






 , Pavel Spirin 1,2, Vladimir Prassolov 1,2, Olga Leonova 1
, Pavel Spirin 1,2, Vladimir Prassolov 1,2, Olga Leonova 11 Department of Cancer Cell Biology, Engelhardt Institute of Molecular Biology, Russian Academy of Sciences, 119991 Moscow, Russia
2 Center for Precision Genome Editing and General Technologies for Biomedicine, Engelhardt Institute of Molecular Biology, Russian Academy of Sciences, 119991 Moscow, Russia
Abstract
In the twentieth century, the textbook idea of packaging genomic material in the cell nucleus and metaphase chromosomes was the presence of a hierarchy of structural levels of chromatin organization: nucleosomes – nucleosomal fibrils –30 nm fibrils – chromomeres – chromonemata – mitotic chromosomes. Chromomeres were observed in partially decondensed chromosomes and interphase chromatin as ~100 nm globular structures. They were thought to consist of loops of chromatin fibres attached at their bases to a central protein core. However, Hi-C and other related methods led to a new concept of chromatin organization in the nuclei of higher eukaryotes, according to which nucleosomal fibrils themselves determine the spatial configuration of chromatin in the form of topologically associating domains (TADs), which are formed by a loop extrusion process and are regions whose DNA sequences preferentially contact each other. Somatic macronuclei of ciliates are transcriptionally active, highly polyploid nuclei. A feature of macronuclei is that their genome is represented by a large number of “gene-sized” (~1–25 kb) or of “subchromosomal” (~50–1700 kb) size minichromosomes. The inactive macronuclear chromatin of “subchromosomal” ciliates usually looks like bodies 100–200 nm in size. The aim of this work was to find out which of the models (chromomeres or TADs) is more consistent with the confocal and electron microscopic data on structural organization of chromatin bodies.
Macronuclear chromatin of four “subchromosomal” ciliate species (Bursaria truncatella, Paramecium multimicronucleatum, Didinium nasutum, Climacostomum virens) was examined using electron microscopy and confocal microscopy during regular growth, starvation and encystment.
Chromatin bodies ~70–200 nm in size observed in the interphase macronuclei consisted of tightly packed nucleosomes. Some of them were interconnected by one or more chromatin fibrils. Under hypotonic conditions in vitro, chromatin bodies decompacted, forming rosette-shaped structures of chromatin fibrils around an electron-dense centre. When the activity of the macronucleus decreased during starvation or encystment, chromatin bodies assembled into chromonema-like fibrils 100–300 nm thick. This data allows us to consider chromatin bodies as analogues of chromomeres. On the other hand, most likely, the formation of DNA loops in chromatin bodies occurs by the loop extrusion as in TADs.
The data obtained is well explained by the model, according to which the chromatin bodies of ciliate macronuclei combine features inherent in both chromomeres and TADs; that is, they can be considered as chromomeres with loops packed in the same way as the loops in TADs.
Keywords
- ciliates
- macronucleus
- chromatin
- chromomeres
- TADs
- electron microscopy
- confocal microscopy
The structural organization of chromatin in the space of the cell nucleus is an important factor regulating genome expression. Large-scale alterations in chromatin structure are associated with cancer, neurological and other diseases (reviews [1, 2]). In the twentieth century, based on electron microscopic and biochemical data, several models of chromatin organization in the cell nucleus and metaphase chromosomes were proposed. The textbook idea of packaging genomic material in the cell nucleus and metaphase chromosomes was the presence of a hierarchy of structural levels of chromatin organization. It was customary to distinguish the following levels of chromatin organization: nucleosomes – nucleosomal fibrils about 10 nm thick –30 nm fibrils – chromomeres – chromonemata (fibrous structures 100 to 200 nm thick) –300 to 700 nm chromatids – mitotic chromosomes [3, 4, 5]. The first level of genomic compaction is the nucleosome. The nucleosome is the basic repeating structural unit of eukaryotic chromatin. The nucleosome core consists of approximately 147 DNA base pairs wrapped around an octamer of histone proteins H2a, H2b, H3, H4. In the nucleosome fibril, the nucleosome cores are connected to each other by linker DNA in a complex with histone H1 and/or other proteins. It has long been assumed that the nucleosome fibre folds into a 30-nm fibril. However, recent cryo-electron microscopy, super-resolution microscopy, and small-angle X-ray scattering studies have not revealed 30 nm fibres in interphase chromatin or mitotic chromosomes in different cells, i.e., supranucleosomal levels of chromatin organization are constructed from 10-nm nucleosomal fibrils that do not fold into 30 nm fibres [6, 7, 8, 9, 10].
Chromomers and chromonemata were observed in the partially decondensed mitotic chromosomes [3, 11, 12]. It has been shown that chromonemata, which look like thick threads 100–300 nm thick, may consist of tightly packed globules about 100 nm in diameter, called chromomeres [3]. Compact chromomeres were also observed in isolated rat hepatocyte nuclei after incubation in 4 mM MgCl2 [13]. It can be estimated that a chromomere 100–200 nm in diameter contains 150–1200 kbp of DNA [14]. When incubated in buffers containing low (0.1–0.2 mM) concentrations of divalent cations, chromomeres gradually decompact, taking the form of rosette-like structures with loop-shaped deoxyribonucleoprotein (DNP) fibrils emerging from the electron-dense core [3]. Rosette-shaped structures have also been observed in ultrathin sections of partially deproteinized metaphase chromosomes [15] and in spread preparations of deproteinized chromatin from interphase nuclei [13, 16, 17] and mitotic chromosomes [16]. It was assumed that loops of chromatin fibrils are attached to the chromomere core, consisting of nuclear matrix proteins or, according to the model proposed by Cook [18], to proteins forming so-called “transcription factories”. Cook proposed a clear and simple model for the formation of chromonemal fibrils, based on the fact that the adhesive properties of the central core of chromomeres and nucleosomal fibrils may differ. When two more adhesive chromomere cores stick together, the less adhesive nucleosomal fibrils looping from them force additional cores to bind at the ends, elongating the chromonema in the form of a cylinder [18].
However, a number of reasons prevent us from considering chromomeres as structures that actually exist in the interphase nuclei of higher eukaryotes. First, chromomeres, like 30 nm fibrils, were not detected in human mitotic chromosomes studied using cryoelectron microscopy [6, 9]. Moreover, it has been shown that in compact metaphase chromosomes, the nucleosomal fibre is in a highly disordered state, comparable to a melted polymer [6]. Second, compact and rosette-like structures were detected only in partially decompacted or deproteinized preparations. Third, the presence of chromomeres in mammalian cell nuclei is not confirmed by modern experimental approaches. Therefore, it is currently accepted that chromomeres are absent in the nuclei of higher eukaryotes, and the structures described above are either artifacts of fixation and preparation of specimens for electron microscopy, or are caused by the presence of divalent cations in the medium.
In the 21st century, the invention of “chromosome conformation capture” (3C) methods [19], including its Hi-C modification, has led to a revolutionary revision of our knowledge about the spatial organization of chromatin in the nucleus. Common to all 3C methods is the step of fixing the cells with formaldehyde, which is a mild fixative compared to the strong fixatives such as glutaraldehyde and osmium tetroxide commonly used in electron microscopy. After a series of complex biochemical procedures, a sequencing step, and computer analysis of the data obtained, a matrix of contact frequencies between different genomic loci (heatmap) is constructed.
An analysis of Hi-C heatmaps shows that the mammalian interphase genome is organized into two distinct classes of domains: topologically associated domains (TADs) and compartments. There is a transcriptionally active A-compartment and a transcriptionally inactive B-compartment. They most likely correspond to the euchromatin and heterochromatin regions observed by electron microscopy in ultrathin sections. The compartments are formed by liquid-solid phase separation (LSPS) and liquid-liquid phase separation (LLPS) processes [20, 21, 22]. The N-terminal tail domains of core histones and their post-translational modifications play an important role in phase separation processes [22].
TADs are currently considered to be the fundamental units of genome organization in eukaryotic nuclei. They have been found in mammals, insects and birds, but are absent in bacteria [23]. TADs are loop genomic domains in which DNA sequences interact with each other more frequently than with sequences outside the domain [24, 25, 26]. The size of TADs in various eukaryotic cells is typically between 100 and 1000 kb [22, 23]. It is currently believed that TADs are formed by a loop extrusion process, which is mediated by the structural maintenance of chromosomes (SMC) protein complexes, such as cohesin and condensin [27, 28]. In contrast to compartments, TADs have distinct boundaries, which are often bound by CCCTC- binding factor (CTCF) [27, 28].
Thus, according to modern concepts, the organization of the genome in the nucleus is determined by the processes of phase separation and loop extrusion, and not by the presence of levels of chromatin organization of different hierarchies.
Hi-C data cannot provide direct information on the structure and size of TADs. However, their size can be estimated to be approximately ~200 nm. For example, Bintu et al. [29] observed spatially separated globular domains of ~200–300 nm in size on a stretch of human chromosome 21 using stochastic optical reconstruction microscopy (STORM) and diffraction-limited microscopy. Miron et al. [30] identified chains of interconnected 200–300 nm chromatin domains consisting of aggregated nucleosomes in living and fixed mouse and HeLa cells using scanning electron and 3D super-resolution microscopy. Punctate chromatin particles of approximately 200 nm in diameter [31] and compact chromatin domains (~160 nm in diameter) consisting of nucleosomes [32] were observed in both interphase chromatin and mitotic chromosomes in experiments with stimulated emission microscopy (STED) [31] and photoactivated localization microscopy (PALM) [32]. In addition, well-defined domains of approximately 160 nm in diameter exhibiting polymeric fractal-like behavior were observed using the Chromatin Scanning Transmission Electron Microscopy (ChromSTEM) method [33].
Although chromomeres and TADs are similar in size and the amount of DNA compacted in them, they differ fundamentally in their structural organization. Chromomeres contain an electron-dense core to which loops of chromatin fibrils are attached. In TADs, loops are formed by loop extrusion involving cohesin and CTCF. The TAD structure appears to correspond to a fractal globule conformation [34, 35]. Fractal globules do not contain entangled nucleosomal fibrils. Such a globule can easily be symmetrically decompacted or expose the genomic locus on its surface for transcription without decompacting [34, 35]. In this case, transcription can occur either on extended regions of nucleosomal fibrils in inter-TADs or on the surface of TADs.
A unique feature of ciliated protozoa is the presence of nuclei of two distinct types in the same cell: one or several transcriptionally inert generative diploid micronuclei (MIC) and usually one large transcriptionally active polyploid somatic macronucleus (MAC). MAC develops from zygotic nucleus after conjugation (sexual reproduction) and during its formation undergoes a set of programmed genome rearrangements which involves stages of DNA amplification, restriction, and elimination of internal eliminating sequences. Then, telomeric sequences are added to the ends of the restricted DNA fragments. Thus, the MAC genome is represented by a large number of relatively small minichromosomes (reviews, [36, 37, 38]). In so called “gene-sized” species MAC DNA molecules are mainly 1 to 25 kb in size, whereas in “subchromosomal” species minichromosomes are ~50–1700 kb. To date, we are aware of only one work on ciliates, in which the structure of MIC and MAC chromatin was studied using the Hi-C method [39]. Hi-C analysis of “subchromosomal” ciliate Tetrahymena thermophila showed that MIC chromosomes have TAD-like structures [39]. Each such TAD-like structure develops into a MAC minichromosome during the MAC development. However, the authors revealed no TADs in the MAC minichromosomes [39].
In ultrathin sections of the interphase nuclei of “subchromosomal” ciliates, inactive chromatin usually looks like chromatin bodies with a diameter of ~100–200 nm [37]. Such chromatin bodies can contain either one or several minichromosomes [14, 40]. It has been shown that in hypotonic solutions, chromatin bodies gradually decompact, forming structures consisting of chromatin fibres emerging from electron-dense centres [41, 42, 43].
Each MAC DNA molecule has telomeric regions at both ends. It has been shown that MAC telomeres of individual DNA molecules can interact with each other by G-quartet formation; the formation of this structure is enhanced and stabilized by the telomere-binding protein (TeBP) [44, 45, 46, 47]. It is known that in different cells, from yeast to humans, telomeres are associated with nuclear matrix proteins [48, 49, 50]. Therefore, it can be assumed that the complex of TeBP, nuclear matrix proteins and telomeric ends of MAC DNA are the “organizing centre” of the chromatin body, around which loops of nucleosomal fibrils are organized.
Taken together, this data allow us to assume that MAC chromatin bodies are analogues of the chromomeres of higher eukaryotes. However, as shown above, the chromomeric model differs from the currently widespread TAD model of eukaryotic chromatin organization. The aim of this work was to find out which of the models (chromomeres or TADs) is more consistent with the confocal and electron microscopic data on structural organization of chromatin bodies.
For this purpose, four species of “subchromosomal” ciliates were studied. First, we tested how fixation conditions affect the structure of MAC chromatin. We then studied changes in the structure of chromatin bodies during incubation in solutions with low ionic strength, and then changes in the structure of chromatin at stages of decreased MAC activity, namely during starvation and at the stage of resting cysts. Finally, we determined the distribution of RNPs in the MACs of the studied ciliate species. We present evidence supporting a model in which the chromatin bodies of ciliate macronuclei combine features of both chromomeres and TADs.
The ciliates Bursaria truncatella were caught in the Tsitsin’s Botanical Garden of the Russian Academy of Sciences. Paramecium multimicronucleatum, Didinium nasutum and Climacostomum virens were from the Resource Centre for Culture Collection of Microorganisms (RC CCM, St. Petersburg University).
P. multimicronucleatum were cultivated in a lettuce medium according to standard protocols [51]. B. truncatella, C. virens and D. nasutum were grown in boiled tap water at 10 °C (B. truncatella) and at room temperature, and fed with Paramecium caudatum cultivated separately. To decrease the activity of MACs during starvation, D. nasutum and P. multimicronucleatum cells were transferred into a food-free medium, and preparations of “starved” cells were obtained 48 h (D. nasutum) or 72 h (P. multimicronucleatum) after the remains of food became undetectable within the ciliate cells. Cyst formation of B. truncatella and C. virens was induced by several days of starvation. One-month-old B. truncatella cysts and one-year-old C. virens cysts were examined.
The interphase MACs of B.truncatella were manually isolated under stereomicroscope from ciliates in the logarithmic growth phase. The MACs were isolated in the solution containing 0.5% NP-40 (Cat.#I8896, Sigma-Aldrich, St. Louis, MO, USA) in 0.1 mM borate buffer (Cat.#221732, Merck, Darmstadt, Germany), pH 8.7, washed in 0.1 mM borate buffer and incubated for 30 min in a drop of the same buffer. The spread chromatin was centrifuged for 10 min at 3000 g onto a freshly ionized carbon-parlodion supporting film through a layer of 0.1 M sucrose containing 4% paraformaldehyde (Cat.#19200, Electron Microscopy Sciences, Hatfield, PA, USA), 0.1 mM borate buffer, pH 8.7, as described in [52]. Then, the grids were washed in 0.4% Kodak Photo-Flo solution (Cat.#5038278, Kodak-Pathé, Chalon-sur-Saône, France), pH 7.8, air-dried and rotary shadowed with palladium-platinum alloy.
For ultrathin sectioning of isolated MACs with partially decompacted chromatin bodies, MACs were isolated in the solution containing 0.1% NP-40, 1 mM MgCl2 (Cat.#M9272, Sigma-Aldrich), 2 mM K2HPO4 (Cat.#P3786, Merck) pH 7.8, incubated in hypotonic buffer consisting of 0.01 mM MgCl2, 2 mM K2HPO4 pH 7.8 for 3 min and fixed.
For ultrathin sectioning, the whole ciliates or isolated MACs were fixed either with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.3) or with 2.5% glutaraldehyde (Cat.#16220, Electron Microscopy Sciences) in the same buffer for 1 h at room temperature. Interphase ciliates and cysts of C.virens were fixed for 1 h on ice in the darkness with freshly prepared 2% glutaraldehyde and 1% osmium tetroxide (Cat.#19150, Electron Microscopy Sciences) in 0.1 M phosphate buffer (pH 7.3). The fixed specimens were dehydrated in a graded series of alcohol and embedded in Epon-Araldite according to standard protocol. The ultrathin sections were cut using the LKB III ultratome (LKB Products, Stockholm, Sweden) and stained with uranyl acetate and lead citrate using the standard procedure. The specimens were examined in a JEM-100CX electron microscope (JEOL Ltd., Tokyo, Japan). To selectively contrast ribonucleoproteins (RNP), we used a differential staining of sections with uranyl acetate (Cat.#22400, Electron Microscopy Sciences) – EDTA (Cat.#E9884, Sigma-Aldrich) – lead citrate (Cat.#17810, Electron Microscopy Sciences) according to Bernhard’s technique [53].
The size of the chromatin bodies was measured on scanned negatives with ImageJ software (http://rsbweb.nih.gov/ij).
Ciliates were fixed in 4% paraformaldehyde in 0.1M phosphate buffer pH 7.3, and mounted in “Slowfade Gold” medium (Cat.#S36936, Thermo Fisher Scientific, Waltham, MA, USA) containing 1 µg/mL 4′,6-diamidino-2-phenylindole (DAPI) (Cat.#D9542, Sigma-Aldrich). Specimens were studied in a Leica TCS SP5 confocal microscope (Leica Microsystems, Mannheim, Germany). Fluorescence was excited by a 405 nm laser, detection range was 416–480 nm.
In this work, we used a combination of four evolutionarily distant
“subchromosomal” ciliate species (Bursaria truncatella, Climacostomum
virens, Paramecium multimicronucleatum and Didinium nasutum), that were
large enough to allow convenient sample preparation for electron microscopy and
confocal studies. First of all we compared the morphology of MAC chromatin in the
cells fixed with 4% formaldehyde (i.e., under conditions used for crosslinking
at the first step of any 3C method) and 2.5% glutaraldehyde. Fig. 1A and Fig. 2A show that chromatin bodies are clearly visible in a confocal
microscope in B. truncatella and in P. multimicronucleatum
cells fixed by formaldehyde. There were also no significant morphological
differences in MAC morphology between ultrathin sections of cells fixed with
formaldehyde and glutaraldehyde (Fig. 1B,C and Fig. 2B,C). For example, the size
and circularity of chromatin bodies of B. truncatella were 177
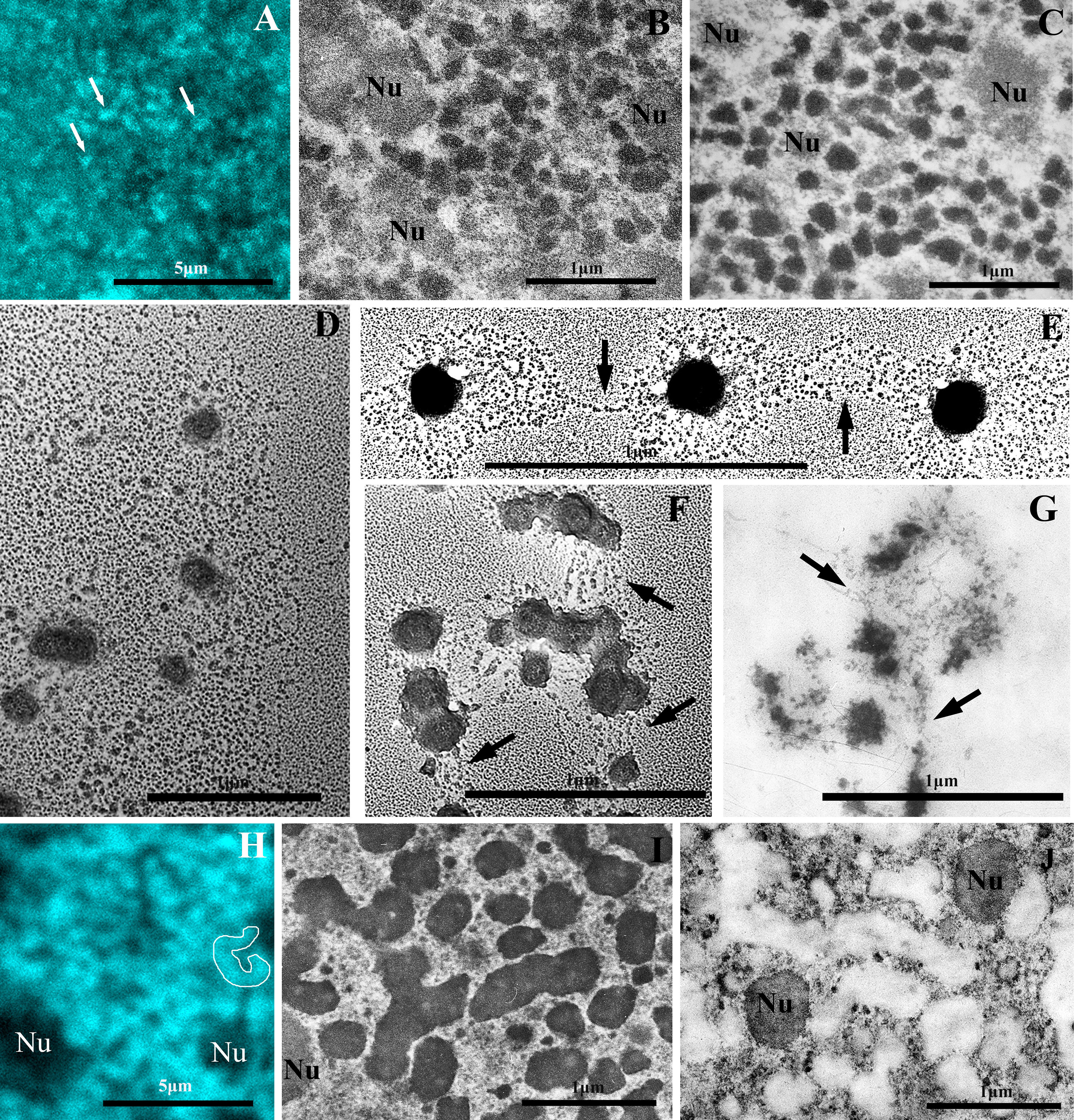 Fig. 1.
Fig. 1.
Electron and confocal microscopy of chromatin structures of the macronucleus (MAC) of the ciliate B. truncatella. (A) Confocal microscopy of the chromatin bodies (white arrows) in the MAC of interphase cell fixed with 4% formaldehyde and stained with DAPI. (B,C) The chromatin bodies 70–250 nm in size in the ultrathin sections of MACs of interphase B. truncatella cells fixed with 4% paraformaldehyde (B) and 2.5% glutaraldehyde (C). (D–F) Isolated MAC chromatin after incubation in 0.1 mM borate buffer for 30 min. (D) An aggregate of partially decompacted chromatin bodies. (E,F) Partially decompacted bodies interconnected with one or more nucleosomal fibrils (arrows). (G) Partially decompacted chromatin bodies in an ultrathin section of isolated MAC fixed with 2.5% glutaraldehyde. Arrows point to nucleosome fibrils connecting chromatin bodies. (H,I) The chromonema-like fibrils 200–300 nm in diameter in the MAC of a one-month-old B. truncatella cyst. (H) Confocal microscopy of a cyst fixed with 4% formaldehyde and stained with DAPI. For clarity, one fragment of the chromonema-like fibril is outlined. (I) An ultrathin section of the MAC of the cyst fixed with 2.5% glutaraldehyde. (J) The same specimen as in (I) after differential staining of nucleic acids according to Bernhard’s technique. RNP structures are contrasted. Nu, nucleolus. Scale bar, 5 µm (A,H); 1 µm (B–G,I,J). DAPI, 4′,6-diamidino-2-phenylindole; RNP, ribonucleoprotein.
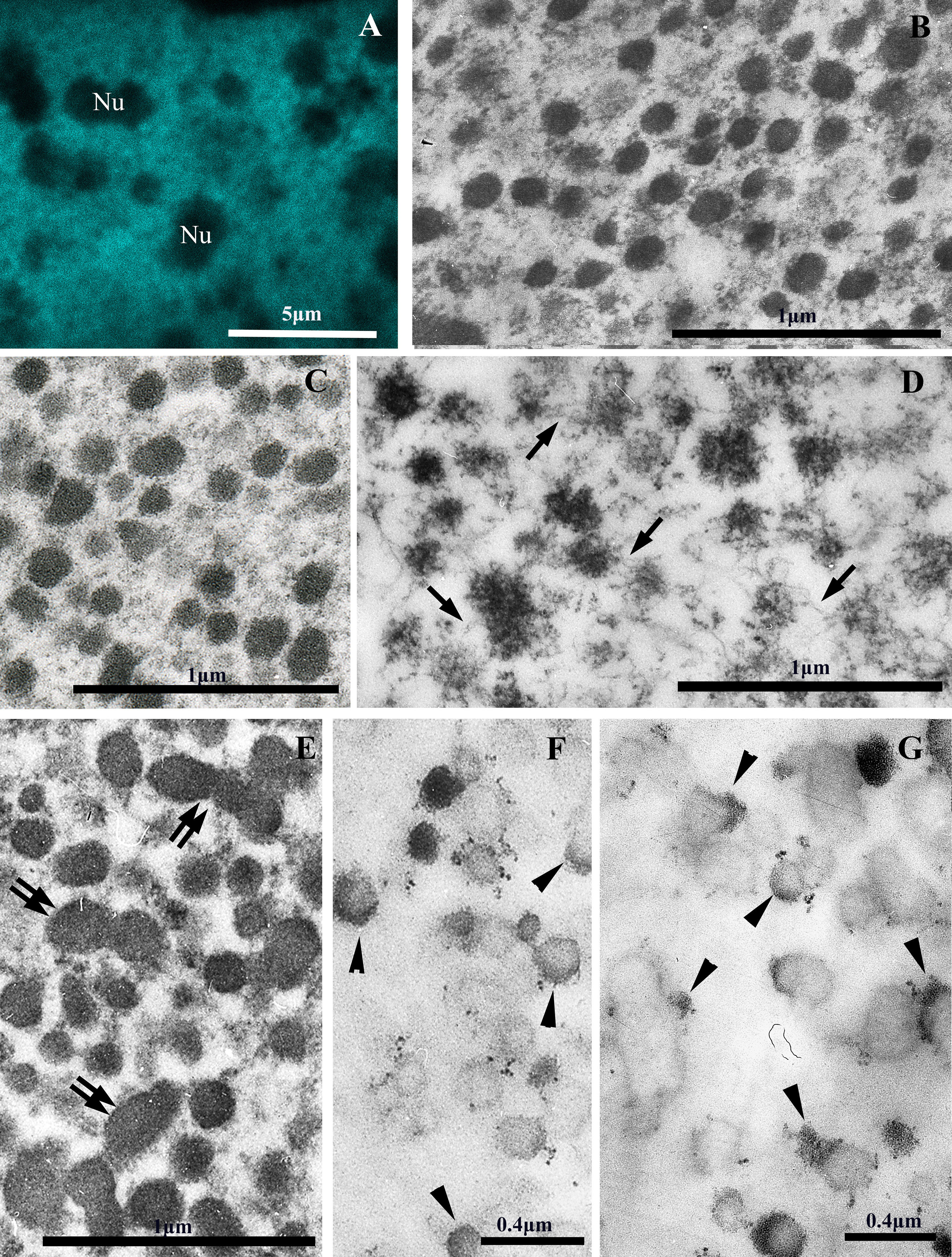 Fig. 2.
Fig. 2.
Electron and confocal microscopy of chromatin structures of the MAC of the ciliate P. multimicronucleatum. (A) Confocal microscopy of the chromatin bodies in the MAC of interphase cell fixed with 4% formaldehyde and stained with DAPI. (B,C) The chromatin bodies 70–250 nm in size are observed in the ultrathin sections of MACs of interphase P. multimicronucleatum cells fixed with 4% paraformaldehyde (B) and 2.5% glutaraldehyde (C). (D) An ultrathin section of isolated MAC with partially decompacted chromatin bodies fixed with 2.5% glutaraldehyde. Arrows indicate nucleosomal fibrils connecting chromatin bodies. (E) An ultrathin section of the MAC of P. multimicronucleatum after starvation for 72 h fixed with 2.5% glutaraldehyde. Chromatin bodies increase in size and form aggregates (double arrows) during starvation. (F,G) The same preparations as in (C) and (E) after differential staining of RNP-containing structures according to Bernhard’s technique. Arrowheads point to RNP structures on the surfaces of single chromatin bodies and aggregates of chromatin bodies. Nu, nucleolus. Scale bar, 5 µm (A); 1 µm (B–E), 0.4 µm (F,G).
At the interphase stage, in all four studied species, the minichromosomes of the MAC were packed into compact, more or less rounded bodies ranging in size from 70 to 250 nm with an average value of 130–180 nm (Fig. 1C, Fig. 2C).
When incubated in a hypotonic solution (0.1 mM borate buffer, pH 8.7–9.0), chromatin bodies gradually decondense to form a halo of nucleosomal fibrils (Fig. 1D,E). In ultrathin sections, such partially decompacted bodies look like “rosette-like” structures (Fig. 1G, Fig. 2D).
Both on preparations of isolated chromatin and in ultrathin sections of MACs, it is visible that some chromatin bodies are interconnected by one or more chromatin fibrils (Fig. 1E–G, Fig. 2D). In addition, decondensation of the bodies occurs with the formation of more or less symmetrical halos (Fig. 1E), which resemble the process of decondensation of fibrils packed according to the principle of the fractal globule [35].
Ciliate MACs divide by amitosis, in which chromatin does not condense into structures similar to the mitotic chromosomes of mammalian cells. Therefore, to study the ability of chromatin bodies to aggregate into higher-level chromatin structures in vivo, we investigated the chromatin structure of MAC under conditions when MAC activity is reduced. For example, this occurs during starvation and encystment (i.e., when cells pass into an inactive state - resting cysts). B. truncatella and C. virens easily form resting cysts under unfavourable conditions.
The morphology of the chromatin structures of MACs changes significantly during encystment. In one-month-old B. truncatella cysts, chromatin bodies aggregate into thick fibrils, clearly visible in a confocal microscope (Fig. 1H). In the ultrathin sections such fibrils 200–300 nm in diameter resemble chromonemata from chromosomes of higher eukaryotes [3, 4, 5]. They are formed by chains of interconnecting chromatin bodies. Their length measured in the sections is up to 1600 nm (Fig. 1I,J).
Even more drastic changes in chromatin morphology were observed between interphase C. virens ciliates and one-year-old cysts of C. virens. In confocal images of the MACs of interphase C. virens ciliates chromatin looks like small granules (Fig. 3A), but in the images of the one-year-old cysts we see thick chromosomal threads in the MAC (Fig. 3C). Electron microscopy shows that MAC chromatin in interphase C. virens cells, similar to other ciliates we studied in this work, is organized in chromatin bodies 70–200 nm in diameter (Fig. 3B). Most of them are located singly (Fig. 3B, single arrows), others form short fibrils of two, three or more rarely of four closely located chromatin bodies (“double” bodies are indicated by double arrows in Fig. 3B).
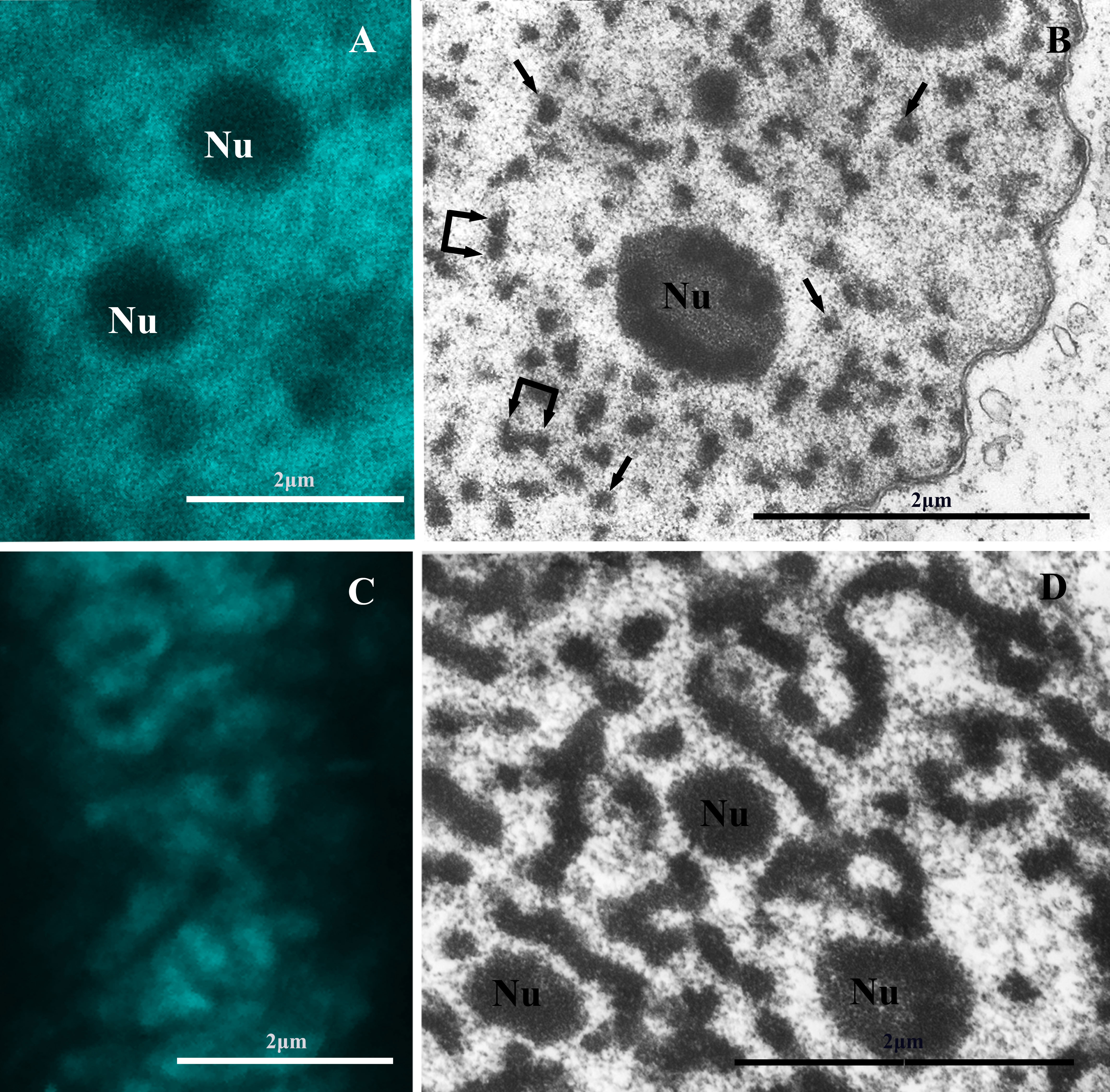 Fig. 3.
Fig. 3.
Electron and confocal microscopy of chromatin structures of the MAC of the ciliate C. virens. (A) Confocal microscopy of the chromatin bodies in the MAC of interphase cell fixed with 4% formaldehyde and stained with DAPI. (B) An ultrathin section of an interphase C. virens MAC. Single arrows point to individual chromatin bodies, double arrows point to two closely located (“double”) bodies. (C) Confocal microscopy of a one-year-old C. virens cyst shows the presence of thick fibrils in the MAC. (D) Chromonema-like fibrils in an ultrathin section of a one-year-old C. virens cyst. Nu, nucleolus. Scale bars, 2 µm.
An ultrathin section of a one-year-old C. virens cyst is shown in Fig. 3D. Long branched chromonema-like fibres about 200 nm thick form an extensive network throughout the entire volume of the MAC. Such fibrils morphologically look like chromonemata observed in the chromosomes of higher eukaryotes [3, 18].
Some ciliate species, e.g., Paramecium, do not have the stage of
resting cysts. However, it is known that the activity of the MAC decreases when
ciliates are cultivated under starvation conditions. This leads to changes in the
morphology of the chromatin structures of the MAC. Fig. 4A,B show confocal
and electron microscopy of MAC chromatin from the ciliate D. nasutum.
The size of chromatin bodies, measured in ultrathin sections, ranged from 70 to
200 nm. Fig. 2E, Fig. 4C show micrographs of chromatin bodies in the
MACs of P. multimicronucleatum and D. nasutum after starvation
for 72 hours and 48 hours, respectively. The mean size of the chromatin bodies of
P. multimicronucleatum increases from 139
 Fig. 4.
Fig. 4.
Electron and confocal microscopy of chromatin structures of the MAC of the ciliate D. nasutum. (A) Confocal microscopy of the chromatin bodies in the MAC of an interphase cell fixed with 4% formaldehyde and stained with DAPI. (B) The MAC chromatin of interphase D. nasutum is mainly organized into chromatin bodies 70–250 nm in size. (C) Chromatin bodies form short fibrils (double arrows) in the MAC of the ciliate D. nasutum after a 48-hour fast. (D,E) The same preparations as in (B) and (C) after differential staining of RNP-containing structures according to Bernhard’s method. Arrowheads point to RNP structures on the surfaces of chromatin bodies and fibrils. Nu, nucleolus. Scale bar, 2 µm (A); 1 µm (B–E).
It should be noted that transcriptional activity in such chromatin structures seems to be preserved. Fig. 2F,G and Fig. 4D,E show the sections of Paramecium and D. nasutum MACs under optimal cultivation conditions and under starvation conditions, contrasted according to the Bernhard method [53]. In such specimens, RNP structures are preferentially contrasted. It is clearly seen that RNP structures are detected on the surface of chromatin bodies, between them and on the surface of 200–300 nm fibrils, too. Notably, transcription does not occur uniformly over the entire surface of bodies and fibrils, but in individual loci (Fig. 2F,G, Fig. 4D,E).
Our data confirms that chromatin bodies 70–250 nm in size are the typical form of inactive chromatin organization in the MACs of “subchromosomal” ciliate species. Chromatin bodies were observed both in confocal and electron microscopes in the cells fixed with formaldehyde (typical fixation conditions for all 3C methods) and with glutaraldehyde (standard for electron microscopy). Thus, the observed structures are not artifacts of fixation and are not a consequence of the presence of divalent cations in the medium.
Such chromatin bodies decompact in a low ionic strength buffer, forming
rosette-like structures consisting of a “halo” of nucleosomal fibrils around
electron-dense centres. It is known from the literature that telomeres of
individual MAC minichromosomes can interact with each other by G-quartet
formation [54] and with proteins of the nuclear matrix; the formation of these
structures is enhanced and stabilized by the TeBP [44, 45, 46, 47]. It was shown that in
Oxytricha and Stylonychia, the TeBP is a heterodimeric protein
consisting of two (
When MAC activity decreases after starvation or during encystment, chromatin bodies aggregate to form thick chromonema-like fibrils resembling the morphology of higher eukaryotic chromonemata. We use the term “chromonema-like fibril” instead of “chromonema” because each higher eukaryotic chromonema is formed from a single long continuous nucleosomal fibril rather than from a large number of subchromosomal minichromosomes. In addition, chromonema-like fibrils have a branched rather than linear structure (Fig. 1J, Fig. 3D).
Thus, our data supports the presence of discrete levels of chromatin organization in the MAC of subchromosomal ciliates: nucleosomal fibres – chromatin bodies – chromonema-like fibrils. The above-described properties of chromatin bodies correspond well to the properties of the chromomeres of higher eukaryotes. However, unlike chromomeres and chromonema of higher eukaryotes, chromatin bodies and chromonema-like fibrils are detected in MACs by electron microscopy without additional procedures of decompactization or deproteinization of preparations. This difference is apparently due to the fact that chromatin bodies and chromonema-like fibrils are formed in MACs of “subchromosomal” ciliates by compaction of relatively short (~50–1700 kb) DNA molecules.
In order to explain the obtained microscopic data, we propose a hypothetical scheme of chromatin bodies organization and formation of branched chromonema-like fibrils in the MACs of ciliates (Fig. 5). In this scheme, chromatin bodies are formed by loops of nucleosomal fibrils around the “organizing centres” according to the principle of a fractal globule, as in TADs. In addition, the scheme takes into account that chromatin bodies can be formed by several minichromosomes, and some of them can be connected by one or more chromatin fibrils.
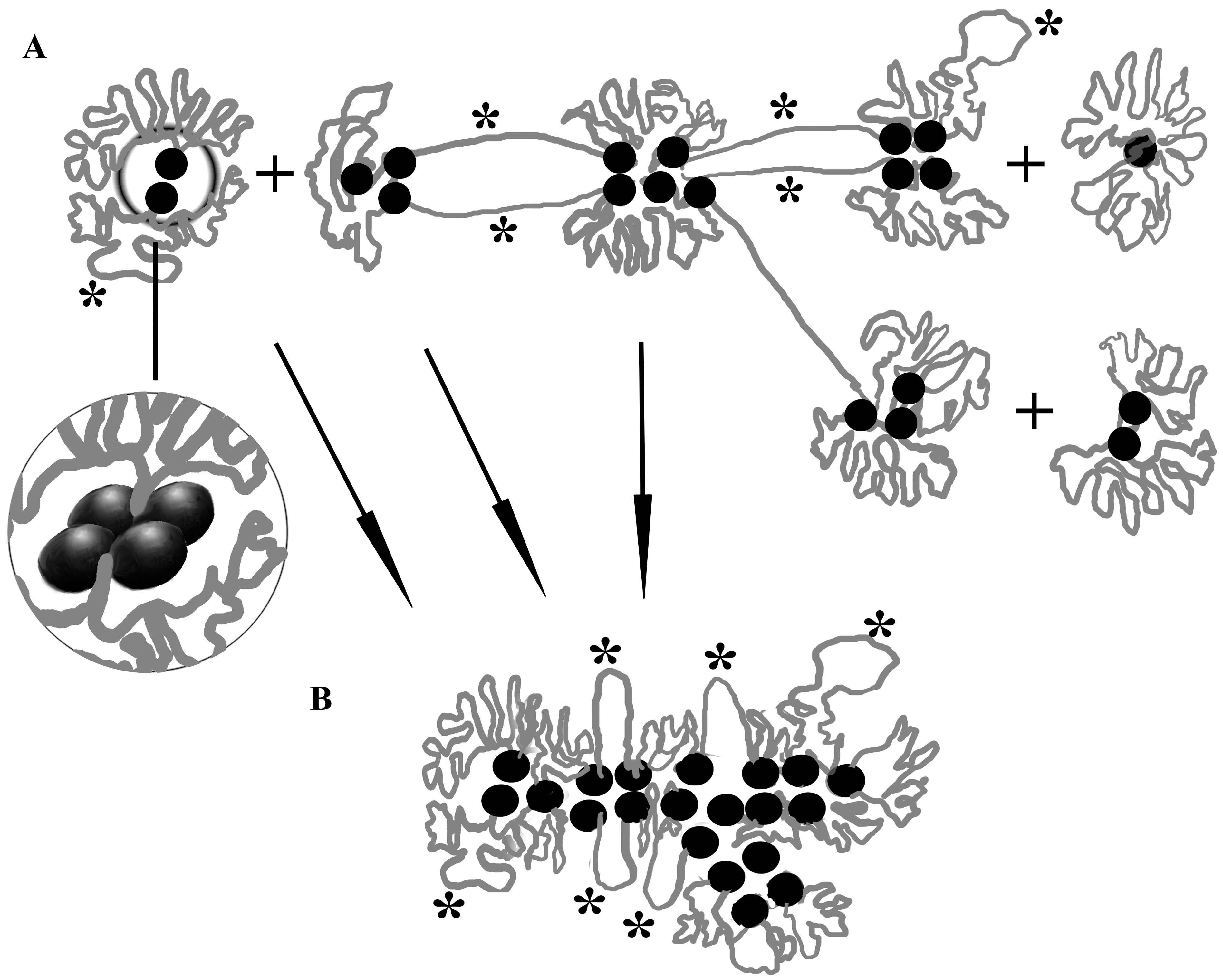 Fig. 5.
Fig. 5.
A hypothetical scheme of chromatin bodies organization and formation of branched chromonema-like fibrils in the MACs of ciliates. (A) Chromatin bodies are formed by loops of nucleosomal fibrils around the “organizing centres” (black circles). Loops in chromatin bodies are organized according to the principle of a fractal globule, as in topologically associating domains (TADs). Asterisks indicate loops and unfolded regions of chromatin where transcription occurs. In the insert circle: complex of four telomeric ends of two minichromosomes and TeBPs in an “organizing centre”. (B) An aggregation of the “organizing centres” of chromatin bodies leads to the formation of chromonema-like branched fibril 200–300 nm in diameter. Transcription can take place on the surface of such fibrils (asterisks).
If the “organizing centres” of chromatin bodies are more adhesive to each other than the nucleosomal fibrils, then first the aggregation of the “organizing centres” will lead to some enlargement of the bodies, and then, due to steric restrictions, further aggregation will occur only in the longitudinal direction, which leads to the formation of 200–300 nm fibrils, similar to the model of the formation of chromonemata from chromomeres [18]. It looks like a phase separation process involving organizing centres and chromatin fibres connecting adjacent chromatin bodies. Branched areas of chromonemata arise where any chromatin body is connected to more than two neighbouring ones. It is noteworthy that transcription processes can also occur on the surface of chromonema-like fibrils. It is currently unclear whether the “organizing centres” are retained as distinct structures in chromonema-like fibrils. This issue requires further study.
This scheme (Fig. 5) also allows us to make some predictions. If chromatin bodies are formed from long DNA molecules and are not interconnected with each other, the starvation or encystment will lead to an increase of the size of chromatin bodies, rather than the formation of chromonema-like fibrils. On the contrary, in the species with short MAC DNAs chromonema-like fibrils should be more common. These predictions will be tested in further studies.
An important question is how the loops of chromatin fibrils are organized in chromatin bodies. Calculations show that minichromosomes with a total size of ~50 kb to ~2300 kb can be packaged in chromatin bodies with a diameter of 70 to 250 nm [14]. Taking into account the fact that the size of DNA loops in the chromatin of higher eukaryotes is 100–200 kb. [23, 24, 25], each chromatin body contains from one to 10–20 DNA loops. According to the “classical” scheme of chromomere organization, such loops should be attached to the core complexes of chromomeres. However, at present there is no confirmed data on the content of sequences homologous to telomeric fragments in the central part of the MAC DNA molecules. Moreover, with such an organization, the transcription of genes located in the central part of the bodies would be difficult.
It is most likely that the loops in chromatin bodies are organized according to the principle of a fractal globule, as in TADs. Such an organization makes it possible to dynamically expose specific parts of DNA molecules for transcription on the surface of the chromatin bodies. In this case, transcription can occur, not only on the unfolded sections of nucleosomal fibrils, but also on the surface of chromatin bodies and chromonema-like fibrils without decompaction of the latter (Fig. 5, asterisks). This is precisely the picture we observed in the ultrathin sections of ciliate MACs after differential staining of nucleic acids: RNPs were detected only in some areas on the surface of chromatin bodies (Fig. 1J, Fig. 2F,G, Fig. 4D,E). Similar data were reported by Jeter et al. [56], who observed rounded chromatin bodies with rather smooth, well defined boundaries in MACs of starved T. pyriformis ciliates. Following refeeding and as the biological activity of the cells increased, the shape of the chromatin bodies became more irregular and appeared to radiate more fibrous material, but no complete decompaction of chromatin bodes was observed [56].
Currently, the most experimentally substantiated model suggests that cohesin and CTCF are involved in the formation of TADs in higher eukaryotes. However, CTCF was not found in T. thermophila. The cohesin complex was only located in the MIC but not in the MAC of T. thermophila [57]. However, it was assumed that the mechanisms of the formation of TADs are actually much more universal than it appears at present [58]. For example, a super resolution chromatin tracing technique showed that cohesin is not required for maintaining the TAD-like structure in single cells [29]. It was also shown that the genome of danio fish contains neither CTCF nor the cohesin complex at most TAD boundaries [59], despite the fact that CTCF in danio and humans shows 86% homology. In addition, it was shown in Drosophila that other proteins of the C2H2 proteins family may play the architectural role in chromatin organization instead of CTCF [58].
The data obtained is well explained by the model, according to which the chromatin bodies of ciliate MACs are structures in which the principles of organization inherent in both chromomeres and TADs are effectively implemented; that is, they can be considered as chromomeres with loops organized according to the principle of TADs, or as TADs with elements of chromomeric organization.
TAD, topologically associating domain; MAC, macronucleus; MIC, micronucleus; TeBP, telomere-binding protein; CTCF, CCCTC-binding factor; DAPI, 4′,6-diamidino-2-phenylindole; RNP, ribonucleoprotein.
All data points generated or analyzed during this study are included in this article and there are no further underlying data necessary to reproduce the results.
VPo and VPr designed the research study. VPo and OL cultivated cells, VPo, OL and PS performed the research. VPo and OL wrote the draft of the manuscript. VPo, PS, VPr and OL participated in the revision of draft and the interpretation of data for the work. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
According to the policy of the Engelhardt Institute of Molecular Biology, Russian Academy of Sciences, ethical approval is not required.
We thank Dr. V.A. Grigoryev for technical maintenance of the electron microscope. We thank Dr. N.A. Lebedeva for providing P. multimicronucleatum, D. nasutum and C. virens cultures. We thank Dr. B.P. Karajan for helpful discussions of our results.
The experiments with electron microscopy were supported by the Russian Science Foundation by grant 22-14-00353. The experiments with confocal microscopy were supported by the Grant 075-15-2019-1660 from the Ministry of Science and Higher Education of the Russian Federation.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.