




 , Yongbiao Ai 1, Tianxiang Zhang 1
, Yongbiao Ai 1, Tianxiang Zhang 11 Department of General Surgery, The Taihe Hospital Affiliated to Hubei University of Medicine, 442000 Shiyan, Hubei, China
Abstract
Circular RNA PVT1 (circPVT1) is involved in malignant tumorigenesis. However, the function of circPVT1 in breast cancer (BC) has not been investigated. The purpose of this research is to explore the mechanisms of circPVT1 in breast cancer from different perspectives.
In this study, the expressions of circPVT1 and microRNA-30b-5p (miR-30b-5p) were analyzed by quantitative real-time polymerase chain reaction (q-PCR). The Kaplan-Meier was adopted to compare disease free survival (DFS) and overall survival (OS). CircPVT1 was depleted with special si-RNA, Cell Counting Kit-8 (CCK-8) to evaluate the cell viability and transwell assays to examine invasion ability. Astrocyte elevated gene 1 (AEG-1) protein level was measured by western blot. The competitive endogenous functions among circPVT1, miR-30b-5p and AEG-1 were verified by bioinformatics analysis and luciferase assay.
In this study, it was found that the circPVT1 was significantly elevated in BC tissues and cell lines compared to the control group. The Kaplan-Meier results indicated that higher circPVT1 expression had a worse prognosis than the low circPVT1 expression group in DFS (χ2 = 7.174, p = 0.007) and OS (χ2 = 3.946, p = 0.047). CircPVT1 regulated the BC cell viability, migration, and invasion ability. Besides, miR-30b-5p was found as a molecular target of circPVT1, and AEG-1 was identified as a target of miR-30b-5p. The suppression of circPVT1 promoted the expression of miR-30b-5p and inhibited AEG-1 expression. Moreover, simultaneous inhibition of miR-30b-5p expression in the circPVT1 knockout group could reverse the inhibition of AEG-1.
Our results indicate circPVT1 regulates AEG-1 expression by binding to miR-30b-5p in breast cancer cells. CircPVT1 promoted AEG-1 expression by inhibiting miR-30b-5p in order to enhance the BC cell viability, migration, and invasion. Our results reveal a new molecular therapy target for breast cancer.
Keywords
- circular RNA PVT1
- breast cancer
- miRNA-30b-5p
- AEG-1
Circular RNAs (circRNAs) includes exon sequences and splicing at classical splicing sites [1]. The biological functions of circRNAs are an important topic of clinical research in recent years, and function in a vital role in malignant tumor progression, acting as an oncogene or tumor suppressor involved in the malignant progression of a variety of cancers [2]. CircRNAs participate in regulating physiological and pathological processes by competitively binding to microRNAs (miRNAs or miRs) and reversing miRNA-mediated inhibition of downstream mRNA protein translation [3, 4].
Circular RNA PVT1 (circPVT1) is located on chromosome 8q24 and plays a carcinogenic role in cancers which include lung, neck, and oral cancers [5, 6, 7]. Several studies have reported that circPVT1 might function in cancer progression by regulating c-Myc protein [8, 9]. It has been demonstrated that circPVT1 regulates cellular radiosensitivity in breast cancer (BC) [10]. However, the detailed regulatory mechanism of circPVT1 in BC has not been fully revealed. This purpose of this study is to investigate the expression of circPVT1 in breast cancer and its biological roles, and to elucidate its potential biological mechanisms.
The Ethical Committee of Taihe Hospital, Hubei University of Medicine approved
the present study (NO.2024KS03). Cancer and para-cancer tissue specimens of
patients undergoing breast cancer surgery in Taihe Hospital, Hubei University of
Medicine from July 2017 to August 2019 were collected. Permission for use of
breast tissues in this research was signed by all the participating patients. A
total of 142 female patients, aged between 35–55 years old, with an average
age of 43.26
Inclusion criteria: (1) pathological diagnosis of invasive breast cancer; (2) complete clinical medical records; (3) no radiotherapy, chemotherapy or endocrine therapy administered prior to surgery. Exclusion criteria: (1) patients with other primary malignant tumors; (2) defecits in clinical data and incomplete treatment. All patients signed informed consent forms. All sample tissues were stored at –80 ℃.
The MDA-MB-231 and MCF-7 cell lines were obtained from Academia Sinica Cell Bank (Shanghai, China), MCF-10A, MDA-MB-468, T47D and BT20 were bought from Shanghai Institute of Cell Biology, Chinese Academy of Sciences (Shanghai, China) and were identified with the short tandem repeat (STR) method. The cell lines used have been tested for Mycoplasma. Cells were cultured under the condition of 37 °C with 5% carbon dioxide. Dulbecco Modified Eagle Medium (DMEM) (11965092, Thermo Fisher Scientific, Waltham, MA, USA) was added with 10% fetal bovine serum (FBS, Life Technologies, Carlsbad, CA, USA) as the culture medium, with antibiotics being streptomycin (100 µg/mL) (Beyotime, Shanghai, China) and penicillin (100 U/mL) (Beyotime, Shanghai, China).
The shRNA that specifically targets for circPVT1 (si-circPVT1) (20 nM) and a negative control that targets nothing (si-NC) were chemical synthesized by Shengong Biotechnology (Shanghan, China). The sequences of siRNA were si-circPVT1#1: 5′-GAGCTTCGTTCAAGTATTT-3′, si-circPVT1#2: 5′-GAAATGTCCTCTCGCCTGC-3′ and si-NC: 5′-UUCUCCGAACGUGUCACGUTT-3′. Lipofectamine 2000 (Invitrogen, Shanghai, China) was added for the transfer of MDA-MB-231 and MCF-7. After 48 h transfection, the silenced cells were harvested and applied for the following experiments.
The breast cancer cell lines MCF-7 and MDA-MB-231 were cultured without serum in 6-well plates. They were divided into si-circPVT1#1, si-circPVT1#2, and siRNA NC with a final concentration of 100 nM. After transfection for 24 h, 1000 cells were inoculated in 96-well plates with a volume of 200 µL per well. The light absorption values of cells were detected at culture time points 1, 2, 3 and 4 days. According to the description of CCK-8 proliferation detection reagent, each well had added 10 µL proliferation detection reagent, incubated for 2 h, and the optical density (OD) value of each well was measured at 450 nm wavelength. The growth curves of different groups of breast cancer cells were drawn.
The TRIzol reagent (Invitrogen, Shanghai, China) was adopted to extract tissue or cell RNA. The NanoDrop ND-1000 (Thermo Scientific, Waltham, MA, USA) was utilized to measure the RNA concentration and quality. The total RNA was reverse transcribed (Takara, Tokyo, Japan). Quantitative real-time PCR was completed with SYBR Green PCR Master Mix reagents (Takara, Tokyo, Japan). The primer sequences were as follows: circPVT1 forward, 5′-CTTCCTGGTGAAGCATCTGAT-3′ and reverse, 5′-TTCAGCCTCCACTTAAAGTACC-3′; and GAPDH forward, 5′-CATGAGAAGTATGACAACAGCCT-3′ and reverse, 5′-AGTCCTTCCACGATACCAAAGT-3′. The expression level of circPVT1 mRNA was relatively normalized to the GAPDH. The 2-ΔΔCt method was utilized to compare the quantification difference.
A wound-healing assay was adopted to observe cell migration. The cells were
seeded with a concentration of 5
The 8 µm pore transwell (BD Biosciences, San Jose, CA, USA) was
utilized to observe cell invasion. Briefly, 1
Sixty micrograms of total protein were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) electrophoresis and electrophoretically transferred onto PVDF (poly vinylidene fluoride) membranes. Five percent milk was adopted to block the membrane. The membrane was incubated with astrocyte elevated gene 1 (AEG-1) or GAPDH (1:1000) antibodies overnight at 4 °C. AEG-1 (1:1000) was purchased from Abcam (No. ab229128, Cambridge, MA, USA) and GAPDH was from Cell Signaling Technology, Inc. (No. #5174, Beverley, MA, USA). The proteins were observed by enhanced chemiluminescence (Millipore, Beijing, China) following the manufacturer’s procedure. Image J 1.53 (https://imagej.net/ij/) to compare western blot (WB) gray value. AEG-1 gray value is normalized to GAPDH.
The luciferase assay kit was purchased from Vigorous Biotech (Beijing, China) and the TOP/FOP Flash luciferase reporter plasmid was bought from Biovector NTCC Ltd (Beijing, China). Luciferase assays were completed following by the manufacturer’s protocol. The miRNA-30b-5p binding site sequence of circPVT1 or AEG-1’s 3′untranslated region (3′-UTR) was cloned into pGL3 vector (wild type), respectively. The 3′-UTR wild type (WT) and mutatin (MUT) luciferase reporter plasmids containing circPVT1 or AEG-1 were synthesized and transfected into MDA-MB-231 or MCF-7 cells cultured in 24-well plates. Simultaneously transfected 100 ng wild type or mutate type plasmid, 100 nm invalid nucleotide or miRNA-30b-5p in the MDA-MB-231 or MCF-7 cells were placed in 24-well plates. The luciferase assay was done after 24 hours transfection. Firefly luciferase was utilized as a reference, and Renilla luciferase was utilized as the internal control.
The 6-week-old female Balb/c mice were obtained from Charles River (Beijing,
China). MDA-MB-231 cells transfected with si-circPVT1 or si-NC and were
separately injected into the dorsum of the mice (n = 7 per group), 5 000 000 cells/200 µL
phosphate buffered saline (PBS) were inoculated. Tumor sizes
were carefully measured weekly and the formula L
Tumor tissue samples were collected, embedded in paraffin with 10% formaldehyde solution.
Antigen retrieval was enhanced by microwaving in sodium citrate solution antigen repair
liquid antigen of 98 °C for 15 minutes to repair, rabbit anti human AEG-1 (Abcam, No. ab229128, Cambridge, MA,
USA) or PCNA monoclonal antibody (Abcam, No. ab92552, Cambridge, MA, USA) (1:100) 4 °C for the night, horseradish peroxidase
labelled goat anti-rabbit second antibody (Shenggong Bioengineering Company, NO. E670020, Shanghai, China) incubation for 1 hour, colour reaction with
3,3-diaminobenzidine (DAB) (NO.DAB-0031, MaiXin Bioengineering Company, Fuzhou, Fujian, China), and then all
the sections were lightly counterstained with hematoxylin and examined by light microscopy. The results
were finally evaluated by the staining intensity and staining area score. The staining intensity was
negative to strong (0–3), negative (0 point), weak positive (1 point), moderate positive (2 points), and
strong positive (3 points). The staining positive area score was 0 (
SPSS 21.0 (SPSS Inc., Chicago, IL, USA) was used to analyse the data. The
measurement data were analyzed with t test, and the Kaplan–Meier method
was used to evaluate disease free survival (DFS) and overall survival (OS).
The Chi-square Test was adopted to compare the difference between circPVT1
expression in breast cancer patients and clinicopathological characteristics.
Values of p
CircPVT1 was expressed in 142 cases of breast cancer and corresponding
paracancer tissues. The results indicated that the relative expression level of
circPVT1 in breast cancer tissues was 4.21
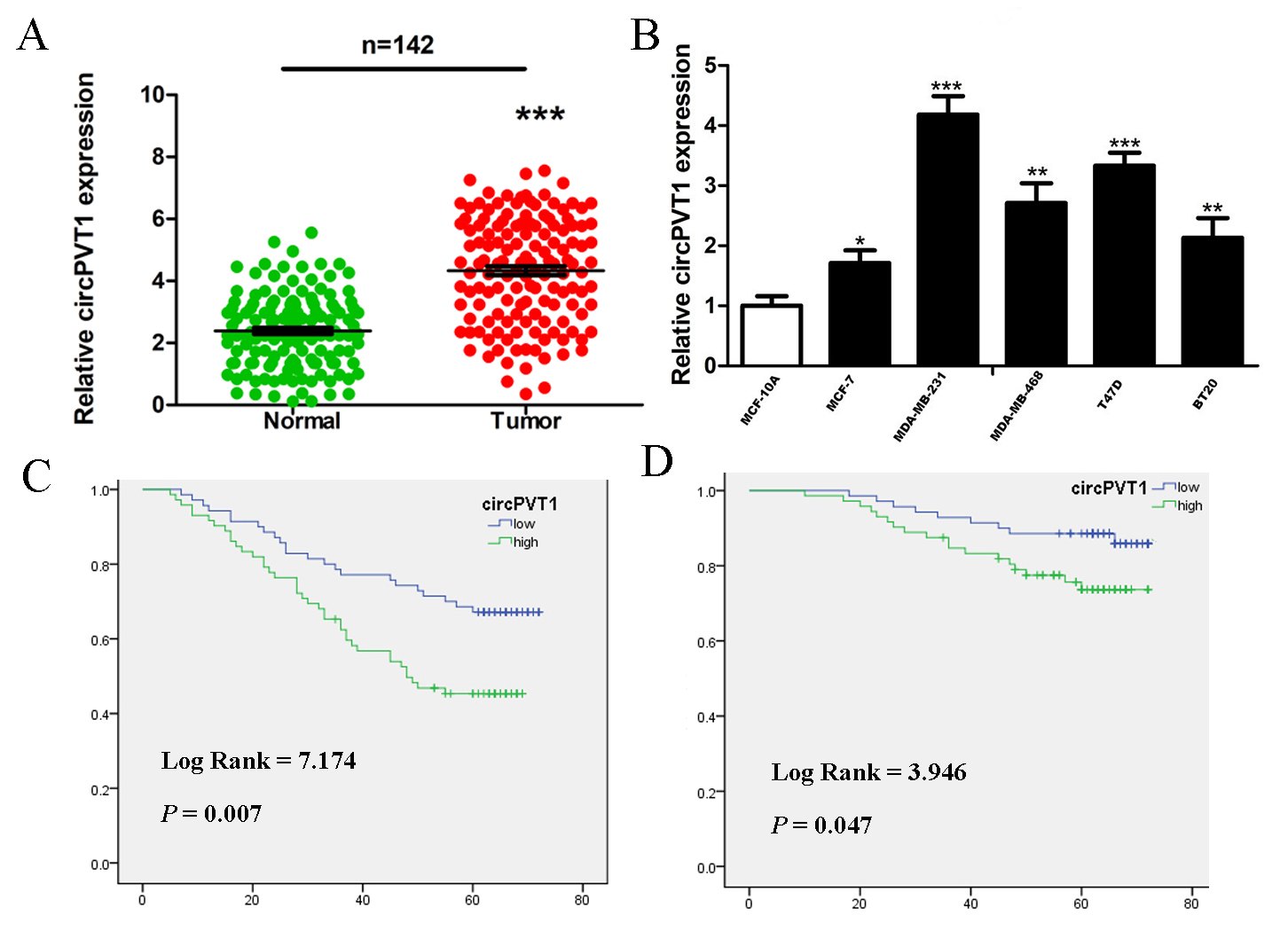 Fig. 1.
Fig. 1.
Increasing expression of circPVT1 in breast cancer tissues and
cells. (A) CircPVT1 expression was elevated in breast cancer tissues compared to
the corresponding paracancer tissues, n = 142. GAPDH was adopted as the internal control.
Data come from triplicate experiments. (B) q-PCR to detect circPVT1 RNA levels in different breast cancer
cell lines. n = 3. MCF-10A is a normal mammary cell. (C) The Kaplan-Meier results indicated
that higher circPVT1 expression correlated with poorer disease free survival. (D) The
Kaplan-Meier results indicated that higher circPVT1 expression correlated with poorer overall survival.
circPVT1, circular RNA PVT1; q-PCR, quantitative real-time polymerase chain reaction. *p
The mean expression level of circPVT1 in breast cancer tissues was 4.21. High
expression of circPVT1 was related to axillary lymph node metastasis
(
| Characteristics | circPVT1 | p value | ||||
| n | Low (n = 70) | High (n = 72) | ||||
| Age (years) | 0.031 | 0.861 | ||||
| 74 | 37 (50.0%) | 37 (50.0%) | ||||
| 68 | 33 (48.5%) | 35 (51.5%) | ||||
| Menopausal status | 0.099 | 0.753 | ||||
| Premenopausal | 81 | 39 (48.1%) | 42 (51.9%) | |||
| Postmenopausal | 61 | 31 (50.8%) | 30 (49.2%) | |||
| LN metastasis | 4.108 | 0.043 | ||||
| Negative | 75 | 43 (57.3%) | 32 (42.7%) | |||
| Positive | 67 | 27 (40.3%) | 40 (59.7%) | |||
| Tumor size | 0.236 | 0.627 | ||||
| 62 | 32 (51.6%) | 30 (48.4%) | ||||
| 80 | 38 (47.5%) | 42 (52.5%) | ||||
| ER status | 0.099 | 0.753 | ||||
| Negative | 61 | 31 (50.8%) | 30 (49.2%) | |||
| Positive | 81 | 39 (48.1%) | 42 (51.9%) | |||
| PR status | 0.001 | 0.973 | ||||
| Negative | 57 | 28 (49.1%) | 29 (50.9%) | |||
| Positive | 85 | 42 (49.4%) | 43 (50.6%) | |||
| HER2 | 0.770 | 0.380 | ||||
| Negative | 107 | 55 (51.4%) | 52 (48.6%) | |||
| Positive | 35 | 15 (42.9%) | 20 (57.1%) | |||
| Ki67 status | 0.127 | 0.721 | ||||
| Negative | 79 | 40 (50.6%) | 39 (49.4%) | |||
| Positive | 63 | 30 (47.6%) | 33 (52.4%) | |||
| Differentiation | 4.823 | 0.028 | ||||
| Well | 54 | 34 (60.7%) | 22 (39.3%) | |||
| Poor | 88 | 36 (41.9%) | 50 (58.1%) | |||
| TNM | 0.671 | 0.413 | ||||
| I, II | 101 | 52 (51.5%) | 49 (48.5%) | |||
| III | 41 | 18 (43.9%) | 23 (56.1%) | |||
circPVT1, circular RNA PVT1; ER, estrogen receptor; PR, progesterone receptor; LN, lymph node; HER2, human epidermal growth receptor 2; TNM, T: tumor, N: lymph node, M: metastasis.
In order to further study the role of circPVT1 in breast cancer, cell lines with
circPVT1 knockout were established in breast cancer cells. The cell lines with
circPVT1 knockout were successfully established by q-PCR (Fig. 2A). Further
cell count assay showed that the elimination of circPVT1 significantly inhibited
the proliferation ability of MCF-7 and MDA-MB-231 cells (Fig. 2B,C) (p
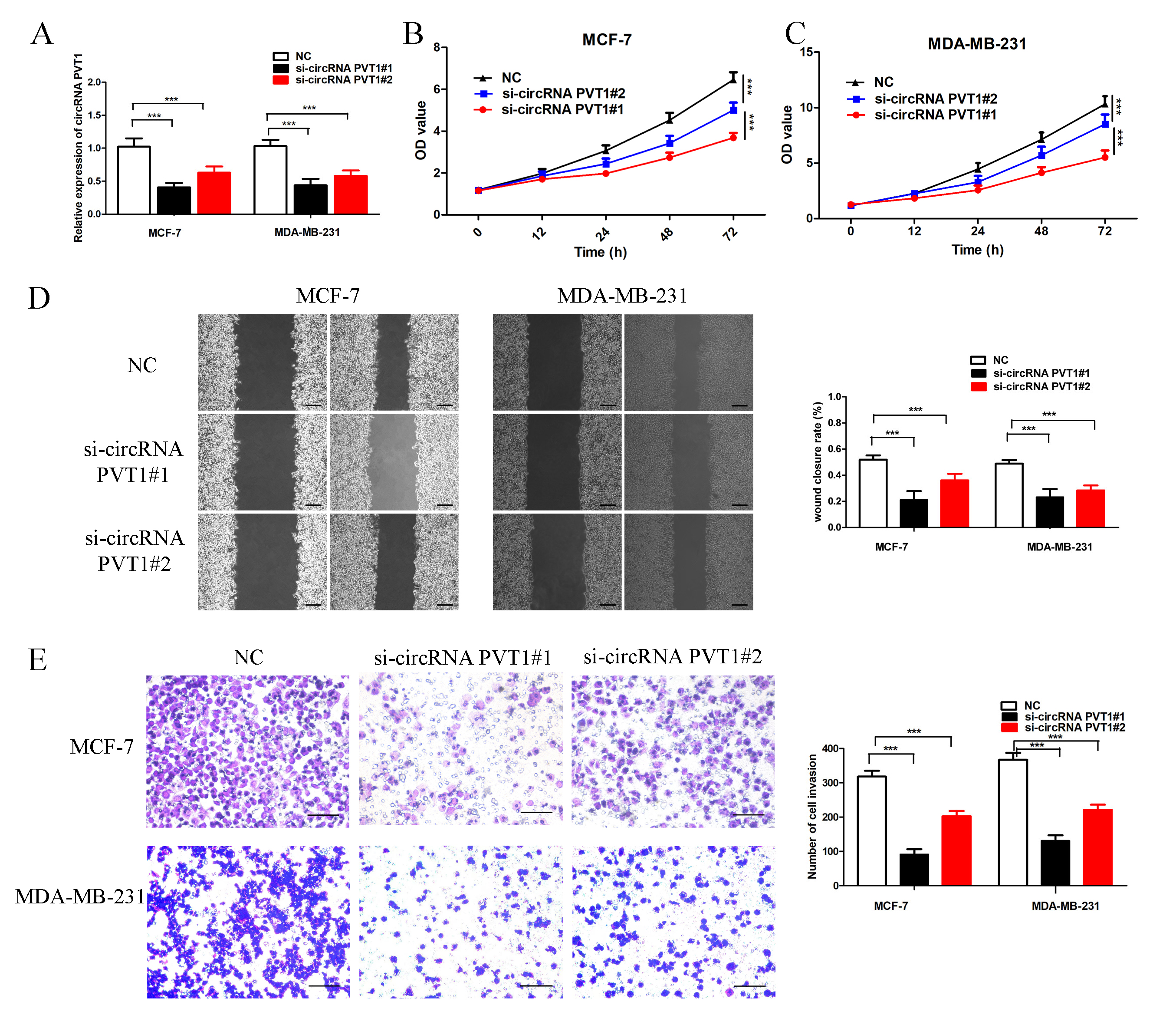 Fig. 2.
Fig. 2.
Knockout of circRNA PVT1 in breast cancer cells. (A) Knockout
of circRNA PVT1 expression compared to negative control (NC) group. (B,C) Cell
proliferation assay results suggested that knockout of circRNA PVT1 inhibited the
breast cancer cell proliferation. (D) Scratch assay suggested that knockout of
circRNA PVT1 inhibited the migration of MCF-7 and MDA-MB-231 breast cancer cells, scale bar = 200 µm.
(E) Cell invasion assay suggested that knockout of circRNA PVT1 effectively
inhibited the breast cancer cell invasion, scale bar = 100 µm. ***p
In order to further study the functions of circPVT1 on breast cancer cell biology, the cell scratch test was applied to study the migration ability. The elimination of circPVT1 significantly inhibit the breast cancer cell migration ability (Fig. 2D). The transwell invasion assay further indicated that the invasion ability of knockout circPVT1 in MCF-7 and MDA-MB-231 cells was significantly suppressed compared to the control group (Fig. 2E).
Since circular RNA can act as a miRNA sponge, by searching the circinteractome
database, we found that the circPVT1 sequence contained the binding site of
microRNA-30b-5p (miR-30b-5p) (Fig. 3A), and further confirmed the relationship
between circPVT1 and miR-30b-5p. The luciferase results indicated that
overexpression of miR-30b-5p could inhibit luciferase activity in circPVT1
3′UTR wild type group. When the circPVT1 3′UTR binding site was
mutated, the overexpression of miR-30b-5p could not effectively inhibit
luciferase activity (Fig. 3A). To confirm the reliability of our results, q-PCR
was applied to detect the expression level of miR-30b-5p in MCF-7 and MDA-MB-231
cells, miR-30b-5p expression was significantly increased after circPVT1 knockout
(Fig. 3B, p
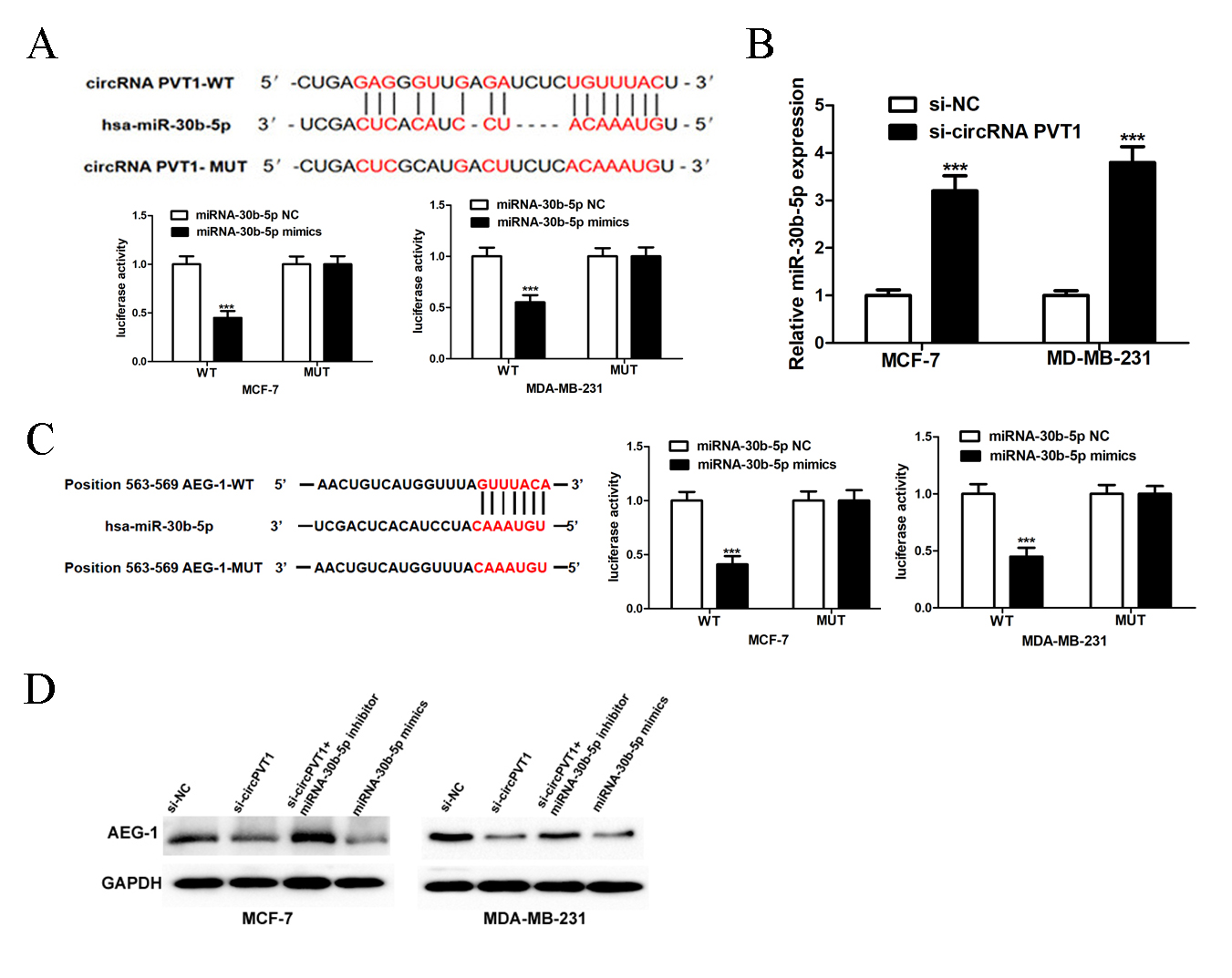 Fig. 3.
Fig. 3.
CircRNA PVT1 regulates miR-30b-5p/AEG-1 pathway. (A) The
binding places between miR-30b-5p and circRNA PVT1. Luciferase assay results
indicated that miR-30b-5p mimics could target circRNA PVT1 3′UTR-WT
(3′-untranslated region-wild type). (B) CircRNA PVT1
knockout by the small interference RNA could up-regulate miR-30b-5p expression.
(C) The binding sites between miR-30b-5p and AEG-1. Luciferase assay results
indicated that miR-30b-5p could target AEG-1 3′UTR-WT. (D) Small si-RNA knockout
circRNA PVT1 in MCF-7 and MDA-MB-231 cells or miR-30b-5p mimics could
downregulate AEG-1 protein expression. MiR-30b-5p inhibitor reversed si-circRNA
PVT1 function. AEG-1, astrocyte elevated gene 1; MUT, mutatin. ***p
By searching the target scan database, we found the presence of miR-30b-5p binding site 3′UTR in AEG-1. Moreover, the biding between AEG-1 and miR-30b-5p was confirmed by luciferase reporter assay and the results revealed that overexpression of miR-30b-5p could inhibit luciferase activity in AEG-1 3′-UTR wild type group. The inhibitory effect disappeared in the mutation group of 3ʹ-UTR binding site of AEG-1 (Fig. 3C). In order to confirm the reliability of our results, we used Western blotting to detect the overexpression of miR-30b-5p in MCF-7 and MDA-MB-231 or the elimination of circPVT1 with si-circPVT1#1, both of which inhibited the expression of AEG-1. However, simultaneous inhibition of miR-30b-5p expression in the circPVT1 knockout group could reverse the inhibition of AEG-1 expression (Fig. 3D).
The expression level of MDA-MB-231 in mice was transfected with si-NC or si-circPVT1#1, and the results demonstrated that si-circPVT1 effectively inhibited the tumor growth (Fig. 4A) and the expression of circPVT1 (Fig. 4B), reduced the tumor volume and weight (Fig. 4C,D), and inhibited the expression levels of AEG-1 and proliferating cell nuclear antigen (PCNA) in nude mice (Fig. 4E).
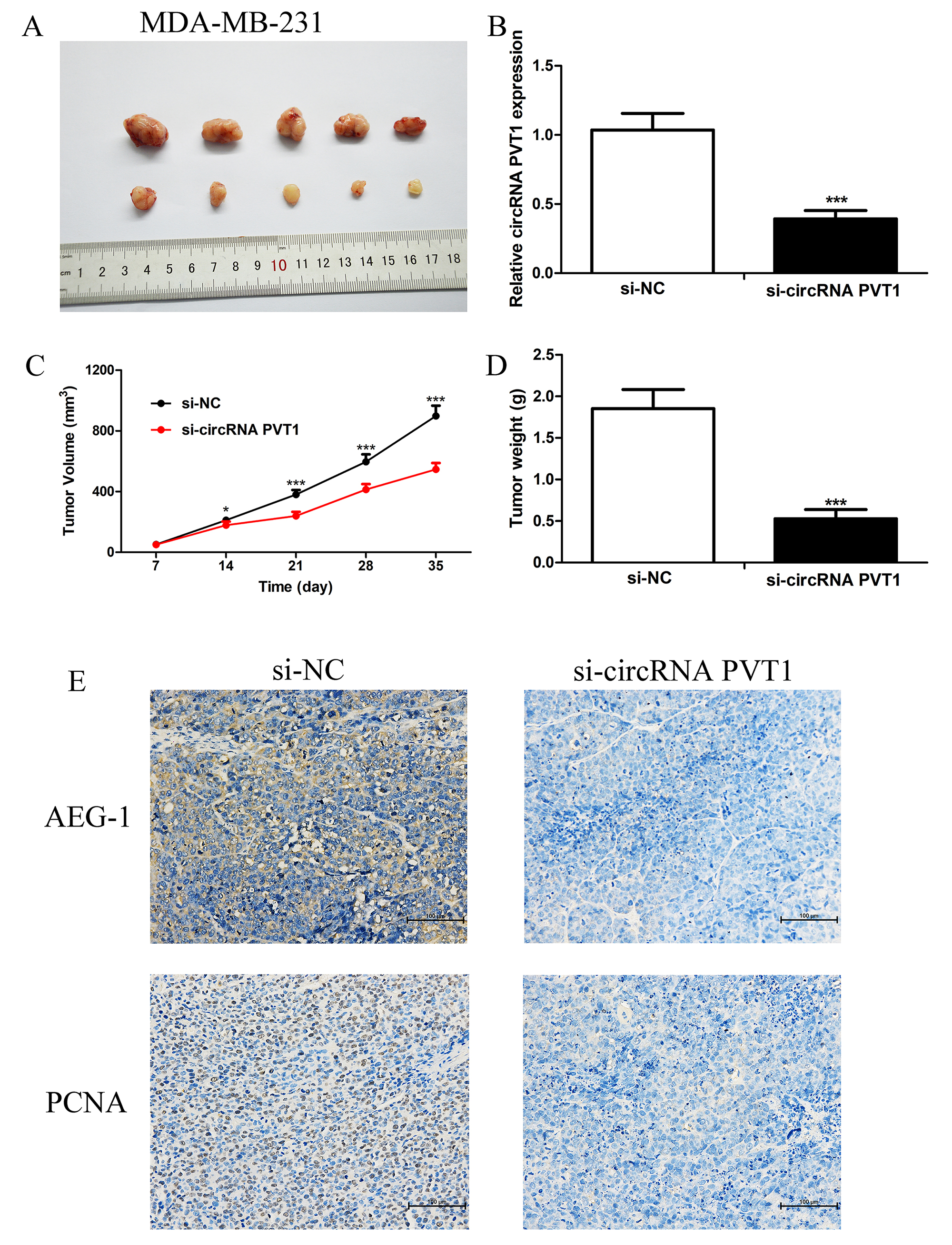 Fig. 4.
Fig. 4.
CircPVT1 knockdown inhibited tumor growth in vivo. (A)
The MDA-MB-231 cells were transfected with si-circPVT1 or si-NC respectively, and
injected into the dorsum of Balb/c mice. Tumor tissues were collected after 5
weeks. (B) q-PCR was used to detect the relative circPVT1 expression in tumor
tissues. (C) Tumor sizes were carefully measured weekly and the formula L
CircPVT1 (circBase ID: hsa_circ_0001821) is located on chromosome 8q24, and the amplification of circPVT1 has been frequently observed in various cancers [8]. CircPVT1 regulates epithelial-mesenchymal transition and tumor stemness in tumor progression, promoting tumor proliferation, invasion and metastasis [9, 10]. Zheng et al. [11] found increased circPVT1 expression in thyroid cancer and it promoted the progression of thyroid cancer. Typically, circPVT1 functions as a miRNA “sponge” and increases special miRNA-targeted mRNA expression, thereby stimulating cancer progression [12]. To elucidate the roles of circPVT1 in breast cancer, we investigated the functions of knockout circPVT1 in breast cancer cells. Our results revealed that high circPVT1 expression was positively correlated with axillary lymph node metastasis and differentiation degree in breast cancer. CircPVT1 can promote breast cancer cell proliferation, migration, and invasion. Further results revealed that circPVT1 inhibits AEG-1 degradation by binding to miRNA-30b-5p. The oncogene AEG-1 demonstrated that it could regulate the expression of genes associated with breast cancer malignant progression [13]. Our data indicates that circPVT1 is a key regulator of AEG-1 in breast cancer and might be a novel target for treatment.
CircRNAs usually localize in the cytoplasm and perform their biological roles via sponging miRNAs. The circRNA-miRNA regulatory networks are involved in the genesis and development of tumors [14]. Our results indicated that circPVT1 was up-regulated in breast carcinoma cells, suggesting that circPVT1 might be involved in the progression of breast cancer through regulation of specific miRNAs. Further dual luciferase reporter experiments indicated that miR-30b-5p was the target of circPVT1. MiR-30b-5p participates in malignant progression in a variety of cancers including pancreatic cancer [15], esophageal carcinoma [16], and papillary thyroid cancer [17]. In our study, miR-30b-5p inhibitors partially reversed circPVT1 depletion-mediated inhibition in breast cancer cells. Therefore, the circPVT1/miR-30b-5p regulatory axis might have important roles in breast cancer.
AEG-1 has approximately a 64 kda molecular weight, also known as MTDH
(metadherin), and the coding gene is located in human chromosome 8q22 [18]. AEG-1
was first reported to be related to lung metastasis of breast cancer in a mouse
tumor model in 2004 [19]. High MTDH expression in breast cancer is closely
associated to high Ki67 expression and poor prognosis, and is related to
invasion, metastasis and EMT of breast cancer [20]. Through activation of
NF-
In our study, the circPVT1 expression is higher in breast cancer tissues compared to corresponding paracancer tissues and higher circPVT1 expression correlates with worse prognosis in breast cancer patients. Silencing circPVT1 could inhibit breast cancer cell malignant biology through sponging miR-30b-5p to promote AEG-1 expression. These findings reveal the key role of the circPVT1/miR-30b-5p/AEG-1 axis in breast cancer malignant biology progression, providing a vital therapeutic target for treatement of breast cancer patients.
circPVT1, circular RNA PVT1; q-PCR, quantitative real-time polymerase chain reaction; si-NC, negative control siRNA; DMSO, dimethyl sulfoxide; EMT, epithelial/mesenchymal transition.
All the data supporting our findings could be found in the “Results” section of the paper. Please contact authors for data request.
YL and WL designed this study. YL and TZ performed the research. YA and TZ collected the patients medical materials and checked the pathological diagnosis. YA analyzed the data. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All subjects gave their informed consent for inclusion before they participated in the study. The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Ethics Committee of Taihe Hospital affiliated to Hubei University of Medicine (approval number: NO.2024KS03).
We would like to express our gratitude to all those who helped us during the writing of this manuscript. Thanks to all the peer reviewers for their opinions and suggestions.
This study was supported by the Shiyan City Scientific Research and Development Project (22Y32).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.