


Frontiers in Bioscience-Elite (FBE) is published by IMR Press from Volume 13 Issue 2 (2021). Previous articles were published by another publisher on a subscription basis, and they are hosted by IMR Press on imrpress.com as a courtesy and upon agreement with Frontiers in Bioscience.
 , Marco Chiera 2, Diego Lanaro 3, Massimo Fioranelli 4
, Marco Chiera 2, Diego Lanaro 3, Massimo Fioranelli 41 Società Italiana di Psiconeuroendocrinoimmunologia, Rome, Italy
2 Società Italiana di Psiconeuroendocrinoimmunologia, Rome, Italy
3 C.O.M.E. Collaboration ONLUS, Pescara, Italy
4 Department of nuclear physics, sub-nuclear and radiation, Guglielmo Marconi University, Rome, Italy
Abstract
The stress response, by virtue of release of glucocorticoids and catecholamines and by modifying the endocrine, neural, and immune responses, can impact the function of the fibroblasts and myofibroblasts that reside throughout the body and more specifically in the fascia, a ubiquitous and multi-functional connective tissue that supports the body. In the present paper, we review these stress-induced responses relying on psychoneuroendocrineimmunology.
Keywords
- Stress
- Glucocorticoids
- Catecholamines
- Immune system
- Fascia
- Connective tissue
- Myofibroblast
- Mechanobiology
- Psychoneuroendocrineimmunology
- Review
The hypothalamic-pituitary-adrenal (HPA) axis and the stress response are paramount for the organism to cope with physical threats or psychosocial adversities. Extensive knowledge has been gained on how:
1. the stress response can affect every endocrine axis (1–3);
2. many cells of several tissues can produce and secrete stress hormones (and also catecholamines) that can impact the central HPA axis and the whole body (4–6);
3. the HPA axis can heavily influence the immune system (7, 8);
4. the stress response can affect the body structure (9, 10).
Fascia is a particular kind of connective tissue – different from blood, cartilage, and bones – having a ubiquitous, multi-functional nature: it provides the structural and functional architecture of the body. The fascia integrates the functions of several systems (e.g., the nervous, immune, and cardiovascular systems) and can transmit mechanical, chemical, and electrical information (11–13). The fascia has no universally accepted definition of it (13), but since defining fascia is not our purpose, we refer the reader to the appropriate references (14, 15).
For the sake of simplicity, we will consider the terms “fascia” and “fascial system” according to the definition given by Carla Stecco and Robert Schleip in an editorial about fascia science and its clinical applications: a fascia is “a sheath, a sheet or any number of other dissectible aggregations of connective tissue that forms beneath the skin to attach, enclose, separate muscles and other internal organs,” whereas the fascial system is “a network of interacting, interrelated, interdependent (fascial) tissues forming a complex whole, all collaborating to perform a movement.” This definition of “fascia” can be useful to describe anatomical features, whereas the term “fascial system” may prove helpful to manual therapists in explaining what occurs during a manual therapy session (16).
Although the stress response can alter the fascia and related tissues (10), it seems no previous research has focused on describing the relationship between the stress response and fascia. On June 13, 2020, for instance, only four studies were retrieved from the PubMed database with the search string ‘fascia AND (“stress response” OR “HPA axis” OR “stress axis” OR “hypothalamic-pituitary-adrenal axis”).’ Only one of these papers briefly discusses the relationship between HPA axis hyperactivity and fascial dysfunction in the case of fibromyalgia (17). Considering the attention paid to the fascia by many manual therapists, we think it is paramount to show how the stress response, fascia, and the immune system can interact.
By understanding the physio-pathological relations between stress, immunity, and fascia, it is possible to better comprehend the interaction between body functions (e.g., circulation, immunity, and neuro-hormonal regulation) and structures (e.g., fascial system, muscles, and bones). Recent advancements in various fields – including medicine, psychology, sociology, biology, mechano-biology, and psychoneuroendocrineimmunology (PNEI), which studies the interactions between mind, body, and environment in determining health or disease (1, 18) – have also recognized that chronic pathologies can benefit from a therapeutic approach that takes into account:
1. the patient’s lifestyle – emotions, diet, and physical activity can affect many diseases (19–22);
2. the systemic influences of every organ – e.g., the vicious cycle connecting dysbiosis, arthritis, low back pain, and Parkinson (23, 24);
3. the benefits of both pharmacological and non-pharmacological therapies (18) – clinicians should cure disease, but also preserve and promote health.
While non-pharmacological therapies like manual and movement therapies (e.g., massage, osteopathic manipulative treatment, and shiatsu, breathing exercises, Taiji Quan, and Qigong) have been proven beneficial in preserving, promoting, and restoring health (25–28), they still lack a strong rationale (29–31). Manual and movement therapists require a better comprehension of how the body can be influenced by the stress response and immune system. Otherwise, they may lack a good grasp of these therapies' potentialities and limitations and look to the patient’s pain only in mechanical terms.
Therefore, the present paper aims to describe (i) the stress response main features and influence on the immune system; (ii) the fascia as connective tissue, explaining the behavior of fibroblasts and myofibroblasts (32); (iii) the main interactions between stress, immunity, and fascia (Figure 1).
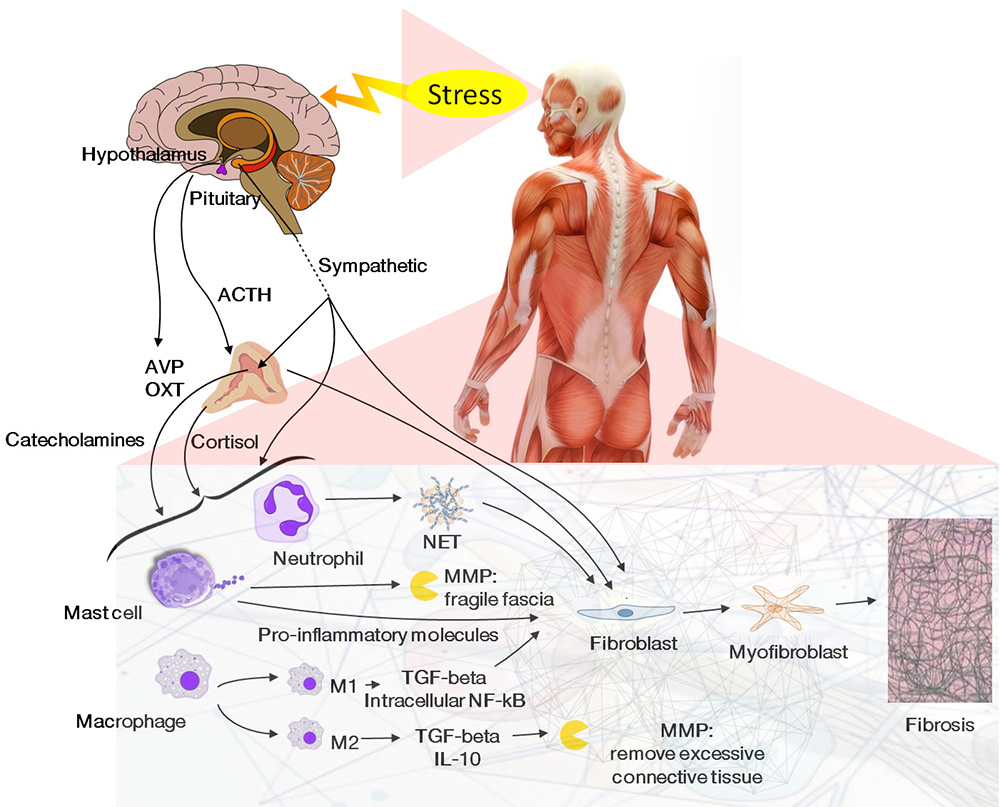 Figure 1
Figure 1The relationship between stress axis, immune system, and fascia: the activation of the HPA axis stimulates the adrenals to release catecholamines and cortisol. These hormones, together with the activation of the sympathetic nervous system caused by stress, stimulate immune cells inside the fascia. Based on the intensity of the stimulus, macrophages, mast cells, and neutrophils release molecules that can influence the structure of the fascia.
The term “stress” was used for the first time by Walter Bradford Cannon (33), whereas the existence of the HPA axis was theorized by Hans Selye (34). Selye viewed stress as the essence of life since the activation of the HPA axis allows living beings to respond to endogenous and exogenous stimuli, such as changes in hydration or tissue pH, infections, environmental temperature, pollutants, etc. (34, 35). Although Selye defined the stress response as “a syndrome produced by diverse nocuous agents” (36), it turned out that the human HPA axis is mainly activated by psychosocial stimuli (37), which can dramatically affect the HPA axis and other related neuroendocrine responses via numerous molecules produced by the brain, with a heavy impact on health and longevity (19, 38–40).
The hypothalamus commands the endocrine system and releases many neurotransmitters that circulate throughout the brain and the body. Its activity can be summarized as follows (35):
1. it secretes arginine vasopressin (AVP) and oxytocin (OXT). These two hormones mainly target the kidneys (41), the brain, the uterus during pregnancy and mammary glands after labor (42);
2. it secretes hormones regulating the anterior pituitary. This part of the hypophysis then releases specific hormones to regulate the activity of gonads, uterus, adrenal cortex, liver, bones, and thyroid (43);
3. it secretes hormones and stimulates the sympathetic nervous system (SNS) to control adrenal medulla, pancreas (44), and pineal gland (45).
The stress response originates from the paraventricular nucleus (PVN) and involves several neuroendocrine circuits. The three main branches of the stress response are (35):
the HPA axis. It originates from the PVN parvocellular nuclei that release CRH, thus inducing the hypophysis to secrete ACTH (35, 46), which in turn stimulates the adrenal cortex to secrete cortisol. The parvocellular nuclei receive information about (i) the body from the nucleus of the solitary tract and the circumventricular organs; (ii) the emotional and affective states from limbic areas such as amygdalae (47), nociceptive centers, and other cortical areas (48);
the hypothalamic-SNS-adrenal medulla axis. It originates from the PVN projection neurons that send information to the brainstem, in particular to the locus coeruleus and the rostral ventromedial medulla (RVM). While the locus coeruleus represents the main noradrenergic center of the brain, the RVM nuclei control the SNS (49). The adrenal medulla can also be activated by several motor areas (premotor cortex, primary motor cortex, supplementary motor area, rostral and caudal areas of cingulate cortex), the medial prefrontal cortices (ventromedial orbitofrontal cortex), and the anterior cingulate cortex (pregenual and subgenual). All these areas are connected to the amygdalae and the hypothalamus and use the corticospinal pathway to transmit information from the brain to the organism (50). This information arrives in the chromaffin cells of the adrenal medulla and stimulates them to secrete adrenaline, noradrenaline, and a small amount of dopamine (43). This direct pathway between cerebral motor areas and the adrenal medullae allows stressors to immediately alert the muscular system (50);
the hypothalamic-posterior pituitary axis. It originates from the PVN magnocellular nuclei that receive information about the body fluids status, including blood volume, and can release AVP and OXT to regulate blood pressure, blood volume, and hydration (35).
These three axes are regulated by negative feedback: the hypothalamus and hypophysis monitor cortisol and catecholamines plasma levels via mineralocorticoids and glucocorticoids receptors (GR) (35).
Thorough knowledge of the effects of stress hormones is necessary to understand their impact on the body structure:
CRH is correlated with anxiety and motor hyperactivity. CRH can inhibit the activity of other hypothalamic hormones, i.e., TRH, increasing hypothyroidism risk; gonadotropin-releasing hormone, facilitating reproductive disorders; and growth hormone-releasing hormone, promoting growth problems in children living under high stress (43). CRH is also produced by immune cells and the autonomic nervous system (ANS). CRH has a pro-inflammatory effect, stimulating mast cells in many tissues and organs such as the gut, brain, respiratory mucosa, prostate, vagina, uterus, heart, and even fascia (51);
ACTH induces the adrenal cortex to produce cortisol, dehydroepiandrosterone, and aldosterone. ACTH can also stimulate adipocytes to produce interleukin-6 (IL-6), thus favoring insulin-resistance under chronic stress (52);
cortisol, the primary hormone released by the HPA axis. During acute stress, cortisol prepares an individual to respond to cognitive or physical stressors: it raises awareness and mobilizes energetic and muscular resources. Under chronic stress, cortisol produces several effects: (i) the dendrites in the hypothalamic and medial prefrontal cortex decrease (35) while they increase in the amygdalae – this neuronal reorganization depends on reversible epigenetic mechanisms (53); (ii) arousal of particular pro-inflammatory immune circuits (54); (iii) cardiovascular system stress, with increased risk of heart diseases (55); (iv) alteration of the circulating lipids, impacting the function of the metabolic system (56); (v) restriction of blood flow to stomach mucosa, favoring gastric ulcers (57); (vi) possible fibrosis and steatosis in the liver (58); (vii) increased risk of ocular pathologies such as glaucoma (59). Furthermore, cortisol affects bones, muscles, and fascia (9);
OXT raises during social interactions with positive effects on mood and represents the hormone of love and empathy – it helps social bonding in many kinds of relationships, such as family and friendship. It can be detected in abundance in the superior temporal sulcus and the fusiform gyrus: the former is involved in cognitive tasks, and the latter in emotion recognition from facial expression. OXT stimulates uterine muscle contraction to induce labor and regulates gamma-aminobutyric acid level to allow the brain development of newborns (35, 60);
AVP affects blood pressure and the metabolism of body fluids, and activates the HPA axis together with CRH (35, 61);
adrenaline, secreted by the adrenal medulla, is the chemical mediator of the adrenergic nervous pathways. It accelerates heart rate, increases blood pressure, dilates coronaries, elicits myocardium activity, induces vasoconstriction, stimulates liver glycogenolysis, raises glycemia, inhibits bronchial and intestinal muscles activity and has a local hemostatic and ischemia-inducing action. Adrenaline acts in particular on alpha and beta-2 receptors (62);
noradrenaline acts on the postganglionic sympathetic nerves and causing intense vasoconstriction accompanied by a rise in diastolic pressure (62). It is stored in large quantities in secondary lymphoid organs, in lymphoid tissues associated with mucosa, gut, and bronchus, and in the spleen, where the two ANS branches merge to create several integrated reflexes necessary to regulate systemic inflammation and immune response (63, 64).
To better understand the effects of the stress response on immunity, we provide a short description of the four main immune subsystems (65). This classification is, however, more conceptual than real since white cells can shift their phenotype depending on external stimuli (e.g., cytokines) and simultaneously express characteristics of two different immune subsystems (65, 66). The four main immune subsystems are:
type-1 immunity protects the organism from viruses and neoplastic cells but also produces major pro-inflammatory effects and can lead to the development of autoimmune pathologies, including type 1 diabetes and multiple sclerosis. The type-1 immunity phagocytes clean up dying cells and debris and release free radicals and enzymes to destroy pathogens, kill infected cells, ingest dying cells and extracellular matrix (ECM). This subsystem also includes type-1 and type-3 innate lymphoid cells (ILCs), which allow tissues to repair and recover their physiological characteristics. Due to its highly inflammatory nature, type-1 immunity acts in conjunction with type-2 immunity, which is typically anti-inflammatory, to achieve optimum tissue repair (67);
type-2 immunity protects the organism from extracellular parasites, venoms, toxins, and can be elicited by antigens. This subsystem is located primarily in the gut, respiratory apparatus, and reproductive organs mucosa, where it controls eosinophils and immunoglobulins recruitment (68). Excess activity of this subsystem can lead to allergic responses (e.g., asthma and dermatitis) (68) and autoimmune pathologies characterized by antibodies production (e.g., lupus). It can also worsen neoplastic disease if present. Type-2 immunity cells, including tissue-resident type-2 ILCs (69), secrete cytokines that regulate processes such as tissue repair, metabolic and temperature control (65, 70). Type-2 immunity acts against organisms too big to be ingested by a single cell (71), whose elimination needs large tissue remodeling. Since they can heavily reshape tissues, type-2 immune cells and cytokines can induce organic fibrosis: under chronic inflammation, they elicit fibroblast activation, myofibroblast differentiation, and ECM secretion and lay down (67);
type-3 immunity acts on extracellular microbes and fungi, regulating the microbiota (72). It protects tissues from damages – by reinforcing the epithelial barriers and making them resistant to microbial activity – but can induce the high inflammation characterizing chronic autoimmune pathologies (e.g., rheumatoid arthritis and psoriasis) (67);
regulatory immunity acts via regulatory lymphocytes – they can show different phenotypes depending on the tissue and its health – to monitor the other types of immunity and control inflammation. Regulatory immunity prevents autoimmune and allergic pathologies by reducing inflammation and protects from infections and tumors by enhancing inflammation. This type of immunity can also affect neuroendocrine transmission (66, 73–75).
Many immune cells, such as natural killers (NKs), macrophages, and ILC-1, express glucocorticoid receptors on their cellular membrane to coordinate their action locally and through the reception of systemic signals (67). Many immune cells, from mast cells to naive Th0 lymphocytes, also express catecholamine receptors (e.g., alpha-2 and beta-2 adrenergic receptors) to sense signals deriving from the nervous branch of the stress response (63). Therefore, through releasing glucocorticoids in the bloodstream, the neuroendocrine system can regulate inflammation strength and duration and protect the organism.
Under acute stress, catecholamines induce immune cells to move into tissues (76) and specialize: Th0 naive cells may become Th1 (type-1 immunity), Th2 (type-2 immunity), Th17 (type-3 immunity), or T regulatory cells. B and T lymphocytes, antibodies, and NKs increase to mount an adequate response: type-1 to contrast viruses, intracellular microbes, and neoplasms; type-2 to contrast parasites and toxins; type-3 to contrast extracellular microbes and fungi (63). Catecholamines and cortisol show, instead, an anti-inflammatory action on already differentiated lymphocytes (8, 63).
Under chronic stress, the situation is far more complex (8, 63, 77):
1. cortisol and catecholamines dampen type-1 immunity and bolster type-2 immunity, thus reducing the ability to fight against viruses and neoplasms. Type-2 and regulatory cytokines such as interleukin-13 (IL-13) and transforming growth factor-beta (TGF-beta) shift from anti-inflammatory to pro-inflammatory and can elicit tissue texture alterations (e.g., demyelination) and fibrosis (the TGF-fibrosis link is discussed in sections 5 and 6). Since type-2 immunity recruits and activates eosinophils, basophils, mast cells, and IgE, allergic responses can manifest (65, 66, 75, 78);
2. the chronic SNS activation can induce the down-regulation of beta2-adrenergic receptors, which favors the hyper-production of inflammatory cytokines such as TGF-beta, and the hypo-activation of the parasympathetic nervous system, which then fails to activate the anti-inflammatory splenic ANS reflexes (79). The HPA axis hyper-activation can make lymphocytes glucocorticoid-resistant due to the high level of circulating glucocorticoids – cortisol loses the ability to suppress type-1 immunity and inflammation, thus facilitating autoimmune diseases (3).
The connective tissue represents one of the four main organic tissues of the body and is essentially represented by the ECM and the cells contained therein (80).
ECM allows connective tissue to perform its various functions: organ shaping, support, protection, communication, etc. Since every cell resides inside the ECM, the form and function of every cell depend on the ECM (80).
The main ECM constituent is procollagen, which becomes collagen after several post-translational enzymatic reactions. Collagen is a protein formed by a triple helix of polypeptides, each constituted by the alternation of three amino acids: glycine, proline, or lysine (81). Collagen formation needs the reaction of glycosylation, which attaches a carbohydrate to the side chain of its constituent amino acids. If an excess of glycosylation occurs, the structure of collagen may become unable to perform its functions (81): both connective tissue and bones may thus be affected, losing elasticity and robustness. This phenomenon may occur in the presence of hyperglycemia: the quantity of carbohydrates circulating in the bloodstream and tissues is so high that glycosylation starts spontaneously, without the intervention of enzymes. This process is called “glycation” and causes the formation of “advanced glycation end-products” (AGEs), which can accelerate aging and degenerative processes (82). AGEs can build up in connective and fascial tissues and cause both inflammation and structural and functional disorders – collagen turnover seems rather slow, showing a half-life of 4-12 months in the liver but of 215 years in the intervertebral discs of an elderly man (83, 84).
Collagen makes up about 30% of the whole-body protein content and can be classified in more than 28 types of different fibers (81). Since every fiber has different characteristics, the collagen composition of the ECM changes according to the stress, especially mechanical, it undergoes. For example, tendons, menisci, and the annulus fibrosus of the intervertebral discs are formed by type I collagen, which is very well suited to resist traction, whereas articular cartilages and the nucleus pulposus are formed by type II collagen, which is very proficient in resisting compression (85–87). Mechanical stress enables the differentiation of connective tissue at any age, and different combinations of compressive and tensional forces induce the formation of different collagen fibers so that connective tissue can better adapt to everyday mechanical stimuli. Therefore, physicians, physical therapists, and manual therapists like osteopaths and chiropractors should comprehend the biomechanical role and function of every collagenic structure, so as to properly choose the optimal kind of force to elicit a good structural and functional recovery (85–87).
ECM also contains elastic and reticular fibers. Reticular fibers constitute the stroma of hematopoietic and lymphoid tissues (thymus excluded), but they are replaced by stronger collagen fibers in the case of wounds (80, 88). Elastic fibers form the ECM of various tissues, such as smooth muscles, especially the vascular ones – elastic fibers constitute 50% of a blood vessel –, lung epithelium, skin, elastic cartilages, and even some tendons and ligaments (about 2-5% of their mass) (80, 81, 89).
ECM is also made of ground substance, an amorphous, aqueous, and viscous gel-like substance rich of long molecular chains: glycosaminoglycans (GAGs), proteoglycans, and multi-adhesive glycoproteins (80). These molecules intertwine with collagen fibers, several proteins located in the cellular membrane such as integrins, and the cytoskeleton to create a reticular structure that shapes and protects the cells it surrounds. This complex structure allows the exchange of substances and the communication between cells, relying upon integrins because of their particular sensitivity to mechanical stimuli: integrins can transduce external mechanical forces in intracellular biochemical signals that can alter gene expressions – therefore, mechanical stimuli can induce epigenetic effects (89).
Proteoglycans and GAGs can absorb compressive shock in tissues such as cartilages and intervertebral discs, protecting the body from sudden or excessive mechanical stress. When proteoglycans link to the water attracted by GAGs, the connective tissue becomes viscoelastic and the collagen fibers can efficiently slide on one another. GAGs and proteoglycans that contain hyaluronic acid prevent undesired movement of substances among tissues: indeed, molecules such as growth factors must be confined within the specific tissue that needs them. They also prevent the infiltration of microbes or neoplastic cells, which have thus to catabolize the ECM to get through the connective tissue (81, 89).
At last, ECM contains multi-adhesive glycoproteins (e.g., fibronectin and laminin) that link all the other fibers and interact with integrins and laminin receptors, thus stabilizing the ECM. Multi-adhesive glycoproteins can convey mechanical forces through the ECM to the cytoskeleton (89) and receive a direct stimulation during physical activity and manual therapies – these mechanical stimuli affect the synthesis of these proteins and collagen (90, 91).
All ECM components are equally important for a healthy connective tissue: given the role of the ECM at the structural, nutritive, and immune level, any disorder affecting one of its components can have serious consequences for the organism (92). In particular, in recent years, mechano-biology has shown that the ECM tensional status has a significant impact on cellular differentiation and proliferation. For example, a mesenchymal stem cell can become a neuron, an adipocyte, a skeletal myocyte, an osteocyte, or another kind of cell depending on ECM rigidity: a low ECM rigidity (low mechanical tension) brings about a neuron, while a high rigidity (high mechanical tension) an osteocyte (93).
Besides, recent studies have discovered that many physiological functions depend upon the mechanical dimension. For instance, sleep and fever stimulate immune response via mechano-sensitive proteins: fever elicits the expression of heat shock proteins on lymphocytes to make them link with other cells’ integrins (94), whereas sleep prevents the accumulation of Gas-coupled receptor agonists, immunosuppressive proteins that block lymphocytes integrins (95).
The ECM and its components derive from the cells that live inside the connective tissue and, in particular, from fibroblasts that regulate ECM structure, function, and turnover (80).
Fibroblasts are highly heterogeneous and show different secretory capabilities: some fibroblasts produce only type I collagen, while others only type III collagen (96). They are fusiform and flat cells, lacking a basal membrane and equipped with several processes, one or two nuclei, prominent rough endoplasmic reticulum, and Golgi apparatus. Their cytoskeleton displays an abundance of intermediate filaments mainly constituted of vimentin, actin, and myosin, which allows them good motility (97, 98). Fibroblasts that reside in different organs can be very dissimilar: fibroblasts found in the heart (97–99) differ from those in the lung (100, 101), peritoneum (102), and elsewhere (99).
Each fibroblast promptly responds to various inputs, either mechanical (e.g., compressive and tensional forces or shear stress due to fluid flow) or chemical (e.g., cytokines, hormones, or hypoxia), thereby influencing the behavior of adjacent cells (100). In the presence of chronic inflammation or repeated wounds, all fibroblasts can promote fibrosis since they are able to secrete a high amount of collagen and substances, including vascular cell adhesion molecules and intercellular adhesion molecules, that tie to leukocytes and increase inflammation (100). As an exception, oral fibroblasts cannot induce fibrosis nor scar tissue: they are similar to fetal fibroblasts, which secrete the highest amount of anti-inflammatory cytokines as compared to all the others (103).
When the organism needs to produce more ECM, for instance during growth or wound repair, fibroblasts increase their cytoplasm to develop larger rough endoplasmic reticulum (protein synthesis) and Golgi apparatus (vesicular transport). As a consequence, they become “activated fibroblasts”: they secrete a larger amount of proteins, GAGs, and procollagen, in particular for the formation of type I and III collagen. Activated fibroblasts also produce fibronectin, periostin, metalloproteinases (MMPs), and pro-inflammatory cytokines such as IL-6, TNF-alpha, TGF-beta, and endothelin-1, which have a local autocrine or paracrine action. In remodeling the ECM, fibroblasts are aided by various other cells, such as myocytes and endothelial cells, and especially by immune cells: neutrophils, macrophages, and mast cells (80, 90).
Leukocytes produce functional proteins such as MMPs, growth factors and cytokines, whereas fibroblasts, myocytes, and endothelial cells secrete mainly ECM structural proteins (90). Conversely, the ECM and the proteins dispersed in it condition the phenotype and activity of fibroblasts. For example, a glycated ground substance due to hyperglycemia increases fibroblasts apoptosis and reduces their ability to proliferate and migrate. It cuts the synthesis of collagen, MMP-1, MMP-2, and hyaluronic acid and slows both angiogenesis and immune cells recruitment – this undermines healing (96).
Fibroblast activity is also affected by extracellular pH: if it falls below 6.7, as in case of neoplasms or ischemia, the circadian cycle of fibroblast becomes accelerated – they start living days of 22 hours (104), which makes them more susceptible to endogenous and exogenous toxins and more prone to induce inflammation (105). Moreover, extracellular acidity can increase the activity of proteases (e.g., MMPs) that can remodel connective tissue during acute stress but induce inflammation and tissue destruction in the long run. Acidity can also bring about deformed, elongated cells, facilitate the activation of integrins, elicit cellular adhesion (e.g., between endothelium and neutrophils) and slow cellular migration rate (106). Large changes in extracellular pH, whether towards acidosis or alkalosis, strongly reduce GAGs secretion (107) and modify several cells such as pericytes (contractile cells located inside blood vessel walls), adipocytes, endothelial, and epithelial cells, transforming them into fibroblasts via the phenomenon known as epithelial-mesenchymal transition (EMT) (96) (Figure 2).
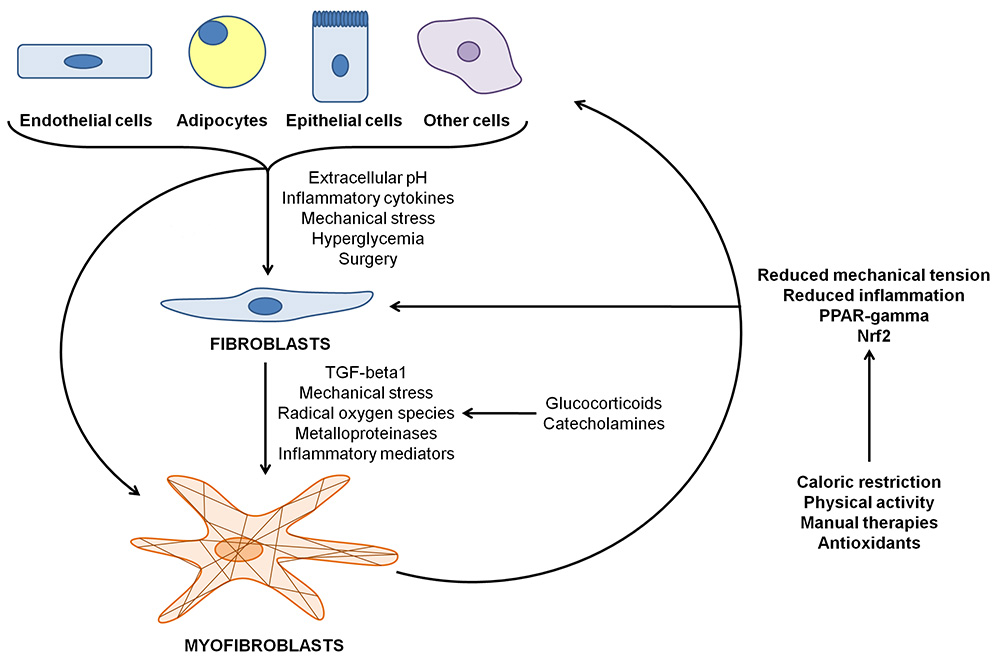 Figure 2
Figure 2Cells can change their phenotypes when stimulated. Endothelial, epithelial, and mesenchymal cells can become fibroblasts due to the effect of inflammatory cytokines, mechanical stress, and extracellular pH; other stimuli such as hyperglycemia and surgery can also induce this phenotype shift since they increase inflammation and mechanical stress. These same stimuli – in particular, TGF-beta1, mechanical stress, and ROS – can elicit fibroblasts and many cells to become myofibroblasts: they can produce contraction since they show a high concentration of alpha-SMA fibers. Fibroblasts and myofibroblasts appear when the organism needs to increase the production of extracellular matrix and to heal a wound: psychophysical stress can facilitate the induction of these cells through the secretion of cortisol and catecholamines, which can increase inflammatory cytokines and ROS. Myofibroblasts can de-differentiate back into their original phenotype through interventions that can reduce the mechanical stress and the inflammation level, or that can activate factors such as PPAR-gamma and Nrf2: some example are caloric restriction, physical activity, manual therapies, and antioxidants.
EMT can be both a physiological (e.g., wound healing) or a pathological process (e.g., fibrosis and neoplasms), and it is facilitated by pro-inflammatory molecules such as NF-kappaB and TGF-beta – TGF-beta strongly induces fibroblast activation and fibrosis (108, 109) – and under conditions such as hyperglycemia and surgical interventions (102). The cells transformed via EMT, however, may return to their original state once the stressor who induced EMT passed (96).
Among the cells that appear through EMT, we can find the myofibroblasts (110) (Figure 2).
Myofibroblasts derive from fibroblasts that developed a cytoskeleton similar to that of myocytes and can be induced by mechanical stress, cytokines, and other stimuli. Myofibroblasts express fibers of alpha-smooth muscle actin (alpha-SMA) similar to those of myocytes, which allow them to produce strong cellular and tissue contractions, which compact collagen and ground substance every time new ECM is secreted (98, 99). These contractions put myofibroblasts in a pre-stress status to better react to internal or external mechanical stimuli (by comparison, the sound produced by a violin depends on the tightness of its strings). When myofibroblasts are in a pre-stress status, also the fascia is in a pre-stress status (111, 112).
Fibroblasts at rest possess actin and myosin filaments, but can’t create cell-cell or cell-ECM adhesions and sustain a high mechanical load: the ECM protects them from excessive mechanical stress (108, 113). Under mechanical or chemical stress, fibroblasts start producing beta-/gamma-cytoplasmic actin filaments and fibronectin and acquire the capability to create focal adhesions, a particular kind of cell junction, and to link with the ECM – proto-myofibroblasts are born (108, 110, 113, 114). If this stimulation continues, as in the case of tissue lesions or chronic inflammation, integrins located in the cellular membrane of proto-myofibroblasts induce the release of TGF-beta1 from the large latent complex (LLC). Proteolytic enzymes such as MMPs, plasmin, thrombin, and elastase can also free TGF-beta1 by “digesting” LLC (115, 116).
TGF-beta1 is usually an anti-inflammatory cytokine similar to IL-10, but it can induce a type-3 immune response, high inflammation, and fibrosis (117) – TGF-beta1 seems the major stimulus for myofibroblasts to differentiate and produce contractions (118, 119). TGF-beta1 increases the secretion of tissue inhibitors of metalloproteinases, which raise connective tissue mechanical tension (110), and induces proto-myofibroblasts to produce alpha-SMA (114) and to create very strong cell-ECM anchoring junctions called fibronexus. When ECM stiffness goes beyond 20kPa, alpha-SMA and beta-cytoplasmic actin filaments wire together and create the typical cytoskeletal structure of myofibroblasts (80, 108, 113, 115, 120).
Whereas muscles produce “contractions” that are fast but energetically expensive, myofibroblasts elicit “contractures” that are slow – they take 30 minutes to reach their maximal intensity –, less energetically expensive, and able to remodel the ECM (113). Various chemical factors, including pH, can induce the production of these contractures by myofibroblasts: an acidic environment increases their responsivity, both to contract or to relax (121). Myofibroblasts seem to be independent of the nervous system (112): it appears they are stimulated to contract by mechanical stimuli and several molecules, but not by electric signals. Acetylcholine, adrenaline, and adenosine have no effect on human myofibroblasts (121); histamine and oxytocin can induce brief contractures, nitric oxide (NO) makes them relax, and mepyramine, a first-generation antihistamine, causes slow and sustained contractures for about 2 hours (112).
Although contractures can be induced by both histamine and an antihistamine, their duration and intensity is different, as well as the resulting outcome. In the case of tissue lesions, for example, histamine favors the production of collagen and alpha-SMA via signaling pathways between mast cells, fibroblasts, and myofibroblasts – wound healing improves (122, 123). Manual therapies that facilitate wound healing can indeed induce mast cells to degranulate many molecules, including histamine (124). On the contrary, several classes of anti-inflammatory drugs, including antihistamines, have deleterious effects on wound repair, in particular in the case of lesions involving tendons and ligaments (85).
Since myofibroblasts are “activated fibroblasts,” they appear especially during wound repair (80), a process that can follow two different paths:
1. a physiological path: whenever new ECM is secreted, remodeled, vascularized, and the wound is healed, myofibroblasts undergo apoptosis. Contractures are maintained by a dense reticulum of collagen that replaces alpha-SMA filaments – the ECM becomes more robust (113);
2. a pathological path: myofibroblasts survive and continue to produce ECM and contractures. High inflammation can occur, become chronic and cause fibrosis: this results in wide and hypertrophic scarring of the tissues that lose their functionality (80, 125). Examples of fibrosis are Dupuytren syndrome and scleroderma, besides the classical ones that affect the pulmonary, renal, cardiac, and hepatic organs (108, 110).
In the latter scenario, inflammation also elicits severe mechanical stress – for example, a pulsating and steady fluid flow – that activates myofibroblasts and prevents their apoptosis (120, 126). The greater the stress, the more severe the contraction brought about by myofibroblasts on the ECM, and the stiffer the ECM, the more intense the production of contractures by myofibroblasts. Therefore, fibrosis is determined by a vicious cycle (115), and a correct treatment should act by both regulating inflammation and reducing mechanical tension (108). For instance, it has been shown that splinting a wound increases the granulation tissue made by myofibroblasts, whereas removing the splint induces the disappearance of that tissue and the apoptosis of its cells (127).
Myofibroblasts may also derive from many cells other than fibroblasts, especially of mesenchymal origin (115) (Figure 2). Due to EMT, smooth muscle cells, pericytes, bone marrow fibrocytes, Kupffer cells, renal mesangial cells, hematopoietic cells, and other cells may lose their phenotype and become myofibroblasts (32, 99, 108, 110, 128). Since EMT is facilitated by mechanical stimuli, pro-inflammatory cytokines, and acidic extracellular pH, as well as by other stimuli such as hyperglycemia and surgical interventions (102, 108, 109), acting on these variables could reverse the myofibroblasts to their original phenotype.
Indeed, even though myofibroblasts are usually considered incapable of changing their phenotype, some authors showed they can lose alpha-SMA and “recover” their original phenotype, whether epithelial or mesenchymal (115, 129, 130) (Figure 2). One potential method could be to induce the anti-inflammatory nuclear receptor PPAR-gamma (129) through, for instance, caloric restriction, physical activity, antioxidants such as resveratrol, probiotics and a healthy microbiota (131). In particular, antioxidants such as capsaicin (the bioactive compound of chili pepper), grape seed extract (GSE), and superoxide dismutase (SOD) can indeed induce the de-differentiation of myofibroblasts: SOD down-regulates TGF-beta1 expression, whereas capsaicin and GSE reduce the inflammation induced by COX-2 and NF-kappaB. Capsaicin also up-regulates PPAR-gamma (129, 132). Other antioxidants such as sulforaphane, the bioactive compound of cruciferous vegetables, can de-differentiate myofibroblasts through the activation of the transcription factor Nrf2 (129).
Manual therapies could also help to revert myofibroblasts: a moderate (20-30% of maximal load) manual stimulation applied to a scar has been shown to decrease TGF-beta1 and collagen production (125). An in vitro simulation of manual medicine treatment showed an increase of TGF-beta3 as compared to TGF-beta1 (133): TGF-beta3 reduces the production of TGF-beta1 and promotes wound healing (134). Besides, many studies showed that manual and mind-body therapies can increase vagal tone (25), which regulates inflammation, and remodel fascia (135) via mechano-transduction pathways such as integrins (136), even in the case of adhesions, fascial fibrosis, and scleroderma (137, 138).
During acute stress, the modifications of the body structure aim to deal with the actual stressor and to elicit adaptation (139, 140).
Under chronic stress, this adaptation could become highly expensive for the fascial system, since chronic stress is often accompanied by chronic inflammation (7, 8) and the immune system principally acts in the connective tissue (80). Fibroblasts, myofibroblasts, adipocytes, mast cells, lymphocytes, and vascular cells can release pro-inflammatory cytokines such as IL-1, IL-6, and TNF-alpha, which increase the production of MMP, proteases, and reactive oxygen species (ROS) (92, 141, 142). We can summarize the effects on the body in two different scenarios:
1. cytokines and enzymes can fragment the ECM network, thus weakening the connective tissue (141). Prolapses and tissue permeability may emerge because the fibers become unable to bear the weight of organs and the epithelial barrier is damaged (143, 144). The facilitation of microbial and neoplastic growth and tissue infiltration may be another undesired result (81);
2. the excessive amount of ECM components excreted by the cells fills more and more space, replacing local tissue (epithelial, muscular, or nervous) with connective fibers and inducing fibrosis (32, 108, 110).
These effects can come from a sustained cytokine increase of only two to three times their normal value – defined “low-grade chronic inflammation” (LGCI). Contrary to acute inflammation, which displays a 100-fold and fast cytokines rise, LGCI takes time to alter organic functions and induce symptoms, however when they finally manifest, inflammation has been acting for a long time (144). LGCI can be avoided or limited through a lifestyle based on healthy nutrition and adequate physical activity; the two latter factors could contribute to LGCI more than any other – the consumption of high-glycemic load foods and exercising beyond one’s capacity, for example, are recognized as highly pro-inflammatory (145, 146).
Besides inflammation, the stress hormones strongly affect the ECM and the status of the fascial system, muscles, and bones (9, 142).
Under acute stress, cortisol activates type-2 immunity with anti-inflammatory effects and cooperates perfectly with mechanical stimuli in remodeling muscles and bones to ensure that these tissues can adapt to everyday stimuli and become stronger. On the one hand, glucocorticoids raise protein degradation in muscles, activate osteoclasts in bones and reduce the blood and water flow through bone canaliculi. On the other hand, the mechanical stimuli elicit muscle contractions that augment that same flow inside bones, blunt osteoclasts activity and induce new bone mass secretion by osteocytes (139, 140, 147). Moreover, even if cortisol reduces intestinal absorption of calcium and increases its renal excretion, the parathyroid glands secrete parathyroid hormone (PTH) to increase the production of vitamin D, which raises both calcium and phosphorus absorption. In order to prevent hyperphosphatemia – dangerous for both brain and heart – bones secrete fibroblast growth factor 23 (FGF23) that increases renal phosphorus excretion (140).
Under chronic stress, cortisol excess inhibits the WNT signaling genetic pathway, which is fundamental for bone development, and increases FGF23 production, which can lead to renal and cardiovascular diseases, thymus atrophy, asthenia and mental status alteration (140). Cortisol excess induces high inflammation and type-3 immunity that greatly activates osteoclasts – which are macrophages (148) – through IL-17 production, thus increasing bone fractures and osteoporosis risk (149). Cortisol excess also prevents bones from producing osteocalcin – a hormone that affects the whole organism. Indeed, osteocalcin (i) increases the number and size of pancreatic cells and facilitates insulin secretion; (ii) induces adipose tissue to release the anti-inflammatory adiponectin and augments insulin-sensitivity and energetic metabolism; (iii) induces testicles to produce testosterone; (iv) stimulates the brain to produce monoamines to protect from anxiety and depression and improve learning abilities and memory (150). After the organism recognizes a sudden danger, osteocalcin can even activate the fight-or-flight stress response in case of adrenal insufficiency (151)!
Even LGCI, hyperglycemia, and obesity can blunt the secretion of osteocalcin (152). Luckily, vitamin D, calcium, and especially vitamin K increase its production and activation (153). There are two main forms of vitamin K, i.e., phylloquinone (K1) and menaquinone (K2), and it seems K2 has a greater protective effect on bones than K1 (153). Good sources of vitamin K1 are green leafy vegetables and brassicas, whereas good sources of K2 are egg yolk, fermented foods (sauerkraut, cheese, fermented soybean), a good microbiota, liver, and butter (153).
Under chronic stress, cortisol induces muscles to produce myostatin that degrades proteins and causes the atrophy of type IIx fast-twitch fibers, which allow muscular strength (139). Cortisol can also induce tendon atrophy since it is a catabolic hormone. Indeed, chronic production of cortisol up-regulates MMPs expression in fibroblasts, facilitates ECM degradation, and reduces the concentration of important growth factors – MMPs can cleave growth factors. Since MMPs are pro-inflammatory, cortisol can greatly modify the connective tissue inside the organs and alter organic functions (154, 155). For example, in most cases, the so-called tendinitis is tendinosis with poor inflammatory infiltrate and highly degraded collagen. Therefore, therapeutic exercise – in particular, eccentric loading –, relative rest, and posture correction can manage these tendinopathies better than local glucocorticoids, which can worsen the situation (156, 157).
At last, cortisol excess causes the growth of adipocytes, with subsequent accumulation of visceral and intramuscular fat that alters the functioning of the involved organs and leads to LGCI (158).
Under chronic stress, catecholamines can have deleterious effects on the body structure. Noradrenaline excess due to high sympathetic tone elicits macrophages to produce ROS, especially superoxide via NADPH-oxidase (142, 159). It also down-regulates beta2-adrenergic receptors (79), induces the differentiation of Th17 immune cells (63) and increases MMPs secretion (160), all effects that lead to a TGF-beta1 increase. ROS and TGF-beta1 can cause tissue damage and induce fibroblasts to differentiate into myofibroblasts (116). Since these cells are not under the direct influence of the nervous system, TGF-beta1, ROS, and MMPs could constitute the link between SNS and myofibroblasts (113, 118, 160, 161).
During acute stress, the release of ROS, MMPs, and TGF-beta1 is extremely helpful: these molecules sustain wound healing and protect from infections (96). But the connective tissue remodeling and the appearance of myofibroblasts induced by chronic SNS activity can cause fibrosis, tissue adherences, and scars, which become even larger and stiffer due to myofibroblasts contractures (113, 118). These mechanical tensions can reach the cellular nuclei, where they alter the expression of several genes, change the cellular biochemistry and increase the production of pro-inflammatory cytokines and growth factors, which can predispose to neoplastic proliferation (113).
Wound healing is a perfect example to understand the different effects of catecholamines under acute versus chronic stress. Usually, adrenaline slows the activity of epithelial cells and induces fibroblasts to proliferate, migrate to the wounded area, change their cytoskeleton, and secrete collagen to heal the lesion. After the stress response stops, the decrease in adrenaline switches off fibroblasts and allows epithelial cells to complete wound healing (162). Under chronic stress, adrenaline excess blocks the migration of epithelial cells towards the wounded area and alters the fibroblasts production of collagen via beta-adrenergic receptors. Adrenaline excess can either inhibit fibroblasts growth or induce the differentiation of myofibroblasts that elicit strong contractures in the lesion surrounding tissues, thus impairing the wound healing process (163, 164).
Since growing evidence shows the presence of sex hormone receptors in the myofascial system – we can find estrogen receptors in synoviocytes, anterior cruciate ligament fibroblasts, and in the blood vessel wall cells of this ligament (165–167) – we choose to dedicate a paragraph to their interaction with fascia.
In vitro, physiological doses of estradiol reduce collagen synthesis and fibroblasts' proliferation by more than 40% in the anterior cruciate ligament. Estradiol inhibits collagen synthesis induced by physical activity through insulin-like growth factor-1 reduction (165). During the follicular phase of the menstrual cycle, estrogens modify the collagen structure and induce an increase in ligament laxity. These hormones also elicit fibroblasts to produce collagenase, an enzyme that degrades collagen and facilitates organ prolapse and ligament rupture in case of excess exposure to estrogens (166). Therefore, clinicians must investigate whether their patients with chronic pain are under hormonal therapies, including contraceptive pills – their assumption being associated with a high risk of low back pain, bone fractures, persistent pelvic pain, and articular pelvic instability (167).
Androgens can elicit leukocytes to produce TNF-alpha, which induces fibroblast to slow down wound healing and collagen deposition. But androgens seem to induce the migration of epithelial cells (especially keratinocytes) towards the lesion site, as if trying to re-epithelialize the wound before the underlying connective tissue has been repaired. We could argue androgens facilitate wound healing from the outside in – internal mucosa appears to be healed better by androgens than by estrogens, maybe because it is highly vascularized (168, 169). As a complement to androgens, estrogens speed up the connective tissue healing in case of cutaneous wounds, and also decrease inflammation and ameliorate the symptoms of diseases such as psoriasis or melanoma (170). However, estrogens – and progesterone – induce the production of type-2 cytokines (171): in case of excessive production or external stimuli such as contraceptive pills, estrogens could thus favor LGCI.
Maybe due to this reason, estrogens correlate with cellulite, which is a typical female phenomenon, even though it can manifest in men with low or null levels of androgens, and can be linked to inflammation (172, 173). Indeed, cellulite seems to manifest when estrogens increase more than progesterone, which naturally happens during puberty, pregnancy, and menopause, and forcefully under contraceptive pills or hormonal replacement therapy (172). Estrogens increase MMPs, whose physiological effects on collagen constitute the origin of cellulite, according to some authors (174). That some degree of cellulite could be natural and beneficial is supported by the evidence that it appears primarily in the typical female adipose tissues, especially gluteofemoral, which function as a reserve for pregnancy and breastfeeding to obtain energy, steroids, etc. Therefore, although not all women have cellulite, it could represent a greater reserve (174).
Cellulite becomes a problem when the organism's health changes (173): inflammation and stress can indeed strongly affect cellulite through MMPs (173, 174). Besides, whereas gluteofemoral superficial adipose tissue (SAT) usually shows a high production of the beneficial adiponectin (175), that same tissue expresses a low adiponectin level in the case of a large amount of cellulite (176). Gluteofemoral SAT affected by cellulite displays damages in the vascular system due to high pressure, which is both high blood pressure due to increased levels of angiotensin II and high mechanical tension due to adipocytes’ growth (173). This results in water and GAGs retention, tissue hypoxia, loss of elastic fibers (177), and more inflammation (176).
Moreover, estrogens increase HPA axis reactivity: since the stress response can increase steroidogenesis, a vicious cycle between psychosocial problems and tissue alterations could occur (178). In this scenario, prolactin secretion could be involved (177) since cellulite shows high inflammation and a large amount of adipose tissue, which can be both induced by prolactin (177, 179). Prolactin allows breastfeeding and is released in large amount during orgasm, but it also increases in combination with several psychosocial stressors such as anger associated with humiliation, loss, neglect, social loneliness, or the feeling of having to take care of lonely others (e.g., little children who lost their parents) (180). Therefore, reducing the burden of these stressors could help to resolve cellulite. Although, as far as we know, no previous research has investigated the association between psychosocial stress and cellulite, these correlations should be evaluated in as complex a paradigm as PNEI.
To show how immune cells can impact the fascia, this section describes the behavior of some specific white cells – macrophages, neutrophils, and mast cells – during acute and chronic responses and their interaction with the ECM.
Macrophages balance the equilibrium between wound healing and fibrosis and regulate inflammation affecting the behavior of fibroblasts, epithelial (e.g., mucosal and cutaneous), and white cells (181). They can circulate (monocytes) or reside in tissues (actual macrophages): since they regulate tissue integrity and the growth of cells and blood vessels, their absence in tissues can cause serious problems in organic growth. It’s worth noting that cells such as osteoclasts and microglia are macrophages (148).
Macrophages show two phenotypes that can link to the four immune responses, although they only represent the two extremes in a continuum of different phenotypes with intermediate characteristics (148, 181, 182).
1. the first phenotype (M1) is associated with type-1 and 3 immunity. M1 macrophages stimulate fibroblasts to transform in myofibroblasts and induce wound contraction and collagen production. They favor myofibroblasts survival via TGF-beta secretion and activate the pro-inflammatory transcription factor NF-kappaB. M1 macrophages secrete antimicrobial molecules as a first-line defense against infection and recruit other white cells;
2. the second phenotype (M2) is associated with type-2 and regulatory immunity. M2 macrophages promote inflammation resolution and tissue healing via leukocytes apoptosis and anti-inflammatory cytokines such as IL-10, which regulates MMPs secretion, reduces ECM production, and allows the removal of connective tissue in excess. M2 macrophages also compete for nourishment (e.g., arginine) against other white cells and myofibroblasts, thus reducing the activity of the latter.
These two phenotypes can self-regulate: for instance, M1 Kupffer cells (the hepatic macrophages) infected by already neutralized microbes undergo apoptosis and release IL-33 and IL-4, which polarise nearby macrophages towards M2 phenotype. The result is good healing of hepatic tissues (181). Although M1 facilitates tissue fibrosis, in a healthy organism M1 is regulated by M2, and thus tissues can heal (148, 181). On the contrary, when an organism displays a hyperactive HPA axis, a high sympathetic tone and LGCI, both M1 and M2 macrophages recruit other leukocytes – in particular, Thelpers and ILCs – and favor fibrosis: M2-secreted IL-13 and TGF-beta stimulate the production of MMPs and activate myofibroblasts (181).
M1/M2 polarization depends on the surrounding environment: it can be induced by both chemical (e.g., cytokines, growth factors, and hormones) and mechanical stimuli (182, 183). All white cells are mechanosensitive, and macrophages are no exception: if ECM is under normal tension, macrophages assume an elongated shape whereby actin is more dispersed at the cell periphery so that anti-inflammatory M2 polarization occurs. Whenever fascia is under high tension (e.g., stiffness due to myofibroblasts contractures, fibrosis), macrophages assume a round shape whereby F-actin accumulates around the nuclei, so that M1 polarisation occurs – stiffness facilitates macrophages attachment and spreading. However, if macrophages are stretched by nearby cells, they can shift towards the M2 phenotype (183). Therefore, as with myofibroblasts, regulating inflammation and mechanical tension become paramount.
Neutrophils represent the major leukocyte population and the first to reach wounded tissues, where they activate and sustain inflammation (65). Neutrophils are constantly produced by the bone marrow and, in the case of chronic inflammation, they can become the main regulators of innate and adaptive immune responses (65, 184). Neutrophils phagocytize debris, degranulate antimicrobial molecules and release opioids in wounded tissues having an analgesic effect (185). Like macrophages, neutrophils show two phenotypes – i.e., the inflammatory N1 and the anti-inflammatory N2 – and recruit lymphocytes via the release of cytokines, alarmins, and cellular DNA. Neutrophils can also dampen tissue damages capturing danger-associated molecular patterns (e.g., uric acid) through releasing “neutrophil extracellular traps” (NETs) (185).
NETs are decondensed chromatin meshes bound to DNA, histones, granular antimicrobial peptides, and enzymes such as cathepsins and elastase, which block and destroy pathogens (185). When produced in excess, NETs can cause conditions such as thrombosis, arteriosclerosis, and autoimmune diseases since they contain inflammatory and auto-antigenic substances such as free cellular DNA (186). Indeed, several autoimmune pathologies that affect connective tissue – e.g., rheumatoid arthritis, lupus, vasculitis, and dermatitis – show a high neutrophils activity (184, 185, 187). A possible reason that explains connective tissue involvement in such pathologies is that neutrophils release NETs after tissue trauma and lesions – even of surgical nature (186).
The signals inducing NETs include ROS, cytokines, and molecules secreted by activated epithelial cells, which are more prone to release pro-inflammatory factors and interact with leukocytes – thus, activated epithelial cells can become myofibroblasts (186). Therefore, the stress response could heavily influence neutrophils behavior. Indeed, physical stress such as moderate exercise can help to control neutrophils activity since it releases DNase, an enzyme that reduces free DNA concentration – it regulates both NETs formation and degradation and decreases the risk of thrombosis (188). On the contrary, an intense physical activity would release a great amount of ROS, with a consequent increase in neutrophils tissue infiltration, inflammation, and pain perception (188). Regulating neutrophils activity should be of the foremost importance since neutrophils can also resolve inflammation, elicit revascularization and limit the onset of fibrosis by excreting IL-1 receptor-alpha, TGF-beta, and specific lipids that mainly originate from omega-3 fatty acids – these lipids inhibit neutrophils afflux and polarise immune cells towards an anti-inflammatory phenotype (184, 185, 189, 190).
Mast cells are greatly influenced by the stress response and appear in a high amount in connective tissue (80). Mast cells can provoke a fast inflammatory response due to the ready-to-be-released pro-inflammatory molecules they contain, i.e., histamine, serotonin, IL-1, IL-6, TNF-alpha, IFN-gamma, TGF-beta, proteases, prostaglandin, leukotrienes, growth factors, NO, heparin, adenosine triphosphate, and calcitonin gene-related peptide (191). Mast cells can be activated by many stimuli, in particular, by mechanical and psychosocial stress: a stiff and fibrotic ECM induces mast cells degranulation and proliferation (192), whereas emotional stress causes mast cells to increase inflammation and the symptoms of allergic conditions such as asthma (193, 194). Mast cells are part of the type-2 immune subsystem, which can be induced by glucocorticoid and catecholamines (195) and, in case of danger, by chemical signals deriving from afferent nerve pathways (e.g., wounds, infections) – it’s the “neurogenic inflammation” (196).
Connective tissue mast cells are different from other mast cells because they contain both tryptases and chymases (80). While tryptases stimulate fibroblasts to produce more proteins and recruit neutrophils and basophils, chymases convert angiotensin I into angiotensin II facilitating hypertension, have anti-parasitic effects and degrade several growth factors (141). Both tryptases and chymases degrade the ECM to remodel it (141) – for example, warm-up exercises increase temperature, activate muscle metabolism and release tryptases and chymases to soften the myofascial system and facilitate the physical effort. These enzymes can also cleave pro-inflammatory molecules (e.g., cytokines, P substance, and endothelin I), thus lowering inflammation: mast cells can indeed blunt neutrophil tissue infiltration through heparin and histamine release (141).
A chronic mast cell activation can make fascia fragile and permeable through releasing MMPs, but it can also induce fibroblasts to transform into myofibroblasts, thus facilitating fibrosis (122, 197, 198). However, without mast cells, the immune system could not mount a timely reaction against infections and venoms (141). Balanced nutrition and healthy gut microbiota greatly help to harness optimal mast cell activity since they can both regulate mast cell degranulation: antioxidants and commensal microbes can protect from ROS, venoms, and pathogens without activating an inflammatory response, thus leaving the mast cells to degranulate only when truly needed (199, 200).
Glucocorticoids and catecholamines can differently affect connective tissue, muscles, and bones. Their effects can be direct or mediated by molecules such as cytokines, MMPs, and ROS or by the recruitment of immune cells such as T lymphocytes, macrophages, neutrophils, and mast cells. In all cases, an organism induces the production of glucocorticoids and catecholamines and activates the immune cells when it is striving to cope with the environment. Hence, these molecules have a purpose and help the organism to remain healthy and become stronger (e.g., the body gains strength and health after a good physical training) (146).
However, the duration of the stress response and the immune activity can bring about deleterious effects on tissues: chronic stress and inflammation can induce muscles and tendons atrophy, induce connective tissue and organ fibrosis, cause osteoporosis and bone fractures, increase fluid retention and inflammation (e.g., cellulite) and facilitate adipocyte growth. These effects can have systemic consequences since they can influence the production of many signal molecules – for example, osteoporosis is linked with reduced osteocalcin secretion (150), adipocytes can induce hypertension (158), and MMPs can reduce growth factors (155) – and thus threaten health.
Therefore, strategies that limit the deleterious effects of chronic stress and inflammation – in particular, anti-inflammatory diet, correct physical activity, good sleep hygiene, social support, and stress relieving – are paramount to regulate for the better the interaction among these systems (3, 7).
The present paper described how stress response and the consequent nervous, endocrine, and immune activations can modify body structure – myofascial system and bones – and how body structure (e.g., the mechanical characteristics of ECM) can influence those same activations. The analysis was based on a systemic approach, i.e., the PNEI paradigm, and aimed to encourage research in this complex field, in particular, in the field of manual and movement therapies.
Papers similar to the present one can help manual therapists in better understanding their practice, as emphasized by the following examples. During the manual palpation of a scar, therapists often feel tissue relaxation, expansion and “breathing,” and myofibroblasts usually manifest a rhythmic contraction every 99 seconds – curiously, this rhythm matches the 100s time defined as “life breath” or “long tide” by Sutherland in the osteopathic field (118). This rhythm is due to intracellular Ca2+ oscillations and increases with inflammation: therefore, myofibroblasts produce faster contractures during inflammation (201). But a manual stimulus can reverse this acceleration via the relaxation of adherens junctions between myofibroblasts (and likely via the reduction of inflammation) (25), thus restoring a slower rhythm – this change can feel like a “tissue release” (118).
In-depth knowledge of the connective tissue can also assist in selecting the most appropriate physical loads and forces to be applied to restore the correct collagen structure after a lesion: different loads induce the production of different types of collagen. Similarly, knowing the mechanisms by which wound healing is affected by health, stress response, inflammation, and manual therapies can be useful for therapists to comprehend which procedures could obtain the best outcome. This information could also help surgeons, improve aesthetics medicine and promote integrative medicine in the context of preemptive healthcare and in the management of patients with chronic musculoskeletal pathologies.
Knowing the relationship between stress, immunity, and fascia helps to understand that the outcome of medical and manual therapies can be boosted or curtailed by the person’s experiences: any beneficial effects of medical and manual treatments can be nullified by psychosocial adversities and lifestyle (e.g., physical inactivity, high glycemic index, and load nutrition) that trigger high inflammation. Notwithstanding that manual therapies can have systemic effects – they can reduce cortisol, SNS activation, and inflammation, increase oxytocin secretion and vagal tone and remodel collagen (11, 25, 29–31, 202) – they remain an isolated event in everyday life that must be integrated with other interventions such as patient education, physical exercise, lifestyle changes, and psychological therapies (as shown by the recommendation in the case of low back pain) (203).
Pain in itself, as a perception, does not strictly depend on local structural alterations but originates from cerebral elaborations of every endogenous and exogenous information, the result of which affects the organism via nervous, immune, and endocrine pathways (10, 202, 204). Inflammation, emotions, beliefs, expectations about pain and body movements, and lifestyle can all greatly impact pain, and knowing their influence can help to assist the patient in coping with pain (202, 205).
Many physicians and manual therapists often treat the painful area or other body structures that are physically connected to it, but this can be noxious if there is no real lesion, because it can reinforce the belief that a lesion does exist, and consequently heighten the patient’s fear and avoidance of healthy movements (205). Physicians and manual therapists should evaluate every possible structural lesion and act accordingly, but should also address the patient’s biopsychosocial needs (1, 206). Since pain is a central response to psychophysical danger (10), the organism can keep producing pain as long as it detects a threat – physical wounds, bad digestion, environmental toxins, inflammation, infections, poor sleep, loneliness, low self-efficacy, fear, low socio-economic status, and so on. Only by addressing all these factors, pain can be managed – our goal should be to understand why the organism thinks it needs pain, and find strategies to support the person in everyday life (10, 204, 205).
For instance, since psychosocial stress can induce fascial alterations and alter immunity responses, a therapy that ignores stress management might never resolve fascial alterations. The organic effects of stress can be countered through pharmacological, surgical, or other interventions; however, this would not address the initial cause – stress – putting the person at risk of developing other pathologies. Indeed, as long as it detects a threat to its integrity, the organism can ceaselessly continue to elicit the stress response (1, 18, 206–209). This reasoning holds for all health conditions (1, 18, 206) and can make us better comprehend what occurs inside patients, why sometimes nothing seems to relieve their pain or condition (205, 206).
More research is required to show the relationships between organic systems, between an organism and its environment, and to define a strong rationale of clinical practice. Too often, we are forced to rely on weak hypotheses in explaining what is happening during therapy, and this increases the likelihood of making an error. This fact is especially true in the fields of manual and mind-body therapies, which struggle in fully satisfying evidence-based medicine criteria (e.g., it is difficult to devise a placebo for manual treatment). Overcoming these limitations through sound biomedical research, both quantitative and qualitative, will improve therapies, healthcare, and health services to augment people’s health and quality of life.
Every author equally contributed to this article. We are grateful to Silvia Clara Tuscano for revising the manuscript.
HPA
hypothalamic-pituitary-adrenal axis
corticotropin-releasing hormone
adrenocorticotropic hormone
arginine vasopressin
oxytocin
sympathetic nervous system
thyrotropin-releasing hormone
paraventricular hypothalamic nucleus
rostral ventromedial medulla
interleukin
autonomic nervous system
extracellular matrix
innate lymphoid cell
natural killer
T helper lymphocyte
interferon
transforming growth factor
advanced glycation end-product
glycosaminoglycan
metalloproteinase
epithelial-mesenchymal transition
nuclear factor kappa B
smooth-muscle actin
large latent complex
nitric oxide
proliferator-activated receptor
nuclear factor erythroid 2-related factor2
grape seed extract
superoxide dismutase
cyclooxygenase
reactive oxygen species
low-grade chronic inflammation
fibroblast growth factor
nicotinamide adenine dinucleotide phosphate
superficial adipose tissue
type-1 macrophage
type-2 macrophage
deoxyribonucleic acid
neutrophil extracellular trap.