


1 Department of Biomedical Sciences, Noorda College of Osteopathic Medicine, Provo, UT 84045, USA
†These authors contributed equally.
Abstract
Immune cells such as macrophages play a significant role in ocular inflammation by activating or inhibiting several cellular pathways. Systemic infections and autoimmune diseases could activate macrophages by releasing various pro-inflammatory cytokines, chemokines, and growth factors, which reach the eyes through the blood-retina barrier and cause immune and inflammatory responses. In addition, environmental pollutants, allergens, and eye injuries could also activate macrophages and cause an inflammatory response. Further, the inflammatory response generated by the macrophages could recruit additional immune cells and enhance the inflammatory response. The inflammatory response leads to ocular tissue damage and dysfunction and affects vision. Macrophages are generally implicated in the clearance of pathogens and debris, generate reactive oxygen species, and initiate immune response. However, uncontrolled immune and inflammatory responses could damage the ocular tissues, leading to various ocular inflammatory complications such as uveitis, scleritis, diabetic retinopathy, and retinitis. Recent studies describe the role of individual cytokines in the mediation of specific ocular inflammatory diseases. In this article, we discussed the potential impact of macrophages and their mediated inflammatory response on the development of various ocular inflammatory diseases and possible treatment strategies.
Keywords
- macrophages
- eye
- oxidative stress
- inflammation
- infections
- uveitis
Uveitis, retinitis, scleritis, and others, such as age-related macular degeneration (AMD) and diabetic retinopathy (DR), are the most common inflammatory complications of the eye. Ocular inflammatory diseases are the major cause of vision problems in patients worldwide. These are different ranges of disorders, each characterized by inflammation in various parts of the eye. The eyes are constantly exposed to inflammatory processes from external stimuli, such as pollutants, pathogens, allergens, and internal stimuli due to systemic infections and autoimmune diseases. The external stimuli directly affect the eyes locally by inducing oxidative stress, increased production of inflammatory cytokines and chemokines, and disruption and damage to various parts of the eye. Some of these events cause infiltration of inflammatory cells and release of cytokines in the aqueous and vitreous humor, leading to difficulty in vision [1, 2]. These inflammatory cytokines damage the ocular tissues, leading to the infiltration of inflammatory cells and other factors in the aqueous and vitreous humor and causing blindness. Some of these complications have still unknown etiology. Therefore, understanding the pathological mechanisms of ocular inflammatory responses is crucial for developing effective treatments that can restore vision and improve the quality of life of affected individuals.
During infections and autoimmune diseases, immune cells such as T-cells, neutrophils, and macrophages are activated and reach the eyes. In most of the cases, immune cells collaborate with each other and contribute to the immune and inflammatory responses in various tissues, including the eyes. Among these, macrophages play a significant role in ocular inflammatory complications because of their ability to initiate, regulate, and resolve inflammation.
Generally, during pathogen attack or injury, monocytes are differentiated into
active macrophages. The active macrophages then migrate to various tissues as a
defense mechanism, including the eyes [3]. Further, it is well known that
macrophages are significantly adaptable and perform different functions based on
the stimuli they receive from external and internal stress conditions [4]. They
also perform various functions, such as pathogen clearance, tissue repair, and
regulation of inflammation. Macrophages play both protective and pathological
roles in the eyes [5]. In general, systemically, macrophages are very critical in
responding to infections. They help phagocytize and clear infectious pathogens
and debris while coordinating the broader immune response with other immune
cells, such as T-cells [6]. Some studies indicate their involvement in
inflammation is necessary for defense and repair [6, 7, 8]. Macrophages could also
lead to chronic inflammation by releasing various pro-inflammatory cytokines and
chemokines. These inflammatory factors in autocrine and paracrine manner cause
ocular tissue damage and dysfunction, exacerbating ocular diseases. Li et
al. [3] have suggested that, unlike other immune cells, macrophages are found in
the various tissues of the eye (uvea, cornea, retina) and play a significant role
in sustaining ocular cell homeostasis and protecting against infection. Usually,
the resident macrophages located in the ocular tissues play an important role in
immune surveillance in the eye. These resident macrophages identify and respond
to potential threats without causing excessive inflammation and damage to the
ocular tissues. However, when eyes are initially exposed to various oxidant
stimuli, activated macrophages release various innate immune inflammatory markers
such as interleukin (IL)-1
Further, macrophage-mediated oxidative and inflammatory signaling plays a significant role in the pathophysiology of ocular inflammatory complications, and understanding their role is very important in developing novel therapeutic strategies. Recently, various studies have shown a substantial correlation between macrophage-mediated immune response and the pathophysiology of ocular inflammatory complications. Although a few recent articles discussed the critical role of macrophages in the pathophysiology of ocular diseases, not specifically in all ocular inflammatory diseases [3, 6, 7, 8]. Further, these researches emphasized on how macrophages or individual cytokines or chemokines contribute to inflammation in individual eye diseases such as uveitis, diabetic retinopathy, and age-related macular degeneration. Further, few studies have only discussed the significance of macrophage polarization in different eye diseases [3, 6, 7, 8]. This review article discusses recent findings on how macrophages are comprehensively involved across various ocular inflammatory diseases. We conducted PubMed and Google scholar search to find articles published in the last ten years or so, using keywords such as macrophages, immune cells, lymphocytes, inflammation, immune response, oxidative stress, autoimmune diseases, infectious diseases, uveitis, scleritis, retinitis, age-related macular degeneration (AMD) and diabetic retinopathy (DR). In this narrative review article, we have included research articles, narrative reviews, systematic reviews, and clinical and pre-clinical researches to discuss the significance of macrophages in ocular inflammatory complications. Understanding the significance of macrophages is very important for the potential development of novel therapies to regulate immune and inflammatory responses leading to ocular inflammatory complications.
During infections and autoimmune conditions, macrophages change their
polarization phenotype (pro-inflammatory M1 and anti-inflammatory M2) in response
to inflammatory stimulus. M1 macrophages are classic macrophages that are mainly
stimulated by pathogen stimuli and cytokines such as interferon-gamma
(IFN-
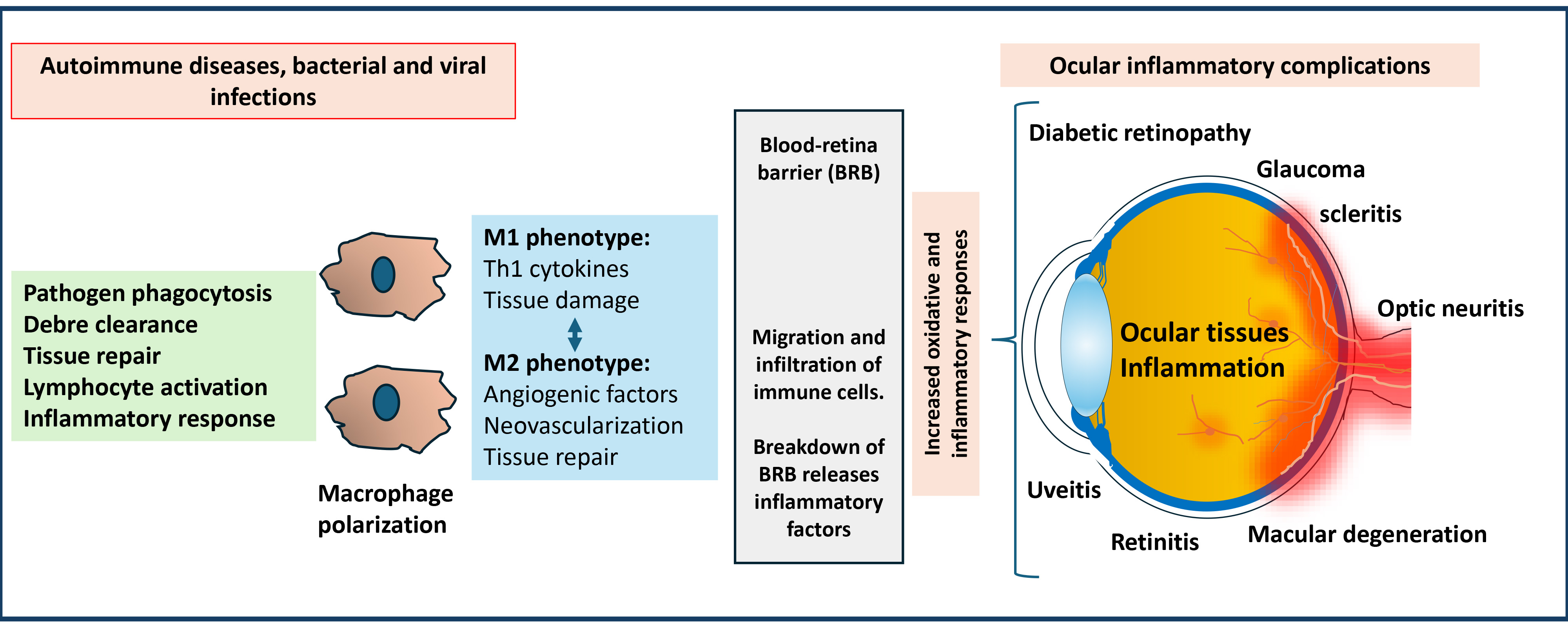
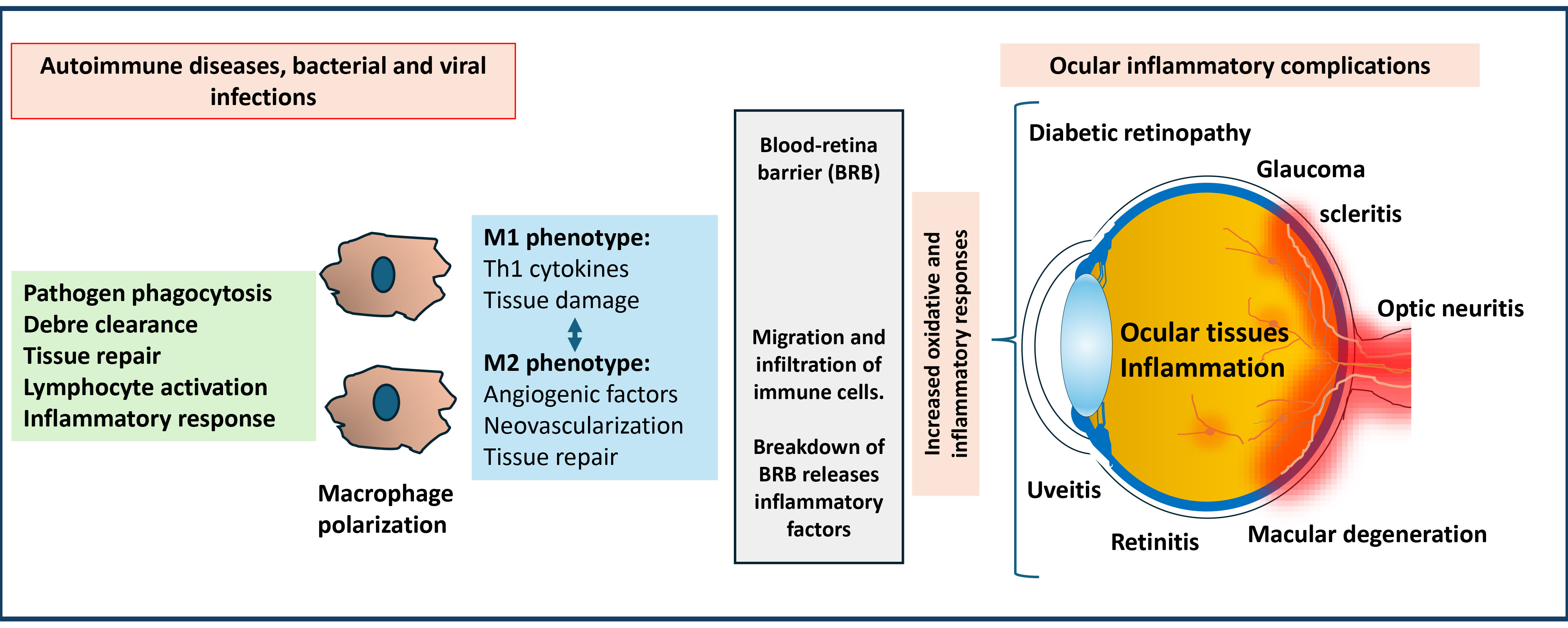 Fig. 1.
Fig. 1.
Significance of macrophage-mediated inflammatory response in developing ocular inflammatory complications. Macrophages polarize into M1 and M2 phenotypes in response to stimuli during autoimmune diseases and infections. M1 macrophages generate pro-inflammatory type-1 T helper (Th1) cytokines. M2 macrophages generate angiogenic factors and anti-inflammatory markers. These factors can damage ocular tissues by crossing the blood-retinal barrier. The polarization disbalance could lead to various ocular inflammatory diseases and cause potential vision loss.
Uveitis, one of the most common and severe inflammatory diseases of the eye, is characterized by uveal tract inflammation, including the iris, ciliary body, and choroid. This inflammation can extend to adjacent tissues such as the sclera, retina, and vitreous humor [12]. Autoimmune uveitis (AIU) involves an inflammatory response in these uveal tissues due to an autoimmune reaction against self-antigens or an innate inflammatory response triggered by external stimuli [13, 14]. The blood-retinal barrier must be disrupted for AIU to develop, activating ocular antigen-specific cluster of differentiation (CD) 4+ T cells. These T cells infiltrate the eye and recruit macrophages, which become classically activated, generate reactive oxygen and nitrogen species, and cause damage to uveal tissues [15, 16]. Ocular antigens such as arrestin (S-antigen), interphotoreceptor retinoid-binding protein (IRBP), and recoverin have been identified in cases of autoimmune uveitis [17, 18]. AIU can occur as an isolated condition or be associated with systemic autoimmune or autoinflammatory diseases, including rheumatoid arthritis, sarcoidosis, Behçet’s disease, multiple sclerosis, and even aging [19, 20, 21, 22, 23]. In autoimmune uveitis, macrophages are among the primary immune cells infiltrating the eye, releasing pro-inflammatory cytokines and acting as mediators that exacerbate ocular inflammation. However, uncontrolled management of inflammatory response could lead to ocular tissue damage and dysfunction, causing vision loss and other ocular complications such as glaucoma, cataracts, and retinal detachment.
The activation of macrophages is a critical determinant of disease outcome,
regulated by inflammatory signals from the microenvironment, including
interferon-gamma (IFN-
Experimental models of AIU have provided insights into how macrophages contribute to this condition. The disease develops when activated CD4+ Th1 and Th17 macrophages infiltrate the eye, leading to the recruitment of neutrophils and macrophages and subsequent structural damage [29]. Depleting macrophage-specific proteins such as CD47 significantly reduces uveitis severity, indicating the significance of macrophages in the mediation of uveitis [30, 31, 32]. Further, Zhao et al. [26] have indicated that the macrophage levels are different across various EIU phases when compared to other immune cells in the retina tissue. This study also demonstrated that during the acute phase, the macrophage levels peaked and decreased during the chronic phase. Further, the change in the shift suggests a polarization of macrophages from pro-inflammatory M1 to anti-inflammatory M2 over time [26]. Glucocorticoids mediate the P38-MAPK/myocyte enhancer factor-2c axis, promoting the transition from M2 to M1 macrophages and releasing anti-inflammatory factors, inhibiting EAU and supporting tissue healing [33]. Several studies suggest that macrophage-mediated activation of Th1 and Th17 responses plays a major role in the pathology of uveitis (Fig. 2) [29, 30, 31, 32].
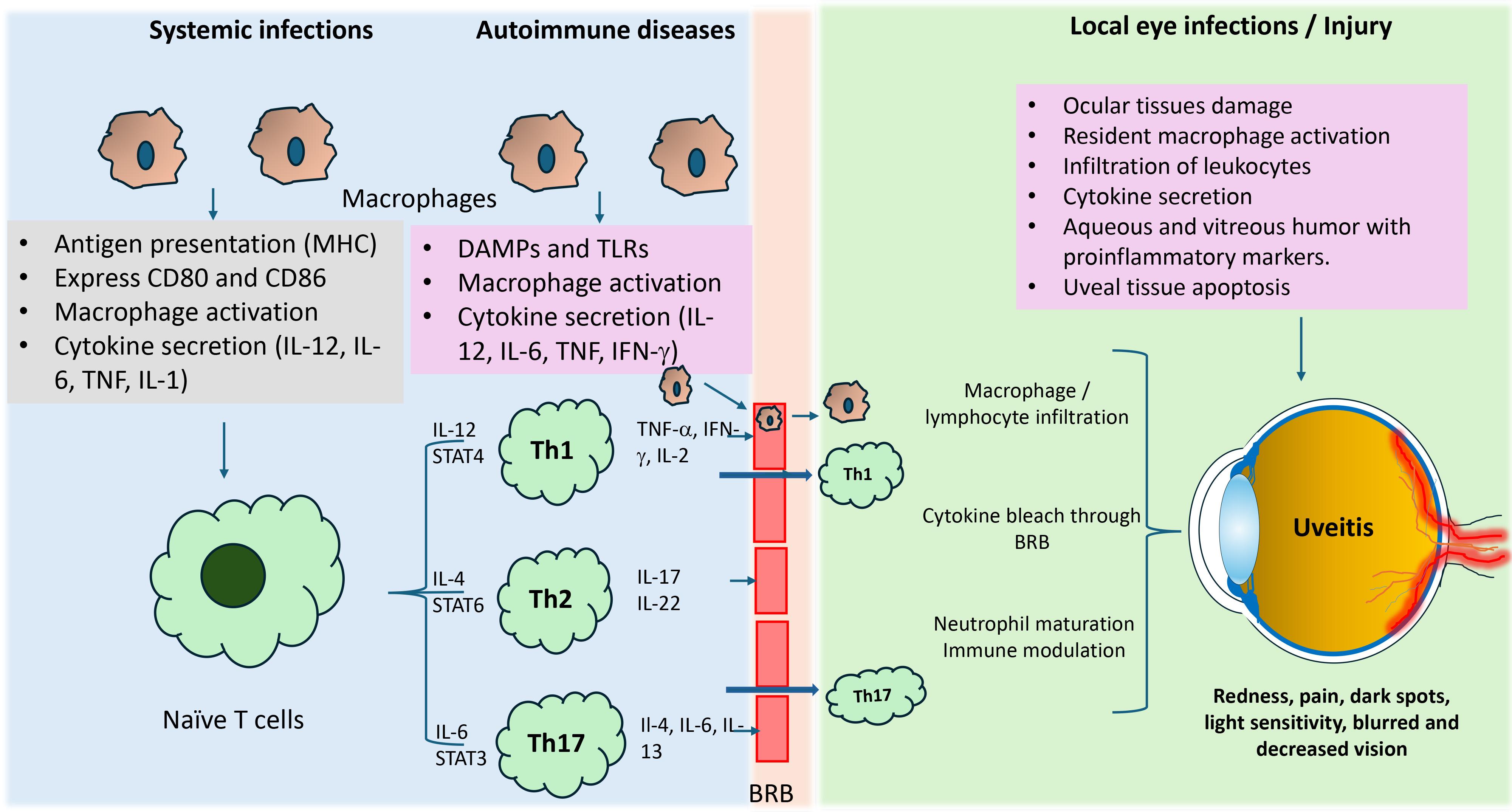
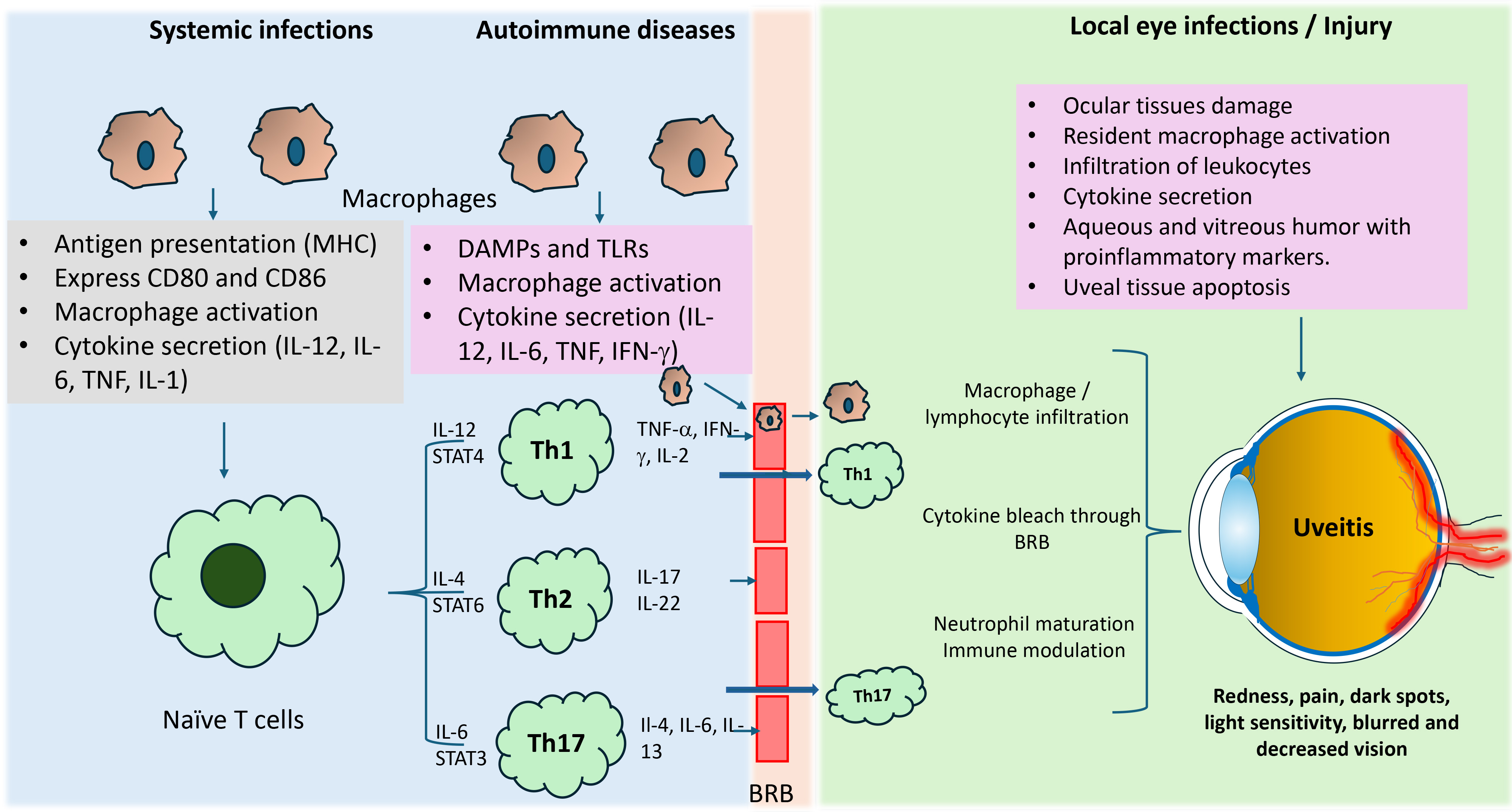 Fig. 2.
Fig. 2.
Macrophage-mediated inflammatory response in uveitis. During
autoimmune diseases and infections, macrophages act as antigen-presenting cells
and, cause their activation and release pro-inflammatory cytokines. These
cytokines activate naive T cells, which differentiate into subsets, such as Th1,
Th2, and Th17. These subsets are formed based on the type of stimulated
cytokines. These cell subsets release various inflammatory cytokines and further
amplify immune response locally and by crossing the blood-retinal barrier
internally at ocular tissues. Th1 and Th17 cell-mediated inflammatory response
are well known to play a significant role in developing uveitis complications.
IL-, interleukin-; TNF, tumor necrosis factor; IFN-
Enhanced M2 macrophage polarization has been shown to be promoted by IL-33,
which signals through the interleukin-1 receptor-like 1 protein receptor (ST2)
receptor [34]. Further, Barbour et al. [35] have shown that after three
weeks of EAU induction, ST2-deficient mice exhibited worse uveitis symptoms than
wild-type mice. They have also shown that interleukin (IL)-33 treatment in
wild-type mice improved uveitis lesions by increasing CD206 and CD273 cells,
indicating that IL-33/ST2 signaling enhances M2 polarization and alleviates EAU
symptoms [35]. Similarly, Huang et al. [36] have shown that activating
the aryl hydrocarbon receptor (AhR) with 2,3,7,8-tetrachlorodibenzo-p-dioxin
through the nuclear factor-kappa binding protein (NF-
Interestingly, suppressors of cytokine signaling (SOCS) proteins, particularly
SOCS1 and SOCS3, play crucial roles in regulating macrophage polarization and
cytokine expression [41]. For example, in bone marrow-derived macrophages, SOCS3
negatively regulates granulocyte-macrophage colony-stimulating factor
(GM-CSF)-induced expression of CCL2, arginase-1 (Arg-1), and matrix
metallopeptidase (MMP) 12 [42]. SOCS3-deficient mice (LysMCre/+SOCS3fl/fl)
exhibit increased GM-CSF in the retina and trigger the release of CCL2 and Arg-1
from macrophages. This leads to enhanced retinal degeneration and angiogenesis
due to inflammation [43]. Further, it has been shown that during the initial
phase of EAU, there is an increased infiltration of neutrophils, decreased
macrophages, and increased inflammatory cytokines such as tumor necrosis
factor-alpha (TNF-
In the context of Experimental Autoimmune Uveitis (EAU), the potential for
therapeutics in treating uveitis has been demonstrated. For example, Chen
et al. [45] have shown that the small molecular integrin inhibitor
(GW559090) prevents the accumulation of Ly6C+ macrophages and blocks the
development of EAU in mice by inhibiting Th17 cell migration through the
blood-retinal barrier. This leads to swift suppression of ocular inflammation and
preservation of the blood-ocular barrier. Further, several studies have shown
that antioxidants and small molecular anti-inflammatory agents could prevent
animal models of EAU and EIU by preventing the activation of NF-
Optic neuritis (ON) can encompass a wide range of conditions that may cause optic neuropathies. However, it is defined as an inflammatory condition that demyelinates the optic nerve and causes the loss of retinal ganglion cells (RGCs), resulting in damaged vision. ON is often studied using the experimental autoimmune encephalomyelitis (EAE) model [54]. ON is an associated effect of Multiple Sclerosis (MS), so many models aim at replicating this autoimmune condition. Funaki et al. [55] have found that upregulated gal-3 controls the NOD-like receptor family pyrin domain containing 3 (NLRP3) signaling in microglia/macrophages in the visual pathways during the peak of an MS flare in EAE mice. Like the retina, the brain is also an immune-privileged site. However, most immune cells in the inner environment of the central nervous system in an inflammatory state are microglial cells and macrophages responsible for most neuronal degradation [54, 56].
In EAE, macrophages infiltrate the optic nerve and contribute to demyelination and axonal damage by releasing pro-inflammatory cytokines and reactive oxygen species [56]. The infiltrated macrophages could promote inflammation and help clear the myelin debris in optic nerves. Further, this intriguing dual role of macrophages regulates the immune cell dynamics between EAE and uveitis. Thus, suppression of the M1 phenotype and activation of M2 phenotype macrophages could prevent retinal inflammation and protect against optic nerve damage and RGC death.
The polarization is still a point of therapeutic interest as fatty acids (FAs), when obtained through diet, can positively impact neuronal health by modulating the macrophage phenotypes from M1 to M2 [57, 58, 59]. Further, cytokines such as IL-12 and IL-23 can significantly contribute to the progression of EAE by promoting the recruitment of M1-macrophages and the release of CXCL-10 and CXCL-11 [60, 61, 62].
Retinitis is inflammation of the retina due to infections by various pathogens such as bacteria, fungi, and viruses. Herpes simplex virus (HSV), herpes zoster virus (HZV), and Cytomegalovirus (CMV) are common viral infections that can lead to retinitis [63, 64]. Further, syphilis and tuberculosis bacterial infections could also lead to retinal inflammation [63]. These and additional infectious agents are known to cause visual impairment by disrupting the retina, retinal detachment, macular edema, and retinal scarring [63]. Most of these cases are also linked with uveitis complications and autoimmune diseases [65]. Macrophage infiltration is one of the major causes of retinal inflammation. Although immune cells generated cytokines play a significant role in retinal damage, the blood-retina barrier provides great support to maintain homeostasis. Yang et al. [66] have indicated that the damage to retinal pigmented epithelial cells could cause cell death, which is sometimes aggravated by the macrophages and T-cell mediated inflammatory response. On the other hand, Taylor et al. [67] have suggested that retinal pigment epithelial cells also regulate immune cells in the eye, as they can cause macrophage apoptosis and altered immune modulation. Most of the studies have shown that infectious uveitis due to viral infections causes retinal apoptosis, necrosis, and inflammation [66, 67, 68]. Viral infections in the retina, such as from CMV, HSV, or varicella zoster virus (VZV), can lead to retinal detachment by causing necrosis of retinal cells. As the virus replicates, it destroys retinal tissue, which is then replaced by scar tissue. The viral infections from CMV, HSV, and VZV could cause damage to the retina and cause retinal necrosis, leading to retinal detachment. Further, the damaged retina tissue can also inflame healthy cells, leading to their detachment from the retina. Indeed, some studies have shown that during acute retinal necrosis and CMV-induced retinitis, the inflammatory response plays a major role in retinal detachment in retinitis [69, 70, 71]. Some viral particles, such as CMV, replicate in the eye due to the lack of functional activity of CD4+ T cells, which could lead to retinal necrosis [69].
Similarly, HSV and Epstein-Barr virus (EBV) could also result in retinal necrosis and inflammation, eventually leading to retinal detachment and visual complications [71, 72]. The role of macrophages here is to respond to the infection and play a dual role by helping to clear debris and remove the virus. At the same time, the uncontrolled release of pro-inflammatory cytokines and chemokines could lead to ocular inflammation. The excessive inflammatory response leads to tissue dysfunction, scarring, and an increased risk of retinal detachment. Indeed, a recent study by Sterling et al. [73] demonstrated that retinal perivascular macrophages, located on post-capillary venules of the eye, play a crucial role in facilitating immune cell migration across the blood-retinal barrier by aiding Ly6C+ monocyte infiltration in a mouse model of retinal inflammation. This study highlights the significance of perivascular macrophages in ocular inflammation.
Furthermore, in retinitis pigmentosa (RP), a broad collection of genetic eye conditions resulting in the deterioration of photoreceptor cells needed to see the light in the retina, microglial cells could also contribute a significant role in the degeneration [74]. This degeneration ultimately leads to a decline in visual acuity. The damage results from the loss of rods and usually starts with night blindness and gradually narrows the visual field, often resulting in tunnel vision. The genetic defects may injure multiple pathways, including apoptosis, ciliary transport, and endoplasmic reticulum stress. Like with uveitis and ON, in RP, there is a disruption in the blood-retinal barrier that results in leakage of macrophages, which play a critical role in the progression of the disease [75]. The microenvironment in RP involves the infiltration of macrophages in the retina that release pro-inflammatory factors, leading to the characteristic degeneration of the retinal photoreceptor layer [76, 77]. Further, during degeneration, the macrophages and resident microglial cells could kill adjacent cells, phagocytize debris, and facilitate regeneration [78]. Thus, infiltrated macrophages in the degenerated retina help in neuroprotection and neurodegeneration.
In addition, depending on macrophage polarization state (M1 or M2), they can exacerbate retinal damage by promoting inflammation and cell death. On the other hand, M2 macrophages are involved in retinal tissue damage repair, and the removal of debris from dying photoreceptors could also contribute to retinal fibrosis. A study by Neves et al. [79] has shown that immune modulation between the M1 pro-inflammatory and the M2 anti-inflammatory can be achieved by using Platelet-Derived Growth Factor (PDGF)-like signaling-induced Mesencephalic Astrocyte-derived Neurotrophic Factor (MANF) for successful regeneration of the retina in flies and mice. Other studies have also found that a diet rich in FA showed benefits in RP related to shifting the macrophage polarization to M2 in mice, while the absence of FAs correlated with increased degeneration in RP [80, 81]. In addition, some studies have also shown the significance of innate immune responses in retinal detachment [82, 83]. Cao et al. [82] have demonstrated that an inhibitor of caspase-1, VX-765, inhibits microglial pyroptosis by changing the M1 phenotype to M2. They have also indicated that caspase-1 inhibition could reduce microglial pyroptosis, shift microglial phenotypes to a protective state, and preserve photoreceptor structure. Thus, the inhibition of caspase-1 could be a potential therapeutic approach for retinal detachment diseases. Similarly, Cao et al. [83] have also shown that P2X7-mediated microglial activation and pyroptosis are critical for photoreceptor degeneration and retinal detachment.
Scleritis is an inflammatory condition of the eye, often linked to systemic
infectious or non-infectious diseases. The most common underlying cause of
scleritis is an autoimmune disease such as rheumatoid arthritis (RA), lupus, and
Wegener’s granulomatosis [84]. Scleritis can be classified into various subtypes,
such as episcleritis, anterior scleritis, and posterior scleritis. Among these,
the posterior scleritis is rare but more severe, potentially leading to
complications such as choroiditis, retinal detachment, and optic nerve damage. On
the other hand, episcleritis affects only the superficial capillaries and is less
severe. Infectious agents are responsible for
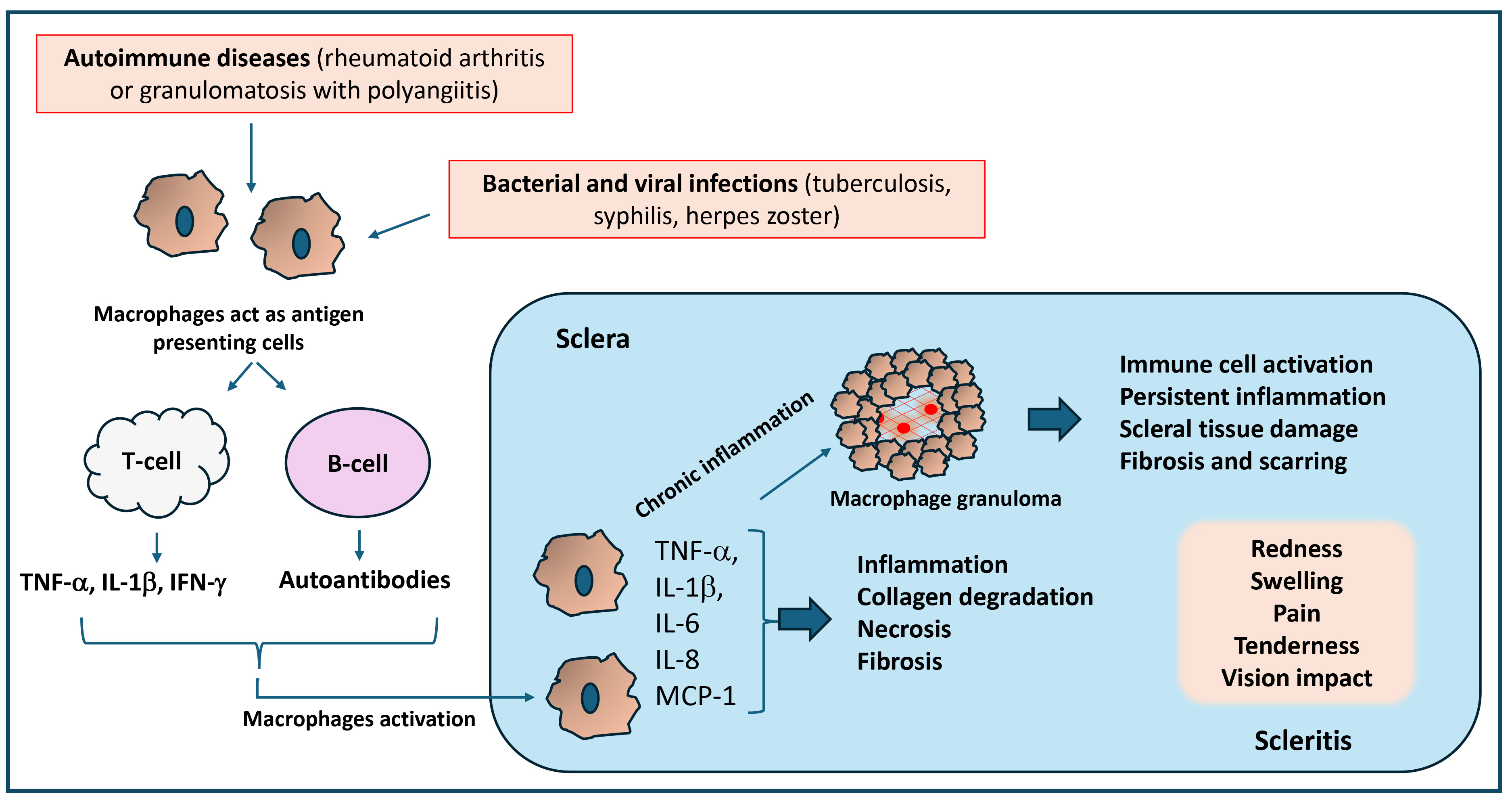
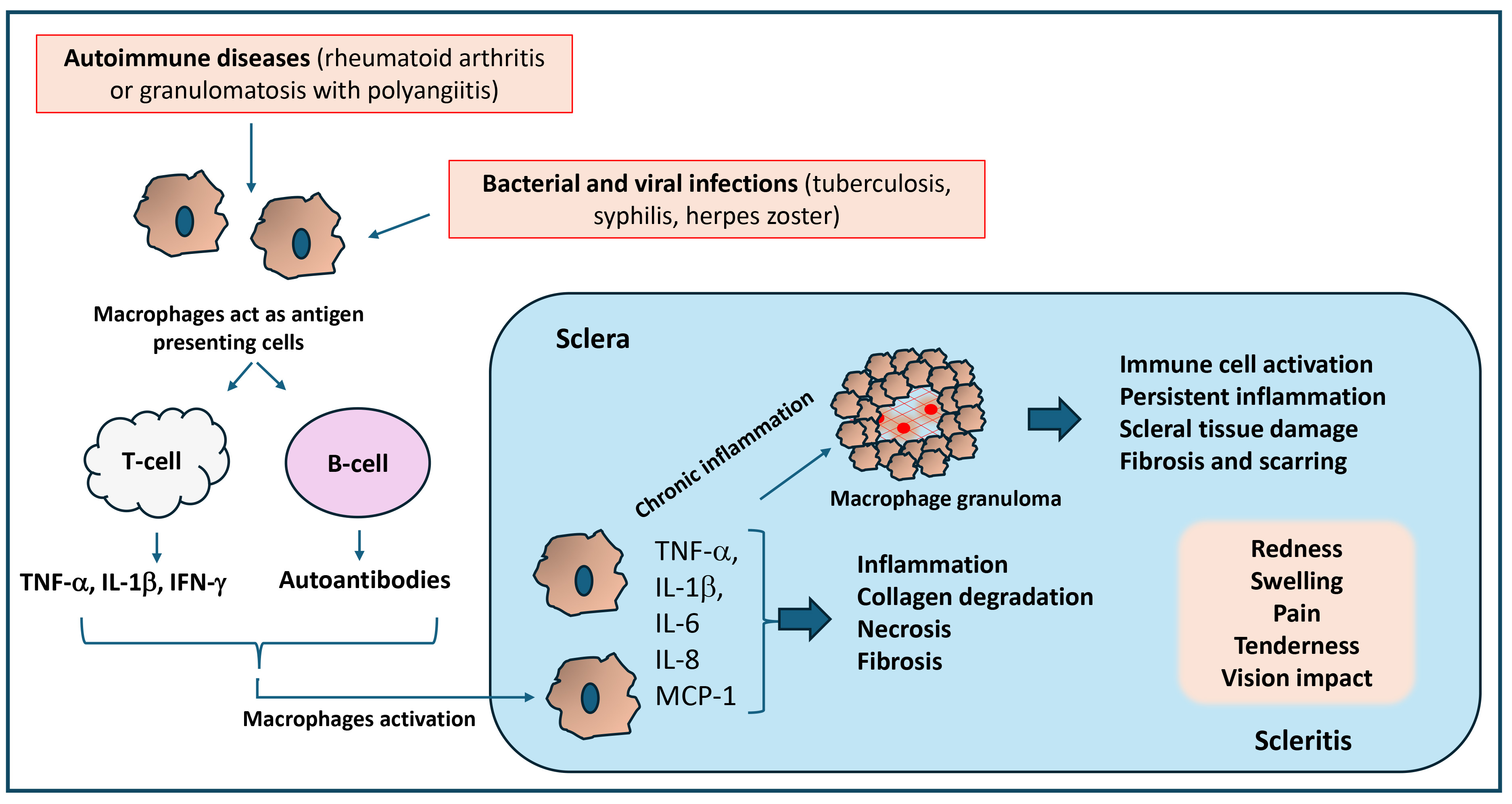 Fig. 3.
Fig. 3.
Significance of inflammatory response in the pathology of scleritis. Macrophage activation during autoimmune and infectious diseases triggers an inflammatory response that can activate T-cells and B-cells, which produce inflammatory cytokines and autoantibodies that damage the sclera and cause necrosis and scleral collagen degradation. Uncontrolled and chronic inflammation could also lead to the formation of granulomas, which promote persistent inflammation, scleral tissue damage, and fibrosis. Thus, the inflammatory response caused by activated macrophages and granulomas contributes to chronic inflammation, leading to pain, swelling, and tissue damage in scleritis. MCP-1, monocyte chemoattractant protein-1.
Further, Fong et al. [90] have found that significantly increased number of macrophages in the conjunctival epithelium of conjunctival and scleral biopsies of scleritis patients. Scleral specimens also showed an increase over controls of macrophages. This study also demonstrated that primary vasculitis plays an important role in the pathogenesis of scleritis. Scleral biopsies taken from scleritis patients showed vascular occlusions and infiltration, as well as evidence of macrophages and T cells. Further, a recent study also suggests that intraocular inflammation associated with scleritis could lead to significant visual impairment [91]. Thus, the role of macrophages in scleritis is still not completely known because of the rarity of the disease and the lack of established animal models.
Although genetic and non-genetic factors contribute to retinal diseases, innate
immune responses and inflammatory responses mediated by the macrophages could
play a critical role in the progression of these complications [92]. The diseases
where the inflammatory response is also a contributing factor include diabetic
retinopathy (DR), glaucoma, and age-related macular degeneration (AMD). Recent
studies have shown that increased NLRP3-mediated release of IL-1
Diabetes mellitus is an autoimmune disease characterized by hyperglycemia resulting from improper insulin production and insulin resistance. Hyperglycemia-mediated oxidative stress and inflammation have been shown to be a major risk for developing diabetic retinopathy (DR), a secondary diabetic complication [94]. Diabetic retinopathy features microvascular retinal lesions and is the leading cause of visual impairment in the middle-aged diabetic population around the world [94]. Diabetic retinopathy is generally divided into non-proliferative and proliferative. Non-proliferative DR is seen at the initial stage, where blood vessels in the retina are weak, leading to mild visual problems. In contrast, proliferative DR is more advanced, where neovascularization is seen on the retina. Early events of diabetic retinopathy include retinal microvascular injury leading to changes in the retinal neurovascular unit and its components. Anti-angiogenic treatments have shown some therapeutic benefits in controlling DR. Ciulla et al. [95] have shown that diabetic macular edema treated using anti-vascular endothelial growth factor (VEGF) therapy could decrease edema and improve visual acuity. However, many patients do not respond to anti-VEGF treatment, indicating that other factors are involved in the pathogenesis of diabetic macular edema [96]. A study by Zhang et al. [97] found that monocyte-derived macrophages also promote diabetic retinopathy progression in mice.
Further, Wang et al. [98] have shown increased levels and density of
macrophage-like cells in the DR subjects with macular edema. The
macrophage-mediated inflammatory response and angiogenesis could lead to diabetic
macular edema. Thus, in DR, macrophages are involved in inflammation, oxidative
stress, pathological angiogenesis, and tissue healing processes. Further, the
breakdown of the blood-retinal barrier during the early stages of DR allows blood
immune cells to enter the retina, causing an inflammatory response mediated by
macrophages. The breakdown of the blood-retinal barrier is influenced by
inflammatory factors and causes vascular damage and neovascularization. Although
macrophages seem to be the primary cells involved in the pathogenesis of
proliferative diabetic retinopathy, they might also be responsible for capillary
occlusion, acellular capillaries, retinal nonperfusion, and retinal ischemia seen
in diabetes conditions. New evidence has emerged to highlight the pivotal role of
macrophage polarization in the pathophysiology of diabetic retinopathy [99].
During hyperglycemia, the balance between M1 and M2 polarization is disturbed,
leading to increased M1/M2 polarization. This polarization imbalance is linked to
insulin resistance and poor glycemic control. However, over time, macrophages in
diabetic conditions shift from protective M2a to pro-inflammatory M1 phenotypes,
contributing to increased inflammation in both human and mouse models of diabetes
[99]. Further, in diabetic retinopathy (DR), both M1 and M2 polarization are
present at the preclinical stage, but M1 polarization dominates in later stages,
where VEGF secretion drives retinal neovascularization. Whereas M2 macrophages
contribute to excessive VEGF release, raising concerns about treatments that
shift M1 to M2 polarization [99, 100]. Thus, additional researches are required to
understand macrophage/microglia polarization in the therapeutic development of
DR. Additionally, macrophages are known to influence cellular proliferation
through the synthesis of various cytokines and growth factors such as
TNF-
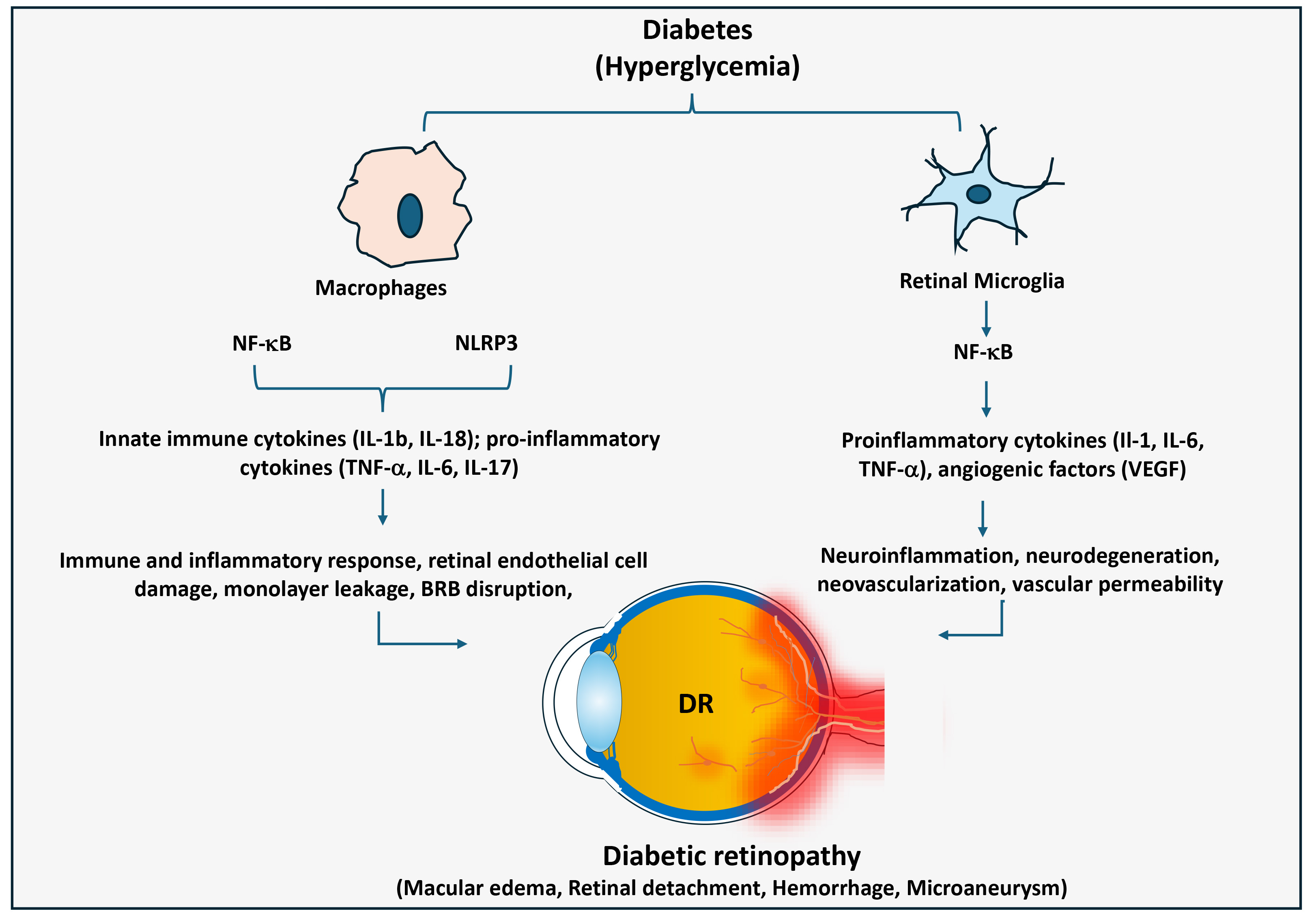
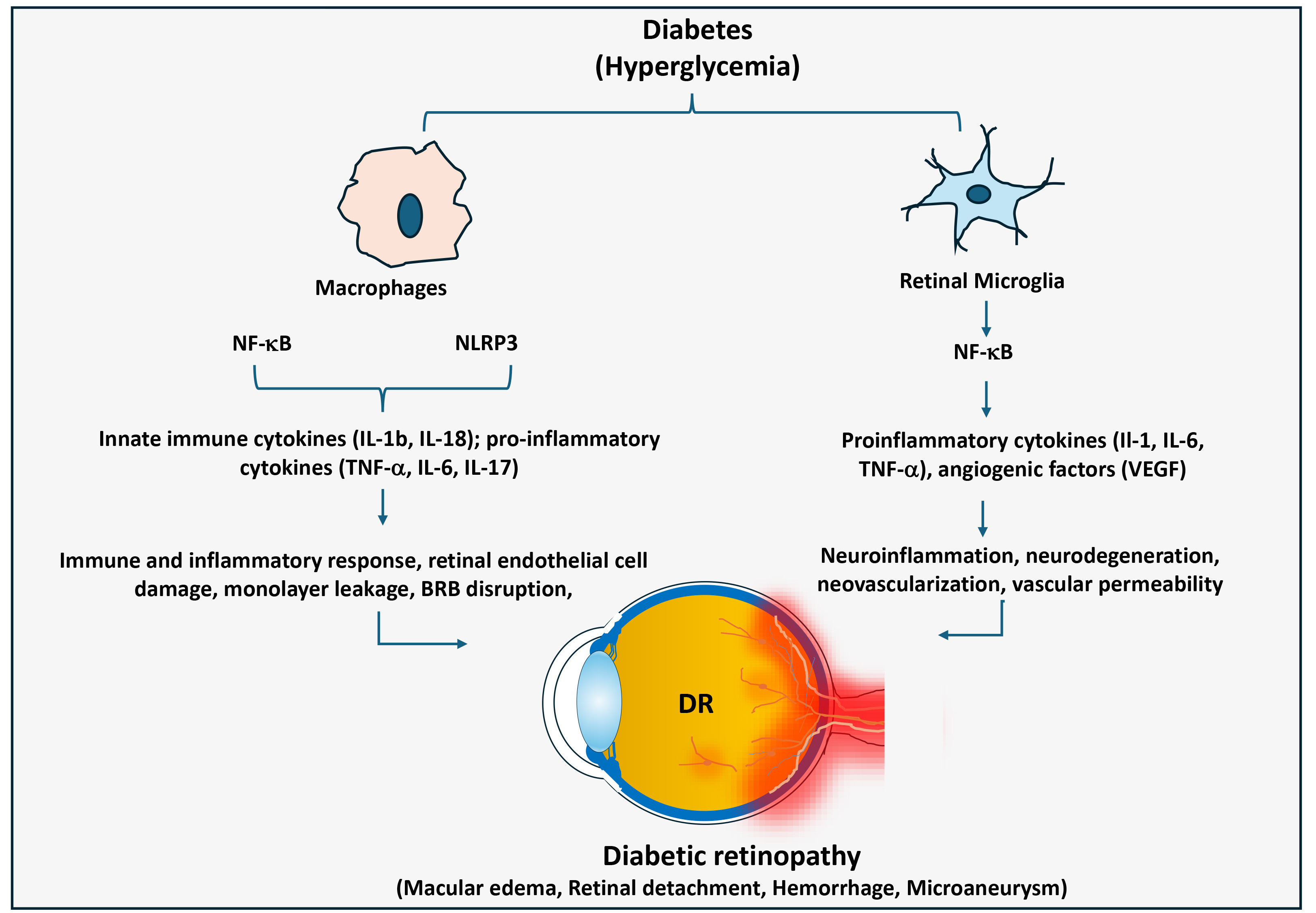 Fig. 4.
Fig. 4.
Significance of macrophage-mediated inflammatory responses in
diabetic retinopathy. In diabetes, macrophages and retinal microglia cells
promote chronic inflammation in the retina through activating NF-
Glaucoma is a progressive neurodegenerative disease characterized by damage and
death of retinal ganglion cells and tapering of the retinal nerve fiber layer
[105]. Glaucoma is one of the most frequent causes of irreversible blindness that
often results from irreversible loss of retinal ganglion cells. The loss of
retinal ganglion cells is caused by a rapid increase in intraocular pressure,
which can lead to acute angle-closure glaucoma [106]. Damage and dysfunction of
the retinal ganglion cells could be categorized as primary or secondary. Primary
damage often results from direct injury to the axon or cell body, whereas
secondary damage results from releasing toxic effectors from adjacent dying
cells. The mechanisms leading to cell death in retinal ganglion cells of glaucoma
include the activation of microglia and macrophages [106, 107]. Macrophages have
been shown to play a protective role in retinal ganglion cells after optic nerve
injury [108]. Although the immune response has been shown to be critical for
normal-tension glaucoma, recent studies suggest that neuroinflammation could be
the significant factor contributing to various glaucomas [105, 109]. However, very
few studies are available showing the significance of macrophage-mediated
inflammatory response leading to glaucoma [110, 111]. Bell et al. [111]
have analyzed the trabecular meshwork of patients with open-angle and acute-angle
closure glaucoma and have found macrophages in the tissue. Few studies also
indicate that macrophages are drawn to the trabecular meshwork of human eyes
following selective laser trabeculoplasty of glaucomatous eyes [112, 113, 114]. At the
same time, microglia in the optic nerve are activated and generate
pro-inflammatory cytokines [112]. In a DBA/2J mouse model of glaucoma, myeloid
cells such as monocytes have been shown to be accumulated at the early stages
before ON damage is detectable [113]. Furthermore, increased serum levels of the
MCP-1, ET-1 but not MMP9, and hs-CRP have been linked to visual field
deterioration in normal-tension glaucoma patients, suggesting peripheral
macrophages play a role in glaucoma development [114]. Further, Lee et
al. [114] have also shown that CD163+ macrophage infiltration is crucial
for the development of glaucoma. In addition, Bell et al. [111] have
observed that a significant increase in the number of macrophages in the retina
was found to follow retinal ganglion cell loss in a rat model of glaucoma. This
study also suggests that the removal of macrophages results in a reduction of
retinal ganglion cell loss, suggesting that macrophages play a role in retinal
ganglion cell death. Further, Bauer et al. [115] have analyzed various
pro-inflammatory markers in aqueous humor of Fuchs uveitis syndrome (FUS)
patients with secondary glaucoma. They found increased levels of TGF-
Age-related macular degeneration (AMD) is a retinal disease that primarily affects the central part of the macula, which causes irreversible decline in central vision [116]. Pathologically, AMD presents with an accumulation of drusen, retinal pigment epithelium degeneration, photoreceptor atrophy, and sometimes choroidal neovascularization. Besides oxidative stress, genetic variations, and angiogenesis, ocular inflammatory response also plays a critical role in AMD pathogenesis. Here, macrophages are the primary immune cells that contribute to increased immune and inflammatory responses in AMD by releasing various pro-inflammatory factors [117]. Macrophage-mediated inflammatory response can promote neovascularization, and their interactions are essential for the pathogenesis of AMD. Blood vessels and mononuclear phagocytes are not present in the subretinal space during normal conditions in adulthood, but many macrophages are found in the subretinal spaces of AMD lesions [116]. Overall, the pathology of AMD lesions has been proven to show the infiltration of macrophages and the accumulation of inflammatory components.
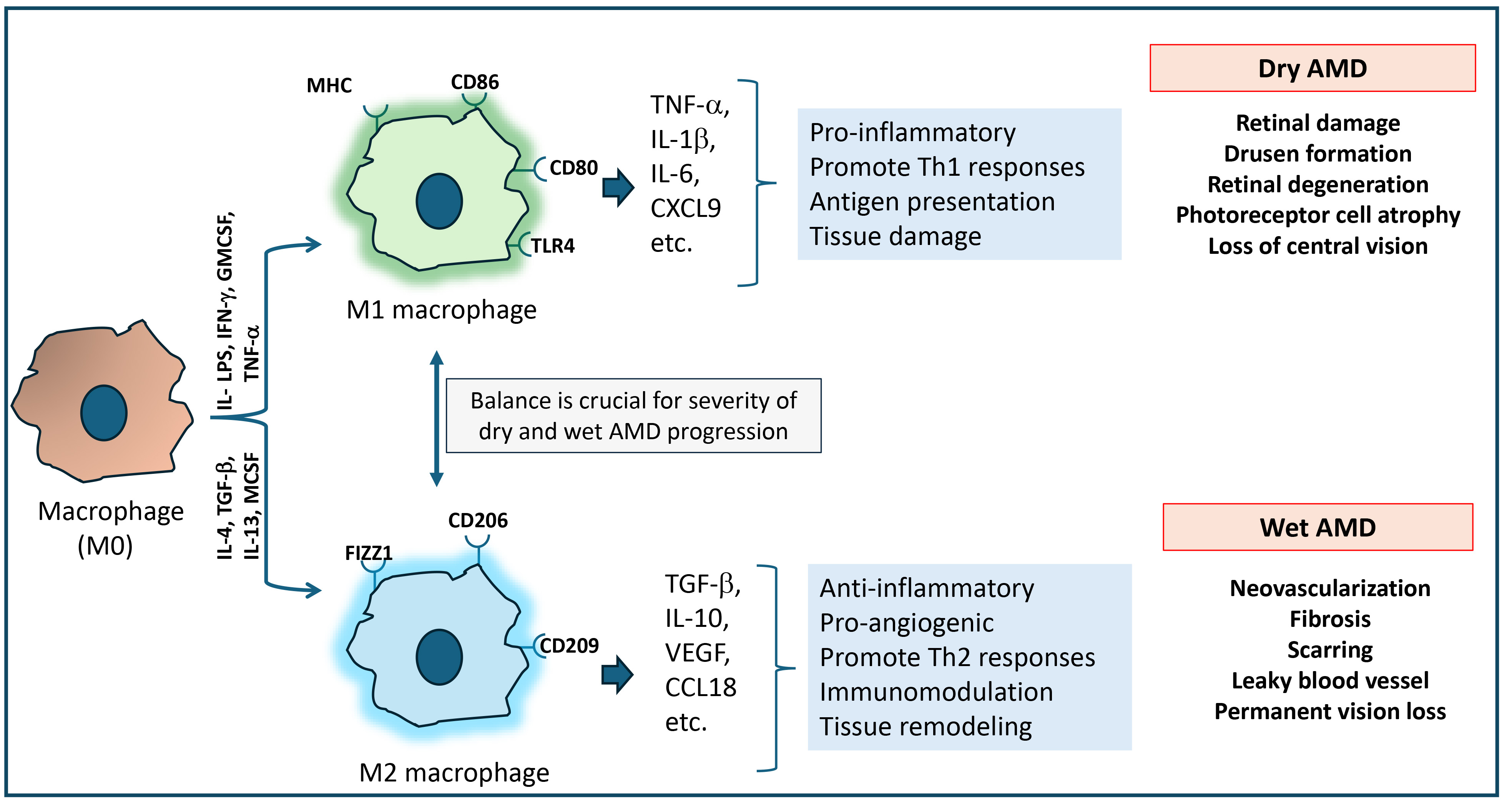
Resident tissue macrophages are found adjacent to retinal pigment epithelium, which sits on Bruch’s membrane, an acellular layer of connective tissue and basal lamina [118]. An inflammatory response occurs when changes occur in Bruch’s membrane and the retinal pigment epithelium. Macrophages have been histologically found near AMD lesions, especially in the breakdown of Bruch’s membrane and retinal pigment epithelium atrophy [118]. As this breakdown continues, retinal pigment epithelium can be lost, and the neural retina can become atrophied, resulting in dry AMD. This can then progress into wet AMD when choroidal neovascularization takes place. Generally, dry AMD is more common, with a gradual breakdown of the light-sensitive cells in the macula, while wet AMD is more severe, involving choroidal neovascularization leading to rapid vision loss [119]. Macrophages help in the clearing of yellow deposits under the retina (drusen). However, uncontrolled activation could lead to a weak inflammatory response, causing retinal damage over time. In wet AMD, macrophage-mediated release of pro-angiogenic factors such as growth factors, specifically VEGF, could increase the formation of new blood vessels, leading to retinal damage and dysfunction (Fig. 5).
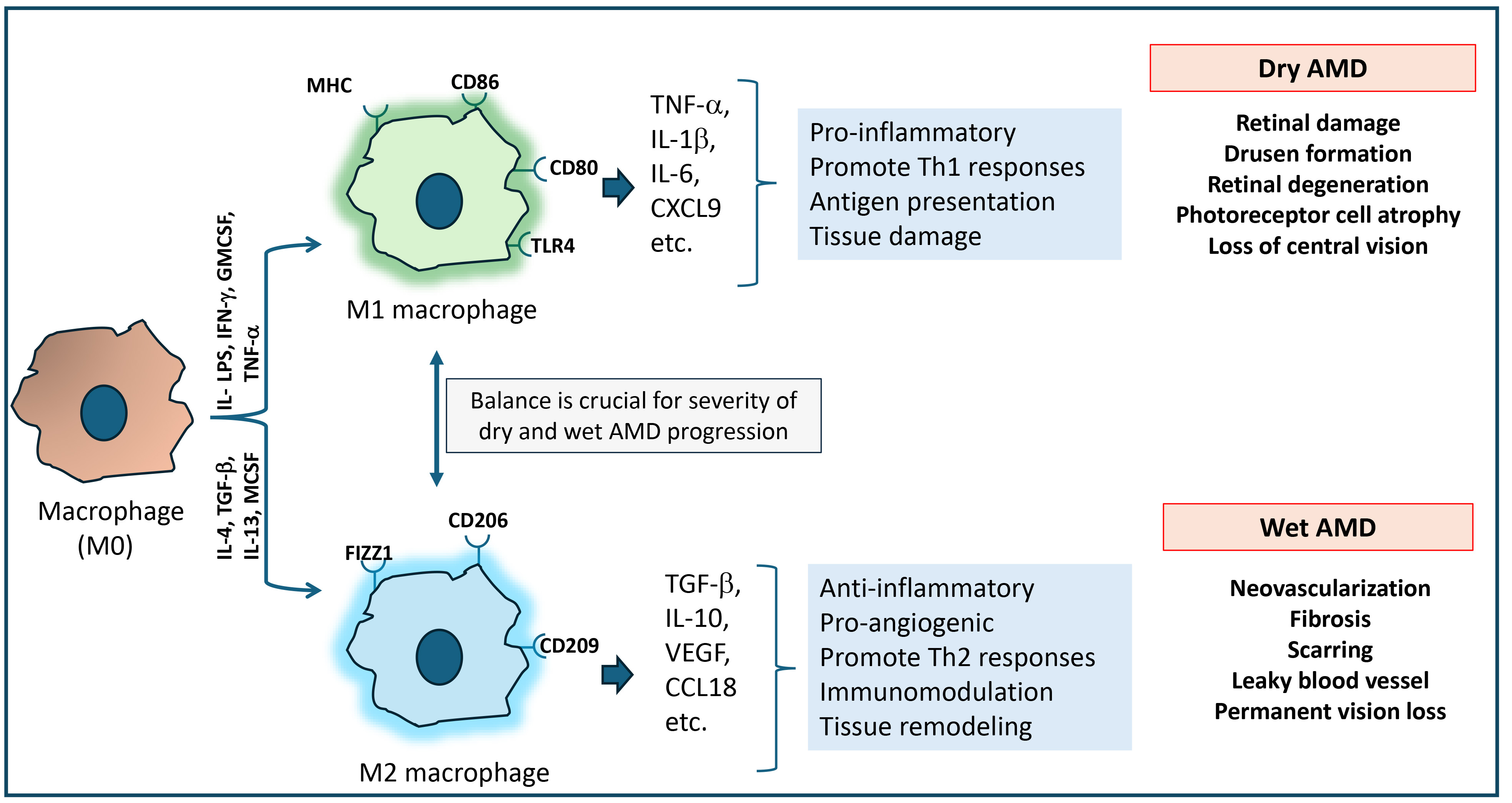 Fig. 5.
Fig. 5.
Macrophage-mediated inflammatory response in the development of dry and wet AMD. In age-related macular degeneration (AMD), macrophage polarization into M1 and M2 phenotypes contributes to two different forms of AMD. M1 phenotype macrophages are pro-inflammatory in nature. They release Th1 cytokines that cause chronic inflammation and tissue damage, leading to dry AMD. Meanwhile, M2 phenotype macrophages are anti-inflammatory in nature. They promote Th2 response, tissue remodeling and could contribute to angiogenesis, leading to wet AMD. The balance between M1 and M2 macrophage activity could play a critical role in the progression of both dry and wet AMD.
Few studies also indicate that pro-inflammatory M1 macrophages induce the inflammatory response to retinal injury and accelerate AMD complications [119, 120]. On the other hand, anti-inflammatory M2 macrophages are found to have a role in the early stages of inflammation and may clear the drusen. While several factors have been associated with the AMD risk, the pathogenesis of AMD is still not well known [120]. Therefore, more researches are required to define macrophages’ precise protective and harmful roles in AMD pathogenesis. However, the pathology of AMD lesions proves that macrophages do play an important role in the inflammation of AMD.
Recently, some clinical studies have investigated the therapeutic significance
of macrophage- mediated immune and inflammatory responses in ocular inflammatory
diseases [121, 122, 123, 124]. However, most of these researches are directed toward certain
diseases such as uveitis, diabetic retinopathy, and age-related macular
degeneration. Since inflammatory cytokines and chemokines released by macrophages
and other immune cells play a role in the development of ocular inflammatory
diseases, anti-cytokine inhibitors have been investigated as a potential therapy.
For example, Jaffe et al. [121] have performed a multinational phase-3
study to examine the therapeutic efficacy of adalimumab, a TNF-
Similarly, inhibition of M2 macrophage - released VEGF has been shown to prevent diabetic retinopathy as well as AMD. Yang et al. [127] have investigated the effect of a bispecific fusion protein (efdamrofusp), which neutralizes the VEGF isoforms and C3b and C4b complement proteins in dry AMD patients in a phase-1 clinical study. They found that the bispecific fusion protein prevents the infiltration and polarization of macrophages into the M2 phenotype and is effective in treating neovascular AMD. Similarly, Jia et al. [128] have also indicated the therapeutic significance of this bispecific fusion protein in treating neovascular AMD in a phase-1b clinical trial. Another study by Chang et al. [129] has demonstrated that intravitreal aflibercept (VEGF-A inhibitor) prevents treatment-resistant neovascular age-related macular degeneration. Similarly, Sarao et al. [130] have also indicated that intravitreal injections of aflibercept reverse the pre-switching trend toward losing vision and improve stable visual acuity for up to 12 months in patients with nAMD who are not responding to ranibizumab. In addition, several recent studies also suggest the significance of anti-VEGF therapies to control AMD [131, 132, 133].
Further, several studies have also suggested the use of anti-VEGF treatment to
control proliferative diabetic retinopathy and macular edema in patients
[134, 135, 136]. For example, in a randomized recovery trial, Wykoff et al.
[134] have shown that alibercept prevents retinal nonperfusion in patients with
proliferative diabetic retinopathy. Similarly, a long-term prospective study
conducted by Chatziralli et al. [135] has shown that intravitreal
treatment of ranibizumab, a VEGF-A inhibitor, prevents proliferative diabetic
retinopathy in patients with coexistent macular edema. In addition, intravitreal
injection of farcimab improves vision in patients with diabetic macular edema
[136]. In addition, several anti-cytokine therapies, such as TNF-
Macrophages have multifaceted functions in the development of ocular
inflammatory complications, such as uveitis, optic neuritis, scleritis, DR, and
AMD (Table 1, Ref. [12, 16, 24, 28, 30, 32, 33, 34, 35, 46, 47, 54, 55, 56, 57, 58, 59, 60, 62, 64, 65, 66, 69, 70, 71, 72, 74, 75, 76, 78, 82, 84, 85, 86, 88, 89, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 111, 112, 114, 115, 116, 117, 118, 120, 127, 128]). Understanding the role of macrophages will
shed light on developing novel immunomodulatory manipulation of macrophages for
future therapeutic advancements for ocular diseases. Further, a better
understanding and manipulation of macrophages is also required to associate
immune and inflammatory responses with other processes, such as oxidative stress
and angiogenesis, which are involved in the progression of ocular inflammatory
complications. Few studies also indicate the importance of macrophage
polarization in these diseases [32, 33, 34]. Generally, macrophages can adopt
pro-inflammatory (M1) or anti-inflammatory (M2) phenotypes depending on their
environment. M1 macrophages secrete TNF-
| Disease | Role of macrophages | Reference |
| Uveitis | [12, 16] | |
| [24, 32] | ||
| [28] | ||
| [30] | ||
| [33] | ||
| [34, 35] | ||
| [46, 47] | ||
| Optic Neuritis | [54, 55] | |
| [56] | ||
| [57, 58, 59] | ||
| [60, 62] | ||
| Retinitis | [66] | |
| [64, 65, 66] | ||
| [69, 70, 71, 72] | ||
| [74, 75, 76, 78] | ||
| [82] | ||
| Scleritis | [84, 86] | |
| [88] | ||
| [89] | ||
| [85] | ||
| DR | [94, 95, 96, 97] | |
| [98, 99] | ||
| [99] | ||
| [95, 100] | ||
| [101, 102, 103, 104, 105] | ||
| Glaucoma | [106] | |
| [111, 112] | ||
| [114] | ||
| [115] | ||
| AMD | [116, 117] | |
| [118, 120] | ||
| [127, 128] |
CCL2, chemokine ligand 2; CXCL10, C-X-C motif chemokine ligand 10; CD, cluster of differentiation; AhR, aryl hydrocarbon receptor; EAE, experimental autoimmune encephalomyelitis; HSV, herpes simplex virus; CMV, Cytomegalovirus; HZV, herpes zoster virus.
It is now well known that external and internal stimuli could activate
macrophages locally or systemically. The activation of macrophages triggers
immune and inflammatory responses, which drive inflammation in various ocular
tissues by releasing cytokines such as TNF-
In summary, recent studies suggest that macrophages could play an important role in the pathophysiology of ocular inflammatory diseases. They can act either independently or in conjunction with other immune cells and modify oxidative and inflammatory responses, leading to ocular tissue damage. Understanding macrophage activation, phenotype change, and immune cell modulation could help control immune and inflammatory responses, promote tissue repair, and improve vision. Developing novel therapeutic targets related to macrophages may hold great potential for improving vision problems associated with ocular inflammatory complications.
During the preparation of this manuscript, the authors used ChatGPT for spelling, grammar checks and language improvements. Afterward, the content was thoroughly reviewed and edited, with the authors take full responsibility for the publication and noting where AI was utilized.
AMD, Age-related macular degeneration; AIU, Autoimmune uveitis; AhR, aryl hydrocarbon receptor; Arg-1, arginase-1; CD4, clusters of differentiation 4; EAU, experimental autoimmune uveitis; EAE, experimental autoimmune encephalomyelitis; EBV, Epstein-Barr virus; CMV, cytomegalovirus; DR, diabetic retinopathy; HSV, herpes simplex virus; IL-1
SM wrote the initial draft, edited; TS wrote the initial draft, edited. KVR edited the final draft, conceptualized the idea on the topic. SM, TS and KVR also contributed to acquisition, analysis and interpretation of the literature and editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This research received no external funding.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.