HA, or any other capsule component, in the development of biofilms by C. neoformans yeasts, it appears that the capsule is also important for that purpose, since acapsular strains do not develop biofilm [280]. Accordingly, it cannot be ignored that CD44 in response to HA has been associated with uncontrolled cell proliferation as well as invasiveness and migration in cancer [281, 282, 283].
Similar to Candida yeasts, also C. neoformans cells within biofilms are less susceptible to antifungal agents as well as to the host immune system [284, 285]. A cryptococcal biofilm consists of a complex network of only yeast cells, enmeshed in a substantial amount of ECM [284, 285]. Its adhesion process was shown to be mediated by the release of one of the capsule polysaccharide, initiating the creation of the biofilm ECM [280, 286]. Whether this is GXM, the same that C. neoformans releases into the colonised tissues, is not known, although both capsule and biofilm have GXM [17]. Accordingly, specific antibodies against GXM inhibited biofilm formation [286]. The capsule could thus act as a facilitator for the biofilm initiation.
The secreted GXM, also designated exo-GXM, has important roles in the development of Cryptococcosis, which include the formation of biofilm and concomitantly protection of the fungal cells against anti-fungal drugs [286, 287], given that exo-GXM-deficient mutant is inefficient in biofilm formation and is avirulent [288]. Exo-GXM accumulates in cerebrospinal fluid, serum and tissue of infected patients [289, 290, 291] and presents several immunomodulatory properties (reviewed in [292]), including that of inhibiting phagocytosis. This occurs probably because GXM is polyanionic. This feature has been associated with the possibility of GXM causing an electrostatic repulsion, preventing host immune cells from interacting with and eliminating fungal cells [293].
S. cerevisiae is the best-studied yeast ever. It is considered a model for human biology due to a high degree of evolutionary conservation of many proteins and molecular processes between humans and yeast. Besides aggregating onto colonies [294] or biofilms/mats [295, 296, 297], S. cerevisiae can also aggregate onto flocs [298]. All provide survival advantages due to the cooperative behaviour of cells [299, 300], and the establishment of common protective mechanisms [301], namely the production and accumulation of the ECM [301, 302]. S. cerevisiae multicellular aggregates of all types produce ECM [42, 303, 304, 305]. This, as for other yeast species, is crucial to prevent desiccation [306], while providing protection against xenobiotics [304], and ultimately a nutrient reservoir [217], as discussed above.
Yeasts essentially develop two major types of colonies. Smooth colonies are formed by most laboratory or domesticated strains [303]. They are composed of tightly attached cells unable to invade semisolid supports, such as agar, and do not harbour any ECM [307, 308]. Alternatively, cells on the surface of the colony pack together forming a protective thin layer of tightly connected cells [303, 307]. Structured colonies formed by wild strains are also designated in the literature as colony biofilms [303]. These are composed by an aerial part, formed mostly of oval shaped cells above the agar, and the subsurface “roots”, formed by pseudohyphae invading the agar [304, 306]. Cells forming colony biofilms are connected by extracellular fibres [303, 304, 309] and are embedded in an abundant ECM which functions as a nutrient reservoir and as a low-permeability barrier, blocking the contact of compounds diffusing through the agar with the cells [304].
S. cerevisiae colonies also differ in size. In fact, they can grow disproportionally on low-density (0.3%) YPD agar, becoming giant colonies that can achieve almost the full size of a Petri dish which are also called mats [295, 310]. These resemble biofilms in terms of spatial organization and properties. They contain distinct cell populations: a central hub composed of adherent cells, from which emanates a network of “cables” that culminates in the formation of spokes bordered by a rim of non-adherent cells [295]. Analogously, Varon and Choder [311], reported that starving cells in the centre of colonies were covered with ECM and united by continuous extensions of the mucous substance forming fibrils, in increasingly higher amounts when cells were purposedly starved. The nature of these fibrils is still unknown, as it is whether they allow actual connection /communication between the cells. Besides forming concentric gradients of nutrients (glucose) and pH [296], which can influence cell properties within the hub and rim of a colony, these also form gradients of ammonia. These signal and confine the apoptotic death of the cells to the centre of the colony [312, 313], thus contributing to the enlargement of the cell population with healthy, reproducing cells at the periphery.
The S. cerevisiae ECM regulates the transition from a relatively
slow-growing colony to a rapidly expanding mat [313, 314], as well as cell-cell
communication, cell-cell and cell-substrate adhesion, and spatial
heterogeneity-mediated cell morphology, growth and multiplication [304, 315, 316]. These processes are accompanied by considerable changes in the expression
of many genes, the most studied of all being the adhesin/flocculin (Flo) proteins
[297, 303, 317, 318, 319, 320, 321]. These are glycoproteins, similar to the adhesion proteins
from other yeasts, including human pathogens such as C. albicans and
C. glabrata [316, 319, 320, 322, 323]. Flocculins share a common
structure [323, 324, 325] but different functions [318, 319], and contribute to the
development of different types of flocs [324, 326]. The most well-known gene
associated with cell adhesion to agar and plastic surfaces is FLO11 [318]. This gene encodes a GPI-anchored cell surface flocculin that contains a
specific domain responsible for cell-surface adhesion [316, 317] and required for
the yeast differentiation into pseudohyphae [327, 328]. It is a key protein,
involved in the formation of structured colony biofilms [329],
flor [330], biofilms [331], and mats [295, 332]. Concurrently, any
flo11
Several other genes besides FLO genes participate in the establishment of S. cerevisiae multicellular forms. The gene BTN2 expresses a v-snare interacting protein involved in intracellular protein trafficking [341]. When this gene was deleted in a Sacharomyces cerevisiae flor strain, the adhesion to plastic was impaired, but the yeast formed a consistently larger biofilm with concomitant increased transcription of FLO11 [342]. Besides, HSP12, which encodes a heat-shock protein, was also reported to have a role in cell adhesion, since the mutation or deletion of this gene resulted in the failure to form a biofilm [343]. Furthermore, Hsp70 molecular chaperones play key roles in mat formation with the assistance of the nucleotide exchange factors Fes1 and Sse1 and the Hsp40 family member Ydj1 since the disruption of these cofactors completely abolished mat formation [344]. Glycosylated cell surface proteins that contribute to the cell surface hydrophobicity, like Ccw14, also contribute to S. cerevisiae biofilms. Deletion of CCW14 resulted in the decrease of the weight of biofilm formed, as well as of the cell adherence to polystyrene [345].
The study of yeast ECM components from planktonic cultures or from biofilms have very distinct purposes. One is identifying economically viable biotechnological processes for pharma or food industries, and another is identifying the molecular players governing the establishment and the biology of yeast biofilms, mainly those that are formed by pathogens. The exopolysaccharides that are produced in planktonic cultures meet the first goal, but cannot be considered to have the same physiological roles as the ones produced in the context of biofilms or other large multicellular aggregates. Therefore, the actual knowledge on the yeast’s ECM composition, supporting multicellular communities and promoting cell differentiation, communication, spatial organization and controlling replication and death, is much scarcer. To this situation also contribute the technical difficulties associated with obtaining substantial ECM amounts, compatible with common analytical procedures, and the use of very different ways to culture the cells, which do not replicate one another. In this regard some efforts are noticeable [309, 310, 345, 346], but much remains to be done.
Each yeast species’ ECM has a characteristic glycoside composition. Polysaccharides differ in size, structure and composing monomers, and in the majority of the studied cases they are chemically and structurally different from those found in the cell wall. Additionally, in some cases, the ECM has been reported to contain large amounts of monosaccharides, which origin remains obscure in view of the often-abiotic biofilm-supporting surroundings. Another important feature of yeasts’ ECM consists in the existence of a large proteome, that was only fully characterized in S. cerevisiae and in C. albicans. The two proteomes are very similar. Noticeably both yeasts share the presence of full metabolic sets from main metabolic pathways like glycolysis, gluconeogenesis, TCA and respiration. The results from both studies also concur in pointing to the putative involvement of acetic and citric acid in the ECM formation, but critical information is absent. For the time being none of these components has been associated to specific functions or physicochemical properties of the ECM, but rather to pharmacological bioactive properties sustaining their possible commercialization [41]. The matrixome of yeast communities needs to be extensively addressed if answers to fundamental questions are to be answered.
The most fundamental of all questions relates with the acquisition of obligatory multicellularity. Yeasts display facultative multicellularity, in the sense that they may form multicellular aggregates either by clonal or aggregation strategies [347]. They may not split after mitosis, remaining ligated [348], or they modulate their adhesive properties by changing the expression of the FLO genes (reviewed by [347]) and stick together. Experimentally, it was possible to turn this facultative into an obligatory multicellular aggregate (called snowflake yeast), which grows spatially in a geometric coherent way, where cells diversify roles and interact socially, and replicates through the formation of multicellular propagules [348, 349]. Does ECM play any role in this behaviour, mirroring the higher eukaryotes cell-ECM mutual regulation? How does a population of millions of yeast cells which are not mobile achieves the orchestrated collective behaviour, spatially oriented? Is ammonia the single communication path between yeast cells or is the ECM a central hub for more complex molecular signals identical to higher eukaryotic tissues? These questions sum to many others underlying all that was summed up in this review. A deeper understanding of the ECM produced by yeasts and its components, as well as its biology, genetics and molecular effectors, and how they change according and react to external stimuli, will be instrumental to a deeper understanding of yeast biology, supporting their many new utilizations in perspective, and the continuous utilization of yeasts as a scientific fundamental model for Biology.
CL—conceptualization, bibliography review and manuscript writing and editing; CS—bibliography review and manuscript writing and editing. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This work was supported by the “Contrato-Programa” UIDB/04050/2020; funded by national funds through the FCT I.P.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.