





1 Department of Physiology, HeartOtago, School of Biomedical Sciences, University of Otago, 9010 Dunedin, New Zealand
†These authors contributed equally.
Abstract
Ischaemic heart disease (IHD) remains the leading cause of mortality worldwide. Current pharmaceutical treatments focus on delaying, rather than preventing disease progression. The only curative treatment available is orthotopic heart transplantation, which is greatly limited by a lack of available donors and the possibility for immune rejection. As a result, novel therapies are consistently being sought to improve the quality and duration of life of those suffering from IHD. Stem cell therapies have garnered attention globally owing to their potential to replace lost cardiac cells, regenerate the ischaemic myocardium and to release protective paracrine factors. Despite recent advances in regenerative cardiology, one of the biggest challenges in the clinical translation of cell-based therapies is determining the most efficacious cell type for repair. Multiple cell types have been investigated in clinical trials; with inconsistent methodologies and isolation protocols making it difficult to draw strong conclusions. This review provides an overview of IHD focusing on pathogenesis and complications, followed by a summary of different stem cells which have been trialled for use in the treatment of IHD, and ends by exploring the known mechanisms by which stem cells mediate their beneficial effects on ischaemic myocardium.
Keywords
- ischaemic heart disease
- stem cells
- clinical trials
- pluripotent stem cells
- adult stem cells
- paracrine mechanisms
Despite global research efforts, cardiovascular disease (CVD) remains the leading cause of mortality worldwide [1]. The term CVD encompasses a multitude of cardiac pathologies including valvular defects, arrhythmias, vasculopathies and congenital malformations. Ischaemic heart disease (IHD) is the most prevalent form of CVD, characterised by insufficient blood supply to myocardium relative to oxygen demand [2]. This ischaemic imbalance results from the formation of atherosclerotic lesions in the tunica intima of coronary arteries, causing progressive stenosis of the coronary lumen and a corresponding decrease in blood flow [3].
As IHD progresses, functional cardiomyocytes are lost owing to ischaemia with remaining myocytes forced to hypertrophy to compensate and maintain heart function [4]. This progressive loss of cardiomyocytes often eventuates in chronic heart failure (CHF)—where the cardiovascular system is unable to supply blood to tissues at normal perfusion pressures. Furthermore, increasing coronary artery stenosis places the patient at risk of plaque rupture and acute coronary syndromes including myocardial infarction (MI), unstable angina and cerebrovascular accident [5]. Current pharmaceutical treatments only delay the progression of IHD, but are unable to reverse existing damage to the myocardium. The replacement of lost cardiovascular cells, along with improving the function and survival of remaining cells is vital for long-term improvement of cardiac function in patients with IHD. Presently, orthotopic heart transplantation is the only way to achieve this goal. However, a limited number of suitable donors and the need for lifelong immune system modulation limit the availability of allotransplantation [6].
Stem cells have garnered global attention as a therapy for a number of pathological conditions following their discovery in 1961 [7]. They have the theoretical potential to both halt the progression of IHD and reverse existing damage by replacing lost cardiac cells, improving the function of resident cells, and the release of beneficial paracrine factors. However, recent studies demonstrate that different stem cell types have varying efficacy in repairing the ischaemic heart [8, 9]. This review will first provide an overview of IHD focusing on pathogenesis and complications, followed by a summary of different stem cell populations trialled for the treatment of IHD and end by exploring the known mechanisms by which these stem cells mediate their beneficial effect on the ischaemic myocardium.
IHD is driven by the formation of atherosclerotic plaques within the tunica intima of coronary arteries, occluding the coronary lumen (Fig. 1). This atherogenesis is thought to stem from a combination of endothelial dysfunction and hypercholesterolaemia [10, 11]. Risk factors for these states include diabetes mellitus, hypertension, smoking, obesity and a lipid-rich diet [12]. Together, these states act to increase the vascular permeability of coronary arteries, allowing the migration of lipids including the cholesterol rich low-density lipoprotein (LDL) through the endothelial lining, into the tunica intima [13]. Here, LDL particles are oxidised by reactive oxygen species (ROS) and become pro-inflammatory, driving the activation of endothelium and expression of vascular adhesion molecule-1 (VCAM-1) and intercellular adhesion molecule-1 (ICAM-1) on the endothelial cell surface [14, 15]. Chemotactic agents including chemokine ligand 2 (CCL2) are released from activated endothelium and recruit mast cells, neutrophils and monocytes which bind to the aforementioned endothelial cell surface adhesion molecules [16].
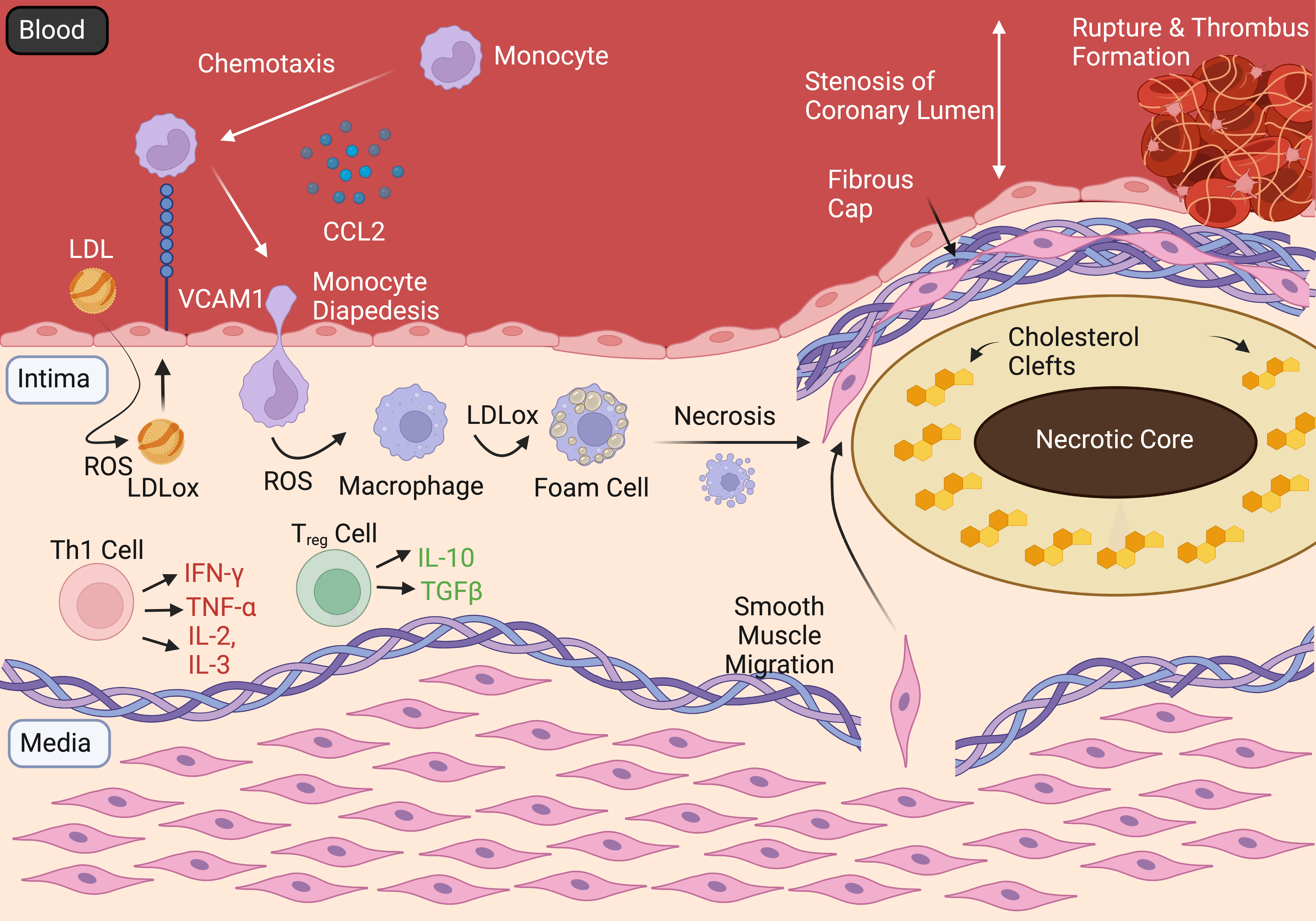 Fig. 1.
Fig. 1.Pathogenesis and Complications of IHD. Figure summarising the
development and progression of the atherosclerotic plaque, which obstructs the
coronary lumen and if unstable—can rupture, with subsueqent thrombus formation
occluding the affected coronary artery—resulting in acute coronary syndromes.
LDL, low density lipoprotein; VCAM1, vascular cell adhesion molecule 1; CCL2,
chemoattractant protein 1; ROS, reactive oxygen species; LDLox, oxidised LDL;
IFN-Y, interferon gamma; TNF-
Following attachment to adhesion molecules, monocytes migrate into the tunica intima, where they differentiate into macrophages in the presence of ROS. These macrophages are involved in further leucocyte recruitment and cytokine release in addition to phagocytosis of oxidised LDL, after which they are referred to as foam cells [17]. Accumulation of foam cells forms a ‘fatty streak’ - the earliest gross pathological sign of atherosclerosis. Over time these foam cells necrose through apoptosis, creating the characteristic ‘necrotic core’ of the atherosclerotic plaque [18]. New blood vessels primarily from the tunica adventitia can grow into the base of atherosclerotic lesions. This can further advance plaque growth as these vessels provide yet another avenue for monocytes and other immune cells to reach the plaque [19]. Meanwhile, adjacent endothelial and smooth muscle cells (SMCs) secrete cytokines and growth factors, causing SMCs to migrate to the luminal side of the vessel wall. This leads to the formation of a fibrous cap composed of collagen, SMCs, macrophages and T lymphocytes (Fig. 1) [20].
Depending on the progression and stability of the atherosclerotic plaque, IHD can progress to develop further complications. The degree of plaque stability is directly proportional to the thickness of the fibrous cap surrounding the necrotic, lipid-rich core [21]. Stable plaques result in stable angina, characterised by ischaemic chest pain induced by exertional stress. In the absence of myocardial scarring, stable angina is rarely fatal and is usually relievable with rest or nitroglycerin [22, 23].
Acute coronary syndromes (ACS) result from atherosclerotic plaque rupture, with subsequent thrombus formation leading to incomplete or total occlusion of the coronary lumen [24, 25]. ACS are medical emergencies, drastically reducing blood flow to the myocardium, resulting in unstable angina and MI. Myocardial infarction can be further classified into ST-elevation myocardial infarction (STEMI) or Non ST-elevation myocardial infarction (NSTEMI). Both NSTEMI and STEMI are characterised by myocardial necrosis subsequent to the onset of sudden ischaemia [26].
In the event of MI, both the renin-angiotensin-aldosterone system (RAAS) and the sympathetic nervous system (SNS) become activated to maintain adequate perfusion of the vital organs [27, 28] (Fig. 2). This is achieved by inducing vasoconstriction of systemic arterioles, increasing total peripheral resistance (TPR), and increasing renal reabsorption of sodium and water to elevate systemic blood pressure. Angiotensin II, the key component of the RAAS system increases the production of ROS within the heart through NADPH oxidase 2 (NOX2) activation which in turn activates downstream pathways involved in cardiac hypertrophy including protein kinase B (Akt), nuclear factor kappa B (NF-kB) and extracellular signal-related kinase (ERK1/2) signalling [29].
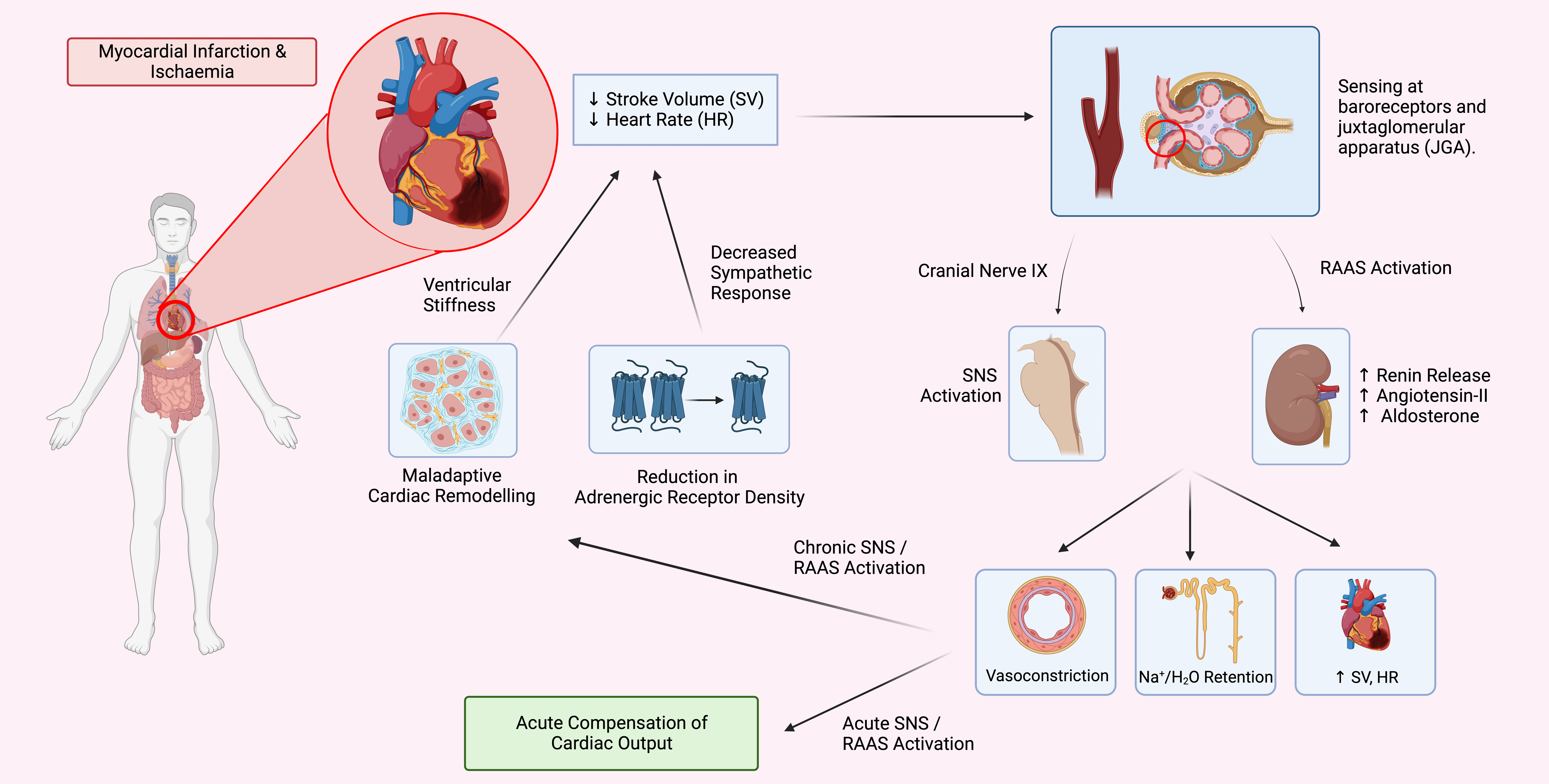 Fig. 2.
Fig. 2.The Cycle of Heart Failure. In response to myocardial injury, neurohumoral mechanisms are activated, compensating for decreased cardiac output. However, in the long term, these mechanisms cause further wall stress and cardiac remodelling, further damaging cardiac tissue and advancing the progression of heart failure. RAAS, renin-angiotensin-aldosterone system; SNS, sympathetic nervous system.
Further, an inflammatory response simultaneously occurs by initiating the recruitment of macrophages to the infarcted area to remove dead cells and matrix debris by phagocytosis [30]. Following MI, resident cardiac fibroblasts (CFs) become pro-inflammatory, increasing their secretion of pro-inflammatory cytokines including interleukin-1 (IL-1). These CFs undergo differentiation into myofibroblasts, increasing the rate of extracellular matrix (ECM) deposition and scar formation [31, 32].
Activation of these compensatory mechanisms are crucial to sustain cardiac output during the initial phase following MI. However, sustained activation of these pathways can eventuate in heart failure. Sustained increases in RAAS and SNS activation increase ventricular wall stress, contributing to maladaptive cardiac remodelling, reducing ventricular function (Fig. 2) [33, 34, 35].
Furthermore, activation of the SNS desensitises
The management of IHD necessitates a combination of lifestyle and pharmacological approaches, with the ongoing involvement of a multidisciplinary healthcare team. The primary aims of IHD management are symptom relief and prevention of disease progression. Lifestyle interventions aim to control for modifiable risk factors including a sedentary lifestyle, hyperlipidaemia, smoking and hypertension [12]. These factors may be controlled by regular exercise, smoking cessation, weight loss and the maintenance of a healthy diet. A recent systematic review of structured lifestyle modification programmes highlighted significant reductions in all-cause mortality, cardiac mortality and cardiac re-admissions, emphasising the importance of lifestyle modifications [38]. However, these changes may prove challenging to elicit in practice due to several barriers the patient may be facing.
Pharmaceutical interventions for IHD range from anti-hypertensives including angiotensin converting enzyme (ACE) inhibitors, antidyslipidaemics such as atorvastatin along with anticoagulant therapy to lower the risk of thrombus formation [39]. In the case of acute MI, the immediate aims of treatment are to promote reperfusion to the myocardium, reducing infarct size. This can be achieved with prompt fibrinolytic, antiplatelet and antithrombotic agents, along with percutaneous coronary intervention (PCI) to restore blood flow mechanically via stenting the affected coronary artery [40].
If the patient has progressed to CHF, the goal of pharmacological therapy is to improve cardiac contractility and to reduce fluid overload [41]. Orthotropic heart transplantation remains the only curative treatment, unfortunately limited by low donor availability [42]. None of the current treatments are able to reverse the disease process or restore lost cardiac cells. As a result, interest has grown globally in seeking novel therapies for IHD, among which cell-based therapies have garnered attention due to their potential to promote cardiac repair and regeneration.
Stem cells are undifferentiated and self-renewing, forming identical clones with the potential to differentiate into an array of specialised cell types, first discovered in 1961 [7]. Progenitor cells are the immediate descendants of stem cells, formed through asymmetric division, but instead give rise to tissue-specific progenitors [43]. Stem cells can be further categorised based on their degree of lineage commitment (Fig. 3). Embryonic stem cells (ESCs) exhibit pluripotency, while adult stem cells exhibit limited differentiation ability (multipotency) and less potent self-renewal [43]. As adult stem cells are derived from adult organs and tissues, they prove easier to obtain, allow for autologous transplantation and are not associated with the same degree of legal and ethical issues ESCs face. To be an ideal candidate for cardiac repair, stem cells should meet the following criteria; straightforward isolation, scalability into large quantities, the capacity to promote vascularisation, ability to reduce ischaemic imbalance and to differentiate into cardiac cell lineages (for integrating-based cell therapies), appropriate long-term electromechanical stability and integration within host myocardium and exertion of positive paracrine effects through the release of bioactive molecules including pro-angiogenic factors. This review will focus on those stem cells which have been extensively investigated for their use in IHD – these being embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs), skeletal myoblasts (SkMs), adipose derived mesenchymal stem cells (ASCs), umbilical cord mesenchymal stem cells (UCMSCs), endothelial progenitor cells (EPCs), cardiac progenitor cells (CPCs) and stem cells which reside in the bone marrow (often referred to collectively as bone marrow mononuclear cells (BMCs)), or separated into haemopoietic stem cells (HSCs) and bone marrow-derived mesenchymal stromal cells (BdMSCs). However, it should be noted that other adult stem cell populations exist including skin stem cells, epithelial stem cells and neural stem cells. Pre-clinical studies on the use of neural crest stem cells and amniotic epithelial cells for cardiac repair warrant further investigation, furthermore human dermal fibroblasts have been incorporated into synthetic scaffolds with iPSC dcrived cardiomyocytes in pre-clincial studies [44, 45, 46, 47].
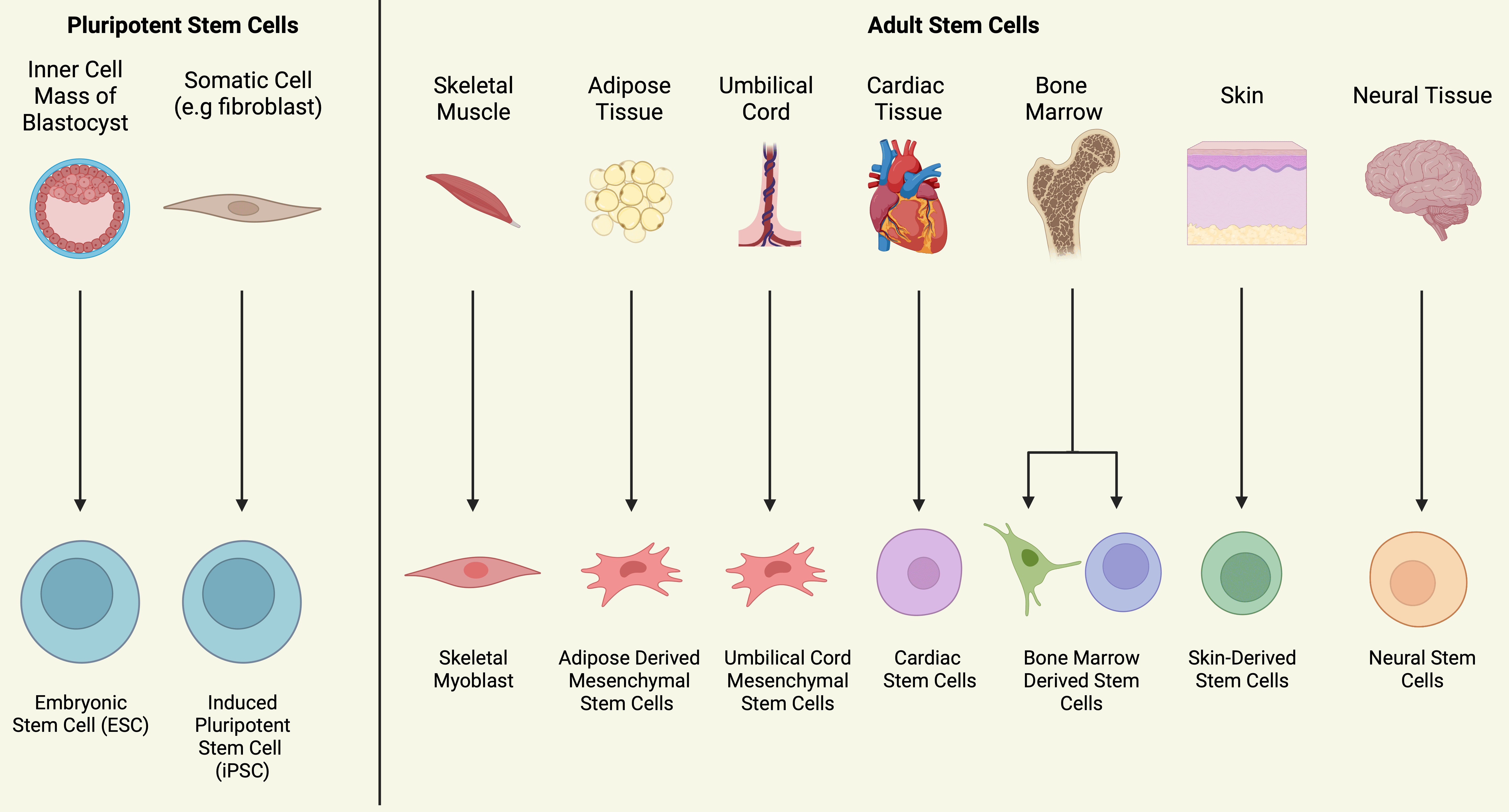 Fig. 3.
Fig. 3.Sources of Stem Cells Used in Cardiac Repair. Figure summarising the most significant sources of stem cells used for cardiac repair. Pluripotent stem cells can be obtained from the inner cell mass or generated from somatic cells by introducing specific transcription factors. Adult stem cells exhibit multipotency and are derived from various adult tissues and organs.
With this as background, we will next review different types of stem cells that have been trialled for their efficacy to regenerate or repair the ischemic myocardium.
ESCs are pluripotent cells extracted from the inner cell mass of human blastocysts. They exhibit the capacity for indefinite symmetrical division; and for asymmetric division into progenitors of mesoderm, ectoderm and endoderm germ layers [48]. Owing to their pluripotency, ESCs are strong candidates for the repair of various adult tissue types including ischaemic myocardium. Several methods have been employed to induce cardiomyogenic differentiation from a pool of ESCs with the resulting cells termed embryonic stem cell-derived cardiomyocytes (ESC-CMs) [49]. ESC-CMs express early cardiac-specific transcription factors including homeobox protein Nkx2.5, GATA binding protein 4 (GATA-4), myocyte enhancer factor 2C (MEF2C) and T-box transcription factors Tbx-5 and Tbx-20 [50]. Furthermore, ESCs give rise to progenitor cells with temporal regulation of foetal liver kinase 1 (Flk1), Islet-1 (Isl-1) and Brachyury (a T-box transcription factor), demonstrating its capacity for differentiation into cardiomyocytes, endothelial cells and vascular smooth muscle cells in vitro [51, 52].
In a seminal study, Caspi et al. [53] transplanted undifferentiated ESCs into infarcted rat hearts, as it was thought the in vivo cardiac environment may be sufficient to induce cardiomyocyte (CM) differentiation. This proved unsuccessful, and teratomas consisting of cells from all three germ layers were observed, highlighting the importance of inducing CM differentiation and ensuring homogeneity of the cell population prior to transplantation, reducing the risk of tumorigenesis. In a follow-up study, ESC-CMs were grafted into the infarcted area 7–10 days following left anterior descending artery (LAD) ligation in a rat model. Improvements were noted in both scar remodelling and left ventricular function, with transplanted cells able to form gap junctions with host cells as assessed by the expression of the left ventricular gap junctional protein connexin-43. However, ESC-CMs exhibited an immature phenotype, which has been identified in subsequent studies from independent laboratories [54, 55]. ESC-CMs are observed as being smaller in size, having a slower action potential upstroke, a less extensive network of T-tubules and poor sarcomeric organisation among other issues [54, 55]. These differences to adult cardiomyocytes may impair the ability of ESC-CMs to effectively integrate within host myocardium, increasing the risk of arrythmia generation due to the difference in electrophysiology. Promising pre-clinical studies including the recent work of Liu et al. [56] have demonstrated that transplantation of ESC-CMs into infarcted myocardium in a non-human primate model improves left ventricular ejection fraction up to 3 months post-transplantation, however this was associated with an increased incidence of ventricular arrythmias.
To reduce the incidence of these events, current studies are aimed at enhancing
the maturity of ESC-CMs in vitro before transplantation. Methods of
improving maturation include inhibiting hypoxia-inducible factor 1-alpha
(HIF-1
In addition, transplantation of ESC-CMs into the recipient heart is allogenous, requiring lifelong immune system modulation [59]. Further, legal and ethical concerns exist as the traditional method of isolating ESCs for in vitro expansion results in the destruction of the human embryo [48]. While alternative, non-destructive approaches are being explored to isolate ESCs, these legal issues effectively rule out the use of ESCs as a viable treatment in many parts of the world.
Recently, the Transplantation of Human ESC Derived Progenitors in Severe Heart Failure (ESCORT) trial was completed (NCT02057900)—assessing the safety of ESC derived cardiac progenitor cells (CPCs) when engrafted into human patients [60]. Despite the knowledge that ESC-CMs exhibit an immature phenotype, the team behind this trial conducted several pre-clinical trials using these ESC derived CPCs, in which no arrythmia or teratoma formation was observed [61]. Six patients with severe ischaemic left ventricle (LV) dysfunction received ESC derived CPCs delivered as epicardial injection during coronary artery bypass graft (CABG) surgery and were followed up for a median of 18 months. All patients who were followed up had an uneventful recovery with no observed arrhythmia development. Importantly, a significant increase in systolic motion was observed in patients receiving ESC-CPCs. These results were encouraging despite a small sample size and warrant further investigation to confirm the efficacy of ESC derived CPCs in human populations. The on-going HECTOR study (NCT05068674) is a phase I trial evaluating the safety of administering varying doses of ESC-CMs to patients with LV dysfunction secondary to MI. This study aims to recruit 18 patients and will provide further insight into the safety profile of ESC-CM transplantation for cardiac repair.
In spite of these recent clinical trials, there are issues that need to be solved regarding the use of ESC derivatives for repair of the ischaemic heart. These include improving long-term engraftment rates, deciding the stage of differentiation for transplantation, increasing in vitro maturity of ESC-CMs and establishing the optimal dosage and safety profile. Further investigations are also required to accurately determine the nature and underlying mechanisms of the beneficial effects observed on ischaemic myocardium—whether this stems from remuscularisation of the heart or through paracrine mediated mechanisms.
In 2006, Takahashi et al. [62] generated pluripotent cells from adult fibroblasts and coined the term iPSCs, a discovery which would go on to win the Nobel Prize in Physiology or Medicine in 2012. As iPSCs are generated from somatic cells, they are not associated with the same ethical and legal issues as ESCs. Furthermore, patient-matched autologous cells can be created—thereby reducing the possibility of immune rejection. The most common method of iPSC production is the viral transduction of genes encoding the transcription factors octamer-binding protein 3/4 (Oct-3/4), sex-determining region Y box 2 (SOX2), Myc proto-oncogene protein (c-Myc) and Kruppel-like factor 4 (KLF4) [62]. Several studies have documented the ability to differentiate iPSCs into cardiomyocytes (iPSC-CMs), with the differentiation efficacy being comparable to that of ESCs, with ESC-CMs and iPSC-CMs sharing near identical transcriptional profiles [63, 64, 65]. In recent years, methods for the derivation and purification of cardiomyocytes from iPSCs have improved, with some studies reporting a purity of the differentiated cell pool for cardiomyocytes of around 95%, with 100 cardiomyocytes generated per input cell [66, 67].
Despite these advantages, differentiation of iPSCs poses some risks. Viral transduction carries with it the inherent risk of genome insertion at unwanted locations, with the potential to promote oncogenesis and disrupt cellular function [68]. As a result, non-viral vectors (e.g., plasmids) have been developed for iPSC production along with RNA and protein delivery. However, these alternate methods tend to be less efficient in iPSC production, as summarised in a review by Rao et al. [69]. While the ability for autologous transplantation using iPSCs is an advantage, this process can be costly and time consuming, with recent studies instead attempting to reduce the immunogenicity of allogenic iPSCs from healthy donors for clinical application [70]. Differentiation of iPSCs into CMs in vitro is achieved through similar protocols to those established for ESC differentiation, with the resulting CMs sharing an immature phenotype and therefore also carrying the risk of arrythmia and teratoma development [55]. This was highlighted in a recent pre-clinical trial conducted by Shiba et al. [71], in which iPSC-CMs were delivered by intra-myocardial injection into the infarcted hearts of non-human primates. Significant improvements in contractile function were reported, however with an increased incidence of ventricular tachycardia in the treatment group.
In a recently published first-in-human clinical trial in Japan, a 51-year-old male with severe ischaemic cardiomyopathy underwent transplantation of three allogenic iPSC-CM patches onto the ischaemic myocardium [72]. These patches were constructed of clinical grade iPSC-CMs which underwent screening for tumorigenesis and arrhythmogenesis risk. Following transplantation, no tumour development, arrythmias or effects related to immunosuppressive treatment were observed. Furthermore, the patient experienced increased quality of life, systolic motion, reduced LV global wall stress (attenuation of fibrosis) and an increase in the coronary flow reserve after 6 months and one year of follow up. The investigators hypothesized the observed benefit was primarily mediated through paracrine stimulation of angiogenesis. In support of this, a pre-clinical study conducted by Tachibana et al. [73] provided evidence that iPSC-CMs release high levels of interleukin-8 (IL-8), granulocyte colony stimulating factor (GCSF) and vascular endothelial growth factor (VEGF) promoting angiogenesis in the ischaemic heart. While the first-in-human clinical trial is promising, the extent of mechanical contribution of the engrafted iPSC-CMs to cardiac contractility and the percentage of cell retention necessitates further investigation. While no randomised clinical trials of iPSC-CM efficacy have been completed, one trial registered in China (NCT03763136) aims to assess the safety, feasibility, and efficacy of intramyocardial delivery of allogenic iPSC-CMs at the time of CABG surgery in patients with CHF. Another registered clinical trial (NCT03759405) will determine changes in quality of life and cardiac function following intravenous injection of iPSC-CMs in three patients with CHF. The recruitment has not started yet, with the trial expected to be completed in 2024. Although early results from case-studies are encouraging, further research needs to be carried out to standardize the protocol for developing mature cardiomyocytes from iPSCs that can electromechanically couple with endogenous cardiomyocytes for its clinical translation to be successful.
SkMs are multipotent stem cells located between the basal lamina and sarcolemma layers of mature skeletal muscle, becoming activated in response to muscle damage and degeneration [74]. SkMs were one of the earliest adult stem cell types explored as a candidate for cardiac repair due to their ability to form mature myofibers, resistance to ischaemia (owing to their skeletal muscle origin), relative ease of harvest and the potential for subsequent autologous transplantation [75, 76].
Pre-clinical murine studies including the pioneering work of Taylor et al. [77] demonstrated beneficial effects of SkM transplantation in improving cardiac contractility, preventing left ventricular remodelling, and decreasing diastolic pressures [76, 77, 78]. However, SkMs are committed to a myogenic lineage and therefore cannot form fully functioning cardiomyocytes, instead differentiating into muscle fibres called myotubes in vivo [79]. Cardiomyocytes behave as an electrical syncytium due to the presence of intercalated discs, more specifically gap junctions between neighbouring cells [80]. Myotubes derived from SkMs fail to form these gap junctions owing to a decreased expression of the intracellular adhesion molecules N-cadherin and connexin-43, therefore remaining electromechanically isolated from the host myocardium [79]. Thus, myotubes do not contract in synchrony with the surrounding myocardium, predisposing the transplant recipient to arrythmia development. Studies using skeletal myoblasts genetically modified to overexpress connexin-43 demonstrated improved myocardial integration and synchronous contractility stemming from an increase in the number of gap junctions formed with host cardiomyocytes [81, 82, 83]. However, subsequent studies have concluded this is insufficient in preventing the development of arrhythmias [84]. Furthermore, the Myoblast Autologous Grafting in Ischaemic Cardiomyopathy (MAGIC) phase II clinical trial showed no improvement in left ventricular ejection fraction (LVEF) in patients treated with skeletal myoblasts compared with the control group, with a higher number of arrhythmic events observed in the SkM-treated group [85]. Due to these major issues, skeletal myoblasts have not progressed as a candidate for cardiac repair.
BMCs are isolated from bone marrow throughout the body and can be further
divided into HSCs and BdMSCs. HSCs exhibit myeloid and lymphoid differentiation,
whereas BdMSCs differentiate into bone, cartilage and adipose tissue lineages
in vivo [86, 87]. Unfortunately, many clinical trials using BMCs have
not specified which sub-population was used, and definitions of these cells
remain controversial. This section will focus on studies reportedly using HSCs,
and more generally BMCs collectively. BdMSCs will be described in further detail
in subsequent sections. One of the earliest studies involving BMC transplantation
for repair of the ischaemic heart was conducted by Orlic et al. [88],
who injected BMCs enriched for Lin
The Reinfusion of Enriched Progenitor Cells and Infarct Remodelling in Acute
Myocardial Infarction (REPAIR-AMI) study was a placebo-controlled phase III trial
assessing the efficacy of BMCs delivered via intracoronary infusion following
acute MI. This study demonstrated a significant improvement in left ventricular
ejection fraction (LVEF) in the treatment group [95]. The PreSERVE-AMI study was
a phase II study in which autologous purified CD34
Mesenchymal stromal cells (MSCs) are multipotent with potential for differentiation into mesenchymal lineages (osteoblasts, chondrocytes, myocytes, adipocytes and fibroblasts), as defined by the International Society for Cell & Gene Therapy [103]. MSCs are attractive candidates for therapeutic cell transplantation due to their low immunogenicity and immunomodulatory capacity resulting from low major histocompatibility complex (MHC) II expression and the secretion of several anti-inflammatory cytokines. This increases the feasibility of allogenic MSC transplantation as an attractive option for large scale clinical implementation [104]. MSCs isolated from several sources have been investigated for their use in cardiac repair. The following sections will focus on bone-marrow derived mesenchymal stem cells along with MSCs derived from adipose tissue and the umbilical cord.
Bone marrow contains a population of BdMSCs with the capacity for differentiation into osteogenic, chondrogenic and adipogenic lineages [103]. Several studies have trialled the isolation and expansion of this cell population for administration into ischaemic or infarcted myocardium. One of the earliest in vivo studies investigating BdMSCs for cardiac repair was conducted by Toma et al. [105], who injected lacZ-labelled human BdMSCs into the left ventricle of adult mice. Despite a high cellular attrition rate, the surviving BdMSCs began to resemble neighbouring cardiomyocytes and expressed proteins classically found within CMs including troponin T (cTnT), a-actinin and desmin. In a subsequent landmark clinical trial, Chen et al. [106] randomised sixty-nine participants who underwent PCI for acute MI to receive intra-coronary administration of autologous BdMSCs, or a saline control at 18 days post-PCI. After three months of follow-up - reduced perfusion defects, decreased left ventricular end-systolic and end-diastolic volumes, along with an increased LVEF were observed in the BdMSC treatment group.
Recently, Lee et al. [107] conducted a randomised, but open–label study assessing the safety and efficacy of autologous BdMSCs administered into the affected coronary artery at 1-month post-MI (n = 80). After a six-month follow up period, significant improvements were noted in the LVEF of participants who received BdMSCs. Additionally, no serious adverse effects were observed during the procedure or in the six months that followed.
Due to low MHC-II expression, transplantation of allogenic BdMSCs is also a
possibility [104]. In a randomised, double blinded, placebo-controlled study,
Hare et al. [108] provided evidence that administration of allogenic
BdMSCs is not associated with an increased rate of adverse cardiac events, and
furthermore improved left ventricular function and attenuated cardiac
remodelling. Despite beneficial effects of BdMSC transplantation observed in
these clinical trials, there is an evident lack of long-term clinical trials
(
As BdMSCs and BMCs are both derived from bone marrow, interest lies in
determining which of these cell populations is more efficacious for repair of the
ischaemic heart. A meta-analysis of clinical trials conducted by
Hosseinpour et al. [109] revealed that although both cell types
significantly increase LVEF following MI, BdMSCs were more effective in improving
cardiac contractility. Currently, a registered phase II, randomised,
double-blinded placebo-controlled trial is aiming to assess functional
improvement in VO
ASCs are located within deposits of adipose tissue, where they comprise around
5% of the cell population and are characterised by expression of the same
surface markers as BdMSCs, while also being CD31
Clinical trials investigating the efficacy of ASCs for repair of the ischaemic
heart have recently begun. In 2014, the placebo-controlled double blinded
Adipose-Derived Regenerative Cells in Patients with Ischaemic Cardiomyopathy
(PRECISE) study tested the feasibility of autologous trans-endocardial ASC
administration to patients with IHD [127]. Safety endpoints were followed for 36
months, with no significant differences found between the ASC and placebo groups,
although both wall motion and viable LV mass significantly increased in the
treated group. This study was followed by the Autologous Adipose-Derived
Regenerative Cells for Refractory Chronic Myocardial Ischaemia with Left
Ventricular Dysfunction (ATHENA) trial, which randomised 31 patients to receive
intramyocardial injections of either autologous ASCs or placebo [128]. Results
showed a trend towards improvement in VO
The umbilical cord is a rich source of UCMSCs with the capacity for differentiation into osteogenic, adipogenic and chondrogenic lineages [130, 131]. UCMSCs present an attractive candidate for repair of the ischaemic heart due to their high proliferative potential, with longer telomeres/less cellular aging than other MSCs cell types [132]. Similar to other mesenchymal stromal cells, UCMSCs display low immunogenicity resulting from a lack of human leukocyte antigen DR (HLA-DR), CD80 and CD86 and have strong immunomodulatory and anti-inflammatory properties [133].
Similar to other MSCs, USMSCs exhibit the capacity for in vitro
differentiation into cardiac linaeges [134, 135, 136]. In vivo, the likely
mechanism of cardiac repair exerted by UCMSCs is the release of paracrine
mediators including transforming growth factor beta 3 (TGF-
The Intravenous Infusion of Umbilical Mesenchymal Stem Cells in Patients with Heart Failure (RIMECARD) trial was a phase I/II study investigating the safety and efficacy of allogenic UCMSCs delivered via intravenous infusion to heart failure patients with reduced ejection fraction (HFrEF) (n = 30) [138]. UCMSCs showed no difference in adverse event rate compared to placebo group, and further no alloantibodies were identified. Significant improvements were observed both in LVEF and quality of life at 12 months of follow up. Future randomized trials include the WANICHD trial (NCT04551456), which aims to recruit 300 participants to investigate UCMSCs efficacy as anti-inflammatory agents in coronary artery disease, and the hUC-MSC trial (NCT04939077) (n = 20) that aims to provide further evidence on the safety and effectiveness of UCMSCs in the treatment of heart failure.
All three MSC types (BdMSCs, ASCs and UCMSCs) discussed here share the benefits of low immunogenicity and immunomodulatory properties. BdMSCs and ASCs have an inherent advantage due to the potential for autologous transplantation, while UCMSCs have demonstrated promise owing to their shorter telomeres and comparatively higher proliferative potential. Furthermore, all MSC types are essentially devoid of ethical issues—furthering their potential for clinical application. In order to determine which MSC type is more efficacious for cardiac repair, clinical trials must be conducted comparing the cell types, and in combination with one another.
EPCs exhibit the capacity for both direct endothelial differentiation and maintenance of existing vasculature through paracrine mediated mechanisms [139, 140]. However, substantial debate exists in the literature regarding the classification and origin of EPC sub-populations. Early studies suggested that EPCs originate from the bone marrow, which has recently been challenged [141, 142]. Although the exact developmental origin of EPCs remains unknown, major sources include both peripheral blood and umbilical cord blood [143].
Two distinct EPC populations have been identified and termed ‘early’ and ‘late’ EPCs [140]. Early EPCs refer to cells of a haemopoietic origin, now termed myeloid angiogenic cells (MACs). These cells express CD45, CD14 and CD31 while being negative for CD146 and CD133 [144]. The major mechanism of endothelial repair from MACs is the release of paracrine mediators promoting angiogenesis including VEGF [145]. Late EPCs are now referred to as endothelial colony forming cells (ECFCs) and are considered the ‘true’ endothelial progenitors. ECFCs display an endothelial phenotype, and are identified by their expression of CD146, CD31 and CD105 while being negative for CD45 and CD14 [144]. It is thought that ECFCs primarily exert their beneficial effects on the vasculature by direct differentiation into endothelial cells, profoundly contributing to de novo blood vessel formation and angiogenesis [145, 146]. The heterogeneity of definitions and the lack of a clear unambiguous marker of EPCs has made it increasingly difficult to draw valid conclusions from trials.
In one of the only completed clinical trials investigating EPCs for repair of the ischaemic heart in humans, Zhu et al. [147] studied the safety and efficacy of EPCs pre-treated with thymosin beta-4 in patients with acute ST segment elevation MI. After six months of follow-up, patients treated with EPCs exhibited increased walking distance and significant improvement in cardiac function compared to the control group. Interestingly, despite these positive results, the use of EPC in the clinical setting for IHD has not advanced further. The primary issues holding back the clinical investigation of EPC therapy include the aforementioned lack of clear and consistent phenotypic classification, extended duration of in vitro expansion due to their low occurrence, and high immunogenicity of the cells [139, 144].
The longstanding dogma that adult mammalian heart was traditionally viewed as a
post-mitotic organ with little capacity for self-renewal was challenged in the
early 2000s, when a group of researchers identified a population of cells in the
myocardium of rats exhibiting classical features of stem cells [148]. These cells
were identified as expressing the tyrosine kinase receptor c-kit while being
negative for common haemopoietic lineage markers such as CD34. Although a number
of early studies demonstrated the efficacy of c-kit
In parallel to research focusing on c-kit
In addition to c-kit
The preclinical studies and clinical trials discussed in this review have provided evidence demonstrating the efficacy of stem cells for cardiac repair. However, the nature of this repair has been a focal point in recent years—with a shift from a theory of cell differentiation and remuscularisation towards paracrine mediated repair. This stems from an inconsistent ability to demonstrate differentiation of transplanted stem cells into cardiac linages in vivo, while nonetheless observing beneficial effects on cardiac contractility and coronary artery reserves [153, 169]. Furthermore, long-term improvements in cardiac performance have been observed at time points where very few transplanted cells remain, and where scar tissue separates the resident and transplanted cells [152]. To date, comparatively little is known about the paracrine effects of pluripotent stem cells, with most published research focusing on adult stem cells (MSCs, EPCs, CPCs, BMCs) – of which MSC sare the most extensively studied. The following sections will review our current understanding of the released paracrine factors and their beneficial effects on ischaemic myocardium (Fig. 4).
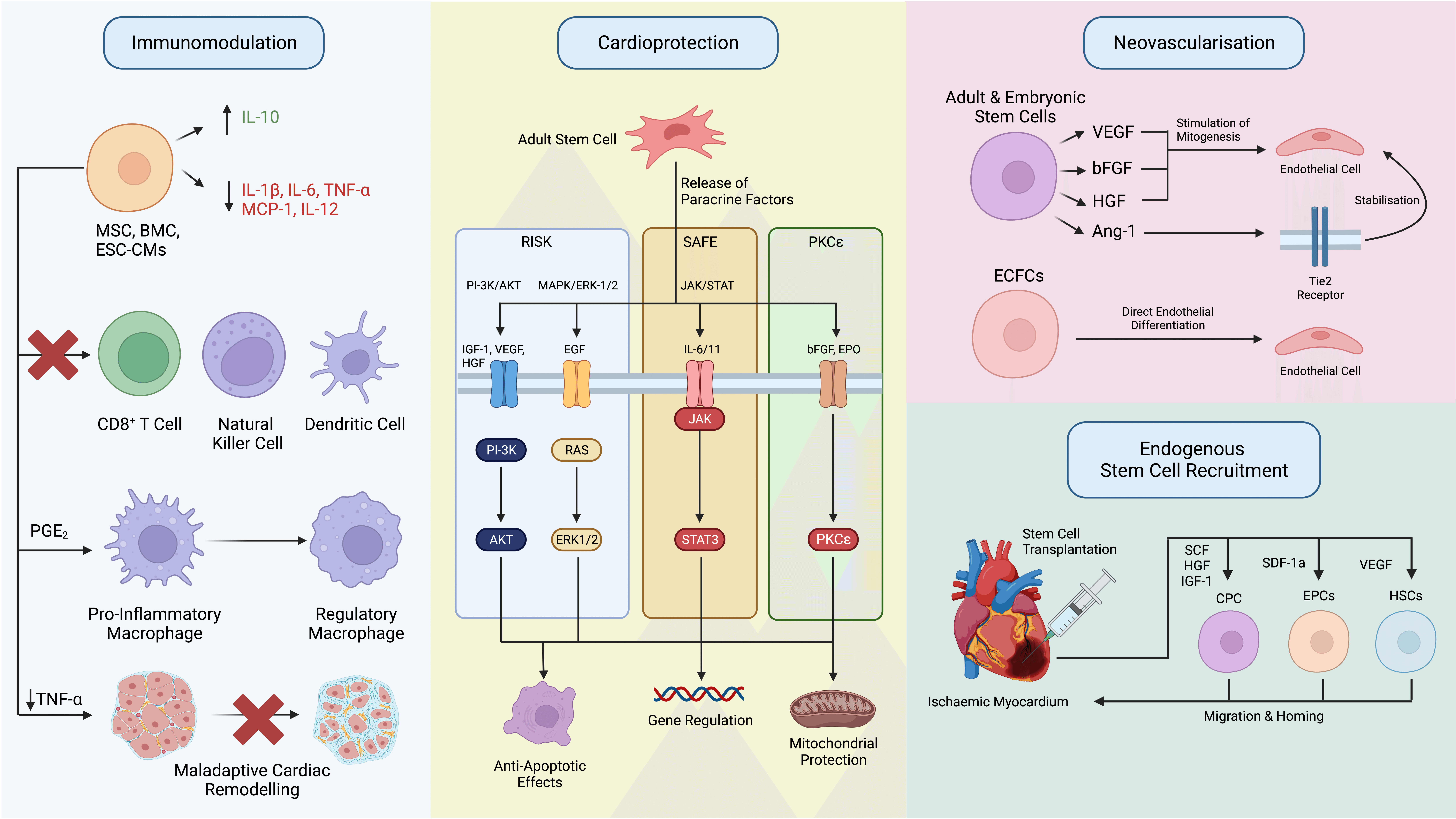 Fig. 4.
Fig. 4.Stem Cell Mechanisms for Cardiac Repair. Summary of the
proposed mechanisms for cardiac repair by transplanted stem cells. Stem cells
secrete various factors with a paracrine effect on other cells promoting
cardioprotection, neovascularisation, immunomodulation and endogenous stem cell
activation, along with an autocrine feedback effect to enhance their survival.
IL-10, interleukin 10; IL-1
While the initial immune response to ischaemic injury is physiologically
essential, a sustained inflammatory response is a direct contributor to adverse
cardiac remodelling and progression to CHF [170, 171]. Both adult stem cells
including MSCs, as well as ESC-CMs, secrete a plethora of anti-inflammatory
cytokines which act to limit deleterious, sustained endogenous inflammation of
the myocardium (Fig. 4). In particular, administration of these cells
downregulates the expression of the pro-inflammatory cytokines tumour necrosis
factor alpha (TNF-
Cardiomyocyte apoptosis is a significant contributor to ischaemic injury and
maladaptive cardiac remodelling [182]. Thus, protecting cardiomyocytes from
apoptosis may attenuate ischaemic injury while promoting their proliferation.
Cultured adult stem cells including BMCs, MSCs and c-kit
Most of the above secreted factors function through the activation of pro-survival kinases Akt and ERK1/2, the downstream signalling cascade of phosphatidylinositol 3-kinase (PI3K) and mitogen-activated protein kinase (MAPK) signalling respectively (PI3K/AKT, MAPK/ERK-1/2), collectively known as the reperfusion injury salvage kinase (RISK) pathway [186, 187, 188, 189]. Acute activation of this pathway mediates cardioprotection, however chronic activation can result in cardiac hypertrophy [190]. A study conducted by Noiseux et al. [169] provided evidence that in rats, activation of the Akt signalling pathway by BdMSCs conferred improved efficacy in enhancing cardiomyocyte survival and preventing apoptosis following myocardial infarction.
Other pathways involved in cardioprotection by stem cells include the surviving
factor enhancement (SAFE) pathway and the protein kinase c epsilon
(PKC
Due to the imbalance between myocardial oxygen demand and supply in IHD,
reperfusion of the ischaemic area is essential for improving clinical prognosis
and halting the progression of IHD. Neovascularisation consists of angiogenesis
(growth of endothelial sprouts from existing vessels), vasculogenesis
(differentiation of angioblasts into endothelial cells) and arteriogenesis
(smooth muscle migration, growth and remodelling) [196]. Secreted paracrine
factors promoting neovascularization from BMCs, MSCs, c-kit
Recent studies have demonstrated the ability of transplanted exogenous stem
cells to activate resident and circulating stem cells, enabling endogenous
cardiac regeneration (Fig. 4) [204]. While the specific paracrine factors
responsible are yet to be identified, Urbanek et al. [205] showed that
c-Met/HGF and IGF-1 receptors expressed by CPCs were able to activate resident
CPCs, forming de novo myocardium in a murine model. In another study
from an independent laboratory, administration of HGF and IGF-1 to CPCs isolated
from a porcine model promoted CPC proliferation, migration and the activation of
downstream signalling pathways including phosphorylation of Akt. Interestingly,
HGF and IGF-1 seem to act synergistically, as the observed effects were far
greater in combination than either alone [206]. When applied to CPCs obtained
from the neonatal rat heart, MSC conditioned medium was able to improve CPC
proliferation and inhibit apoptosis [204]. In addition, MSCs also express bone
morphogenetic proteins (BMPs), Wnt pathway modulators and FGF, all of which are
involved in regulating CPC differentiation and commitment, suggesting these
molecules may contribute to the regeneration of the myocardium through the
activation of endogenous stem cells [207]. Along with activation of CPCs,
circulating stem cells including MSCs, BMCs and HSCs home to ischaemic myocardium
following insult from the bone marrow and circulation [208, 209]. Further,
studies have suggested the ability of transplanted MSCs to recruit circulating
EPCs, c-kit
Transplantation of stem cells into ischaemic myocardium exposes them to a
hypoxic environment, triggering activation of transcription factors which have
pro survival and proliferative potential [213, 214]. A key transcriptional factor
overexpressed in MSCs among others is hypoxia-inducible factor-1
IHD and its various complications remain the leading cause of mortality worldwide. Despite advances in the discovery of novel therapeutics for IHD, no widespread clinical translation has occurred of a treatment able to regenerate ischaemic myocardium, thereby restoring cardiac function. It has become increasingly clear that cell-based therapies primarily exert their beneficial effects within ischaemic myocardium through the release of paracrine mediators, rather than remuscularisation of the heart. As outlined in this review, a number of cell-based therapies have demonstrated great promise in early clinical studies, with others including SkMs no longer investigated due to their adverse effects. To reach clinical translation, there is an immediate need to undertake clinical trials with larger sample sizes, a longer duration of follow up and clear, standardised phenotypic classification of cells.
DT and AYG did the literature search, wrote the first draft of the manuscript and created images. RK conceptualized the idea, developed the outline for the review, critically revised the manuscript and figures for submission in its final form. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript.
Not applicable.
Not applicable.
This study is supported by a project funded by the Health Research Council of New Zealand to RK (Grant #22/632). DT was supported by the Phyllis Paykel Memorial Scholarship and the Department of Physiology, the University of Otago, during his MBChB/PhD study. AYG was supported by the Otago Medical School Scholarship and the Department of Physiology, the University of Otago, during his BMedSc(Hons) study.
The authors declare no conflict of interest. Rajesh Katare is serving as one of the Editorial Board members and Guest editors of this journal. We declare that Rajesh Katare had no involvement in the peer review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Giuseppe Boriani.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.