
Frontiers in Bioscience-Elite (FBE) is published by IMR Press from Volume 13 Issue 2 (2021). Previous articles were published by another publisher on a subscription basis, and they are hosted by IMR Press on imrpress.com as a courtesy and upon agreement with Frontiers in Bioscience.

1 University Guglielmo Marconi, University of Salerno, Italy
Abstract
Gastrointestinal (GI) health of newborns is maintained by the gastrointestinal microbiome, and the PNEI (psychoneuroendocrinoimmunology) system and can be altered by the delivery mode of the milk to the newborn such as breast versus bottle feeding, exposure to antibiotics in the milk, or exposure to environmental pollutants. The health of GI tract is particularly is maintained by breast feeding since the maternal milk provides micro- and macronutrients essential to growth, as well as multifunctional small oligosaccharides, particularly, 2’-fucosyllactose which maintain the GI health.
Keywords
- 2’-fucosyllactose
- 2’FL
- Human Milk Oligosaccharides
- HMOs
- Breast Feeding
- Review
Psychoneuroendocrinoimmunology (PNEI) is a discipline investigating the bidirectional relationships between human psyche and the biological systems (1). This relationship can strongly influence the overall health status of a human being. Starting from this assumption, and in order to contextualize the gut in relation to PNEI, the following have to be examined: connections with the biological systems, the psyche, the environment, and food coming in direct contact with the gut since the early stages of life. In recent years, an increasing interest has arisen on the connections between the central nervous system and enteric nervous system. This has forced to re-evaluate the perception of the relationships between emotional and cognitive centers in the brain and intestinal gut functions. The reciprocal connections between these two organs can also be mediated through biochemical signals transmitted by the microorganisms themselves residing in the gastroenteric tract. For this reason, the intestinal microbiota, or gut flora, has been defined as the “forgotten organ” (2).
This communication network includes the central nervous system (CNS), including the brain and spinal cord; the autonomous nervous system (ANS); the enteric nervous system (ENS); the immune system (IS); and, finally the hypothalamic pituitary adrenal (HPA) axis. This latter communication pathway, whose main actor is cortisol, can be activated by environmental factors, emotions, or stress, all of which are able to change the composition of gut flora regardless of stimulus duration (3).
The gastrointestinal tract differs from all other peripheral organs in that it has a widespread intrinsic (or enteric) nervous system containing 200-600 million neurons distributed into thousands of small ganglia. This articulated neural network allows ingested food to advance from the esophagus to the anus with peristaltic movements (determining progression and mixing). It has an autonomous organization (peristaltic movements are not “decided” by the brain), and communicates with the brain through the vagal nerve and sympathetic system (4).
Recent research has highlighted how the microbial ecosystem can also influence communication between central and enteric nervous systems by using both signals coming from the microbiota itself, and signals coming from the brain. This transmission exploits the neural, humoral endocrine, and immune networks (5). One of the determining factors for adequate gut colonization is maternal milk. It stimulates proliferation of a well-balanced and diversified microbiota (6). Maternal milk can influence the transition from a Th2 dominance during intrauterine life to a balanced Th1/Th2 response with Treg cell activation by specific organisms stimulated by maternal milk action (7). Bacteria are not only able to stimulate lymphoid elements, but also the components of the epithelial layer forming the mucosal barrier, thus stimulating the development of intestinal microvilli (8) in the above mucus layer (9) and tight junctions (10). The gastrointestinal mucus system is the first line of defense against bacteria, and its organization changes throughout the gastrointestinal tract. The main elements making up the mucus are mucins. These are highly glycosylated proteins generally consisting of more than 80% carbohydrates with an amino acid-rich protein core where serine, proline, and threonine are dominant (called PTS sequences). If mucus is not perfectly healthy, or has any kind of defects, it can determine a pathophysiological condition leading to gastrointestinal problems (11).
Breastfeeding is defined as the Gold Standard in newborn nutrition. It can be considered as an extraordinary “living tissue” changing its composition during feeding to satisfy the baby’s needs. These needs go well beyond the mere nutritional aspect. Maternal milk also contains cells and immune cytokines, and growth and gene regulation factors (fig. 1) (12).
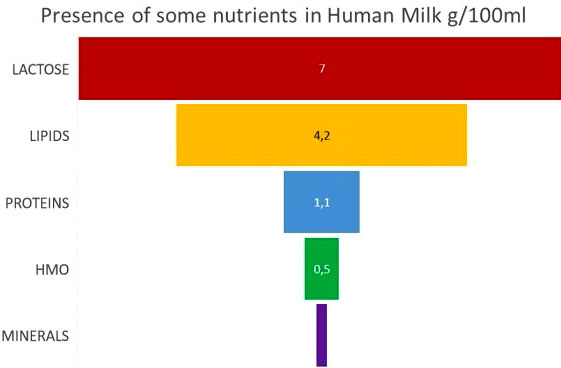
Quantitatively, the most abundant class of substances after water and lactose are small oligosaccharides called HMOs (Human Milk Oligosaccharides). 5-15g/L can be detected in colostrum and mature maternal milk (13-14-15). In human maternal milk, about 150-200 different HMOs can be found, whereas only approximately 30 are present in cow milk (15-16). Only 1-2% of these short sugar chains are absorbed by the baby. Since the baby does not have the enzymatic makeup to digest them, they reach the systemic circulation. The remaining ingested HMOs reach the colon, where they are metabolized by the gut flora (15-16-17). Although the percentage of systemic absorption is very low, it still seems to perform a neurotrophic action by promoting regulatory factors such as brain derived neurotrophic factor (BDNF) (15). The presence of HMOs’ metabolites in the excretory apparatus could also somewhat have a role in modulating the genito-urinary microbiota of the newborn.
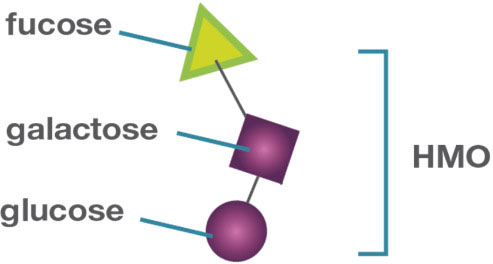
HMOs are natural bioactive substances that are synthesized by the human mammary gland. Their complex structure essentially consists of 5 elemental blocks that are differently linked together. There are: Glucose, galactose, N-acetylglucosamine, fucose, and sialic acid. Fucose is an unusual sugar as it has an L -configuration, whereas the other sugar molecules in the body have a D-configuration (27). Essentially, HMOs are based on a basic lactose structure. They however differ for the decoration in their terminal positions. Elongated or differently branched structures can also occur, thus creating a series of HMOs with different architectures and molecular weights (15-20). Fucosylated HMOs are the main components (about 77% of HMOs), whereas sialylated HMOs account for about 16% of all HMOs. Fucosylated HMOs are neutral molecules, whereas sialylated HMOs are acidic molecules. To simplify, one can classify HMOs based on their terminal sugar: Fucosylated HMOs such as 2’FL; non-fucosylated HMOs such as LNnT; and sialylated HMOs such as 3’SL (15-20). 2F’ lactose is the most represented HMO in maternal milk (fig. 2) with a percentage above 60% of all maternal HMOs (15-20).
For a long time, scientists have wondered about the biological meaning of these oligosaccharides, as they are non-digestible for the newborn. Nonetheless, the fact that breastfeeding is linked to a reduced risk of gastrointestinal conditions and infections in the newborn stage, as well as in the following stages of life, is widely recognized (21-22). HMOs could be partly responsible for this protective effect, as maternal milk has shown to support immune defenses in the child (18-23-24-25-26). The type of terminal sugar influences the types of pathogens or toxins that can adhere thereto. There is actually a lectine-glycane-kind binding specificity (27).Researchers have discovered that the kind and amount of HMOs in maternal milk is related to maternal ABO blood group and Lewis antigens (28). HMOs bind to specific receptors on immune cell surface such as type C lectins, Siglecs, Galectins, and Selectins, thus modulating immune response (81).
HMOs ingested with maternal milk or as a novel food in formulas for babies, or as supplements for adults remain unmodified through the gastric barrier since they are resistant to hydrochloric acid and pepsin. They are not modified by pancreatic enzymes and bile salts. Only a percentage around 1% is absorbed in the small intestine. HMOs can be found in the feces and urine of babies and adults who took them (29).
The relative abundance of fucosylated HMOs depends on the activity of transferases glycosylating lactose or transmembrane mucines. These fucosyltransferases are coded by genes such as FUT2. About 80% of Italian women are secretors, i.e. they are able to produce and secrete a wide abundance of 2’FL and other HMOs in maternal milk (30-31-32). Limiting polymorphism of FUT2 gene doesn’t have implications for females only (in relation to maternal milk production). Non-secretory FUT2 polymorphism in males as well as in females, will also result in less fucosylation of intestinal transmembrane mucines with negative effects on microbiota equilibrium and immune system maturation (33). A recent study HMO profile showed to modulate the infant gut microbiota response to fortificant iron; compared to infants of secretor mothers, infants of non-secretor mothers may be more vulnerable to the adverse effect of iron but also benefit more from the co-provision of GOS (34).
Beneficial effects of HMOs can be grouped into 2 classes, namely Effects related to prebiotic action and Specific immunomodulatory and regulatory effects (35-36) (tab. 1).
In the past thirty years, there is enough evidence to show that breastfed children have a more stable microbiota with superiority of bifidobacteria in the first week of their life. This relative superiority penalizing other groups such as bacteroides is undoubtedly favored by fucosylated and syalilated oligosaccharides in maternal milk (44-45-46-47-48). Further evidence of this is the fact that children being breastfed by non-secretors, develop a more varied bacterial flora; the lack of a relevant superiority of bifidobacteria can be observed as the increase of Streptococcus and other potential pathogens (48-49).
One of the most important mechanisms by which the selective action of HMOs on children’s microbiota is exerted is through the trophic action on some bacterial groups that are able to digest HMOs. These bacteria create a complex trophic network in which some bacteria internalize and digest them, others externalize the enzymatic digestion makeup providing monosaccharides to other strains. The final product of this trophic network is the production of satisfying amounts of SCFA which favor the development of a healthy microbial environment and exert a regulatory action on the gut-brain axis and other metabolic regulation axes (lipolysis) (48-49).
Children’s microbiota and immune system are both immature at birth, and have a critical development period during the first 1000 days of life. In this period, trophic microbiota conditions and interactions between the microbiota and gut-associated lymphoid tissue (GALT), can influence the health condition of an individual all through his/her life (15-49-50).
An important evidence of the protective function of HMOs in the child come from studies where children being breastfed with milk rich in fucosylated HMOs have developed less diarrhea episodes due to Campylobacter jejuni or enterotoxigenic E. coli. These babies anyway had less diarrhea episodes for all causes (51-52).
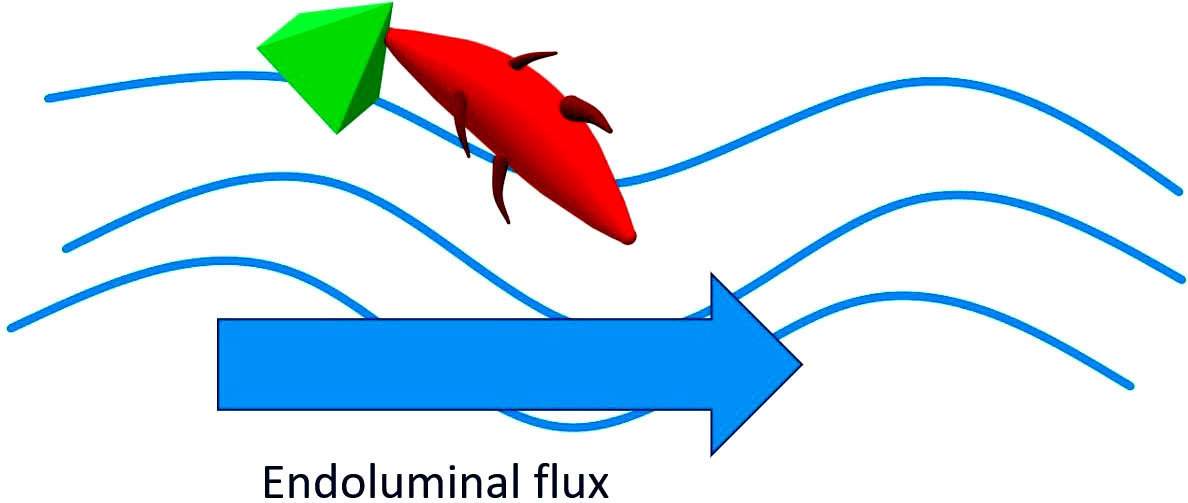
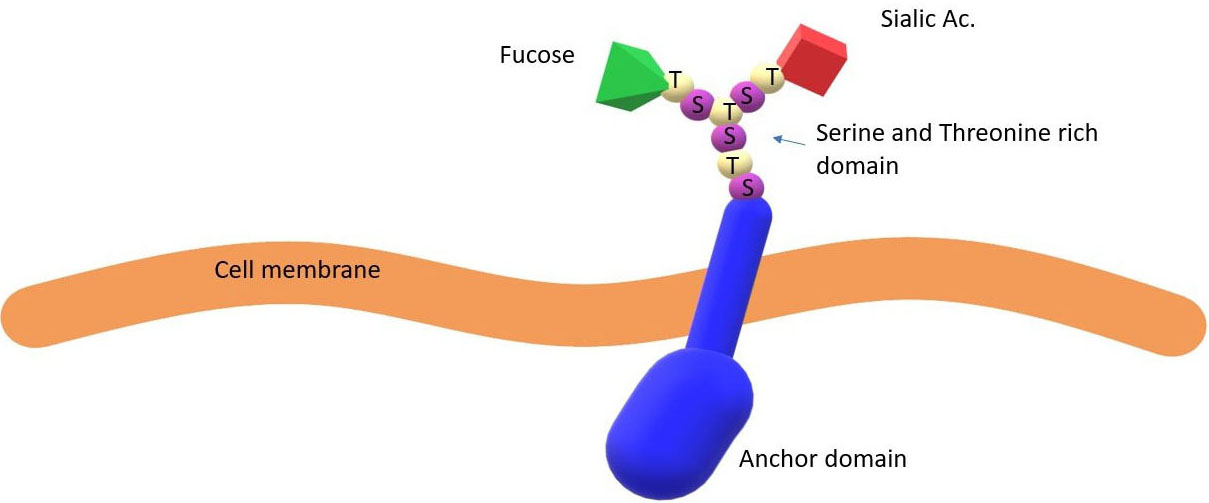
HMOs are able to prevent attacks to epithelial cells in the gastrointestinal tract by toxins and pathogenic agents such as Campylobacter jejuni, Escherichia coli, Vibrio cholerae, Salmonella fyris, Helicobacter pylori, Entamoeba histolytica, and viruses (fig. 3). This process reduces the pathogenicity of such agents since the binding to receptors on epithelial cells is a prerequisite for infection (25). HMOs resemble glycans of transmembrane adhesion proteins (lectins and mucins) to which pathogens must adhere to colonize and infect the host. Due to this similarity, free HMOs in the intestinal lumen (whether coming from maternal milk or ingested as supplements) act as a selective bait for toxins and intestinal pathogens that bind to them and are then inevitably dragged along into the intestinal lumen (fig. 4) (15-25-53-54-55-56-57). HMOs can directly influence the production of proinflammatory cytokines and other immune cytokines in the intestinal mucosa (55-56-57). Early clinical evidence underline that types of milk supplemented with HMOs produce immunomodulatory effects on cytokines, which are similar to those we can find in children being breastfed by secretors, which is not the case for infants who received feeding not enriched in HMOs (58).
Several HMOs have a relevant effect one gastrointestinal motor contractions, as highlighted by an in vitro model of murine colon peristalsis. 2'-Fucosyllactose (2'-FL) and 3'-fucosyllactose (3'-FL) reduced contractility in a concentration-dependent mode (43).
Fucose and fucosylated molecules had an immediate effect in decreasing colon smooth muscle contractility within 5-10 minutes from their application. It is unlikely that these effects of HMOs occur by bifidobacteria stimulation. Rather, they represent a direct action on neuron-dependent motor complexes (43).Results of clinical studies done so far suggest a specific interaction of fucose and/or fucosylated HMOs with tissue receptors which, on their turn, regulate intestinal motility and can also show antinociceptive activity. These observations support the hypothesis that fucosylated HMOs can be useful as a therapeutic or preventative support in diseases involving gut motility or intestinal pain and can further have beneficial effects on the central nervous system.
Researchers have started to investigate the protective effect of HMOs with regard to allergies. Feeding with a milk rich in 2’FL has shown to lower the risk of eczema related to IgE and risk of allergy at the age of 2, even for infants born though Caesarian section (59-60-61). In allergic newborns, the bifidobacteria flora is very different from healthy ones. This might also be explained by a lack of supply of certain HMOs for polymorphisms of maternal FUT2 (59-62). HMOs have shown to modulate human epithelial cell responses related to allergic disease. In particular, 6-sialyl-lactose (6'-SL) could inhibit inflammatory chemokine release, which, in turn, would inhibit inflammatory cell influx into the gut, potentially decreasing food allergy symptoms. The effect of two HMOs, 2' FL and 6’-SL, has been studied on anaphylactic symptoms induced by oral ovalbumine in a murine model of food allergy (mice sensitized to ovalbumine). Daily oral treatment with 2'-FL or 6'-SL has decreased food allergy symptoms among which diarrhea and hypothermia. The results suggest that 2’-FL and 6'-SL reduce food allergy symptoms by inducing regulatory TIL-10(+) cells and indirect mast cell stabilization (61).
A very important application field for HMOs is prevention of necrotizing enterocolitis in newborns. Animal model (rat) studies are encouraging (63). A cohort study in humans showed that a particular HMO (disialyllacto-N-tetraose, DSLNT) could have a more relevant role in counteracting this severe pathology: (64).
The use of HMOs as ingredient for food supplementation in adults is very recent, and is justified by clinical studies on the efficacy and safety of its use (62).Oral 2'-FL administration to adult subjects at doses up to 20 g/day has proven definitely safe and well tolerated, and is an effective way to repopulate the amount of bifidobacteria in adults showing dysbiosis or a reduction in the amount of bifidobacteria (frequent in elderly people). In a recent study, 100 adult subjects have been randomized to 10 groups, each receiving chemically synthesized HMOs in several daily doses (5, 10, or 20 g), or 2 g glucose as the placebo for 2 weeks. Physical parameters, including pulse and blood pressure, remained unchanged during and after taking HMOs. Routine hemato-chemical tests were also stable during the study. Tolerability was good, and adverse events were mild. Sequencing analysis of 16S rRNA revealed that HMO supplementation specifically modified the gut flora in adults. The immediate effect was a substantial increase in Actinobacteria and Bifidobacterium, and a reduction in Firmicutes and Proteobacteria. Bifidobacterium increase, which was more than 25% in some subjects, was dose-dependent but independent from the initial amount (62-65). In a double-blind, placebo-controlled study, the effects of HMO supplementation in patients with IBS were studied. Sixty-one patients were randomized, and the treatment group received 5g and 10g/day HMOs for 4 weeks. This treatment significantly increased the presence of intestinal bifidobacteria without worsening symptoms (83). The same authors evaluated the effects of 5g and 10g/day of a mixture of HMOs on the fecal microbiota by means of a GA-map Dysbiosis Test, and observed, besides optimal tolerability of the prebiotic mixture, an increase in Bifidobacterium, Alistipes, Bacteroides spp. & Prevotella spp., and Parabacteroides. HMOs modify the intestinal microbiota by acting on microbial trophic network. The action is both direct and indirect, because some strains are able to internalize and digest HMOs as such. Conversely, other strains externalize digestive enzymes thus producing simpler metabolites that are made available to the microbial community. In this way, a selective trophic pressure is exerted on the microbiota, together with other modulatory mechanisms (84).
HMO showed in several studies to have a modulatory effect of infants gut microbiota increasing the alfa diversity better than other prebiotics like fructooligosaccharides and galactoligosaccharides. HMO increases the population of beneficial bacteria like Akkermansia muciniphila and reduced the population of pathogens like Pseudomonas sp. (84). In adults with IBS HMOs could be specific dietary modifiers of IBS microbiota, in particular by specifically increasing bifidobacteria. In addition, HMOs increase Alistipes, Bacteroides spp. & Prevotella spp., Parabacteroides and Firmicutes either directly or by cross-feeding (84-85).
Probiotic effectiveness of many strains is limited by conditions that are not fully known. Recent studies have shown that some individuals seem to be more permissive than others towards probiotic treatments that are able to positively modulate gut flora. The integration of a probiotic with an HMO supplement may increase the possibilities of determining positive results in secretors, and even more so in non-secretors thanks to the above described numerous bioactive mechanisms (66).
Due to the complex chemical structure and important technical production issues, the use of HMOs in the production of enriched food for infants and production of food supplements took place only in recent years. Engineering processes on E. coli allowed to obtain highly purified 2’FL starting from lactose and glucose (65). From a regulatory standpoint, only a few HMO mixtures obtained the status of Novel Food, and received authorization to be used for human feeding, supplementation and production of AMFS in Italy and Europe (69-70-71). Whether they come from chemical synthesis or fermentation, HMOs require a special authorization procedure before they can be recognized as safe substances (GRAS in the USA, or Novel Food in the EU), and used in human feeding (69-70-71). Pharmaceutical-grade production processes allow one to obtain ultrapure HMOs without allergens, bacterial antigens, and with residual lactose between 1 and 2.5%. In the future, the production quality coming from industrial ingredient purification processes will differentiate quality products from ingredients unable to guarantee effectiveness and tolerability.
Clinical studies in children and adults showed the high safety profile of HMOs, and particularly of its most abundant trisaccharide, 2’FL (72-73-74-75-76-77). The studies in adults showed an optimal tolerability profile with a daily dose up to 20g/day (62-65). The use as supplement is currently proposed at doses of 250 mg/day 1 to 4 times a day, which represents the amount that a mother would give her newborn baby during a breastfeeding day. We are particularly interested in HMOs’ effect on infant microbiota because we are aware that gastrointestinal health of adults depends on this effect. Early clinical studies showed encouraging results. Newborns fed with 2’FL- and NnT-enriched mixtures developed a similar microbiota to that of breastfed infants (76-77-78-79-80). A non-secretory genotype is known to be related to a higher risk of developing intestinal chronic inflammatory diseases such as Crohn's disease (78), the risk of developing infections (79), and type I diabetes (80). Considering its optimal tolerability (81), we’ll hopefully be able to understand and gain insight into the potential of HMO supplementation in non-secretory adults and secretors with intestinal dysbiosis.
We just started to understand the complex biological signals coming from HMOs, and how these small oligosaccharides are able to act on the biology of the whole body from a PNEI’s point of view. HMO supplementation could be useful for non-secretory breastfeeding mothers, and infants fed with formulated milk not containing HMOs. The prevention and therapeutic potential involved in an early restoration of HMOs through milk strengthening and food supplementation in adults has to be investigated. Moreover, HMOs also represent a possible tool to strategically manipulate the microbioma and microbiota for health and therapeutic purposes (66).
The potential synergies with other substances able to contribute to gastrointestinal barrier health, have to be analyzed. This barrier regulates transport and host defense mechanisms at the mucosal interface with the outside environment. Such synergies could represent a new therapeutic approach to the gastrointestinal health entailing the use of probiotics, galacto-oligosaccharides (GOS), fructo-oligosaccharides (FOS), glutamine, zinc, and bioactive anti-inflammatory and antioxidant substances able to support gastrointestinal health. Further studies are needed to understand the interactions of HMO with micronutrients and other active substances.
| HMOs Effects | Action | References |
|---|---|---|
| Effects related to prebiotic action | Exert a selective pressure on microbial equilibrium, thus fostering beneficial bacteria (bifidogenic action) and penalizing opportunistic pathobionts | (15-30-37-38) |
| Support short-chain fatty acid (SCFA) production | (39) | |
| Specific immunomodulatory and regulatory effects | Limit adhesion of pathobionts to intestinal mucosa of the baby with a selective bait action | (40-41) |
| Modulate the barrier function aiding in the maturation process of innate immunity in the child | (15-40-41-42) | |
| Directly modulate some immune mechanisms such as lymphocyte rolling/adhesion | (40-41-42) | |
| Modulate mucosal neuromuscular contractility | (40-41-42-43) |
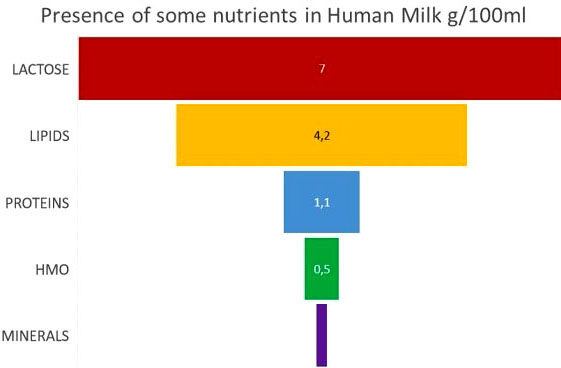 Figure 1
Figure 1Presence of some nutrients in human milk g/100 ml. Breast milk is mainly made up of macronutrients such as: lactose, lipids and proteins. In small quantities we also find HMOs and minerals.
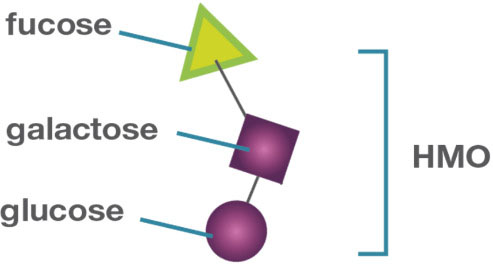 Figure 2
Figure 22’-Fucosyllactose structure. 2’FL consists of a lactose molecule (glucose and galactose), bound to a fucose molecule.
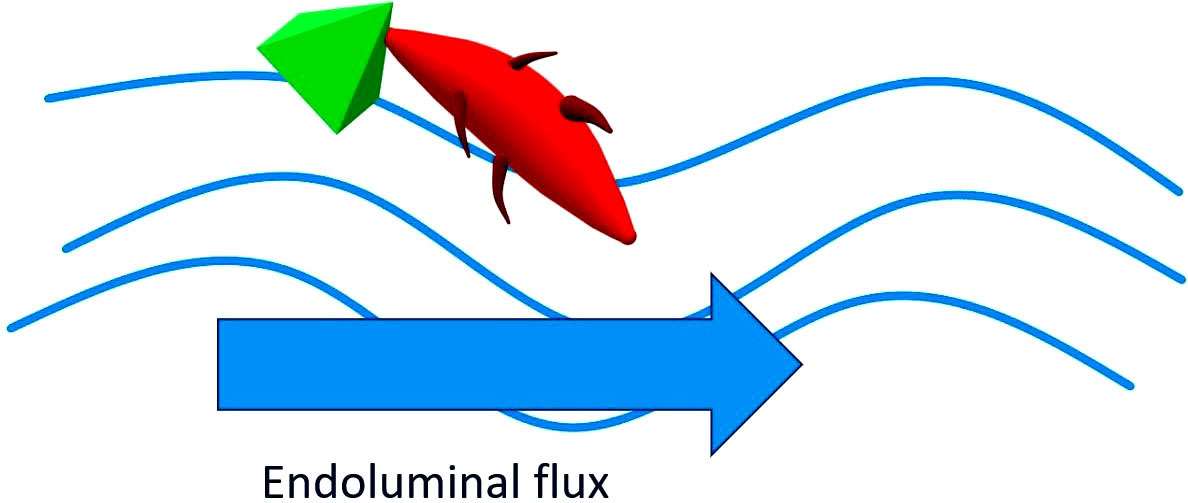 Figure 3
Figure 3Pathogens linked to HMO are then flushed away in the endoluminal flux. The HMOs resemble the structure of some glycans on the surface of the epithelial cells of the host and can therefore act as "bait" receptors able to prevent the adhesion of pathogens to the epithelial surfaces and the subsequent translocation. Once the link with the HMO has occurred, the pathogen or its toxin is dragged outwards through the endoluminar flow.
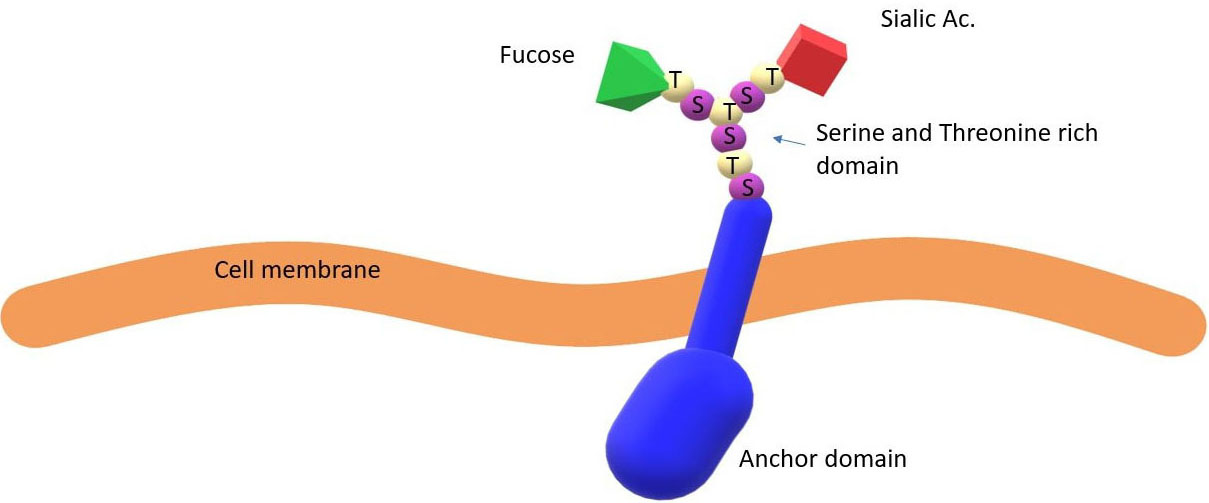 Figure 4
Figure 4Membrane glycoprotein with adhesion sites. In secretory subjects, membrane glycoproteins have binding sites decorated with a fucose molecule that represents the main binding sites for pathogens and their toxins.
GI
Gastrointestinal
Human Milk Oligosaccharide
Human Milk Oligosaccharide
2'-fucosyllactose
6-sialyl-lactose
Psychoneuroendocrinoimmunology
Central nervous system
Enteric nervous system
Immune system
Hypothalamic pituitary adrenal
Brain derived neurotrophic factor
Gut associated lymphoid tissue
Disialyllacto-N-tetraose
Galacto-oligosaccharides
Fructo-oligosaccharides
Support short-chain fatty acid