

 , Shuyang Zhang 1,2,*
, Shuyang Zhang 1,2,*1 Department of Cardiology, Peking Union Medical College Hospital, Chinese Academy of Medical Sciences & Peking Union Medical College, 100730 Beijing, China
2 State Key Laboratory of Complex Severe and Rare Diseases, Peking Union Medical College Hospital, Chinese Academy of Medical Sciences & Peking Union Medical College, 100730 Beijing, China
†These authors contributed equally.
Abstract
Coronary thrombosis events continue to be the leading cause of morbidity and mortality worldwide. Recently, emerging evidence has highlighted the role of gut microbiota in cardiovascular disease, but few studies have systematically investigated the gut microbiota variation associated with atherothrombosis.
We conducted multi-omics analysis (metagenomics sequencing and serum metabolomics) on 146 subjects from Peking Union Medical College Hospital-Coronary Artery Disease (PUMCH-CAD) cohort. We analyzed the key strains and metabolic pathways related to coronary artery disease (CAD) development, explored the bacterial functional pathway which contributes to atherothrombosis at strain level in depth. Single strain colonization procedures on germ free mice demonstrated the promotion of platelet activation and thrombotic phenotypes of the disordered gut microbiota.
Gut microbiome and serum metabolome shifts were apparent in cases of CAD progression, Bacteroides spp. disturbed the development of CAD by participating in lipopolysaccharide (LPS), menaquinone and methanogenesis pathways. Particularly, coronary thrombosis is characterized by increased circulatory levels of L-tryptophan, which correlate with Bacteroides thetaiotaomicron that has enriched biosynthetic potential. In germ free mice we demonstrate that Bacteroides thetaiotaomicron colonization could induce thrombosis, aggravate platelet hyperreactivity and augment fecal levels of L-tryptophan.
The disordered gut microbiota of CAD contributed to the occurrence and development of atherothrombosis. The key members of the bacterial and metabolic features may become biomarkers for predicting the cardiovascular thrombosis event. Targeting the microbial pathway may have the potential to reduce the incidence of cardiovascular disorders.
ChiCTR2000033897, https://www.chictr.org.cn/showproj.html?proj=55023.
Keywords
- gut microbiota
- coronary artery disease
- myocardial infarction
- atherothrombosis
- Bacteroides thetaiotaomicron
- L-tryptophan
Coronary artery disease (CAD), a major burden of cardiovascular diseases (CVDs), is a chronic inflammatory pathophysiological process of the artery arising at sites of disturbed blood flow. CAD is attributed to atherosclerosis progression where thrombus formation may cause fatal complications such as myocardial infarction (MI) [1]. Despite timely reperfusion by primary percutaneous coronary intervention, MI is still the main cause of death worldwide [2]. Many researchers have attempted to find novel targets to predict and prevent this critical complication. Since Wang et al. [3, 4] reported a metabolite derived from gut microbiota, trimethylamine N-oxide (TMAO), was predictive of CVD events. Gut microbiota has been implicated as a novel ‘endocrine organ’ that plays an important role in CVDs. Thus, further studies are warranted to increase understanding of pathophysiological mechanisms between gut microbiota and CAD, in order to shed light on novel risk biomarkers and interventions.
Many researchers have suggested that commensal microbiota functioned as an environmental factor contributing to metabolic disease [5], atherosclerotic lesions [3, 6] and arterial thrombosis [7, 8, 9]. For instance, Kelly et al. [10] analyzed gut microbiota from patients suffering from heart disorders and found Alloprevotella, Prevotella, and Paraprevotella were linked to increasing CVD risk. In another metagenomics study on patients diagnosed with symptomatic atherosclerotic plaques, Collinsella was significantly enriched, whereas Eubacterium and Roseburia were more abundant in healthy volunteers [11]. Our previous study proved that the bacterial co-abundance group (CAG) in the different stages of CAD were dominated by Roseburia, Clostridium IV, Klebsiella and Ruminococcaceae spp. [12]. However, the current model cannot effectively differentiate between stable CAD and an MI event, which limits its broad utilization in clinical settings. Therefore, it remains controversial how gut microbiota influences the rupture of atherosclerotic plaque with subsequent thrombus events.
Gut microbiota is responsible for a number of factors related to platelet function and thrombosis, including serotonin [13], vitamin K [14] and von Willebrand factor [15]. Additionally, TMAO can alter calcium signaling in platelets, enhancing platelet reactivity and thrombotic potential [7]. Since gut microbiota are the major source of physiologically pattern-recognition receptors (PRR) agonists, it is crucial to explore the mechanism of thrombogenesis in the microbial ecosystem [16]. Bacterial irritant molecules from microbiota can affect immune vigilance, thereby increasing the risk of thrombosis. By combining metagenomics studies with fecal microbiota transplantation experiments, it can provide relevant microbial-relatedevidence for thrombosis prevention.
To overcome these challenges, we launched interplay analysis of gut microbiome, serum metabolome and clinical indicators in a population of 146 subjects from Peking Union Medical College Hospital CAD cohort (PUMCH-CAD, ChiCTR2000033897, https://www.chictr.org.cn/showproj.html?proj=55023). We characterized multi-omics associations with dominant clinical CAD phenotypes including atherosclerosis burden and thrombotic event type. Especially Bacteroides enriched in CAD populations showed a strong correlation with thrombotic events by interfering with L-tryptophan and methanogenesis pathways. Last, we evaluated the ability of microbial signatures including Bacteroides thetaiotaomicron (B.t) to modulate platelet function.
This is a cross-sectional study, where recruited patients were individuals who
had been hospitalized for coronary angiography in our center, as previously
described [12]. This study was approved by the local Medical Ethics Committee of
Peking Union Medical College Hospital (PUMCH) (JS-1195). Prior to their participation in the study, all subjects provided
written informed consent. CAD definition:
The burden of coronary atherosclerosis was estimated using the Gensini and
Syntax scores, as previously reported [12]. Blood pressure was measured with a
standard mercury sphygmomanometer when the patient was in a seated position after
Continuous, normal distributed variables were expressed as means
We provided a stool sampler and detailed instructions for sample collection. Freshly collected stool samples were immediately frozen at –80 °C. DNA extraction was performed using the Qiagen QIAamp DNA Stool Mini Kit (51504, Qiagen, Hilden, Germany) according to the manufacturer’s instructions. DNA quantity was determined using a NanoDrop spectrophotometer, Qubit Fluorometer (with the Quant-iT™dsDNA BR Assay Kit, Q33130, Eugene, OR, USA), and gel electrophoresis.
DNA library construction was mainly performed following the manufacturer’s
instruction (Illumina, San Diego, CA, USA). We constructed one paired-end (PE) library with an insert
size of 350 bp, followed by high-throughput sequencing with PE reads of length 2
Employing the same parameters to construct the Metagenomics of the Human Intestinal Tract (MetaHIT) gene catalogue [20], we performed gene prediction using SOAPdenovo v1.06 (Beijing Genomics Institute, Hongkong, China) [21] and GeneMark v2.7 (Georgia Institute of Technology, Atlanta, GA, USA) [22]. Genes were aligned using BLAT (BLAST-like alignment tool, Kent Informatics, Inc, Mountain View, CA, USA) and genes with over 90% of their length aligned to another with more than 95% identity were removed as redundancies.
We performed taxonomic assignment using an in-house pipeline, we collected the microbial reference genomes from the Integrated Microbial Genomes (IMG) database to make alignment. We used 85% identity as the threshold for genus assignment, and threshold of 80% for the alignment coverage based on sequence similarity across phylogenetic ranks by MetaHIT [23]. For genes, the highest scoring hit(s) above these two thresholds were chosen for the genus assignment.
We calculated
Differentially enriched KEGG (KEGG database release 59.0, genes from animals and plants removed) pathways were identified according to their reporter score as follows. Wilcoxon rank-sum tests were performed on all KOs of our samples and adjusted for multiple testing using the Benjamin–Hochberg procedure. An absolute reporter score value higher than 1.6 (95% confidence on either tail, according to normal distribution) was used as threshold.
For instance, leucine biosynthesis-related KEGG pathway details were obtained
from KEGG database ‘M00432’ (list of KO genes) under the ‘Valine, leucine and
isoleucine biosynthesis’ category. Prevalence of genes higher than 5% were
analyzed for any of the stages (SCAD, MI) compared to the controls. Pathways
shown in our results were manually modified according to KEGG database
or referring to the literature. We showed KO genes with significant differences
between group comparisons (p
Metabolomics analysis was performed on a Waters ACQUITY ultra-high-performance liquid chromatography system (Waters Corporation, Milford, MA, USA) coupled with a Waters Q-TOF Micromass system (Waters Corporation, Milford, MA, USA) as previously described [25, 26]. We used databases such as KEGG, MetaboAnalyst, Human Metabolome Database, and METLIN to identify metabolic pathways. Next, SIMCA-P 14.0 software (Umetrics AB, Umea, Sweden) was implied to acquire metabolite information and variables.
A total of 7061 serum metabolic features were yielded after pre-processing
above. We conducted a “cross-comparison scheme” as follows to downsize
metabolic features associated with disease status. (i)
Multi-comparisons set A: various stages of CAD were compared with healthy control
(HC) and to each other using the Wilcoxon test and T test, the
metabolites will be retained for the next stage of analysis both statistics
results satisfied the adjusted p value (q value)
We cluster metabolites into metabotypes using the R package WGCNA [27] as
previously described. For parameters, scale-free topology threshold was set for
All standards were obtained from Sigma-Aldrich (St. Louis, MO, USA), Steraloids
Inc. (Newport, RI, USA) and TRC Chemicals (Toronto, ON, Canada). All the
standards were accurately weighed and prepared to obtain individual solutions at
a concentration of 5.0 mg/mL. Formic acid was of analytical grade obtained from
Sigma-Aldrich (St. Louis, MO, USA). Methanol (Optima LC-MS, Thermo Fisher Scientific, Waltham, MA, USA), acetonitrile (Optima
LC-MS), and isopropanol (Optima LC-MS, Thermo Fisher Scientific, Waltham, MA, USA) were purchased from Thermo-Fisher
Scientific (FairLawn, NJ, USA). Sample preparation was performed as previously
described [26]. Targeted quantitation was performed to determine the
concentration of metabolites as previously described by reverse-phase UHPLC using
a Prominence 20 UFLCXR system (Shimadzu, Columbia, MD, USA) with a Waters
(Milford, MA, USA) BEH C18 column (2.1
Spearman correlations between functional modules, serum metabolites, and clinical parameters were calculated using R (version 4.3.0), and both differential abundances of AS- or Thrombosis-associated functional modules and metabolites were tested using the Wilcoxon rank sum test. Wherever mentioned, the Benjamini-Hochberg method was used to control the false discovery rate (FDR).
Germ-free (GF) ApoE -/- mice were provided by the Institute of Laboratory Animal Sciences (ILAS) at the Chinese Academy of Medical Sciences and Peking Union Medical College [research license no. SYXK (Beijing) 2015-0035], which is a member of (and accredited by) the American Association for the Accreditation of Laboratory Animal Care. All experiments were performed in accordance with the guidelines of the Institutional Animal Care and Use Committees of the ILAS. The mice were maintained under standard GF conditions with a 12:12-hour light:dark cycle. GF mice were maintained in flexible film isolators and their GF status was checked weekly by aerobic and anaerobic culture. All GF ApoE -/- mice were male. All mice were used after reaching the age of 6 weeks. All GF ApoE -/- mice received sterilized high fat diet (40 kcal% fat and 1.25% w/w cholesterol; product number D12108S) which was purchased from Changzhou SYSE Bio-Tec. Co., Ltd (Changzhou, Jiangsu, China).
Bacteroides thetaiotaomicron (ATCC 29741) and Prevotella copri
(control strain, ATCC 33547) were cultured in tryptic soy agar/broth supplemented
with defibrinated sheep blood (ATCC medium 260) under anaerobic conditions.
Strains were harvested in logarithmic phase, aliquoted using fresh broth and
concentrated to 109 CFU/mL and diluted into 10% glycerol under anaerobic
conditions and stored at –80 °C until use. GF ApoE-/- mice
were kept in individual isolators and received a high-fat diet. In the next three
weeks, the mice were gavaged twice a week, each time with 100 µL of
bacterial solution containing B. thetaiotaomicron or P. copri
(5
Measurement of acute thrombus formation were performed as described earlier [29]. Some mice did not survive meaning there were ultimately 8 mice in the B.t group and 7 mice in the Prevotella copri (P.c) group. After the mice were anesthetized, the left jugular vein and right carotid artery were carefully dissected. The mice were injected intravenously with Rhodamin B to label the platelets. The ferric chloride injury model was performed by placing 7.5% FeCl3 solution for 1 min laterally to the common carotid artery. Then, the resulting thrombus formation and occlusion of the artery was recorded using a high-speed wide-field Leica M205 FCA fluorescence stereo microscope with Leica Application Suite X (LAS X) software (Leica Microsystems GmbH, Wetzlar, Germany). We recorded the occulusion time. The experiment was terminated if the carotid artery did not occlude within 30 minutes.
Mouse platelet-rich plasma (PRP) was prepared as described previously with minor
modifications [30, 31, 32]. Mouse whole blood (
The final platelet suspensions (100 µL PRP dilute to 700
µL with HEPES Tyrode’s buffer (pH 7.4); 2
For the platelet spreading functional study, we largely followed procedures as
previously described [31]. Microfluidic shear flow experiments were performed
using the rectangular flow chamber assay (31-010, Glycotech, Rockville, MD, USA) equipped with single
channel syringe pump NE-1000 and flexcell VP750 vacuum pump. Briefly, cover
glasses were degreased (24 by 60 mm) by incubating overnight in undiluted
chromosulfuric acid. Then the glass coverslips were coated with collagen (100
µg/mL; C7661-10MG, Sigma, St. Louis, MO, USA) and fibrinogen (100 µg/mL;
F3879, Sigma). The coated coverslips were blocked with Tyrode’s HEPES buffer
containing 1% bovine serum albumin (BSA). The rested and washed platelets were stained by FITC labeled
anti-CD41Mab (1:100; 133903, Biolegend) in the dark at room temperature for 30
min. Unbound antibodies were washed away and prostacyclin was added at a final
concentration of 10 ng/mL to prevent platelet activation. The platelet pellet was
reconstituted in HEPES Tyrode buffer to achieve a platelet concentration of
150,000 platelets/µL. After the incubation, platelet samples were
perfused over chips coated with or without agonist at a physiological shear rate
(7.5 µL/min) using a microfluidic device for 20 min. Immediately
after performing the flow experiments, samples were fixed for 15 minutes in 4%
paraformaldehyde in PBS at room temperature in the dark. Microscopic images were
acquired using a Nikon (Tokyo, Japan) confocal microscope with a 60
To explore the role of gut microbiota in mediating cardiac events, we conducted
a prospective cohort study of patients with CAD at PUMCH, China. In total, 36 control subjects and 110 CAD patients who
underwent coronary arteriography were included, CAD patients were divided into a
SCAD subgroup (N = 64) and an MI subgroup who were suffering from a coronary
thrombosis event (MI, N = 46). The inclusion and exclusion criteria were
previously summarized in methods. The MI subgroup showed a more severe
atherosclerosis burden than the SCAD group in terms of the Gensini score and the
Syntax score (evaluate severity of plaque burden under coronary angiography). The
number of stenosed vessels was also significantly increased (Gensini score,
p
| Group | HC (N = 36) | SCAD (N = 64) | MI (N = 46) | p value | |
| Gender (Male, %)§ | 22 (61.1%) | 51 (79.7%) | 36 (78.3%) | 0.097a | |
| Age, years* | 58.24 |
60.86 |
61.28 |
0.177 | |
| SBP, mmHg* | 125 |
129.8 |
128.9 |
0.161 | |
| BMI, kg/m2* | 24.6 |
26.6 |
26.1 |
0.005ab | |
| Current smoker§ | 14 (38.9%) | 39 (60.9%) | 26 (56.5%) | 0.097a | |
| Gensini score† | NA | 40.8 (33.1, 57.8) | 74.5 (56.6, 93) | ||
| Syntax score† | NA | 9 (5.3, 13) | 17 (10.8, 23.1) | ||
| No. of stenosed vessels§ | 0.089 | ||||
| 1 | NA | 18 (38.1%) | 1 (2.2%) | ||
| 2 | NA | 21 (32.8%) | 10 (21.7%) | ||
| 3 | NA | 25 (39%) | 35 (76.1%) | ||
| NYHA class§ | 0.13 | ||||
| 1 | 36 (100%) | 43 (67.2%) | 33 (71.7%) | ||
| 2 | 0 | 18 (28.1%) | 12 (26.1%) | ||
| 3 | 0 | 3 (4.7%) | 1 (2.2%) | ||
| TIMI risk score | 6.8 (3.5, 9.9) | 9.9 (6.8, 14.5) | 9.9 (6.8, 14.5) | ||
| Laboratory data | |||||
| WBC |
6.47 |
7.07 |
7.13 |
0.209b | |
| ALT, U/L† | 19.5 (15, 28.5) | 23.5 (19.3, 33.8) | 27.5 (20, 37) | 0.004ab | |
| LDL-C, mmol/L† | 2.65 (1.9, 3.3) | 1.9 (1.6, 2.5) | 2.3 (1.9, 2.9) | ||
| Hs-CRP, mg/mL† | 1.03 (0.42, 2.3) | 1.3 (0.5, 2.6) | 2.7 (1.1, 6.3) | 0.001bc | |
| FBG, mmol/L† | 6 (5.2, 7.1) | 6.2 (5.3, 8.2) | 7 (5.9, 7.9) | 0.059b | |
| CK-MB, µ/L† | 0.5 (0.4, 0.85) | 0.7 (0.5, 1) | 0.75 (0.5, 1.3) | 0.028ab | |
| cTnI, µg/L† | 0 | 0.00 (0, 0.01) | 0.28 (0.08, 0.78) | 0.015ac | |
| Cr, umol/L† | 73.5 (66, 85.3) | 79 (69.3, 89.5) | 81.5 (70, 91) | 0.033ab | |
| eGFR, mL/min/1.73 m2† | 94.7 (83.2, 111.7) | 91.7 (82.7, 105.2) | 89.2 (81.6, 111.9) | 0.533 | |
| PCI history§ | 0 | 45 (70.3%) | 36 (56.5%) | ||
| Stroke§ | 0 | 2 (3.1%) | 6 (13%) | 0.020a | |
| Diabetes mellitus§ | 5 (13.9%) | 18 (28.1%) | 17 (37%) | 0.066b | |
| HTN§ | 7 (19.4%) | 41 (64%) | 34 (73.9%) | ||
| PAD§ | 1 (2.8%) | 20 (31.3%) | 13 (28.3%) | 0.003ab | |
| Drugs | |||||
| Aspirin§ | 0 | 60 (93.8%) | 46 (100%) | ||
| Clopidogrel§ | 0 | 49 (76.6%) | 38 (82.6%) | ||
| 4 (11.1%) | 49 (76.6%) | 40 (87%) | |||
| ACEI/ARB§ | 5 (13.9%) | 43 (67.2%) | 23 (50%) | ||
| Statin§ | 1 (2.8%) | 60 (93.8%) | 40 (87%) | ||
| Metformin§ | 2 (5.6%) | 11 (17.2%) | 7 (15.2%) | 0.251 | |
| PPI§ | 2 (5.6%) | 10 (15.6%) | 7 (15.2%) | 0.309 | |
†median (IQR), *mean
Continuous and distributed variables were analyzed by one-way analysis of
variance. The Kruskale Wallis H-test was applied for data which were not normally
distributed. Student’s t-test were used to analyse continuous, normally
distributed data. Mann-Whitney U test was applied for data of this type that were
not normally distributed. Categorical variables were compared by the
ap
A total of 146 fecal samples were subjected to metagenomic shotgun sequencing in
order to investigate the variations in the gut microbiome. Following the removal
of low-quality reads and human DNA reads, 52.4 million high-quality reads were
aligned to a comprehensive gene catalogue, resulting in an average mapping rate
of 87.2
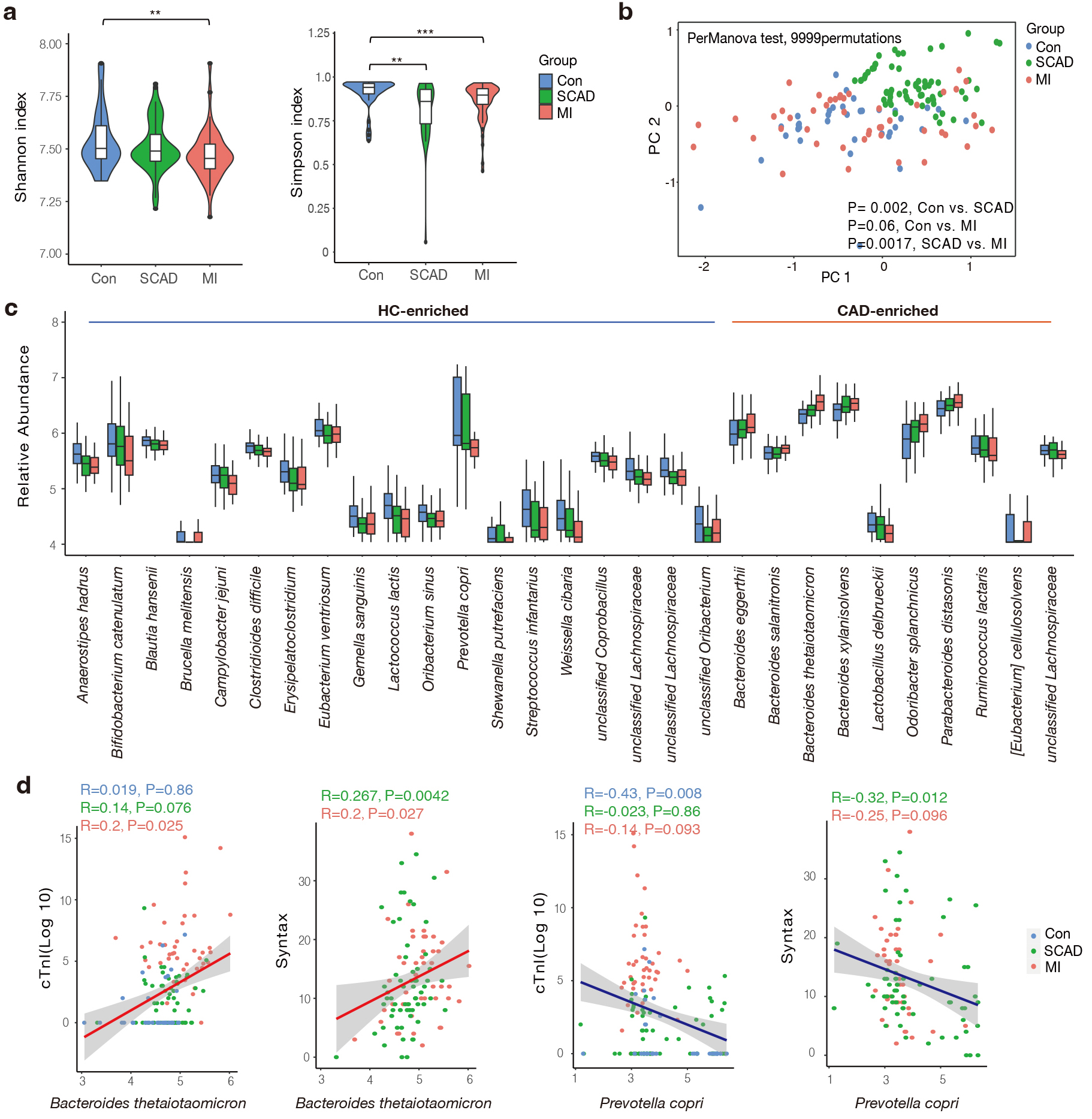
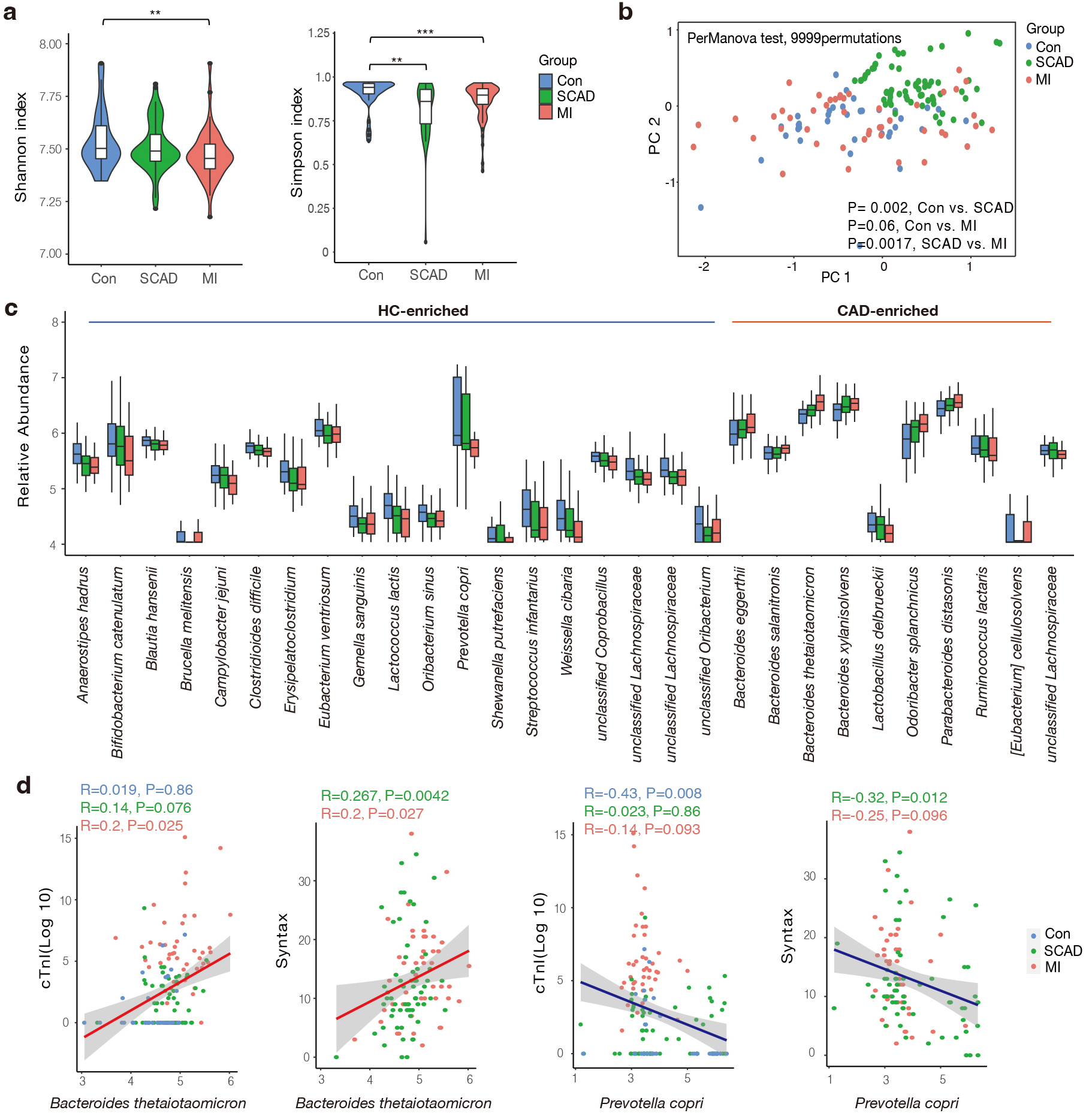 Fig. 1.
Fig. 1.
Gut microbial characteristics of the cohort. (a) SCAD and MI
patients were characterized by lower microbial richness in Shannon and Simpson
indexes based on genera level compared to healthy controls. (b) Differentially
changed bacteria feature in SCAD, MI and Control according to distance-based
redundancy analysis based on the Bray-Curtis distance. perMANOVA test, 9999
permutations, p values were annotated. (c) Species significantly changed
in the CAD compared to Control group (p value
Then, we aimed to screen dominant bacteria with prognostic value for CAD progression (Supplementary Table 2). In total, 19 species were elevated in the control group while 10 species were more abundant in the CAD group, respectively (Fig. 1c). These CAD-enriched bacteria, including Bacteroides thetaiotaomicron, Bacteroides xylanisolvens were positively correlated to AS burden and atherothrombotic risk represented by the thrombolysis in myocardial infarction (TIMI) risk score (Fig. 1d and Supplementary Fig. 1) [34]. In accordance with the findings of previous studies, members of the Lachnospiraceae family are the primary producers of short-chain fatty acids, showed dominant relevance in healthy subjects [35]. We found Prevotella copri was more abundant in the control group, which was negatively associated with Syntax score and cTnI levels (Fig. 1d). It has been reported that prevotella copri was more common in the gut microbes who prefer plant-rich diet consumption and mediate glucose response [36]. In summary, we found gut microbiota composition and structure differed between the SCAD and MI groups, and closely correlated with the phenotype of CAD severity.
Since gut microbiota interacts with their host through circulatory metabolic exchange, we integrated gut microbial functions and serum metabolomics in the states of CAD progression. The metabolic potential of gut microbiota was represented by KEGG modules, which are manually curated pathway units consisted of KOs. 21 KEGG modules were remarkably associated with one or more of the CAD phenotypes (Supplementary Table 3). Next, we examined 146 serum samples under polar ionic/lipid mode for untargeted metabolomic analysis, and 562 metabolites were further binned into 35 co-abundance metabotypes (Supplementary Table 4 and Supplementary Fig. 2). In accordance with previous reports [37], lipid metabotypes enriched in the control subjects mainly included phosphatidylcholine (PC), phosphatidylethanolamine (PE) and phosphatidylserine (PS). Meanwhile, metabotypes elevated in different stages of CAD included amino acids, prenol lipids, benzenoids, fatty acyls and furanones (Supplementary Fig. 2). Importantly, these bacterial KEGG modules were also correlated to CAD phenotype filtered metabotypes (Fig. 2a), with the majority also differing in abundance in the expected direction in the cohort. The bacterial pathways positively associated with CAD progression contained enzymes for lipopolysaccharides (LPS), menaquinone (Vitamin K2) biosynthesis, citrate cycle and various transport systems, which have been proven closely linked to CVD [38, 39]. In contrast, the microbiome negatively associated with CAD progression contained bacterial genes important for leucine and trehalose biosynthesis. Notably, methanogenesis and tryptophan pathway showed a positive correlation with cTnI levels which is an indicator of atherothrombosis. Research has shown methane may exert cardioprotective effects via its anti-oxidative and anti-inflammatory activities [40]. As an essential aromatic amino acid, tryptophan is the precursor of indole derivatives which normally under the direct regulation of the gut microbiota [41].
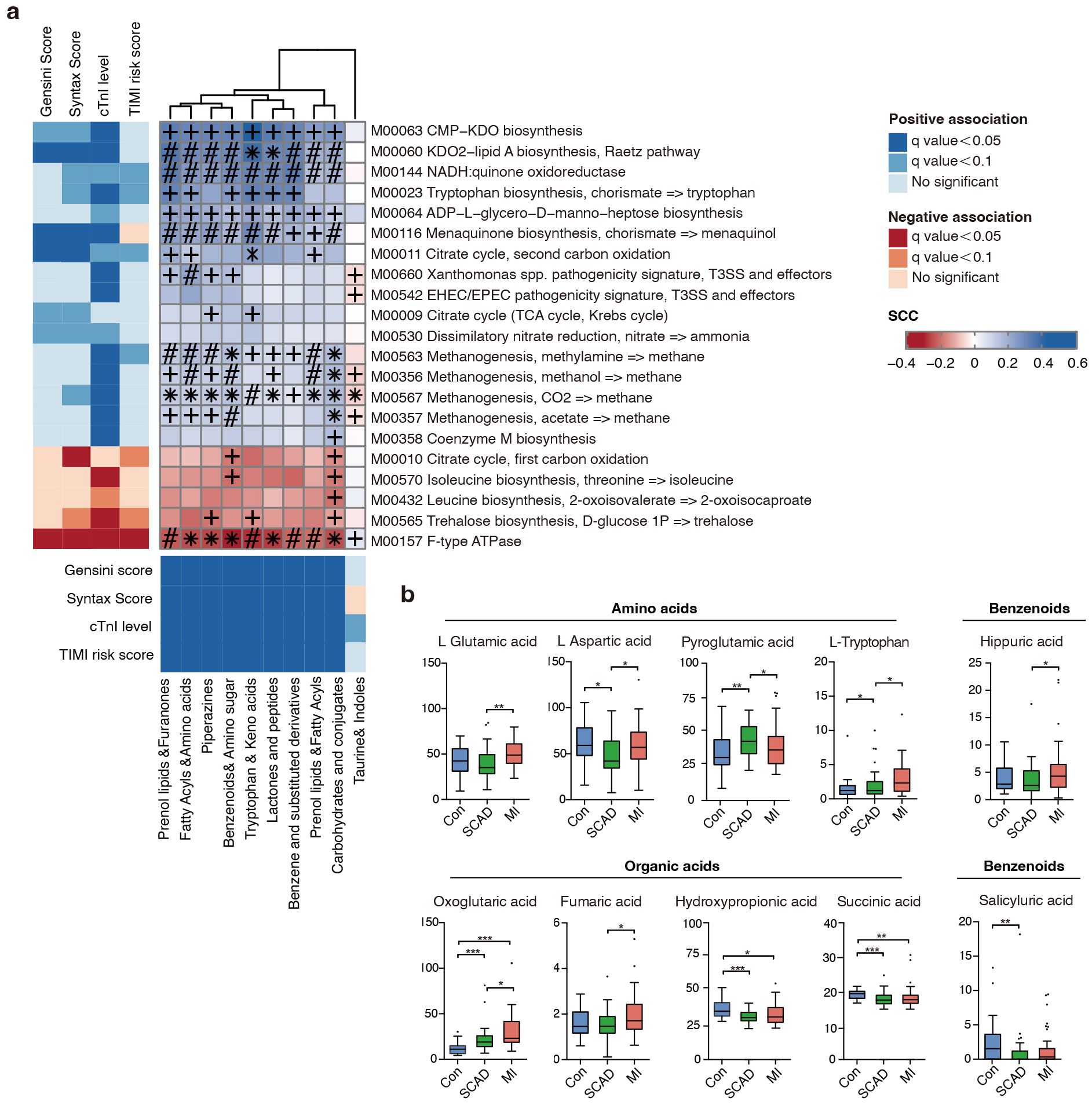 Fig. 2.
Fig. 2.
Fine-grained correlation profile of metagenomics, metabonomics
and clinical traits in 146 subjects. (a) The left and under panel showed
associations (Mann-Whitney U-test; FDR
Since amino acids, bile acids, benzenoids and elements of organic acids were focuses of attention. We next measured 132 serum metabolites using targeted metabolomics in 10 categories and 10 compounds were finally identified as being significantly associated with CAD progression (Fig. 2b, Supplementary Table 5). We noticed that L-tryptophan increased with CAD severity in the serum, reflecting a CVD predictable prognosis value. Overall, we investigated the microbial modules in relation to serum metabolites and CAD phenotypes using cross-domain associations, indicating gut microbiota regulate cardiometabolic disease as a novel endocrine organ.
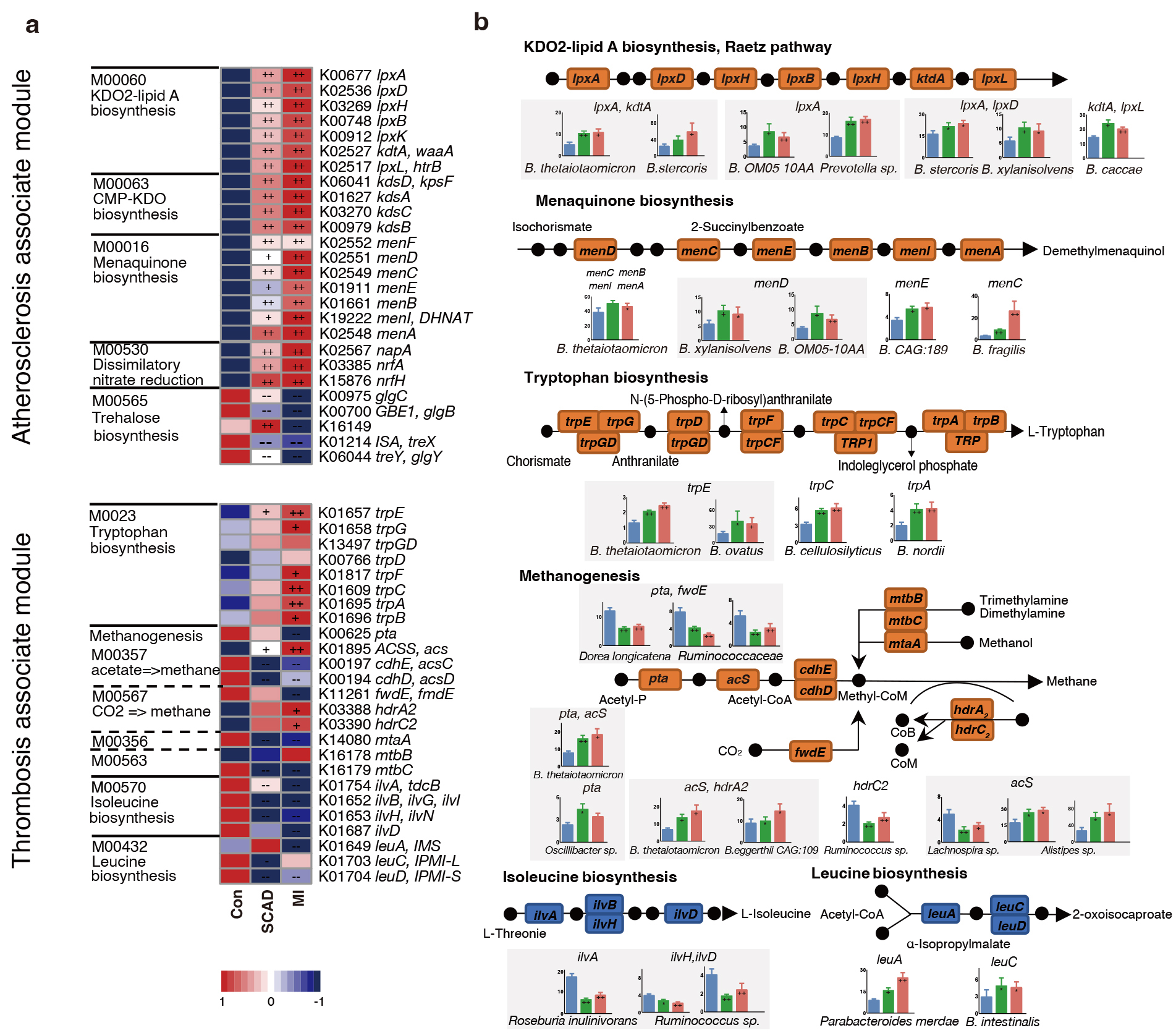
In order to check the roles of microorganisms in metabolic pathways, we dissected the critical KOs related to CAD progression and identified the key bacteria which were responsible for KO variation. Among the 5 main atherosclerosis-associated microbial modules, a significant elevation was observed in 21 KOs and a significant depletion in 5 KOs in at least one of the disease stages when compared to the control (Fig. 3a). The alterations in the gene were illustrated in a pathway representation, which was constructed manually by modifying KEGG pathway maps (Fig. 3b). Bacteria expressed lipopolysaccharide (LPS) biosynthesis enzyme A (lpxA) and 2-Keto-3-deoxyoctonate transferase A (kdtA) gene in 2-keto-3-deoxyoctulosonic acid (KDO2)-lipid A biosynthesis including Bacteroides thetaiotaomicron, Bacteroides stercoris and Bacteroides xylanisolvens, were also found significantly elevated in CAD patients. Bacteroides were also strongly related to menaquinone biosynthesis pathway, Bacteroides sp. OM05-10AA, Bacteroides fragilis and Bacteroides thetaiotaomicron were identified as top marker expressing gene menaquinone biosynthesis enzyme C (menC) and menaquinone biosynthesis enzyme D (menD). Ruminococcaceae and Faecalibacterium prausnitzii in control subjects showed a positive relationship with AS-negative modules such as trehalose biosynthesis. Bacteroidetes may promote thrombosis since they were involved in the synthesis of L-tryptophan for expressing abundant tryptophan biosynthesis gene E (trpE), tryptophan biosynthesis gene C (trpC) and tryptophan biosynthesis gene A (trpA). Overall, Bacteroidetes seem to affect the progression of CAD at different stages by participating in LPS, menaquinone and methanogenesis pathways.
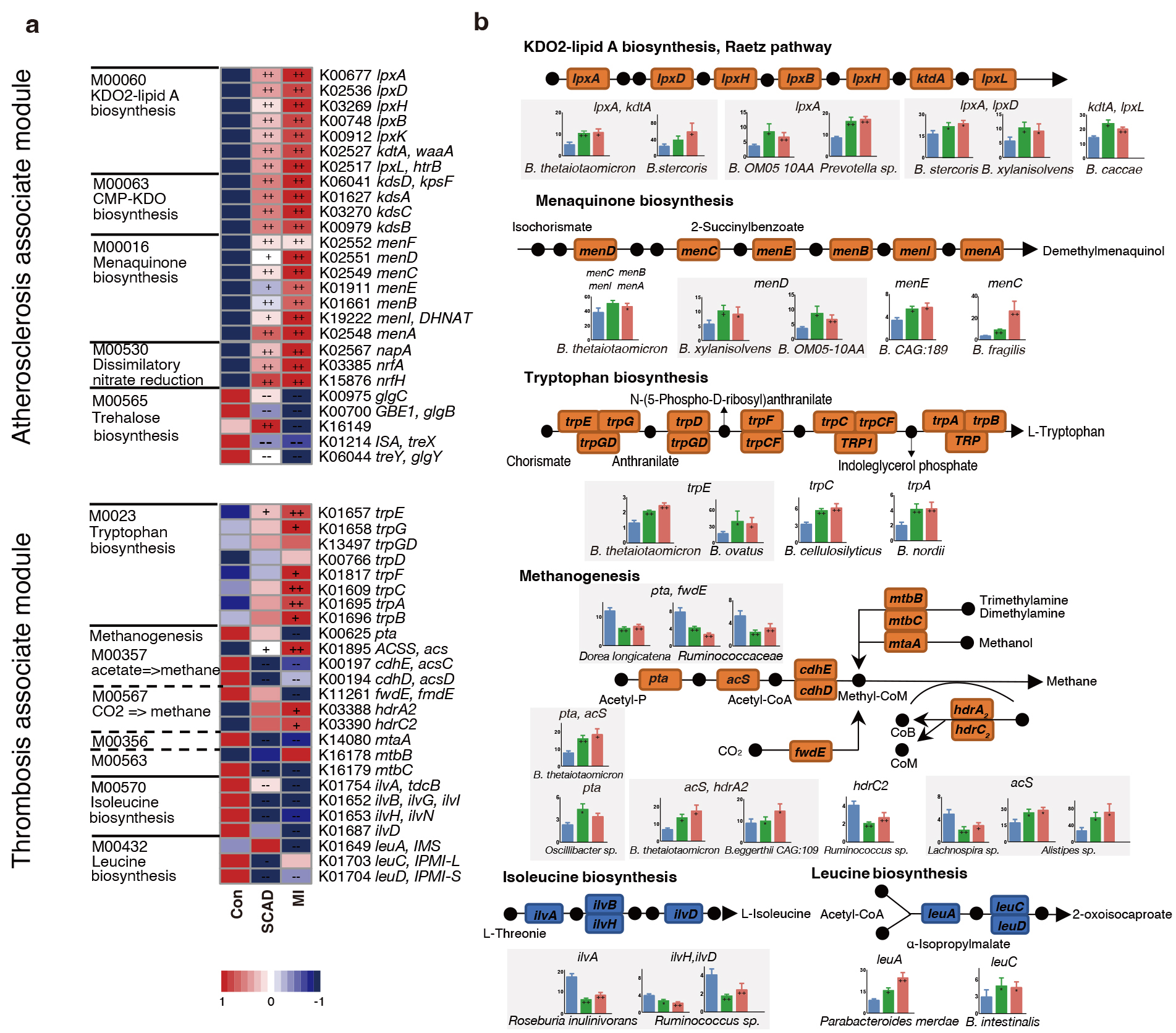 Fig. 3.
Fig. 3.
CAD progression-associated changes in bacteria summarized in KO
genes and KEGG pathways. (a) Relative abundance of KOs altered in each of CAD
stages compared to the Control were shown in the heat map. Significant elevation
or depletion were denoted as follows: ++, elevation with p
In the pathways related to myocardial damage, most of the methane-producing KOs decrease in CAD such as phosphotransacetylase (pta) and forward transferase E (fwdE). These KOs were negatively correlated with Ruminococcaceae spp. while positively correlated with Bacteroidetes and Alistipes sp. Meanwhile, genes (ilvA, leuA) involved in the biosynthesis of leucine and isoleucine were significantly depleted in SCAD and MI. Roseburia inulinivorans was found to be active in iso-leucine production, while Parabacteroides merdae and Bacteroides intestinalis reduced amino acid generation. In conclusion, Bacteroides spp. were found to exhibit a dominant functional role in CAD progression through metagenomic analysis.
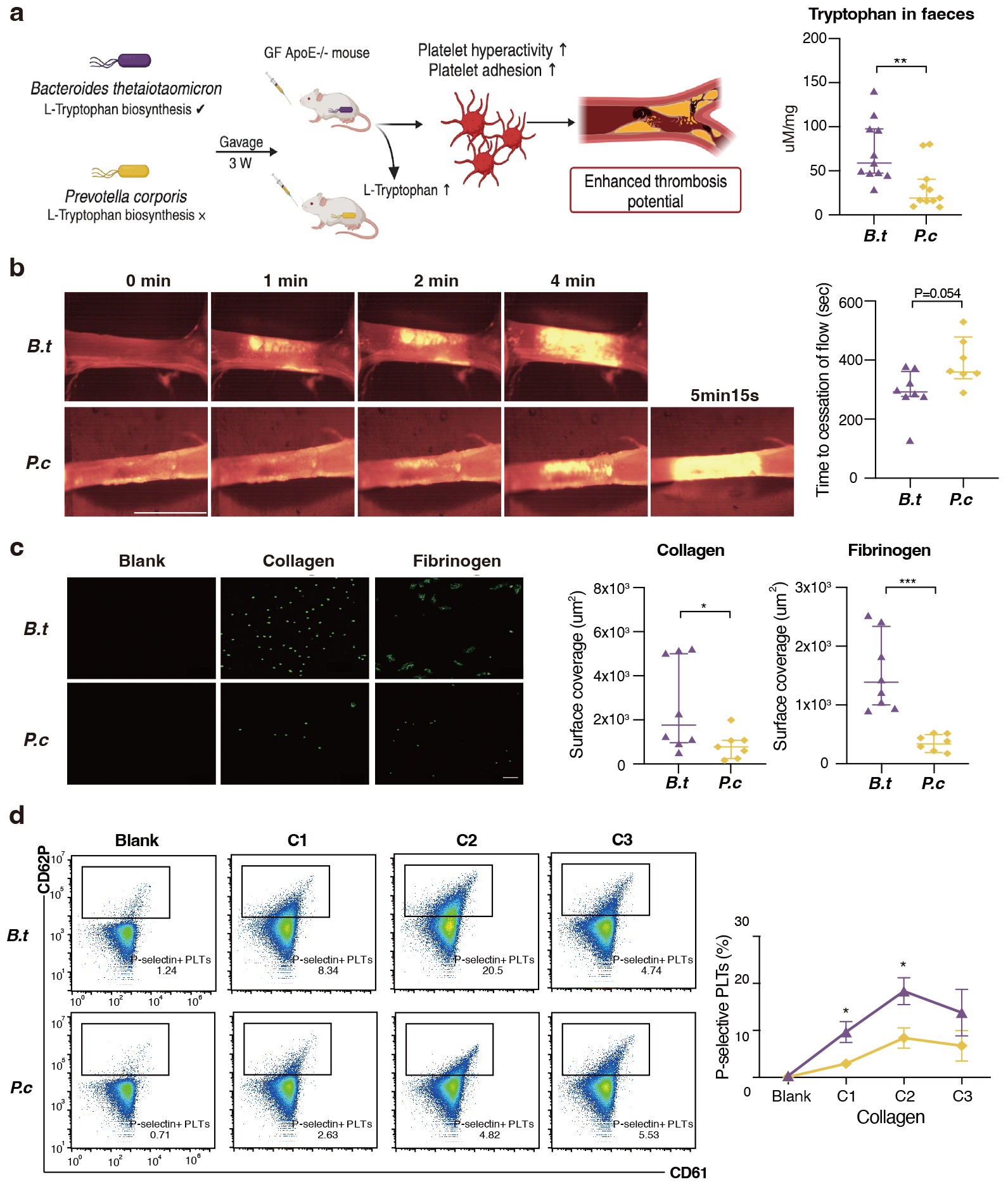
Since B. thetaiotaomicron was the strongest driver strain for
L-tryptophan biosynthesis, suggesting a possible causal relationship. Meanwhile,
we have observed a robust association between increasing B.
thetaiotaomicron and major adverse cardiac events in our prospective cohort
(Data not published), served as impetus for research aimed at verifying the
hypothesis that B. thetaiotaomicron may modulate platelet activity. We
chose another LPS-producing Gram-negative bacterium Prevotella copri as
a control strain since this strain does not contain genes encoding
Trp biosynthesis. Both Bacteroides and Prevotella have
been identified as dominant contributors to human gut enterotypes [23]. To
experimentally address the issue, we colonized B. thetaiotaomicron and
P. copri with germ free ApoE-/- mice fed on high-fat diet for
three weeks. The concentration of fecal L-tryptophan levels in B. thetaiotaomicron mice was observed to be approximately twice that of the
concentration observed in the P. copri mice (Fig. 4a). Notably, mice
kept with B. thetaiotaomicron showed enhanced in vivo
thrombosis potential, as we observed that time to blood flow cessation was
shortened (indicating pro-thrombotic phenotype) compared to P. copri
(286.1
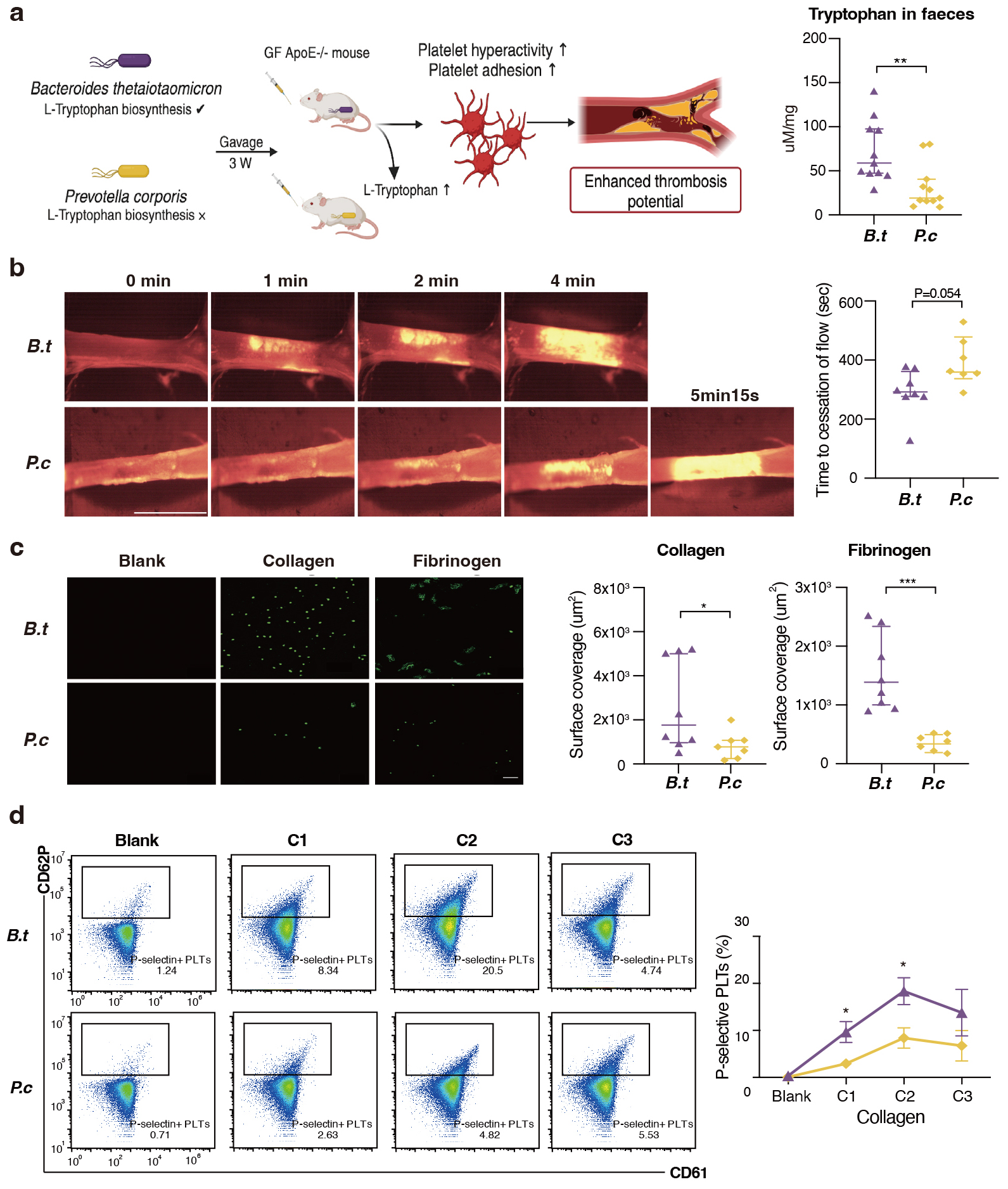 Fig. 4.
Fig. 4.
B. thetaiotaomicron induce L-tryptophan biosynthesis
and provoke thrombosis by microbiota transplantation in germ free mice. (a) We
colonized Bacteroides thetaiotaomicron and Prevotella copri
with germ free ApoE-/- mice fed on high-fat diet for three weeks (n = 11
pergroup). L-tryptophan levels in mice faeces gavaged with B.
thetaiotaomicron or p. copri (11 per group). (b) Intravital
epifluorescence video microscopy of thrombus formation at different time
(representative images) in the FeCl3 -injured carotid artery of germ free
ApoE-/- mice gavaged with B. thetaiotaomicron (n = 8) or p. copri (n = 7) with analysis of occlusion times. Scale bar 2 mm.
(c) Standardized blood flow chamber for platelet adhesion on collagen and
fibrinogen. End-stage representative images of platelet coverage area were shown.
Scale bar 10 µm. (d) Expressions of platelet activation markers
P-selectin (CD62P) in vitro by increasing collagen concentrations (10
µg/mL, 15 µg/mL, 20 µg/mL). Data were
presented as median with interquartile range except for flow cytometric results.
For (a–c), p values were determined by either Mann-Whitney U-test; for
(d), data were examined by two-way ANOVA. *, p
Research recently has revealed an unexpected interaction between gut microbial metabolism and the host to modify the risk of developing CVD [44, 45]. We strengthened the novelty to dissect clinical CAD stage–specific microbial features associated with disease progression by integrating multi-omics for clinical phenotype, gut microbiome, and fasting serum metabolome. The current study directly demonstrated that CAD-associated functional components of gut microbiome: notably the upregulated potential for lipopolysaccharide, L-tryptophan and methanogenesis pathways. Further, B. thetaiotaomicron colonization studies confirm that gut commensal may modulate platelet hyperresponsiveness and thrombosis potential. These conclusions contribute to evaluating the relationship between adverse cardiovascular outcomes and imbalanced microbial homeostasis.
Atherosclerotic plaque rupture and thrombosis can lead to severe ischemic events, result in MI and death [46]. Our results demonstrate that microbiome shifted with the progression of CAD compared to controls and highlighted the remarkable role of Bacteroidetes in the onset and development of CAD. Members of Bacteroides spp. are potential commensals and mutualists of the human colon [47], and they only function as “providers” for microbes residing close to them, but also profoundly affect the susceptibility of the host to inflammatory diseases [48, 49]. While few other studies reported a negative association between Bacteroidetes abundance and CVD populations [50, 51, 52], the inconsistency may be due to single case-control comparison analysis design and the CAD patients recruited in these researches contained various subtypes. Meanwhile, CAD patients exhibited higher carbohydrate intake according to our previous study [12], which shed light on the opinion that diet habit may causally select for gut microbial features. As recent research reinforced that gut microbiota modulates the interplay between diet and cardiometabolic disease risk [53]. Based on the current conclusions, the structure in bacteria between different stages of CAD might represent distinct metabolic roles for atherosclerotic plaque growth at early stages and plaque rupture or intra-arterial thrombus at late stages of CAD.
The human metabolome consists of endogenous metabolites and bacteria-derived metabolites. The metabolic patterns observed in patients with varying stages of CAD indicate that atherothrombosis may be associated with a universal metabolic disturbance. Similar to our previous findings, integrating metagenomics and metabolome profiles showed that benzenoids and menaquinols, which are normally biosynthesized by bacteria, significantly perturbed with the development of CAD [12]. The observed associations suggest that Bacteroides thetaiotaomicron involved in the synthesis of tryptophan may reflect CVD prognosis linked to MI, potentially play a unique role in the maintenance of platelet function and artery thrombosis. L-tryptophan and corresponding catabolites have been widely studied as key regulators of cardiovascular homeostasis, as well as immune homeostasis [41, 54, 55]. Several cohorts [56, 57] mainly consisting of individuals from Western countries found circulatory tryptophan was inversely associated with cardiac mortality and thrombotic event while downstream indoles metabolites were positively correlated to major adverse cardiac event. We speculate that the inconsistency is attributed to diet difference, and have conducted work using a verification cohort including more than 2000 patients and analyzed the important role of diet.
More emerging evidence has confirmed the relationship between bacteria and thrombosis formation [7, 58, 59]. Gut microbiota, an actuating trigger of a systematic immune response, could affect atherothrombosis and subsequent adverse prognosis. Our data revealed severe clot formation in the B. thetaiotaomicron colonization GF mice compared with their P. copri counterparts, indicating a stimulatory effect of gut microbe on atherothrombosis. We analyzed platelet hyperresponsiveness on platelet activating matrices to mimic physiological arterial flow conditions, in order to detect differences in platelet activation. Similar to our in vivo results, we found B. thetaiotaomicron to promote prothrombotic platelet adhesion function. More importantly, vascular inflammation not only “fuels” atherosclerosis but also creates the milieu for episodes of thromboses [60]. In conclusion, given the pivotal role of thrombi in immunothrombosis, it is plausible that the mutualistic relationship between gut microbiota and host not only impairs immunovigilance but also influences arterial thrombus formation under steady-state conditions.
The present study had several limitations that should be considered when interpreting the results. Despite the numerous findings of B. thetaiotaomicron on thrombosis potential, the mechanistic insights for bacteria-derived tryptophan within platelets function remains unknown. Although we selected P. copri as control strain in order to minimize the interference of LPS, other immune factors may be involved in the formation of thrombi and further research is needed. The current research mainly focused on the association of B. thetaiotaomicron with platelet function, and we have also evidenced that long-term colonization of gut microbiota cause formation of atherosclerotic plaques in another study (Data unpublished).
The gut microbial ecosystem is capable of producing a variety of metabolites that are carried via circulation and influence host biological processes by distributing to distant body sites. The microbiome in the human gut may survive, decline or flourish in response to endoenvironmental perturbations. Multi-omic studies could provide a global understanding of bacterial variations that occur in CVD populations as a consequence. Collectively, our findings have implications on future studies that aim to depict microbiome-health association network across disease stages and may enable the design of non-invasive diagnostic tools.
Since CAD is a chronic, long-term pathologic disease that is closely associated with inflammatory cascades [61]. The underlying mechanism responsible for the sudden transformation of a stable atherosclerotic plaque to thrombosis may cause life-threatening death, which normally occurs after decades of progression [62]. Therefore, seeking novel and effective biomarkers for monitoring plaque rupture is important for prevention of cardiovascular death. Our results show that gut microbiota is closely correlated to CAD progression via the mediation of circulatory metabolites. Furthermore, the present study highlights that coronary thrombosis may be influenced by the tryptophan output of the Bacteroides spp., as well as the presence of microbial-related methanogenesis pathways. The current study provides ideas for understanding the interaction of host-intestinal microbiota in the pathogenesis of atherothrombosis, and modulating gut microbiota as a therapeutic target.
The microbiome data in this study are available at (https://db.cngb.org/cnsa/), project number: CNP0001804.
HHL, MYT and SYZ conceived the project. HHL, MYT and SQF collected the samples and metadata. HHL, MYT and RT performed the computational analysis and processed the metabolomic samples. HHL, SQF designed and performed the mouse experiments. HHL and RT wrote the manuscript, SQF, MYT and SYZ revised the manuscript. SYZ supervised the project. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
This study was approved by the Medical Ethics Committee of Peking Union Medical College Hospital (JS-1195). All patients or their families/legal guardians gave their written informed consent before they participated in the study.
Not applicable.
This work was supported by National Natural Science Foundation of China (grant 82170486 and 81670329 to SY.Z.) (grant 82300382 to HH.L.); National Key Research and Development Program of China (grant 2022YFC2703100 to SY.Z.); China Postdoctoral Science Foundation (2021TQ0050 to HH. L.).
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/j.rcm2511395.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.