




 , Podoprigora Irina Viktorovna 1, Anyutoulou Kitio Linda Davares 1, Kezimana Parfait 2, Smolyakova Larissa Andreevna 1, Hippolyte Tene Mouafo 3, Manar Rehailia 2, Yashina Natalia Vyacheslavovna 1, Smirnova Irina Pavlovna 4, Irma Aurelia Monique Manga 5, Das Milana Sergueïevna 1
, Podoprigora Irina Viktorovna 1, Anyutoulou Kitio Linda Davares 1, Kezimana Parfait 2, Smolyakova Larissa Andreevna 1, Hippolyte Tene Mouafo 3, Manar Rehailia 2, Yashina Natalia Vyacheslavovna 1, Smirnova Irina Pavlovna 4, Irma Aurelia Monique Manga 5, Das Milana Sergueïevna 11 Department of Microbiology and Virology, Institute of Medicine, RUDN University, 117198 Moscow, Russia
2 Department of Agrobiotechnology, Peoples Friendship University of Russia (RUDN University), 117198 Moscow, Russia
3 Centre for Food and Nutrition Research, Institute of Medical Research and Medicinal Plants Studies, 7210 Yaounde, Cameroon
4 Department of Biochemistry, Institute of Medicine, RUDN University, 117198 Moscow, Russia
5 Higher Institute of Medical Technology, 188 Yaounde, Cameroon
Academic Editor: Jinwei Zhang
Abstract
Background and Aim: Antibiotics’ resistance is the leading cause of complications in the treatment of urinary tract infections. This study aimed to screen the antimicrobial potential of 8 plants from Cameroon against multi-resistant uropathogenic (MRU) bacteria and to investigate their antibioresistance reversal properties. Method: Bioactive compounds were extracted from leaves of Leucanthemum vulgare, Cymbopogon citratus, Moringa oleifera and Vernonia amygdalina; barks of Cinchona officinalis and Enantia chlorantha barks and seeds of Garcinia lucida and leaves and seeds of Azadirachta indica using water and ethanol as solvents. The extracts were tested against Escherichia coli ATCC 25922, Staphylococcus aureus ATCC 6538 and Candida albicans 10231 using the well diffusion and the broth microdilution methods. The antibiotic-resistance reversal activity was assessed against selected MRU bacteria. The phytochemical composition and the elemental composition of the most active extracts were assessed respectively using HPLC-MS/MS and X-ray fluorescence (XRF) spectrometry. Results: Among the most active plants, in decreasing order of antimicrobial activity we found ethanolic (EE) and aqueous extracts (AE) of E. chloranta bark (ECB), EE of L. vulgare leaves and G. lucida seeds. The best synergies between common antibiotics and extracts were found with EE-ECB which well-modulated kanamycin nitrofurantoin and ampicillin. All the compounds identified in EE-ECB were alkaloids and the major constituents were palmatine (51.63%), columbamine+7,8-dihydro-8-hydroxypalmatine (19.21%), jatrorrhizine (11.02%) and pseudocolumbamine (6.33%). Among the minerals found in EE-ECB (S, Si, Cl, K, Ca, Mn, Fe, Zn and Br), Br, Fe and Cl were the most abundant with mean fluorescence intensities of 4.6529, 3.4854 and 2.5942 cps/uA respectively. Conclusions: The ethanol extract of the bark of E. chlorantha has remarkable, broad-spectrum antimicrobial and contains several palmatine derivatives.
Keywords
- medicinal plants
- antimicrobial
- synergy test
- antibiotics
- multiresistant
- uropathogenic bacteria
Antibiotic resistance is defined as the ability of a bacteria to resist the inhibitory or destructive activity of an antimicrobial to which it was not resistant [1, 2, 3]. This adaptation phenomenon is mainly due to the enzymatic degradation of antibiotics by bacteria, the modification of the antibiotic target, the change in membrane permeability, and alternative metabolic pathways [4].
Antibiotic resistance is a public health problem with an impact on human and animal health, agriculture, the economy and the environment [1, 2, 3, 4]. Recent estimates have shown that antibiotic resistance is responsible for 700,000 annual deaths worldwide, 230,000 of which have resulted from multidrug-resistant bacteria [2]. The World Health Organization estimates that if nothing is done to address this problem, drug-resistant diseases may cause 10 million deaths each year by 2050 and damage to the economy as catastrophic as the 2008–2009 global financial crisis [2]. Furthermore, economically (linked directly or not to agriculture and animal breeding), antimicrobial resistance could force up to 24 million people into extreme poverty by 2030 [2]. The search for new antimicrobials is therefore essential to address this worldwide public health issue [1, 2, 3, 4]. This situation affects all areas requiring the use of antibiotics including the management of diseases such as urinary tract infections (UTIs).
UTIs are very common infections in the human population (especially in women) and can be defined as any infection, commonly of bacterial origin, which occurs in any part of the urinary system [3]. Nowadays, UTIs are serious public health issues and are responsible for nearly 150 million disease cases every year worldwide [3]. Most UTIs (80–90%) are caused by Escherichia coli while other germs like Staphylococcus saprophyticus, Pseudomonas aeruginosa, Staphylococcus aureus, Klebsiella pneumoniae, Proteus mirabilis, Acinetobacter baumannii, Streptococcus, and Enterococcus faecalis are rarely involved [5].
Resistance to antibiotics has made these infections more difficult to treat. Medicinal plants are among the most promising solutions to address this problem and each year, studies carried out in the 4 corners of the globe are intended to exploit their antimicrobial potential. In this context, the Cameroonian flora, known for its abundance of plants with multiple therapeutic virtues, can significantly contribute to this fight against antibiotic resistance and the development of new antimicrobials.
In this study, the herbal medicines investigated were leaves of
Cymbopogon citratus (DC.) Stapf (C. citratus), Moringa
oleifera Lam (M. oleifera), Leucanthemum vulgare (L.
vulgare) and Vernonia amygdalina Delile (V. amygdalina); barks
of Cinchona officinalis (C. officinalis) and Enantia
chlorantha Oliv (E. chlorantha); barks and seeds of Garcinia
lucida Vesque (G. lucida) and Leaves and seeds of Azadirachta
indica (Neem) (A. indica). These medicinal plants are among
the most famous in Cameroon. For example, C. officinalis is a shrub of
the Rubiaceae family whose bark is well known for its very bitter taste and its
antimalarial properties. This plant is rich in alkaloids such as quinine,
dihydroquinine, cinchonidine, epiquinin, quinidine, dihydroquinidine, cinchonine
and epiquinidine [6, 7]. G. lucida Vesque is also a well-known herbal
medicine whose seeds, fruits and barks were reported to possess cardioprotective
and nephroprotective effects [8] and are recognized to be useful in the treatment
of gastric and gynecological infections, diarrheas, cure for snake bites as well
as an antidote against poison [9]. Otherwise, E. chlorantha also called
Epoue (Baka), Peye (Badjoue), and Nfol (Bulu), is used in the management of
various infections including dysentery, malaria, typhoid fever, jaundice, wounds,
high blood pressure, urinary infection, leprosy spots and convulsions [10].
Furthermore, A. indica known as Neem, is a monoecious tree of the
Meliaceae family whose oil produced from its seeds is widely used for its
medicinal properties in the northern part of Cameroon. It is known that compounds
in Neem extracts have anti-inflammatory, anti-hyperglycaemic, anti-carcinogenic,
antimicrobial, immune-modulator, anti-mutagenic, antioxidant, anti-ulcer, and
antiviral effects [11, 12]. Recent studies by Baildya et al. [12] even
found 19 compounds from this plant which may be used as anti-COVID-19. Finally,
M. oleifera, V. amygdalina and C. citratus are all
edible and medicinal plants. Every part of the M. oleifera, from the
leaves to the roots, has been reported to possess potential health benefits [13].
Besides its nutritional properties, M. oleifera is traditionally used to
treat skin infection, asthma, diabetes, diarrhea, arthritis, inflammation, cough,
fever, and headache. It has also been reported to have antioxidant,
anti-inflammatory, antitumor, antimicrobial, hepatoprotective and anti-arthritic
properties [14, 15, 16]. V. amygdalina (known in Cameroon under the popular
name of Ndolè) have been reported to have anticancer and antitumor activity
[17, 18]; antihepatotoxic activity [19]; hypoglycemic activity [20]; antibacterial
activity [21]; anti-inflammatory [22] as well as antioxidant property [23].
Moreover, C. citratus (lemongrass) is widely used as a tea and is rich
in minerals, vitamins, macronutrients (including carbohydrate, protein, and small
amounts of fat) and its leaves are a good source of various bioactive compounds
such as alkaloids, terpenoids, flavonoids, phenols, saponins and tannins that
confer C. citratus leaves pharmacological properties such as
anti-cancer, antihypertensive, anti-mutagenicity, anti-diabetic, antioxidant,
anxiolytic, anti-nociceptive and anti-fungal [24]. Like C. citratus, all
plants investigated in this study have various phytocompounds such as terpenoids
and xanthones products, alkaloids (such as protoberberines and phenanthrene
alkaloids), aporphines, zeatin, quercetin,
Therefore, the aim of this study was to evaluate the antimicrobial potential of aqueous and hydro ethanolic extracts of thirteen samples (bark, leaf, seed) of the eight above-mentioned plants from Cameroon and to assess their synergy with common antibiotics against various multiresistant uropathogenic bacteria.
The vegetal materials used in this study were the same used in our previous study [27]. They were barks and seeds of G. lucida Vesque, leaves of C. citratus (DC.) Stapf, L. vulgare, M. oleifera Lam and V. amygdalina Delile; barks of C. officinalis and E. chlorantha Oliv; and Leaves and seeds of A. indica (Neem).
Three reference cultures (purchased from American Type Culture Collection) were used to screen the antimicrobial activity of the different vegetal materials. They were S. aureus ATCC 6538 as Gram positive model, E. coliATCC 25922 as Gram negative model and C. albicans ATCC 10231 as fungi model. To assess the synergy between common antibiotics and the extracts, 11 strains of uropathogenic bacteria provided by the Department of Microbiology and Virology of the Peoples’ Friendship University of Russi were used. These strains were A. xylosoxidans 4892, C. freundii 426, E. avium 1669, E. coli 1449, K. oxytoca 3003, K. rizophilia. 1542, M. catarrhalis 4222, M. morganii 1543, P. aeruginosa 3057, S. aureus 1449 and S. agalactiae 3984. From frozen stock, the different strains were subcultured twice in BHI broth for bacteria and SAB broth for C. albicans ATCC 10231.
Brain Heart Infusion Broth (BHIB), Muller Hinton Agar (MHA), and Sabouraud Dextrose Broth (SDB) were purchased from HiMedia™ Laboratories Pvt. Ltd., India while Dimethyl sulfoxide (DMSO) was purchased from BDH Laboratories, VWR International Ltd., USA.
Plant extracts were prepared following the protocol described in previous studies by Mbarga et al. [27]. Briefly, fifty grams of plant material was weighed and introduced into separate conical flasks containing 450 mL of the solvents which were distilled water and ethanol/water (80:20, v/v), The mixture were shaken at 200 rpm for 24 h and 25 °C (Heidolph Inkubator 1000 coupled with Heidolph Unimax 1010, Germany), filtered (Whatman n°1), and concentrated at 40 °C using a rotary evaporator (IKA RV8, Germany) The semi-solid extracts obtained were kept at 4 °C for analyses.
Strains were cultured for 24 h in their appropriate medium and temperature.
Cells were collected by centrifugation (7000
The different dried extracts were dissolved in DMSO 5% (v/v) in order to obtain
a stock solution of 521 mg/mL. The solution was sterilized by microfiltration
(0.22
The antimicrobial activity of the extracts was screened using the well diffusion
method previously described by Mbarga et al. [28]. Briefly, 100
MIC is the lowest concentration of antibacterial agent that completely inhibits
the bacterial growth. The MIC of the different extracts was assessed using the
broth microdilution method [29, 30]. Briefly, the wells of a U-bottom 96-well
microplates were filled with 100
The tolerance level of the different pathogens to the plant extracts used in this study was performed according to the method described by Mondal et al. [31]. The following formula was used:
Tolerance value
The susceptibility of the uropathogenic bacteria to antibiotics was assessed with the modified Kirby–Bauer’s disk method exactly as described in previous study by Mbarga et al. [29] and the inhibition diameters were interpreted referring to the Clinical & Laboratory Standards Institute [32]. The antibiotics used are presented in Table 1 (Ref. [4, 32]). Resistant R, Intermediate I, and Susceptible S interpretations were obtained automatically using algorithms written in Excel software [Microsoft Office 2016 MSO version 16.0.13628.20128 (32 bits), USA] with the parameters described in Table 1 [4].
| Antibiotics | Inhibition diameters (mm) | ||
| R | I | S | |
| Ciprofloxacin, 30 |
d |
16–20 | d |
| Cefazolin, 30 µg/disk (CAZ) | d |
15–17 | d |
| Amoxicillin, 30 |
d |
14–17 | d |
| Ceftriaxone, 30 |
d |
14–20 | d |
| Trimethoprim, 30 |
d |
14–15 | d |
| Tetracyclin, 30 |
d |
15–18 | d |
| Nitrofurantoin, 200 |
d |
14–17 | d |
| Ampicillin, 25 |
d |
14–16 | d |
| Imipenem, 10 |
d |
14–15 | d |
| Cefazolin/clavulanic acid, 30/10 per disk (CAC) | d |
15–17 | d |
| Fosfomycin, 200 |
d |
13–15 | d |
| D, inhibition diameter. | |||
The antibiotics which gave inhibition diameters of less than 20 mm after
susceptibility test against the uropathogenic bacteria were modulated with 5
mg/mL of each extract. The test was performed as described by Rolta et
al. [33] with slight modifications. Briefly, in Petri dish, inoculated the test
bacteria, a sterile disc paper and an antibiotic disc were placed aseptically.
Then, 20
Where, “A” is the increase in fold area, “Y” the inhibition diameter of extract + antibiotic and “X” is the inhibition diameter of antibiotic alone.
The checkerboard method, commonly used for the determination of synergy between
the antibiotics and natural antibacterial compounds, was used for the antibiotic
modulation assay [28]. Modulations of ampicillin, benzylpenicillin, cefazolin,
ciprofloxacin, nitrofurantoin, and kanamycin were performed with extracts whose
MIC were successfully determined (Not those with MIC
The FIC index was interpreted as follows FIC
1.0 mg of the dried ethanolic extract was placed in an Eppendorf, 1.0 mL of a mixture of methanol:water (70:30) was added, and extraction was carried out in an ultrasonic bath for 30 minutes. The complete dissolution of a sample was noted, and it was transferred to a chromatographic vial for analysis.
Extract was analyzed by 6030 series HPLC-MS/MS (Agilent, USA). HPLC (Agilent
1290), consisting of a binary pump, an autosampler, and a thermostatted column
compartment, was performed with a Shim-pack FC-ODS C18 column (150
An EDX-7000 Shimadzu energy dispersive X-ray fluorescence (XRF) spectrometer was
used to identify the minerals present in the E. chlorantha extract . The
equipment settings were as followed: range of measured elements — 11Na - 92U;
X-ray generator — a tube with a Rh-anode, air cooled; voltage 4–50 kV, current
1–1000
All the experiments were carried out at least in triplicate. The statistical
significance was set at p
Fig. 2 presents the inhibition diameters of the different extracts on the tested pathogens. All extracts were not active against the pathogens. It is the case of the extracts (both ethanolic and aqueous extracts) from C. officinalis and G. lucida leaves. The rest of extracts were actives with inhibition diameters ranging from 5 to 36 mm.
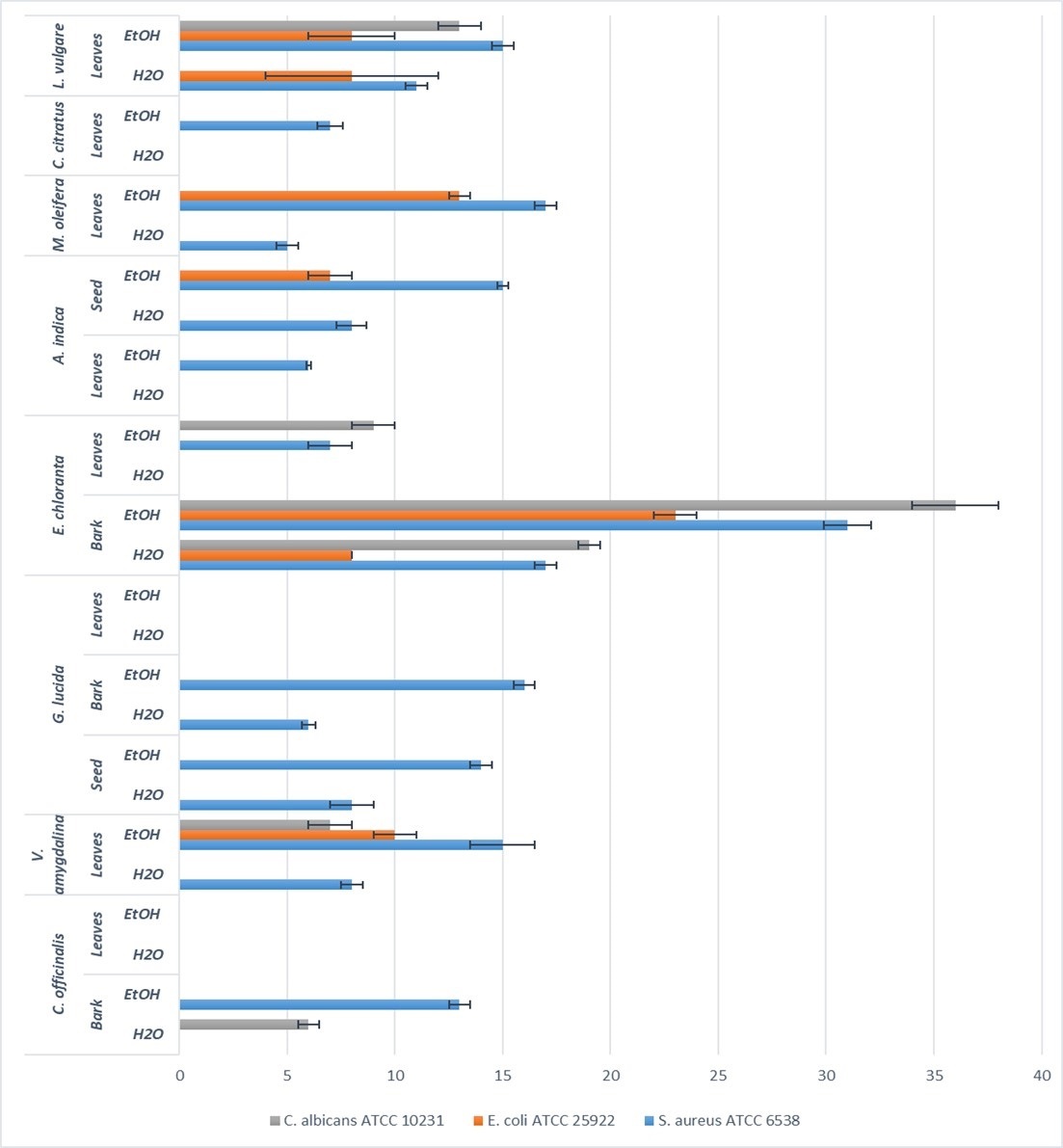 Fig. 2.
Fig. 2.Inhibition diameter (mm) resulting from the screening of antimicrobial activity by well diffusion method with 100 mg/mL of each extract.
Taking into consideration the extraction solvents, the highest inhibition diameters were mainly recorded with ethanol as solvent. Ethanol therefore appears as the solvent which extracted more antimicrobial compounds compared to water although the extraction yields were globally more important with water as solvent [27]. This observation could be ascribed to the insoluble nature of metabolites extracted with ethanol as solvent opposite to water. Indeed, most of bioactive compounds endowed with antimicrobial activity such as flavonoids, polyphenols, tannins and alkaloids are generally insoluble in water [35, 36, 37]. In a study conducted by Mouafo et al. [35], it was highlighted that ethanol extracted more antimicrobial compounds from plant materials opposite to water. A similar conclusion was also stated by Evbuomwan et al. [38].
With regards to the part of the plant material, extracts from bark were more actives independently of the pathogens and the extraction solvents. The highest activities on both bacterial and yeast strains were noticed with bark from E. chloranta. This could be attributed to the presence of high amount of antimicrobial alkaloids such as protoberberines (berberine, canadine, palmatine, jatrorrhizine, columbamine and pseudocolumbamine), phenanthrene alkaloids (atherosperminine and argentinine) and aporphines (7-hydroxydehydronuciferine and 7-hydroxydehydronornuciferine) in that plant as reported in the literature [26]. Several studies also highlighted the interesting antibacterial and antifungal properties of dried and fresh barks from E. chlorantha [10, 39, 40]. The extracts from E. chloranta therefore appears as a source of antimicrobials with a broad-spectrum activity. Similarly, ethanolic extracts of L. vulgare and V. amygdalina showed a broad-spectrum antimicrobial activity against all the tested pathogens. This could be ascribed to the richness of these extracts in phytochemicals such as saponins, sesquiterpenes, flavonoids, steroid glycosides, and lactones [20] for which recent studies reported its antibacterial activity [21].
Amongst leaves extracts, those derived from M. oleifera were more actives against the bacterial strains E. coli ATCC 25922 and S. aureus ATCC 6538 while extract from L. vulgare leaves was more active against C. albicans ATCC 10231. This result could be explained by the great variability of the phytochemical composition of the two vegetal materials and thus, the possible different action mechanisms against microorganisms.
When the tested pathogens are considered, it appears from Fig. 2 that S. aureus ATCC 6538 was more sensitive than E. coli ATCC 25922. This observation can be explained by the nature and composition of both cell wall and membranes which differ between Gram positive and Gram-negative bacteria. In fact, Gram-negative bacteria possess a lipopolysaccharides layer in their external membrane. This layer act as a barrier against the permeability antimicrobials [41].
A surprising observation was noticed in this study as the yeast C. albicans ATCC 10231 was more sensitive to the ethanolic and aqueous extracts of both bark and leaves extracts from E. chlorantha compared to the bacterial strains S. aureus ATCC 6538 and E. coli ATCC 25922. This result might arise from the antimicrobial action mechanisms of the bioactive compounds found in that plant. In fact, eukaryotic cells are known for their ability to resist to several antimicrobials as opposite to prokaryotic cells, they possess less phospholipids in their membranes. Phospholipids due to their anionic nature are mostly involved in the preliminary interaction with antimicrobial which will ease their penetration into cells [42, 43].
All the extracts inhibited the pathogens with MIC values which varied significantly from one plant material to another. As previously observed with qualitative tests, the extract of E. chloranta was the most active independently of the solvents and the tested pathogens. As shown in Table 2, the ethanolic extract of E. chloranta with MIC value lower than 2 mg/mL against the three pathogens was the most active extract. This important activity could result from the presence of alkaloids such as palmatine, coloumbamine and jatrorrhizine in its composition [44]. Indeed, these compounds have the ability to penetrate cells and intercalate DNA of microorganism leading to their death [45]. MIC values higher than 256 mg/mL were observed with some aqueous and ethanolic extracts against the different pathogens. This observation suggests that further analysis at concentrations higher than 256 should be performed in order to quantify the antimicrobial activity of these extracts. Moreover, extracts which have showed no activity in well diffusion qualitative test were active in liquid medium against the pathogens. Thus, suggesting that these extracts might contain antimicrobial compounds which cannot diffuse in the Muller Hinton agar.
| Plants | Part used | Solvents | S. aureus ATCC 6538 | E. coli ATCC 25922 | C. albicans ATCC 10231 | ||||||
| MIC | MBC | MBC/MIC | MIC | MBC | MBC/MIC | MIC | MBC | MBC/MIC | |||
| Cynchona officinalis | Bark | H |
256 | - | - | - | |||||
| EtOH 80% | 64 | 256 | 4 | 256 | - | 256 | - | ||||
| Leaves | H |
- | - | - | |||||||
| EtOH 80% | 256 | - | - | - | |||||||
| Vernonia amygdalina | Leaves | H |
128 | 256 | 2 | 256 | - | - | |||
| EtOH 80% | 32 | 128 | 4 | 64 | 256 | 4 | 256 | - | |||
| Garcinia lucida | Seed | H |
64 | 256 | 4 | 128 | 256 | 2 | 256 | - | |
| EtOH 80% | 8 | 64 | 8 | 8 | 128 | 16 | 256 | 256 | 1 | ||
| Bark | H |
128 | 256 | 2 | 256 | 256 | 1 | - | |||
| EtOH 80% | 32 | 128 | 4 | 128 | 256 | 2 | 256 | - | |||
| Leaves | H |
- | - | - | |||||||
| EtOH 80% | - | - | - | ||||||||
| Enantia chlorantha | Bark | H |
8 | 32 | 4 | 8 | 64 | 8 | 4 | 16 | 4 |
| EtOH 80% | 4 | - | 8 | - | 4 | - | |||||
| Leaves | H |
128 | 256 | 2 | - | - | |||||
| EtOH 80% | 32 | 256 | 8 | - | - | ||||||
| Azadirachta indica | Leaves | H |
- | - | - | ||||||
| EtOH 80% | 128 | - | - | - | |||||||
| Seed | H |
64 | 128 | 2 | 256 | - | - | ||||
| EtOH 80% | 8 | 32 | 4 | 64 | - | 256 | - | ||||
| Moringa oleifera | Leaves | H |
256 | - | 128 | - | - | ||||
| EtOH 80% | 16 | 64 | 4 | 32 | 256 | 8 | - | ||||
| Cymbopogon citratus | Leaves | H |
256 | - | - | - | |||||
| EtOH 80% | 128 | - | 256 | - | - | ||||||
| Leucanthemum vulgare | Leaves | H |
32 | 128 | 4 | 64 | 256 | 4 | 32 | 256 | 8 |
| EtOH 80% | 8 | 8 | 1 | 16 | 32 | 2 | 8 | 16 | 2 | ||
With water as solvent, the most important activity was recorded with extract from E. chloranta for which MIC of 8 mg/mL was noticed against S. aureus ATCC 6538 and E. coli ATCC 25922, and 4 mg/mL against C. albicans ATCC 10231. These activities were higher compared to those reported by Adesokan et al. [46]. They obtained with aqueous extract of E. chloranta, MIC values of 25 and 100 mg/mL against S. aureus and E. coli, respectively. This can be ascribed to several factors which influence the plant composition such as climate and soil composition, as well as to the tested strains.
The majority of extracts showed MIC values higher or equal to 256 mg/mL against
the yeast strain C. albicans ATCC 10231 independently of the extraction
solvent. Only ethanolic (MIC = 8 mg/mL) and aqueous (MIC = 32 mg/mL) extracts of
L. vulgare leaves, and the ethanolic (MIC
An observation of Table 2 revealed that S. aureus ATCC 6538 was the most sensitive strain as lower MIC values of most extracts were generally recorded against that strain. Thus, it clearly appears that the bacterial cell walls and membranes are involved in the antimicrobial activity mechanism of these extracts. This conclusion is different to that stated by Etame et al. [10] who found no significant difference in the MIC values of plant extracts against Gram positive and Gram-negative bacteria.
According to the classification established by Kuete [47] and Kuete and Efferth [48], the different extracts could be considered as deserving a weak antimicrobial activity independently of the extraction solvent and the tested strain as they scored MIC value higher than 0.625 mg/mL.
Minimum bactericidal concentration (MBC) and minimum fungicidal concentration (MIC) of the different extracts against three pathogenic strains were assessed and results are presented in Table 2. The MBC values ranged from 4 to more than 256 mg/mL. The strongest MBC value (4 mg/mL) against C. albicans ATCC 10231 and S. aureus ATCC 6538 was obtained with the ethanolic extract of E. chloranta bark. Against E. coli ATCC 25922, the strongest MBC value of 8 mg/mL was recorded with the same ethanolic extract of E. chloranta bark. Globally, lowest MBC values were recorded against S. aureus ATCC 6538, thus confirming its higher sensitivity to the different plant extracts.
The MBC values against E. coli ATCC 25922 and S. aureus ATCC 6538 obtained in this study with the aqueous extracts of E. chloranta were lower than that reported by Adesokan et al. [46]. The authors found with aqueous extract of E. chloranta, MBC values of 90 and 130 mg/mL against S. aureus and E. coli, respectively. In the same way, the MBC values of the ethanolic extract of V. amygdalina leaves against E. coli ATCC 25922 and S. aureus ATCC 6538 were lower than the 100 and 200 mg/mL obtained respectively against E. coli and S. aureus by Evbuomwan et al. [38] using the ethanolic extract from the same plant. This difference could be explained by the variability of phytochemical profile of the plant according to the geographical origin. Besides, the fact that microbial strain developed different resistance mechanism to antimicrobials as highlighted by several authors in the literature [49, 50] could also explained the variability in the MBC values.
It is established in the literature that an antimicrobial compound is considered
as bactericidal/fungicidal against a microbial strain when the ratio MBC/MIC or
MIC/MIC is
The use of combination therapy has been suggested as a new approach to improve
the efficacy of antimicrobial agents by screening crude extracts from medicinal
plants with good indications for use in combination with antibiotics [53]. The
checkboard method was applied to assess the synergy between some conventional
antibiotics and plant extracts which showed a valid MIC. After determining the
MIC of the plant materials (Table 2), we determined the MICs and MBCs of the
antibiotics (ampicillin, benzylpenicillin, cefazolin, ciprofloxacin,
nitrofurantoin, and kanamycin) against S. aureus ATCC 6538 and
E. coli ATCC 25922. As shown in Table 3, the MICs of the different
antibiotics varied from 4–64
| Pathogens | MIC (µg/mL) | |||||
| CIP | NIT | BP | AMP | Ka | CZ | |
| E. coli ATCC 25922 | 4 | 32 | 16 | 32 | 64 | 16 |
| S. aureus ATCC 6538 | 4 | 16 | ND | 4 | 32 | ND |
| MBC (µg/mL) | ||||||
| E. coli ATCC 25922 | 16 | 256 | 64 | 64 | 256 | 64 |
| S. aureus ATCC 6538 | 8 | 128 | ND | 4 | 32 | ND |
| AMP, ampicillin; BP, benzylpenicillin; CZ, cefazolin; CIP, ciprofloxacin; NIT, nitrofurantoin; and Ka, kanamycin; MIC, minimum inhibitory concentrations; MBC, minimum bactericidal concentration; ND, not determined. | ||||||
Antibiotics with high and quantified MICs values against the both pathogens
(ampicillin, nitrofurantoin, and kanamycin) were selected for modulation assays.
Similarly, regarding the plant extracts, we decided to work with extracts with
low and determined MICs. Thus, it was performed a modulation of ampicillin,
nitrofurantoin, and kanamycin with aqueous extract of E. chlorantha bark
and ethanolic extract of G. lucida seed, A. indica seed and
L. vulgare leaves. As shown in Table 4, the fractional inhibitory
concentration (FIC) ranged from 0.125 to 0.750. No antagonism (FIC
| Plants | ATB | S. aureus ATCC 6538 | E. coli ATCC 25922 | ||||||||
| MIC extr (mg/mL) | MIC ATB (µg/mL) | MIC’ extr (mg/mL) | MIC’ ATB (µg/mL) | FIC | MIC extr (mg/mL) | MIC ATB (µg/mL) | MIC’ extr (mg/mL) | MIC’ ATB (µg/mL) | FIC | ||
| E. Chlorantha bark H |
AMP | 8 | 4 | 0.5 | 1 | 0.313 | 8 | 32 | 0.5 | 2 | 0.125 |
| Ka | 32 | 0.5 | 2 | 0.125 | 64 | 1 | 8 | 0.250 | |||
| NIT | 16 | 1 | 2 | 0.250 | 32 | 0.5 | 4 | 0.188 | |||
| G. lucida seed EtOH | AMP | 8 | 4 | 2 | 1 | 0.500 | 8 | 32 | 2 | 4 | 0.375 |
| Ka | 32 | 4 | 4 | 0.625 | 64 | 4 | 16 | 0.750 | |||
| NIT | 16 | 2 | 4 | 0.500 | 32 | 2 | 8 | 0.500 | |||
| A. Indica seed EtOH | AMP | 8 | 4 | 1 | 1 | 0.375 | 64 | 32 | 4 | 8 | 0.313 |
| Ka | 32 | 4 | 4 | 0.625 | 64 | 8 | 16 | 0.375 | |||
| NIT | 16 | 1 | 2 | 0.250 | 32 | 4 | 2 | 0.125 | |||
| L. vulgare leaves EtOH | AMP | 8 | 4 | 2 | 1 | 0.500 | 16 | 32 | 4 | 4 | 0.375 |
| Ka | 32 | 2 | 2 | 0.313 | 64 | 8 | 16 | 0.750 | |||
| NIT | 16 | 1 | 2 | 0.250 | 32 | 2 | 4 | 0.250 | |||
| ATB, antibiotics; FIC, fractional inhibitory concentration; MIC, minimum
inhibitory concentrations; MBC, minimum bactericidal concentration; AMP,
ampicillin; BP, benzylpenicillin; CZ, cefazolin; CIP, ciprofloxacin; NIT,
nitrofurantoin; and Ka, kanamycin. FIC | |||||||||||
The solid-medium modulation test using commercial antibiotic discs is a less complex means for synergy testing. In this study, after having observed in liquid medium (checkboard method) that most of the extracts had a synergistic effect with antibiotics on non-resistant bacteria, we undertook to perform modulation tests in solid medium to assess the extent to which extracts from the tested plants could potentially enhance the performance of conventional antibiotics against a wide range of resistant bacteria. So, we started by determining the effectiveness of antibiotic discs alone. The sensitivity of the eleven uropathogenic bacteria used in this study to eleven (11) antibiotics was determined (Table 5) and the multidrug resistance (MDR) index of each bacterium was calculated. No bacteria were resistant to imipenem and amoxiclav. Regarding the other antibiotics, we found resistance in 10/11 bacteria to ampicillin, 8/11 to trimethoprim and tetracyclin, 6/11 to cefazolin + clavulanic acid, 5/11 to ceftazidime, 4/11 to nitrofurantoin and 1/11 to ceftriaxone and ciprofloxacin. The highest MDR index (0.54) was found in E. coli 1449 which was resistant to ampicillin, ceftazidime, cefazoline + clavulanic acid, ceftriaxone, tetracycline and trimethoprim. St. agalactiae 3984 and K. rizophilia 1542 scored the same MDR index of 0.45. K. rizophilia 3984 was resistant to Ampicillin, Ceftazidime, Cefazoline + clavulanic, Nitrofurantoin and tetracycline while St. agalactiae 3984 was resistant to trimethoprim, ampicillin, ceftazidime, cefazoline + clavulanic and tetracycline The lowest MDR index (0.27) was recorded with C. freundii 426 and S. aureus 1449. This result is consistent with those obtained by other authors on the resistance of clinical strains to antibiotics [61, 62, 63].
| Pathogens | NIT | TE | CTR | AMC | FO | CAZ | IPM | CAC | CIP | AMP | TR | MDR Index |
| Ac. Xylosoxidans 4892 | 6 |
11 |
23 |
36 |
6 |
16 |
32 |
16 |
20 |
20 |
6 |
0.36 |
| C. freundii 426 | 21 |
30 |
27 |
35 |
40 |
12 |
37 |
10 |
30 |
6 |
22 |
0.27 |
| E. avium 1669 | 21 |
6 |
30 |
25 |
31 |
23 |
27 |
24 |
15 |
6 |
6 |
0.36 |
| E. coli 1449 | 24 |
11 |
8 |
27 |
30 |
7 |
22 |
12 |
26 |
6 |
6 |
0.54 |
| K. oxytoca 3003 | 20 |
8 |
22 |
24 |
25 |
15 |
27 |
6 |
30 |
6 |
6 |
0.36 |
| K. rizophilia 1542 | 10 |
13 |
22 |
22 |
28 |
10 |
23 |
6 |
30 |
13 |
21 |
0.45 |
| M. catarrhalis 4222 | 12 |
22 |
24 |
36 |
27 |
16 |
27 |
21 |
32 |
10 |
15 |
0.18 |
| M. morganii 1543 | 15 |
6 |
33 |
17 |
13 |
23 |
22 |
23 |
22 |
10 |
6 |
0.36 |
| P. aeruginosa 3057 | 6 |
13 |
21 |
16 |
27 |
15 |
34 |
22 |
30 |
6 |
6 |
0.36 |
| S. aureus 1449 | 16 |
25 |
18 |
27 |
27 |
6 |
25 |
6 |
26 |
6 |
6 |
0.27 |
| St. agalactiae 3984 | 18 |
6 |
27 |
27 |
27 |
10 |
30 |
12 |
18 |
6 |
6 |
0.45 |
| AMC, Amoxycillin; AMP, Ampicillin; CZ, Cefazolin; CAC, Cefazolin/clavulanic acid; CAZ, Ceftazidime; CTR, Ceftriaxone; CIP, Ciprofloxacin; FO, Fosfomycin; IMP, Imipenem; NIT, Nitrofurantoin; TE, Tetracyclin; TR, Trimethoprim; MDR, multidrug resistance. | ||||||||||||
Various means have been implemented in recent years to provide effective
solutions to the antibioresistance issue, and the studies carried out target
bacteria that are resistant or not. Here we focused on evaluating the modulatory
effect of ethanolic extracts of plant materials on antibiotics which presented an
inhibition diameter lower than 20 mm (Table 5) against uropathogenic bacteria.
Tables 6,7,8,9,10 present, respectively, in the form of increase in fold area
(IFA), the modulating effect of plant extracts with ampicillin (AMP) (Table 6),
ceftazidime (CAZ) (Table 7), tetracycline (TE) (Table 8), nitrofurantoin (NIT)
(Table 9) and trimethoprim (TR) (Table 10). As with synergy tests using the
checkboard method on non-resistant bacteria (Table 5), we found that E.
chlorantha bark (ECB) (indenpendently of the solvent used) had the best
modulating properties on most of the antibiotics tested. Its IFA was
significantly different (p
| Pathogens | C. officinalis | V. amygdalina | G. lucida | E. chlorantha | A. indica | M. oleifera | C. citratus | L. vulgare | |||||
| Bark | Leaves | Leaves | Seed | Bark | Leaves | Bark | Leaves | Leaves | Seed | Leaves | Leaves | Leaves | |
| C. freundii 426 | 0.31 | 0.10 | 0.00 | 0.53 | 0.36 | 0.10 | 1.47 | 0.14 | 0.34 | 0.42 | 0.15 | 0.05 | 0.42 |
| E. avium 1669 | 0.31 | 0.10 | 0.10 | 0.53 | 0.36 | 0.00 | 1.47 | 0.14 | 0.31 | 0.42 | 0.15 | 0.05 | 0.65 |
| E. coli 1449 | 0.27 | 0.09 | 0.27 | 0.46 | 0.31 | 0.09 | 1.25 | 0.12 | 0.31 | 0.36 | 0.13 | 0.04 | 0.57 |
| K. oxytoca 3003 | 0.32 | 0.10 | 0.00 | 0.56 | 0.27 | 0.00 | 1.56 | 0.14 | 0.11 | 0.44 | 0.16 | 0.05 | 0.70 |
| K. rizophilia 1542 | 0.72 | 0.21 | 0.96 | 1.25 | 0.56 | 0.21 | 3.84 | 0.30 | 0.42 | 0.96 | 0.32 | 0.10 | 1.59 |
| M. catarrhalis 4222 | 0.58 | 0.17 | 0.36 | 1.01 | 0.46 | 0.17 | 3.00 | 0.25 | 0.46 | 0.78 | 0.27 | 0.09 | 1.28 |
| M. morganii 1543 | 0.46 | 0.14 | 0.00 | 0.78 | 0.36 | 0.07 | 2.24 | 0.20 | 0.36 | 0.60 | 0.21 | 0.07 | 0.98 |
| P. aeruginosa 3057 | 1.30 | 0.36 | 1.25 | 2.36 | 1.15 | 0.17 | 8.00 | 0.52 | 1.01 | 1.78 | 0.56 | 0.17 | 3.07 |
| S. aureus 1449 | 0.27 | 0.13 | 0.00 | 0.72 | 0.38 | 0.06 | 2.06 | 0.18 | 0.21 | 0.56 | 0.20 | 0.06 | 0.91 |
| St. agalactiae 3984 | 0.23 | 0.11 | 0.49 | 0.63 | 0.34 | 0.06 | 1.78 | 0.16 | 0.10 | 0.49 | 0.17 | 0.06 | 0.79 |
| Pathogens | C. officinalis | V. amygdalina | G. lucida | E. Chlorantha | A. indica | M. oleifera | C. citratus | L. vulgare | |||||
| Bark | Leaves | Leaves | Seed | Bark | Leaves | Bark | Leaves | Leaves | Seed | Leaves | Leaves | Leaves | |
| Ac. Xylosoxidans 4892 | 0.27 | 0.13 | 0.13 | 0.73 | 0.32 | 0.03 | 1.64 | 0.13 | 0.06 | 0.56 | 0.56 | 0.06 | 1.07 |
| C. freundii 426 | 0.46 | 0.00 | 0.17 | 1.09 | 0.44 | 0.10 | 2.36 | 0.09 | 0.09 | 1.25 | 1.01 | 0.00 | 1.53 |
| E. coli 1449 | 1.94 | 0.31 | 0.65 | 1.94 | 0.65 | 0.31 | 4.90 | 0.31 | 0.15 | 1.47 | 0.31 | 0.15 | 3.01 |
| K. oxytoca 3003 | 0.60 | 0.00 | 0.14 | 0.78 | 0.35 | 0.00 | 1.78 | 0.00 | 0.00 | 0.78 | 0.44 | 0.00 | 1.15 |
| K. rizophilia 1542 | 0.96 | 0.21 | 0.69 | 1.25 | 0.44 | 0.00 | 3.00 | 0.21 | 0.10 | 1.56 | 0.96 | 0.10 | 1.89 |
| M. catarrhalis 4222 | 0.72 | 0.27 | 0.41 | 0.49 | 0.32 | 0.00 | 1.64 | 0.00 | 0.06 | 0.72 | 0.41 | 0.13 | 1.07 |
| P. aeruginosa 3057 | 0.28 | 0.14 | 0.44 | 0.71 | 0.35 | 0.14 | 1.78 | 0.14 | 0.00 | 0.87 | 0.60 | 0.07 | 1.15 |
| S. aureus 1449 | 3.00 | 0.78 | 0.56 | 1.31 | 0.56 | 0.36 | 6.11 | 1.25 | 0.36 | 3.00 | 0.78 | 0.78 | 4.69 |
| St. agalactiae 3984 | 0.96 | 0.21 | 0.32 | 1.25 | 0.32 | 0.44 | 3.00 | 0.44 | 0.10 | 1.25 | 0.96 | 0.10 | 1.89 |
| Pathogens | C. officinalis | V. amygdalina | G. lucida | E. chlorantha | A. indica | M. oleifera | C. citratus | L. vulgare | |||||
| Bark | Leaves | Leaves | Seed | Bark | Leaves | Bark | Leaves | Leaves | Seed | Leaves | Leaves | Leaves | |
| Ac. Xylosoxidans 4892 | 0.80 | 0.00 | 1.36 | 2.47 | 1.04 | 0.19 | 2.64 | 0.62 | 0.40 | 1.68 | 0.00 | 0.40 | 1.39 |
| E. avium 1669 | 1.70 | 0.17 | 1.72 | 4.44 | 2.18 | 0.00 | 5.25 | 1.25 | 1.01 | 3.69 | 0.17 | 0.25 | 4.44 |
| E. coli 1449 | 0.77 | 0.19 | 1.36 | 1.12 | 0.81 | 0.00 | 2.64 | 0.62 | 0.40 | 1.68 | 0.00 | 0.19 | 1.12 |
| K. oxytoca 3003 | 1.53 | 2.30 | 2.30 | 3.20 | 1.67 | 0.00 | 6.11 | 1.51 | 0.36 | 3.69 | 0.00 | 0.78 | 3.69 |
| K. rizophilia 1542 | 0.52 | 0.14 | 0.50 | 1.14 | 0.61 | 0.08 | 2.13 | 0.51 | 0.42 | 1.37 | 0.16 | 0.33 | 0.92 |
| M. morganii 1543 | 1.43 | 0.00 | 1.20 | 2.36 | 1.45 | 0.00 | 7.03 | 2.06 | 0.78 | 3.69 | 0.00 | 0.00 | 3.00 |
| P. aeruginosa 3057 | 0.51 | 0.16 | 1.11 | 2.00 | 0.59 | 0.00 | 2.13 | 0.51 | 0.51 | 1.37 | 0.51 | 0.16 | 1.61 |
| St. agalactiae 3984 | 1.38 | 0.00 | 1.25 | 2.36 | 1.45 | 0.17 | 4.44 | 0.78 | 0.00 | 3.69 | 0.00 | 0.36 | 3.00 |
| Pathogens | C. officinalis | V. amygdalina | G. lucida | E. chlorantha | A. indica | M. oleifera | C. citratus | L. vulgare | |||||
| Bark | Leaves | Leaves | Seed | Bark | Leaves | Bark | Leaves | Leaves | Seed | Leaves | Leaves | Leaves | |
| Ac. Xylosoxidans 4892 | 1.13 | 0.15 | 0.95 | 1.64 | 0.83 | 0.00 | 7.03 | 0.78 | 0.61 | 4.48 | 0.21 | 0.00 | 0.58 |
| K. oxytoca 3003 | 0.63 | 0.00 | 1.21 | 0.93 | 1.51 | 0.10 | 1.40 | 0.32 | 1.37 | 0.96 | 0.63 | 0.06 | 0.77 |
| K. rizophilia 1542 | 1.10 | 0.10 | 1.09 | 2.97 | 0.76 | 0.16 | 3.41 | 1.25 | 0.68 | 1.27 | 0.28 | 0.30 | 1.15 |
| M. catarrhalis 4222 | 1.52 | 0.72 | 0.98 | 1.03 | 1.21 | 0.00 | 2.67 | 0.00 | 0.74 | 0.78 | 0.38 | 0.05 | 1.01 |
| M. morganii 1543 | 0.68 | 0.13 | 0.69 | 0.83 | 0.81 | 0.00 | 2.00 | 0.14 | 0.74 | 1.43 | 0.48 | 0.06 | 0.88 |
| P. aeruginosa 3057 | 1.06 | 0.81 | 0.97 | 1.55 | 1.02 | 0.00 | 3.31 | 0.78 | 0.99 | 3.64 | 0.31 | 0.05 | 1.83 |
| S. aureus 1449 | 0.93 | 0.29 | 1.00 | 1.47 | 0.79 | 0.00 | 1.85 | 0.00 | 0.86 | 1.08 | 0.86 | 0.16 | 0.85 |
| St. agalactiae 3984 | 1.06 | 0.67 | 1.07 | 1.08 | 1.10 | 0.15 | 1.60 | 0.36 | 0.41 | 0.50 | 0.31 | 0.11 | 1.02 |
| Pathogens | C. officinalis | V. amygdalina | G. lucida | E. chlorantha | A. indica | M. oleifera | C. citratus | L. vulgare | |||||
| Bark | Leaves | Leaves | Seed | Bark | Leaves | Bark | Leaves | Leaves | Seed | Leaves | Leaves | Leaves | |
| Ac. Xylosoxidans 4892 | 1.63 | 0.78 | 1.42 | 1.07 | 1.47 | 0.56 | 7.49 | 1.28 | 1.01 | 2.47 | 0.46 | 2.06 | 0.78 |
| E. avium 1669 | 1.13 | 1.72 | 0.90 | 2.91 | 0.70 | 1.78 | 1.86 | 0.82 | 0.36 | 0.70 | 1.25 | 2.04 | 3.69 |
| E. coli 1449 | 1.96 | 0.00 | 1.61 | 2.03 | 1.32 | 2.67 | 3.87 | 1.75 | 0.56 | 1.32 | 1.87 | 1.69 | 2.36 |
| K. oxytoca 3003 | 2.06 | 0.40 | 0.77 | 1.67 | 1.12 | 0.36 | 3.13 | 0.50 | 0.78 | 1.12 | 1.71 | 1.78 | 1.78 |
| M. morganii 1543 | 1.18 | 1.78 | 0.87 | 1.30 | 0.92 | 0.00 | 2.46 | 0.64 | 0.36 | 1.92 | 1.56 | 1.65 | 1.25 |
| P. aeruginosa 3057 | 1.78 | 0.45 | 1.25 | 2.45 | 1.28 | 0.78 | 3.77 | 1.28 | 1.25 | 2.28 | 1.05 | 1.67 | 3.00 |
| S. aureus 1449 | 2.06 | 0.43 | 0.75 | 1.84 | 0.86 | 1.51 | 2.31 | 0.50 | 2.36 | 0.86 | 1.79 | 1.78 | 2.06 |
| St. agalactiae 3984 | 1.56 | 0.27 | 0.94 | 2.97 | 0.76 | 0.78 | 2.06 | 0.86 | 1.78 | 0.76 | 1.15 | 1.78 | 3.34 |
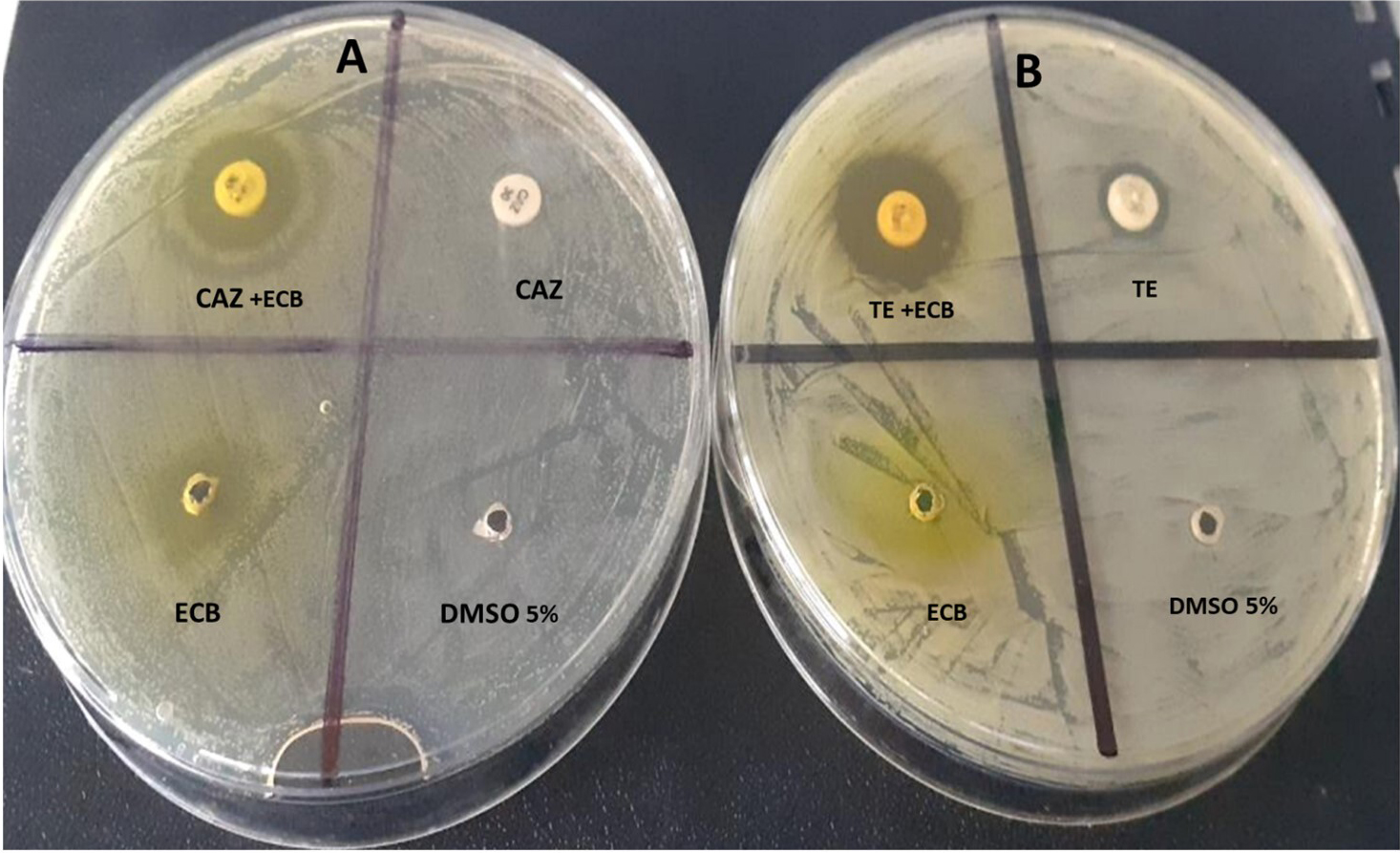 Fig. 3.
Fig. 3.Modulation of ceftazidime (CAZ) and tetracycline (TE) with E. Chlorantha bark (ECB) respectively against S. aureus 1449 (A) and K. oxytoca 3003 (B).
The ECB-TR and ECB-NIT combinations also showed a strong increase in activity especially against Ac. Xylosoxidans 4892 (both) K. rizophilia 1542 (only ECB-NIT) P. aeruginosa 3057 (both), and K. oxytoca 3003 (only ECB-TR). Otherwise, L. vulgare leaves (LVL) and G. lucida seed (GLS) have also demonstrated some promising results when combined with certain antibiotics. For example, the combinations LVL-AMP, LVL-CAZ, LVL-CAZ, LVL-TE, LVL-TE, LVL-TE, LVL-TE, LVL-TR, LVL-TR and LVL-TR were respectively very active against P. aeruginosa 3057, E. coli 1449, P. aeruginosa 3057, E. avium 1669, K. oxytoca 3003, M. morganii 1543, St. agalactiae 3984, E. avium 1669, P. aeruginosa 3057 and St. agalactiae 3984. Interestingly, C. citratus well-modulated trimethoprim while with other antibiotics the activity was lower compared to ECB, LVL and GLS.
Generally, the activity of antibiotics to which bacteria were resistant was interestingly increased with E. chlorantha bark, L. vulgare leaves and G. lucida seed while the action of ATB-extract combinations was moderate with G. lucida bark and weak with the other extracts. Besides mechanisms previously mentioned for checkboard method, another explanation of the modulation effect observed in this study in solid media could be the fact that some plant-derived compounds enhance the activity of antimicrobial compounds by inhibiting MDR efflux systems in bacteria [54]. Bacterial efflux pumps are responsible for a significant level of resistance to antibiotics in pathogenic bacteria [5]. Indeed, efflux pumps allow bacteria to flush antibiotics out of bacterial cells and therefore reduce their sensitivity to conventional antibiotics [5]. It is likely that ethanolic extracts from our plants may contain potential efflux pump inhibitors which are likely to be broad considering that the synergistic effect of the extract was observed on both Gram-positive and Gram-negative organisms. Several studies reported the isolation of some broad-spectrum efflux pump inhibitors from plant materials [5, 54, 62, 64]. A good example of this is the work of Smith et al. [64] who reported one efflux inhibitor (ferruginol) from the cones of Chamaecyparis lawso-niana, which inhibited the activity of the quinolone resistance pump (NorA), the tetracycline resistance pump (TetK) and the erythromycin resistance pump (MsrA ) in Staphylococcus aureus.
Finally, the management of bacterial infections should be done by aggressive empiric therapy with at least two antimicrobial agents [54]. Empiric combination antimicrobial therapy is usually applied to expand antibacterial spectrum and reduce the selection of resistant mutants during treatment [54]. In addition, combinations of agents that exhibit synergy or partial synergy could potentially improve the outcome for patients with difficult to treat infections [54]. However, this approach while viable, has limitations because it might not be effective for a long period of time due the possible alteration in the susceptibility of bacteria. Therefore, the development of new classes of antimicrobial compounds is of significant importance.
The HPLC–MS/MS chromatogram of the ethanolic extract from Enantia chlorantha bark showed a total of 11 peaks (Fig. 4). The compounds corresponding to these peaks were identified based on their retention time, peak area (%), height (%) and mass spectral fragmentation (m/z, MS and m/z, MS/MS) (Table 11). After comparing the data obtained with that previously reported in the literature [65, 66], it was found thar all the identified compounds belonged to the alkaloids family and the major constituents were palmatine (51.63%), columbamine + 7,8-dihydro-8-hydroxypalmatine (19.21%), jatrorrhizine (11.02%) and pseudocolumbamine (6.33%). Four (4) compounds (compounds 1, 2, 3 and 6) were not clearly identified and seemed to have been never reported in the literature.The structures of these compounds were only predicted since without isolation and NMR research, reliable description of new component structure is impossible. However, the composition of this plant explains its strong antimicrobial activity because the compounds it contains have already been individually reported for their antimicrobial activities in other studies [40, 44, 46].
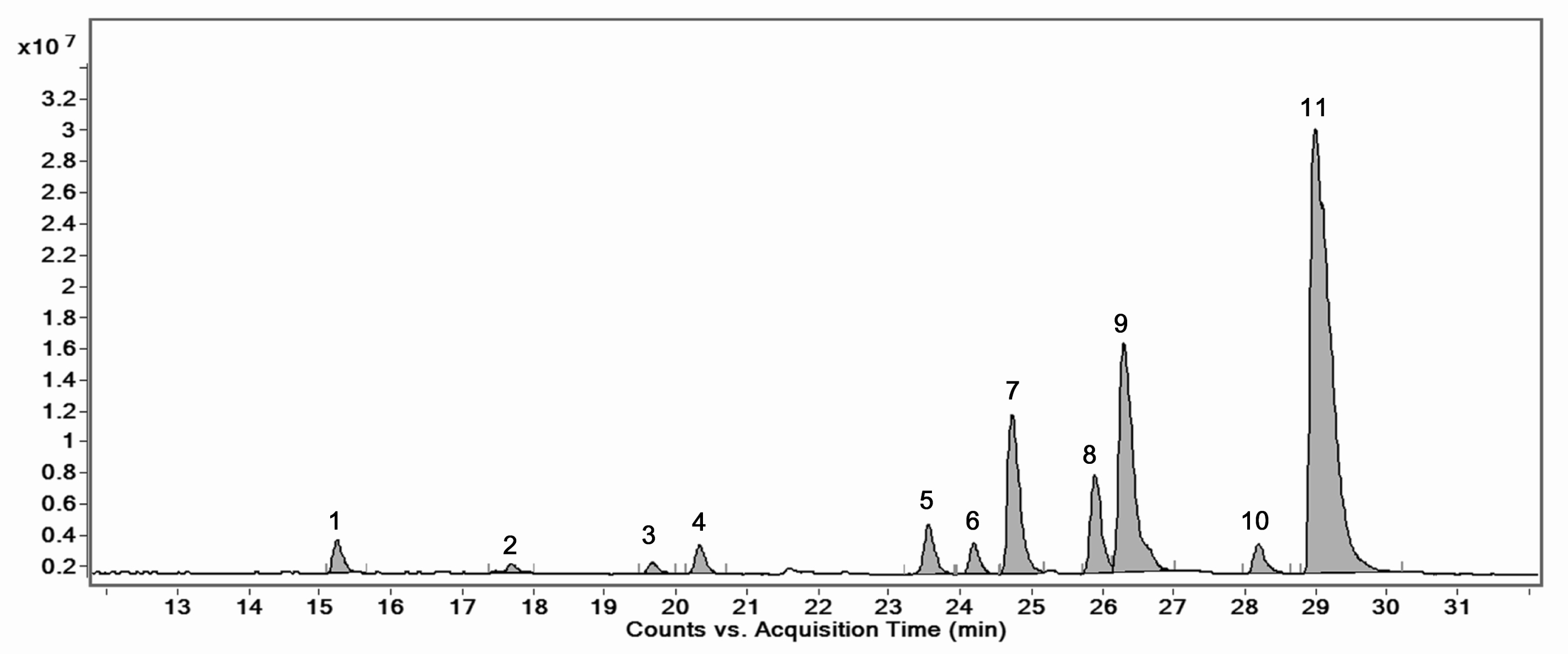 Fig. 4.
Fig. 4.HPLC-MS/MS chromatogram of the optimized extract of Enantia chloranta bark.
| № | Name | RT, min | Structure | m/z, MS spectra | m/z, MS/MS spectra (CE 30 eV) | % |
| 1 | Compound 1 | 15,24 | 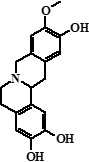 |
314 | 269, 253, 237, 211, 209, 192, 175, 160, 145, 143, 137, 121, 107 | 1.86 |
| 2 | Compound 2 | 17,68 | 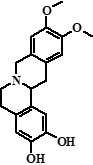 |
328 | 283, 269, 253, 237, 189, 174, 151, 121, 107 | 0.64 |
| 3 | Compound 3 | 19,66 | 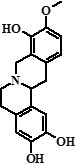 |
314 | 269, 253, 237, 211, 209, 192, 175, 145, 143, 137, 121, 107 | 0.69 |
| 4 | Pseudorotundine | 20,33 | 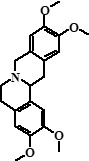 |
356 | 192, 190, 177 | 1.68 |
| 5 | Tetrahydropalmatine (Rotundine) | 23,54 | 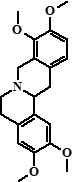 |
356 | 192, 190, 177 | 3.12 |
| 6 | Compound 6 + Unknown compound | 24,19 | 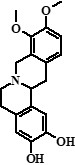 + ??? + ??? |
328 (78%), 368 (22%) | 283, 268, 252, 237, 189, 174, 145, 121, 107 + 353, 352, 338, 336, 324, 310, 307 | 1.83 |
| 7 | Jatrorrhizine | 24,73 | 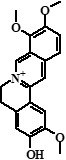 |
338 | 323, 322, 308, 294, 279 | 11.02 |
| 8 | Pseudocolumbamine | 25,88 | 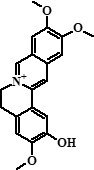 |
338 | 323, 322, 308, 294, 279, 265 | 6.33 |
| 9 | Columbamine + 7,8-dihydro-8-hydroxypalmatine | 26,29 | 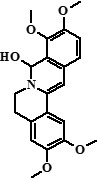 + +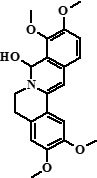 |
338 (95%), 370 (5%) | 323, 322, 308, 306, 294, 279, 277, 265 + 355, 354, 340, 326, 312, 311 | 19.21 |
| 10 | Pseudopalmatine | 28,19 | 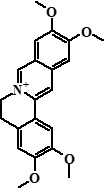 |
352 | 336, 320, 308, 294, 292, 279 | 1.99 |
| 11 | Palmatine | 28,98 | 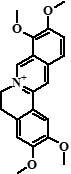 |
352 | 336, 322, 320, 308, 294, 292, 278 | 51.63 |
In addition, X-ray fluorescence spectrum made it possible to qualitatively assess the microelement present in O-ECB. As shown in Table 12, the minerals found were sulfur (S), Silicon (Si), Chlorine (Cl), Potassium (K), Calcium (Ca), Manganese (Mn), Iron (Fe), Zinc (Zn) and Bromine (Br). The highest mean fluorescence intensities were recorded for Br (4.6529 cps/uA), Fe (3.4854 cps/uA) and Cl (2.5942 cps/uA), which could mean that these minerals are the most abundant in O-ECB.
| Chemical element | Mean fluorescence intensity, cps/uA | Standard deviation |
| Si | 0.0581 | 0.0024 |
| S | 0.2052 | 0.0028 |
| Cl | 2.5942 | 0.0097 |
| K | 0.6511 | 0.0010 |
| Ca | 0.6185 | 0.0253 |
| Mn | 0.1482 | 0.0033 |
| Fe | 3.4854 | 0.0127 |
| Zn | 0.1624 | 0.0014 |
| Br | 4.6529 | 0.0114 |
We found that only L. vulgare leaves, G. lucida seed, and E. chlorantha bark possessed exploitable and promising antimicrobial properties. E. chlorantha bark was the most active and had strong activity against both Gram-positive and Gram-negative bacteria as well as fungi. The results of this study clearly demonstrated that E. chloranta bark, L. vulgare leaves and G. lucida seed act synergistically with most common antibiotics and hence increases drug efficacy. Finally, under the limitations of this study, it can be concluded that E. chlorantha bark, which contains a high proportion of alkaloids, especially palmatine and its derivatives, should be considered for further studies in the search for new antimicrobials.
MIC, Minimum Inhibitory Concentration; MBC, Minimum Bactericidal Concentration; EE, Ethanolic extract; AE, Aqueous extract.
MMJA designed the research study. MMJA, AKLD, PIV, DMS & SIP performed the research. MMJA analyzed the data. SLA, MMJA, KP, MR, HTM, IAMM, YNV and SIP wrote the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript.
Not applicable.
Not applicable.
This study has been supported by the RUDN University strategic Academic Leadership Program.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.