






 , Cheng Lu 2,*
, Cheng Lu 2,* , Mingming Lv 2, Shengwang Du 1, Xinjun Wu 1
, Mingming Lv 2, Shengwang Du 1, Xinjun Wu 11 Department of General Surgery, Lianyungang Affiliated Hospital of Nanjing University of Traditional Chinese Medicine, 222000 Lianyungang, Jiangsu, China
2 Department of Breast, Nanjing Maternal and Child Health Hospital, 210094 Nanjing, Jiangsu, China
Abstract
Breast cancer (BC) is a prevalent malignancy among women, and numerous investigations have reported that platelet aggregation may play a role in BC progression. Thus, identifying new targets for BC is essential. In this regard, we focused on nucleolar protein 6 (NOL6), located on chromosome 9p13, which is implicated in tumor development.
To investigate NOL6 expression in BC, examine its role in platelet aggregation and angiogenesis, and elucidate the underlying mechanisms.
Bioinformatic analyses, immunoblotting, and quantitative real-time polymerase chain reaction (qPCR) were performed to assess NOL6 expression in BC. Cell counting kit-8 (CCK-8) and 5-ethynyl-2′-deoxyuridine (EdU) assays were conducted to determine the impact of NOL6 on BC cell proliferation. Immunostaining, enzyme-linked immunosorbent assay (ELISA), and flow cytometry (FCM) assays were utilized to analyze the effects of NOL6 on platelet aggregation. Tube formation and transwell assays were performed to examine angiogenesis and invasion, immunoblot assays were used to confirm the underlying mechanisms, and tumor growth assays in mice were conducted to validate the findings in vivo.
NOL6 was found to be highly expressed in BC and was associated with patient prognosis, platelet aggregation, and angiogenesis. Its knockdown inhibited BC cell proliferation and reduced platelet aggregation induced by BC cells. Additionally, NOL6 depletion impaired angiogenesis and migration of BC cells. In vivo studies confirmed that NOL6 promotes tumor growth. Mechanistically, NOL6 enhances the Twisted spiral transcription factor 1 (Twist1)/galectin-3 axis, contributing to BC progression.
NOL6 can promote tumor progression by facilitating platelet aggregation and angiogenesis in BC cells through the Twist1/galectin-3 axis.
Graphical Abstract
Keywords
- breast cancer (BC)
- Nucleolar Protein 6 (NOL6)
- platelet aggregation
- angiogenesis
- Twist1/galectin-3
Breast cancer (BC) is a prevalent malignancy among women worldwide, and although advances in its early detection have led to a decline in mortality rates, metastasis remains the leading cause of death among BC, accounting for over 90% of cancer-related fatalities [1]. Despite extensive research into deciphering its metastatic intricacies, the precise mechanisms are still not fully understood. Recently, platelet activation has emerged as a potential therapeutic target for BC [2], as inhibiting platelet function has been shown to suppress metastasis [3]. Although current literature suggests that platelet aggregation plays a significant role in BC progression [4], the mechanisms through which tumor cells instruct platelets to contribute to metastasis are not yet clear.
Nucleolar Protein 6 (NOL6) is a nucleolar protein located on chromosome 9p13 with a total length of 11434 bp [5]. It is mainly localized in the nucleolus of cells and plays a significant role in tumor biology. NOL6 is aberrantly overexpressed in prostate cancer, where it promotes cell proliferation and inhibits apoptosis [6]. In gastric cancer, NOL6 overexpression enhances cell viability and reduces apoptosis [6, 7]. Additionally, NOL6 supports endometrial cancer cell growth by regulating Twisted spiral transcription factor 1 (Twist1) expression [8]. Despite these findings, the role of NOL6 in BC remains unexplored, and its mechanisms are not well understood.
Analysis of The Cancer Genome Atlas (TCGA) data indicates that NOL6 is highly expressed in BC patients, with elevated levels correlating with poor prognosis [8]. NOL6 has been shown to upregulate TWIST1, which promotes the malignant phenotype of BC cells by enhancing their motility [7]. Furthermore, TWIST1 increases the expression of galectin-3 [4, 9]. Galectin-3, an anti-receptor for Glycoprotein VI (GPVI) on BC cells, facilitates platelet adhesion and aggregation, contributing to tumor progression [10, 11]. Despite these associations, the exact role of NOL6 in BC progression remains unclear.
In this study, we investigated the role of NOL6 in BC and found that NOL6 promoted the Twist1/galectin-3 axis, thereby facilitating platelet aggregation and angiogenesis in BC cells, which may contribute to the malignant phenotype of the disease. Thus, we propose NOL6 as a novel therapeutic target for BC.
The expression of NOL6 in breast cancer tissues was analyzed using the TCGA database (https://portal.gdc.cancer.gov/) and prognostic analysis was conducted using the Gene Expression Profiling Interactive Analysis (GEPIA) tool (http://gepia.cancer-pku.cn/).
This study was conducted in accordance with the principles outlined in the Declaration of Helsinki. The serum samples were collected from 40 BC patients and 20 healthy controls at Lianyungang Affiliated Hospital of Nanjing University of Traditional Chinese Medicine. This study was approved by our hospital’s Ethics Committee. The informed consent has been obtained by the patients or their families/legal guardians. Peripheral blood was processed using Ficoll lymphocyte isolation solution, and the clinical and pathological characteristics of the BC patients and healthy controls were assessed (Table 1).
| Cancer patients | Healthy subjects | p value | z value | t value | ||
| Female, no. (%) | 40 (100%) | 20 (100%) | / | |||
| Age, years | 60.00 (22.50) | 58.00 (14.50) | 0.236 | 1.185 | ||
| Median (IQR) | ||||||
| Stage, no. (%) | / | |||||
| I | 5 (12.5) | |||||
| II | 13 (32.5) | |||||
| III | 7 (17.5) | |||||
| Metastasis | 15 (37.5) | |||||
| Tumor size, no. (%) | / | |||||
| 10 (25) | ||||||
| 2–5 cm | 27 (67.5) | |||||
| 3 (7.5) | ||||||
| P-selectin, ng 10–8/cell | 66.50 (24.00) | 46.50 (8.50) | 5.915 | |||
| Median (IQR) | ||||||
| Blood coagulation function | ||||||
| Prothrombin time (PT), s | 8.35 (1.05) | 15.20 (1.35) | 6.263 | |||
| Median (IQR) | ||||||
| Activated partial thromboplastin time (APTT), s | 29.60 (2.83) | 43.25 (3.90) | 6.275 | |||
| Median (IQR) | ||||||
| Fibrinogen (FIB), g/L | 6.70 (1.73) | 3.50 (0.28) | 6.285 | |||
| Median (IQR) | ||||||
| D-D, µg/L | 876.28 |
331.00 |
45.162 | |||
| Mean |
||||||
| Indicators of Angiogenesis | ||||||
| VEGF, ng 10–8/cell | 102.00 (23.00) | 51.00 (12.50) | 6.196 | |||
| Median (IQR) | ||||||
| Ang-1, ng 10–8/cell | 31.00 (6.75) | 20.50 (9.00) | 5.063 | |||
| Median (IQR) | ||||||
IQR, Interquartile range; VEGF, vascular endothelial growth factor.
BC cell lines, including MCF-7 (SCSP-531), MDA-MB-231 (TCHu227), as well as the normal breast cell line HMEC (GNHu46), were purchased from Cell Bank, Chinese Academy of Sciences (Shanghai, China). SW527 (5922-60-1) was bought from ATCC (Manassas, VA, USA). They were maintained in Dulbecco’s Modified Eagle Medium (DMEM; Gibco, Carlsbad, CA, USA, Cat# 11965092, Lot# 2123456) supplemented with 10% fetal bovine serum (FBS; Gibco, Cat# 10099141, Lot# 3456789) and incubated at 37 °C in a 5% CO2 atmosphere. All cell lines were authenticated by short tandem repeats (STR) profiling and tested negative for mycoplasma. NOL6 shRNA plasmids were constructed in our lab. For gene knockdown or overexpression, plasmid containing DNA (pcDNA) plasmids or shRNA constructs targeting NOL6 (5′-CTGACCCTGGGACTCCTTC-3′), and negative control (sh-NC) (5′-GCAGGAATTGGAACATTAT-3′) were transfected into MCF-7 cells using Lipofectamine 2000 (Invitrogen, Shanghai, China). Transfection efficiency was assessed by quantitative real-time polymerase chain reaction (qPCR) and immunoblotting 48 hours post-transfection.
The cell samples were lysed using RIPA buffer (Beyotime, Beijng, China). Protein concentration was assessed using a Bovine serum albumin (BCA) protein assay kit (Thermo Fisher Scientific, Waltham, MA, USA). Equal amounts of protein (30 µg) were separated by 10% SDS-PAGE and transferred to PVDF membranes (Millipore, Burlington, MA, USA). Membranes were blocked with 5% dry milk and incubated with primary antibodies against Nucleolar Protein 6 (NOL6) (Abcam, Cambridge, UK, ab228836; 1:500), Twist1 (Abcam, ab50887; 1:1000), galectin-3 (Abcam, ab76245; 1:500), platelet-derived growth factor (PDGF) (Abcam, ab178439; 1:1000), Vascular endothelial growth factor (VEGF) (Abcam, ab291246; 1:500), and beta-actin (Abcam, ab8226; 1:3000). After incubation with Horseradish peroxidase (HRP)-conjugated secondary antibodies (Goat Anti-Rabbit antibody, Abcam, ab6721; 1:2000), the protein bands were visualized using the enhanced chemiluminescence (ECL) detection system (RPN2106, GE, Marlborough, MA, USA).
RNA was extracted from tissue and cell samples using Trizol reagent (TaKaRa, Kusatsu City, Japan), RNA concentration and purity were assessed using a NanoDrop spectrophotometer (Thermo Fisher Scientific, USA), total RNA was reverse transcribed into cDNA using the RT reagent Kit (Takara), and quantitative PCR was performed using SYBR Ex Taq™ II (Takara). The primer sequences used were: NOL6: Forward: 5′-AGGAAGCGTACATTGGCTGAA-3′, Reverse: 5′-GTCTCCCGAAGGCGATTAAGC-3′; TWIST1: Forward: 5′-GTCCGCAGTCTTACGAGGAG-3′, Reverse: 5′-GCTTGAGGGTCTGAATCTTGCT-3′; Galectin-3: Forward: 5′-CTTATTAACCTGCCTTTGCCTGG-3′, Reverse: 5′-GCAACATCATTCCCTCTTTGGA-3′; GAPDH: Forward: 5′-TGTGGGCATCAATGGATTTGG-3′, Reverse: 5′-ACACCATGTATTCCGGGTCAAT-3′. In addition, the relative gene expression was calculated using the 2-ΔΔCt method, with GAPDH as the normalization control.
For the cell counting kit-8 (CCK-8) assay, the cells were seeded into 96-well plates and incubated at 37 °C. After 1, 2, 3, 4, 5, and 6 days, the cells were treated with CCK-8 reagent and incubated for 4 hours at 37 °C. OD value at 450 nm was measured using a Bio-Rad spectrophotometer (Berkeley, CA, USA).
For the 5-ethynyl-2′-deoxyuridine (EdU) assay, the cells were plated in 24-well plates and incubated at 37 °C for 24 hours. Subsequently, the cells were treated with EdU reagent (10 µM; RiboBio, Guangzhou, China) for 2 hours. Cells were then fixed with 4% paraformaldehyde, permeabilized with 0.5% Triton X-100, and stained with Apollo® reaction cocktail (RiboBio, China) according to the manufacturer’s instructions. Nuclei were counterstained with DAPI (Sigma-Aldrich, Shanghai, China), and images were captured using a fluorescence microscope (Zeiss, Oberkochen, Germany).
BC cells were fixed with 4% paraformaldehyde (PFA) and blocked with 5% bovine serum albumin (BSA). Cells were then incubated with primary antibodies against CD41 (Abcam, ab134131; 1:200) and subsequently with secondary antibodies conjugated with Alexa 555 (Invitrogen, 1:1000, CA, USA) and phalloidin-FITC (Abcam, ab235137, 1:1000, UK). The nuclei were counterstained with DAPI for 5 minutes. Images were acquired using a fluorescence microscope (Zeiss, Germany).
The MCF-7 cells were washed with PBS and fixed. They were then stained with anti-GPIb-IX (CD42b) antibody (BD Biosciences, Franklin Lake, NJ, USA, Cat# 551061, Lot# 123456) and anti-GP-IIb (CD41) antibody (BD Biosciences, Cat# 551061, Lot# 789012) for 20 minutes, and their expression levels were measured using a BD FACSCalibur (Franklin Lakes, NJ, USA) according to the manufacturer’s instructions.
The levels of T-box Protein 2 (TXB2) (Abcam, ab133054) and ATP production (Abcam, ab113849) in serum samples were measured using Enzyme-Linked Immunosorbent Assay (ELISA) kits (Abcam) following the manufacturer’s guidelines.
Transwell inserts (BD Falcon) were used with 24-well plates as the lower
chambers. For migration assays, the inserts were left uncoated. For invasion
assays, inserts were coated with 100 µL of Matrigel (diluted 1:3 with
serum-free media). Cells (1
HMEC cells were plated onto 24-well plates pre-coated with 50% Matrigel and cultured in medium from the MCF-7 cells upon the indicated transfections. Tube formation was assessed 4 hours later by capturing images with a fluorescence microscope (Carl Zeiss Inc, Oberkochen, Germany).
Nude mice (6-week-old female BALB/c nu/nu, purchased from Beijing Vital River
Laboratory Animal Technology Co., Ltd., Beijing, China) were subcutaneously
injected with 1
Tissue samples were sliced, dehydrated through absolute alcohol, and rehydrated. The sections were fixed with 4% paraformaldehyde for 30 minutes, blocked with 2% BSA and then incubated with primary antibodies against CD41 (Abcam, ab134131; 1:200) and CD31 (Abcam, ab28364; 1:500) for 2 hours. All sections were incubated with HRP antibody for 1.5 h followed by adding diaminobenzidine in 5 min, and all sections were then counterstained with hematoxylin.
Statistical analysis was performed using GraphPad 6.0 (GraphPad Software, Inc., Boston, MA, USA). For
measurement data, the data were first assessed for normality using the
Shapiro-Wilk test. For data that did not conform to a normal distribution,
results are presented as medians with interquartile ranges, non-parametric tests
were employed. Specifically, the Mann-Whitney U test was used to compare
differences between two independent groups, and the Kruskal-Wallis test was
applied for comparisons across multiple groups. Data are presented as mean
To investigate NOL6 expression in BC, we analyzed transcript per million (TPM)
values from the TCGA database. The analysis revealed that NOL6 expression was
significantly elevated in BC tissues compared to normal tissues (p
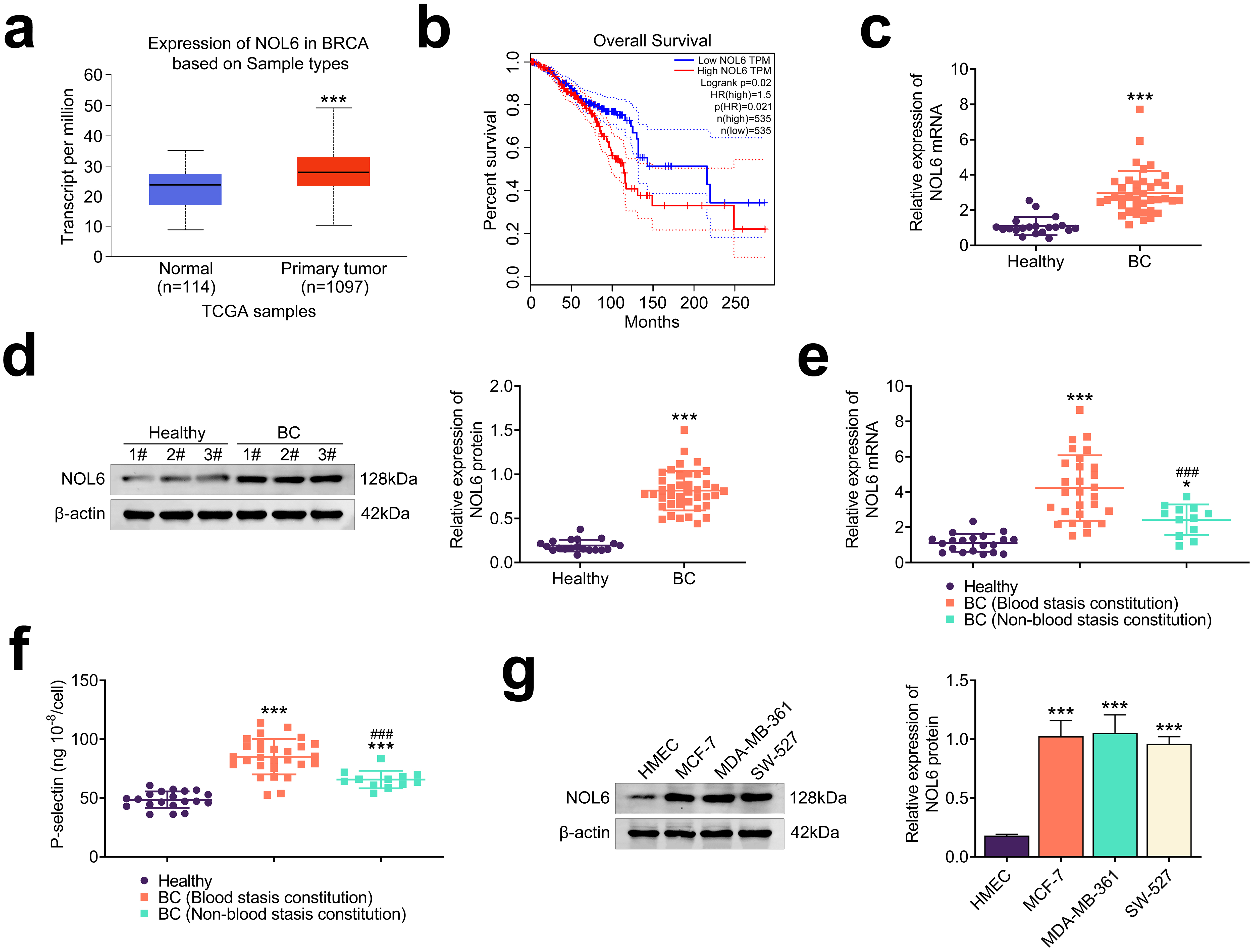 Fig. 1.
Fig. 1.
NOL6 is highly expressed in BC. (a) TCGA database analysis of
TPM values for NOL6 in 1097 tumor tissues and 114 normal tissues. ***p
To elucidate the role of NOL6 in BC cell proliferation, we used shRNA and
overexpression plasmids to modulate NOL6 expression in MCF-7 cells. We found that
transfection with NOL6 overexpression plasmids resulted in a significant increase
in NOL6 expression, while transfection with NOL6 shRNA plasmids led to a marked
decrease in its expression (p
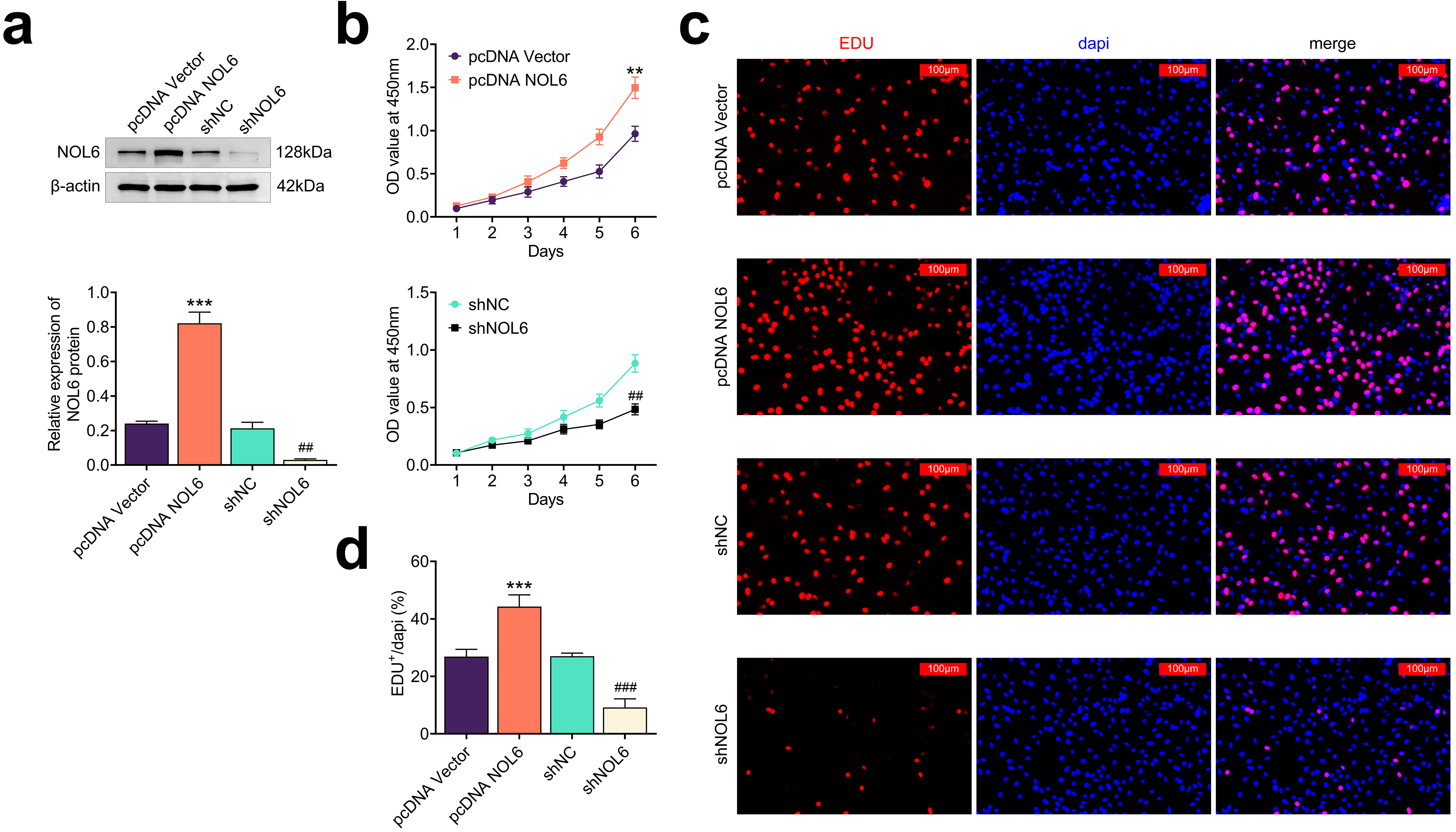 Fig. 2.
Fig. 2.
Knockdown of NOL6 inhibits the growth of BC cells. (a)
Immunoblot analysis showing NOL6 expression levels in MCF-7 cells following the
indicated transfections. (b) CCK-8 assay results depicting the growth of MCF-7
cells with different NOL6 expression levels over 6 days, measured by OD450. (c)
Edu assay images showing MCF-7 cell proliferation following the indicated
transfections, with (d) quantification of Edu-positive cells per field. Scale
bar, 100 µm. Each experiment was conducted in triplicate. **p
Building on the observation that NOL6 promotes platelet aggregation in BC
patients (Fig. 1f), we investigated how NOL6 influences platelet aggregation
induced by BC cells. Initially, MCF-7 cells were co-cultured with human
platelets, and immunostaining assays were performed to assess the expression of
the platelet aggregation marker CD41. The results showed that NOL6 overexpression
in MCF-7 cells resulted in elevated CD41 levels (p
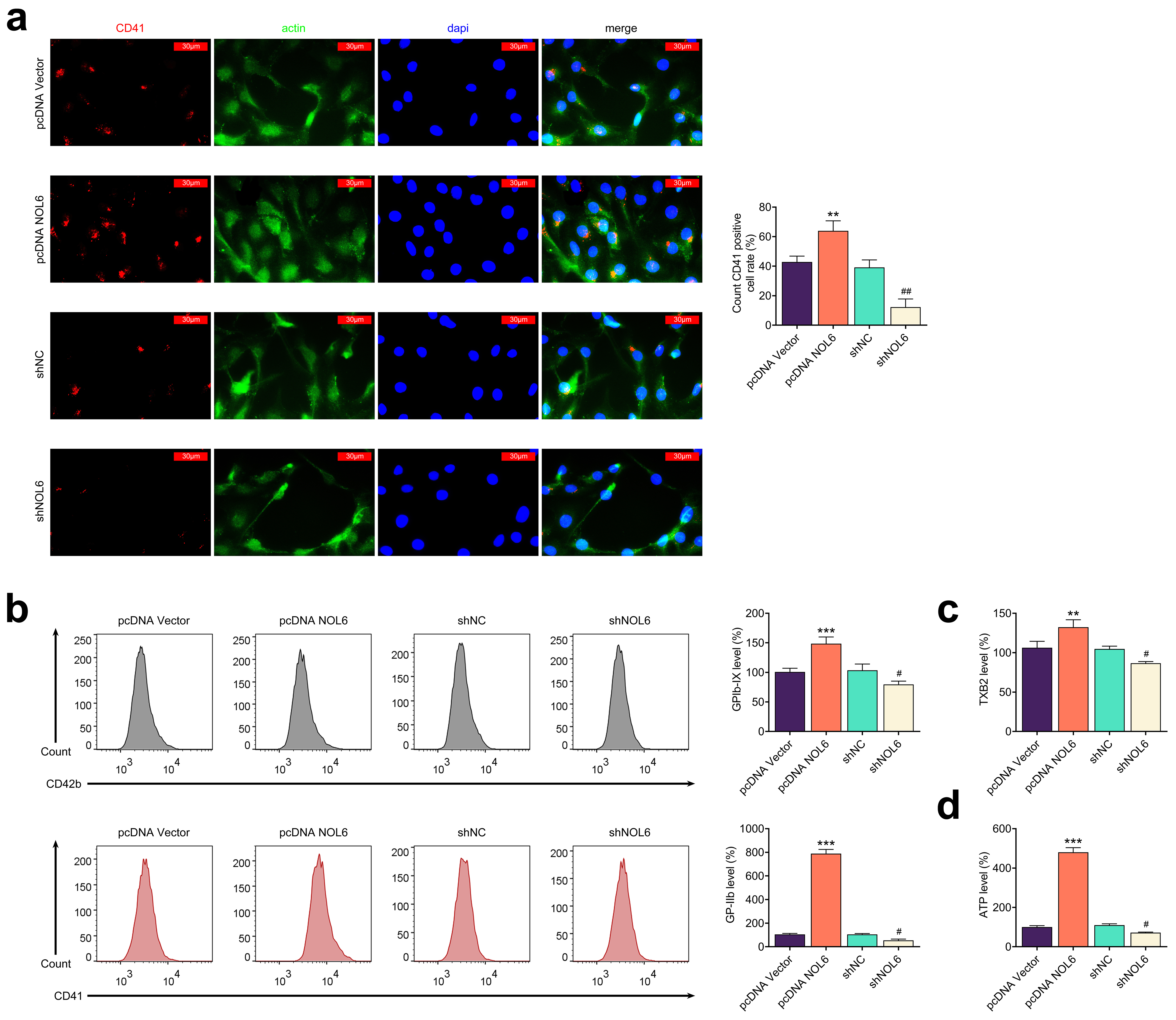 Fig. 3.
Fig. 3.
Knockdown of NOL6 inhibits platelet aggregation induced by BC
cells. (a) Immunostaining of CD41 in MCF-7 cells with the indicated
transfections. Red panel indicates CD41, green panel indicates actin, and blue
panel indicates DAPI. Scale bar, 30 µm. (b) Flow cytometry (FCM) analysis
of GPIb-IX and GP-IIb expression levels in MCF-7 cells following different
transfections. (c) Enzyme-Linked Immunosorbent Assay (ELISA) results showing TXB2
levels in MCF-7 cells with altered NOL6 expression. (d) Measurement of ATP levels
in MCF-7 cells upon indicated transfections. Each experiment was conducted in
triplicate. **p
To evaluate the impact of NOL6 on BC cell motility and angiogenesis, we
performed several assays. Transwell assays revealed that NOL6 overexpression
significantly enhanced the migration and invasion of MCF-7 cells (p
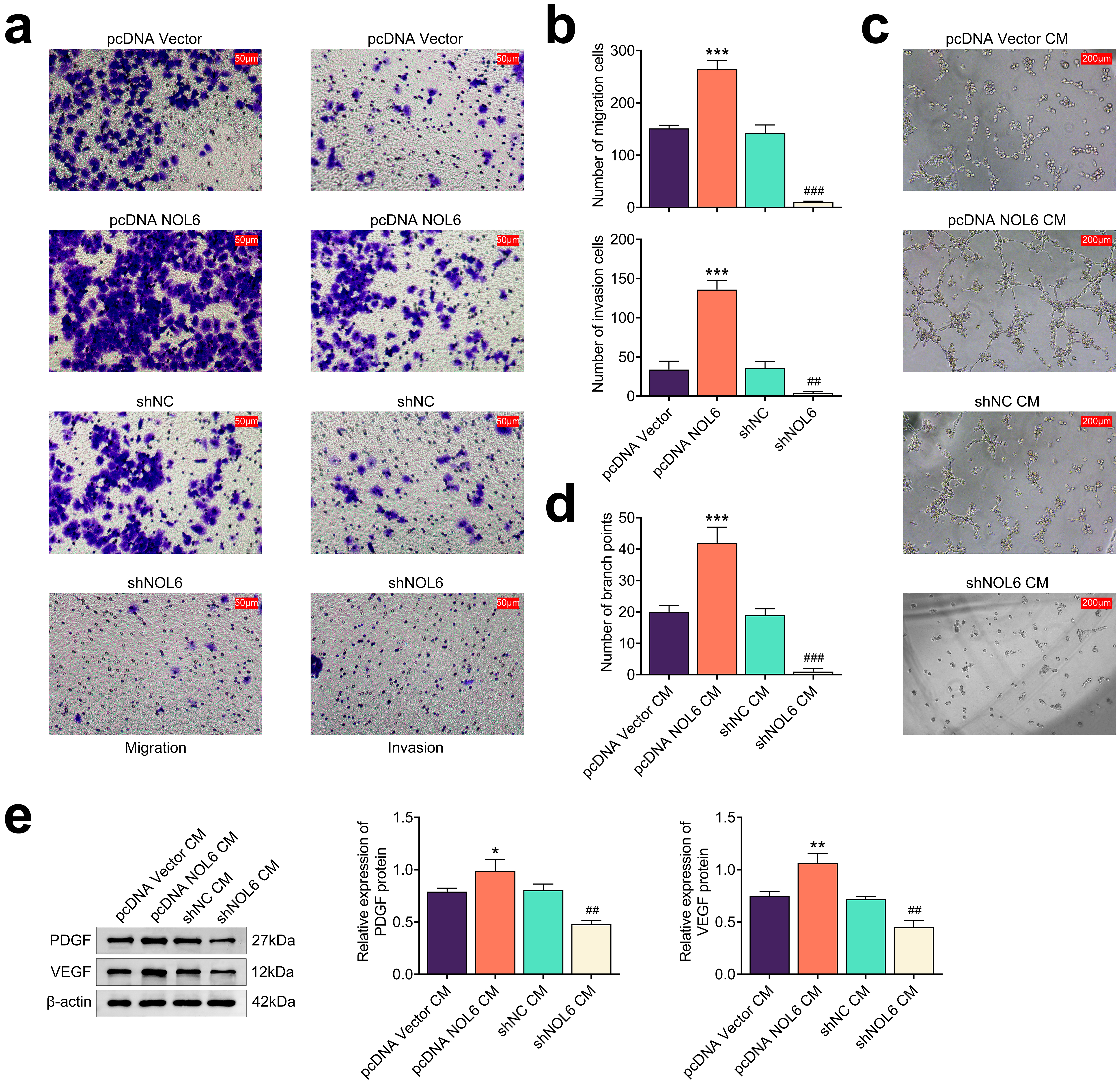 Fig. 4.
Fig. 4.
Knockdown of NOL6 inhibits angiogenesis and motility of BC
cells. (a) Transwell assays assessing the migration and invasion of MCF-7 cells
following NOL6 modulation. Scale bar, 50 µm. (b) Quantification of
migration and invasion cell counts from panel (a). (c) Tube formation assays
illustrating the effects of NOL6 expression of MCF-7 cells and its culture medium
on human umbilical vein endothelial cell (HUVEC) angiogenesis. Scale bar, 200
µm. CM, culture medium. (d) Quantification of branch points from panel (c).
(e) Immunoblot analysis showing platelet-derived growth factor (PDGF) and
vascular endothelial growth factor (VEGF) expression levels in HUVECs in response
to NOL6 modulation in MCF-7 cells. Each experiment was conducted in triplicate.
*p
Next, we explored the underlying mechanism by which NOL6 influences BC through
immunoblot analyses. We observed that overexpression of NOL6 resulted in
increased levels of Twist1 and galectin-3 in MCF-7 cells, while NOL6 depletion
led to reduced expression of these proteins, suggesting a regulatory role of NOL6
in the Twist1/galectin-3 axis (p
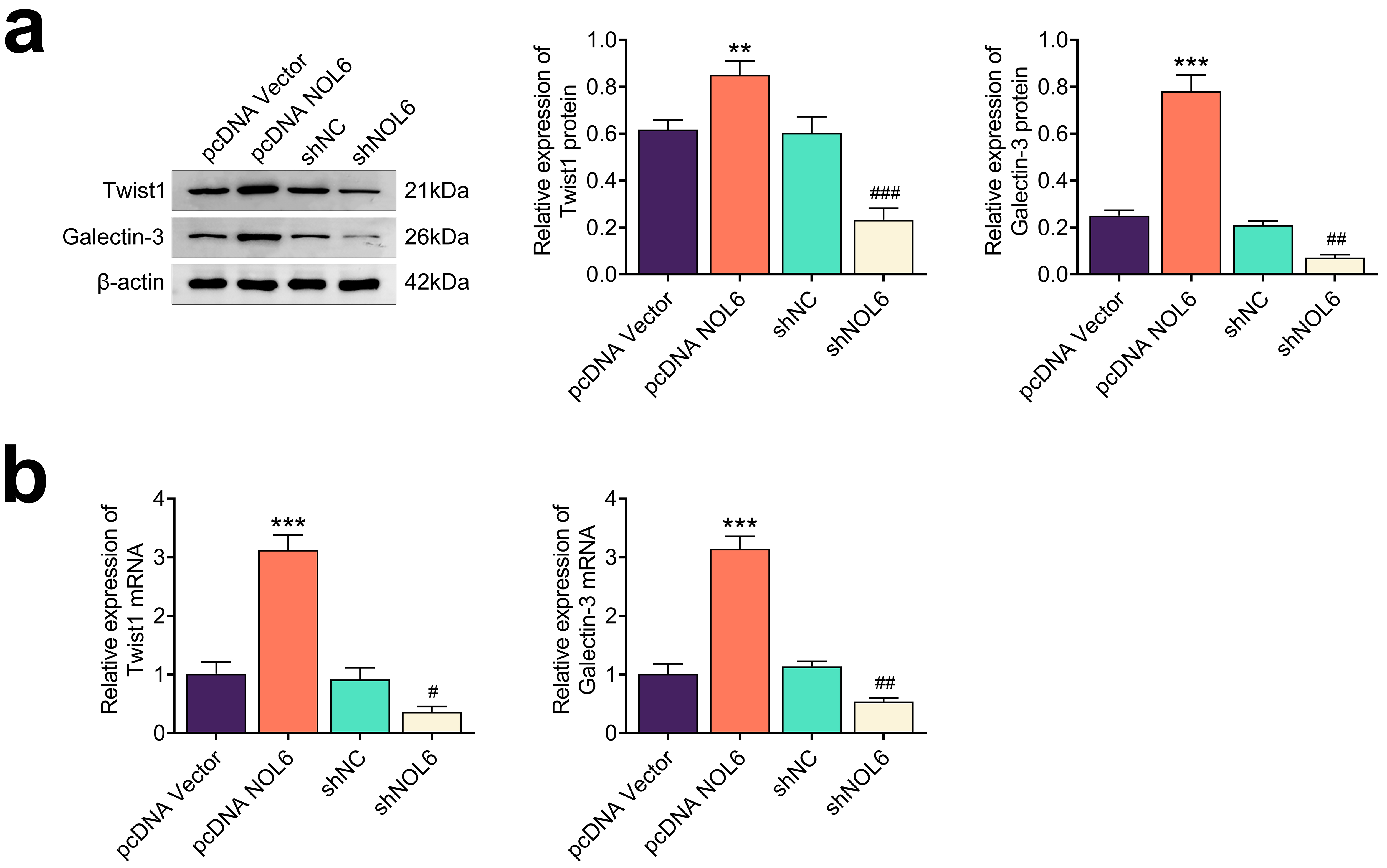 Fig. 5.
Fig. 5.
NOL6 promotes the Twist1/galectin-3 axis in BC. (a) Immunoblot
analysis of Twist1 and Galectin-3 expression in MCF-7 cells following NOL6
modulation. (b) qPCR assays assessing the mRNA levels of Twist1 and
galectin-3 in MCF-7 cells upon NOL6 modulation. Each experiment was
conducted in triplicate. **p
Additionally, we investigated whether the effects of NOL6 depletion could be
reversed by Twist1 overexpression. The results from both immunoblot and qPCR
assays demonstrated that Twist1 overexpression rescued the expression levels of
Twist1 and galectin-3 reduced by NOL6 depletion (p
To assess the impact of NOL6 on BC tumor growth in vivo, we utilized
shRNAs targeting NOL6 to decrease its expression in MCF-7 cells. These
NOL6-depleted MCF-7 cells were then injected subcutaneously into the abdomen of
nude mice. After 20 days, the results demonstrated that NOL6 ablation led to a
significant suppression of tumor growth, as evidenced by reduced tumor volume and
weight (p
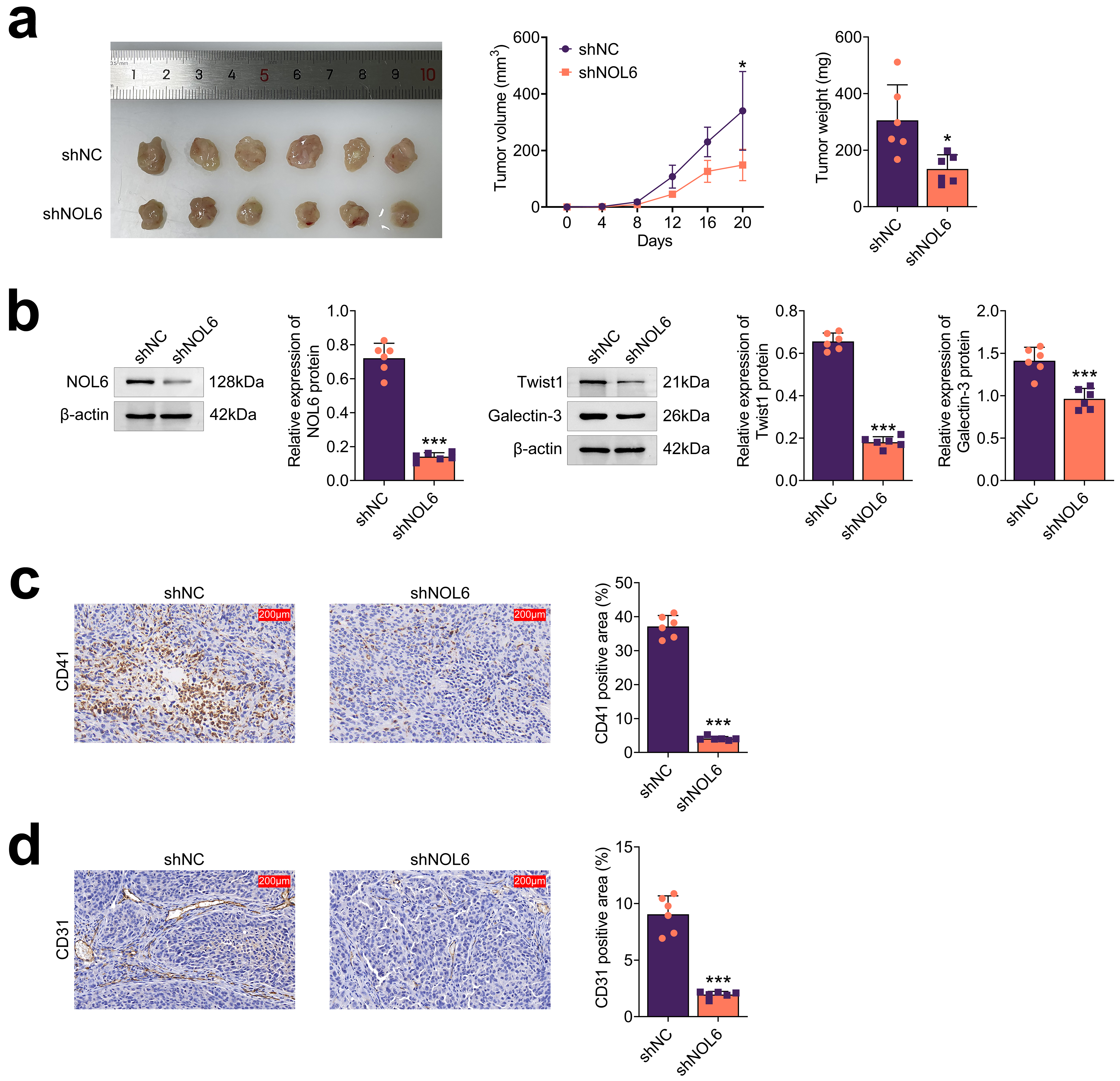 Fig. 6.
Fig. 6.
NOL6 inhibits the growth of BC in vivo. (a) Tumor
growth assays illustrating the effects of NOL6 on BC progression in
vivo. Representative images of tumors are shown (left), with tumor volume and
weight measured every 4 days over 20 days (n = 6 per group). (b) Immunoblot
analysis displaying the expression of NOL6 (left), Twist1, and Galectin-3 (right)
in tumor tissues from the indicated groups. (c) Immunohistochemical (IHC) assays
showing CD41 expression in tumor tissues, with quantitative analysis included.
Scale bar, 200 µm. (d) IHC assays showing CD31 expression in tumor tissues,
with quantitative analysis included. Scale bar, 200 µm. Each experiment was
performed in triplicate. *p
BC treatment typically involves a comprehensive approach tailored to individual patient profiles [12]. While targeted therapy represents a promising strategy for advanced BC, further research into the underlying mechanisms and the development of novel therapeutic targets is crucial. Notably, abnormal coagulation indices in BC patients suggest a dysregulation in coagulation, which may contribute to tumor progression and metastasis [13]. Platelets play a significant role in the survival and dissemination of tumor cells. The interaction between circulating tumor cells and platelets promotes metastasis through physical contact and bidirectional activation [14]. Tumor cell-induced platelet aggregation (TCIPA) facilitates immune evasion and enhances tumor cell adhesion and invasion [15]. Additionally, tumor-induced platelets release soluble mediators that increase tumor cell invasiveness and bone metastasis in animal models [16, 17]. In this present study, our findings indicate that NOL6 knockdown inhibits platelet aggregation induced by BC cells, thereby suppressing both tumor progression and metastasis.
Angiogenesis plays an essential role in BC development. This complex biological process involves various molecules, including cytokines, growth factors, receptor proteins, cell membrane molecules, transcription factors, and signaling pathway molecules [18]. Understanding the roles and interactions of these molecules is essential for gaining insights into the biological characteristics of BC angiogenesis. Such knowledge can offer new approaches and strategies for BC treatment. Endothelial cell growth and directional migration are critical aspects of angiogenesis [19]. In this study, we demonstrate that the ablation of NOL6 impairs both angiogenesis and the migration of BC cells.
NOL6 is a nucleolar protein that has been highly conserved throughout evolution. It is closely associated with chromosome condensation during mitosis. Additionally, NOL6/NRAP protein stress has been reported to activate p53/CP-1-mediated immune mechanisms, thereby influencing tumor progression [6]. It has been shown that NOL6 plays an important role in cell cycle maintenance and tumor cell invasiveness. Investigating the relationship and mechanisms between NOL6 expression and BC is crucial for developing effective treatment strategies [5]. Our data confirm the high expression of NOL6 in BC, which contributes to various processes, including platelet aggregation and angiogenesis, which facilitate BC progression.
Galectin-3, a multifunctional proto-oncoprotein, plays a significant role in regulating tumor cell growth, invasion, angiogenesis, and apoptosis. It has potential as a novel diagnostic marker for tumor metastasis and is a candidate for targeted cancer therapies [20]. Additionally, Twist1, an upregulated marker in dormant BC cells [9, 21], is closely associated with invasion, metastasis, prognosis, drug resistance, and angiogenesis in BC [22]. Twist1 enhances the expression of the proto-oncogene AKT2 by binding to its promoter, which in turn activates YB-1, a protein involved in cell growth, thus promoting tumor cell proliferation [23]. Our findings indicate that NOL6 influences platelet aggregation, angiogenesis, motility, and tumor growth in BC through the Twist1/galectin-3 axis.
In conclusion, our study demonstrates that NOL6 is highly expressed in BC and facilitates platelet aggregation and angiogenesis through the Twist1/galectin-3 axis, supporting NOL6 as a potential therapeutic target in BC.
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.
TZ, CL, ML, SD and XW contributed to the study conception and design. Material preparation and the experiments were performed by TZ. Data collection and analysis were performed by CL, ML and SD. The first draft of the manuscript was written by XW and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript. All authors contributed to editorial changes in the manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
The serum samples study has been approved by the Ethics Committee of the Lianyungang Affiliated Hospital of Nanjing University of Chinese Medicine (Approval No.: 2023-13). Written informed consent was obtained from legally authorized representative(s) for anonymized patient information to be published in this article. This study was conducted in accordance with the principles outlined in the Declaration of Helsinki. All animal experiments were approved by the Ethics Committee of Lianyungang Affiliated Hospital of Nanjing University of Traditional Chinese Medicine for the use of animals and conducted in accordance with the National Institutes of Health Laboratory Animal Care and Use Guidelines.
Not applicable.
This work was supported by the present study was financially supported by the Natural Science Foundation of Jiangsu Province (Grant No. BK20211018) and Science and Technology project of Lianyungang Administration of Traditional Chinese Medicine (Grant No. LZYYB2 02306).
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/FBL25361.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.