




 , Mingkui Zhang 2,*
, Mingkui Zhang 2,*1 Department of Vascular Surgery, The First Hospital of Tsinghua University, 100016 Beijing, China
2 Department of Cardiac Surgery, The First Hospital of Tsinghua University, 100016 Beijing, China
3 Department of Cardiovascular Surgery, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, 430022 Wuhan, Hubei, China
† These authors contributed equally.
Academic Editor: Brian Tomlinson
Abstract
The purpose of this study was to explore the pathomechanism of human
myxomatous valve degeneration by investigating changes in the phenotype of
valvular cells, the metabolism of the extracellular matrix and their mechanical
properties. Mitral valve specimens were harvested from patients who had undergone
valve replacement, and divided into two groups: patients with a myxomatous mitral
valve and a control group. Histological investigation showed that the morphology
of the extracellular matrix was looser and less coordinated in myxomatous valves
than in controls.
Keywords
- Myxomatous mitral valve
- Degeneration
- Valvular interstitial cell
- Extracellular matrix
- Mechanical property
Due to continuous improvements in global medical care over many decades, the incidence of rheumatic valvular heart disease has been decreasing year by year. In contrast, myxomatous degenerative heart valve disease has gradually become a serious cardiovascular disease that affects the quality of life and life expectancy of the elderly worldwide [1, 2, 3]. The exact cause of myxomatous degenerative valve disease is still unknown. At present, research on this disease has focused on morphology, while the study of its pathological mechanism at the mechanical and cellular level has received little attention. The aim of this study was to explore the mechanism of valvular degenerative pathological changes by detecting changes in the phenotype of valvular interstitial cells, extracellular matrix (ECM) metabolism, valve structure, and mechanical properties, and therefore provide a theoretical basis for the prevention and treatment of myxomatous degenerative valve disease in the future.
A total 26 of mitral valve specimens (anterior leaflet) were selected from
patients who underwent valve replacement surgery, which were then divided into
two groups: A myxomatous mitral valve group (hereafter referred to as diseased,
n = 20, specimens were obtained from patients with myxomatous degenerative mitral
valve which could not be reconstructed, 12 males and 8 females aged 70.3
Valve specimens were fixed in 10% neutral formalin, embedded in paraffin, and
cut transversely in 5-
Serial 5-
Excessive water on the surface of fragments from the valve specimens was removed
by clean absorbent paper. The specimens were then weighed, ground, and
homogenized in PBS, followed by centrifugation (3000 r/min) for 5 minutes. After
the addition of 500
The surface of the valve specimens was dried with absorbent paper. Specimens were weighed and tested in accordance with the operating instruction of the hydroxyproline reagent cartridge for processing. Elx800uv microplate reader (Bio-Tek, Milton, VT, USA) was used to test the absorbance of samples at 550 nm, and the hydroxyproline content of the samples was calculated.
Valve specimens were cut into 1.5
Valve fragments were placed in an autoclavable mortar and soaked in DEPC, 1
mL of Trizol was added, the fragments were then ground down and transferred to
an Eppendorf tube. After standing at room temperature for 5 minutes, 0.2 mL of
chloroform was added to the sample and shaken vigorously for 15 seconds, then
placed on ice for 5 minutes. Following centrifugation (14000 r/min) at 4
Reverse transcription was carried out according to the instructions of the
reverse transcription kit (Fermentas) and
samples after reverse transcription were stored in a –20
Primer designs are shown in Table 1 and the PCR reaction system is shown in
Table 2. Conditions for PCR amplification were: an initial denaturation at 95
| Primers | Sequence | Length (bp) | Product | Annealing temperature | |
| Length (bp) | |||||
| SMemb (human) | |||||
| Upper primer | 5′-CCGCCCAGAAGAGTGACAATG-3′ | 21 | 376 | 64.2 | |
| Down primer | 5′-TGGAGTTTACGCCGAGATGC-3′ | 20 | |||
| MMP-13 (human) | |||||
| Upper primer | 5′-TGGCTGCCTTCCTCTTCT-3′ | 18 | 235 | 49.3 | |
| Down primer | 5′-TCAAGTTTGCCAGTCACCT-3′ | 19 | |||
| MMP-1 (human) | |||||
| Upper primer | 5′-GAAGAATGATGGGAGGCAAGT-3′ | 21 | 523 | 57.4 | |
| Down primer | 5′-ATGAGCCGCAACACGATG-3′ | 18 | |||
| TIMP-1 (human) | |||||
| Upper primer | 5′-AATTCCGACCTCGTCATCAG-3′ | 20 | 195 | 54.5 | |
| Down primer | 5′-GTTGTGGGACCTGTGGAAGT-3′ | 20 | |||
| Upper primer | 5′-CTCATCACAGGCAAGGAAGAT-3′ | 21 | 410 | 55 | |
| Down primer | 5′-TTAAGGTAAGTGTAGGTTGGG-3′ | 21 | |||
| 10 × PCRbuffer (Mg |
| dNTP mixture (2.5 mM each) 4 µL |
| sense primer (10 pmol/µL) 2 µL |
| antisense primer (10 pmol/µL) 2 µL |
| Taq DNA polymerase (5 U/µL) 0.25 µL |
| dH |
| cDNA 3 µL |
| total volume 50 µL |
| *10 |
5
Statistical data are presented as standard deviations from the mean. Unpaired Student t-test was used to compare means of two experimental groups, and statistical significance was set at the 0.05 level. Statistical analyses were performed with SPSS ver. 23.0 (SPSS Inc., Chicago, IL, USA).
Histological examination of non-myxomatous valve leaflets demonstrated that cells were distributed in all three layers, of an integrated and compact three-tier structure (a fibrous layer, a sponge layer, and a ventricular layer) which was interconnected by dense fibrous tissue. The diseased valve leaflets showed a slight increase in number of cells. The three-tier structure was still intact. However, fibrous tissue was loose and disordered, and the sponge layer was deeper with more voids (Fig. 1).
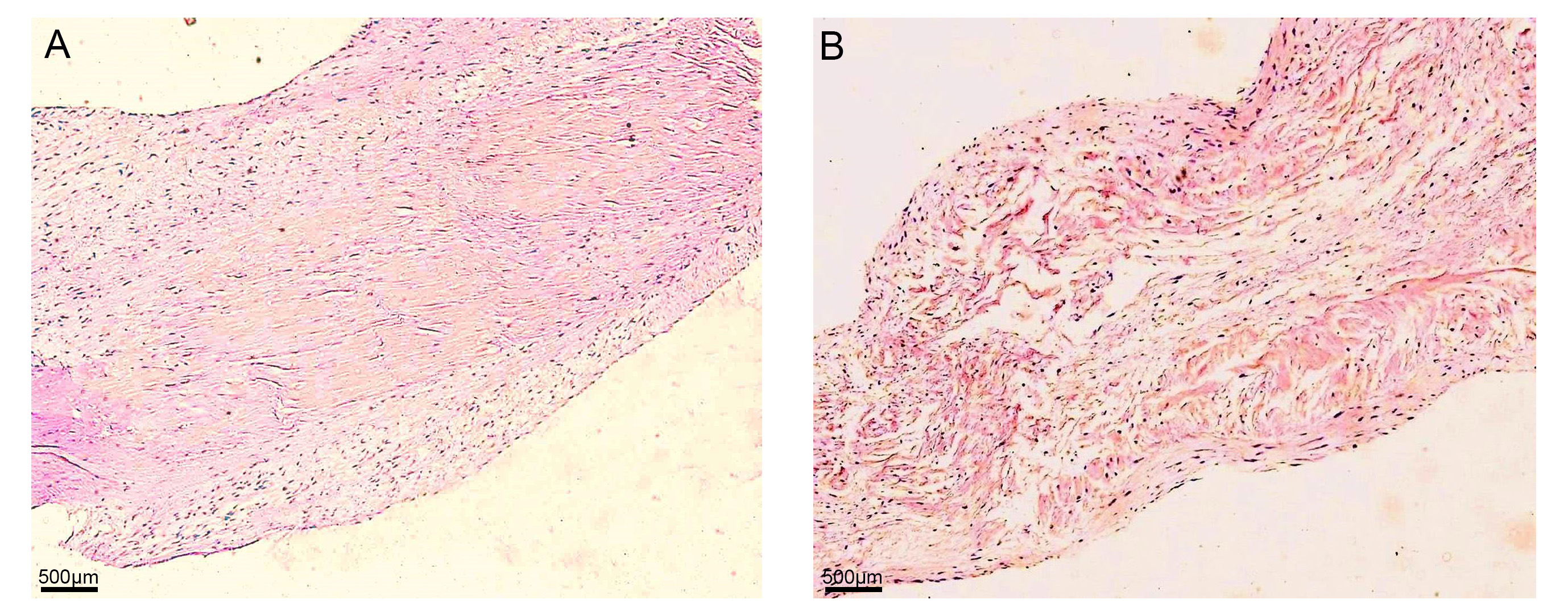 Fig. 1.
Fig. 1.Histologic examination of valve leaflets. (A) Control
group: Cells were distributed in all three layers, of an integrated and compact
three-tier structure (fibrous layer, sponge layer, and ventricular layer) which
was interconnected by dense fibrous tissue. (B) Diseased group: The number of
cells increased slightly. The three-tier structure was still intact, however,
fibrous tissue was loose and disordered, and the sponge layer was broader with
more voids (A and B: hematoxylin and eosin; original magnification
Immunofluorescence for
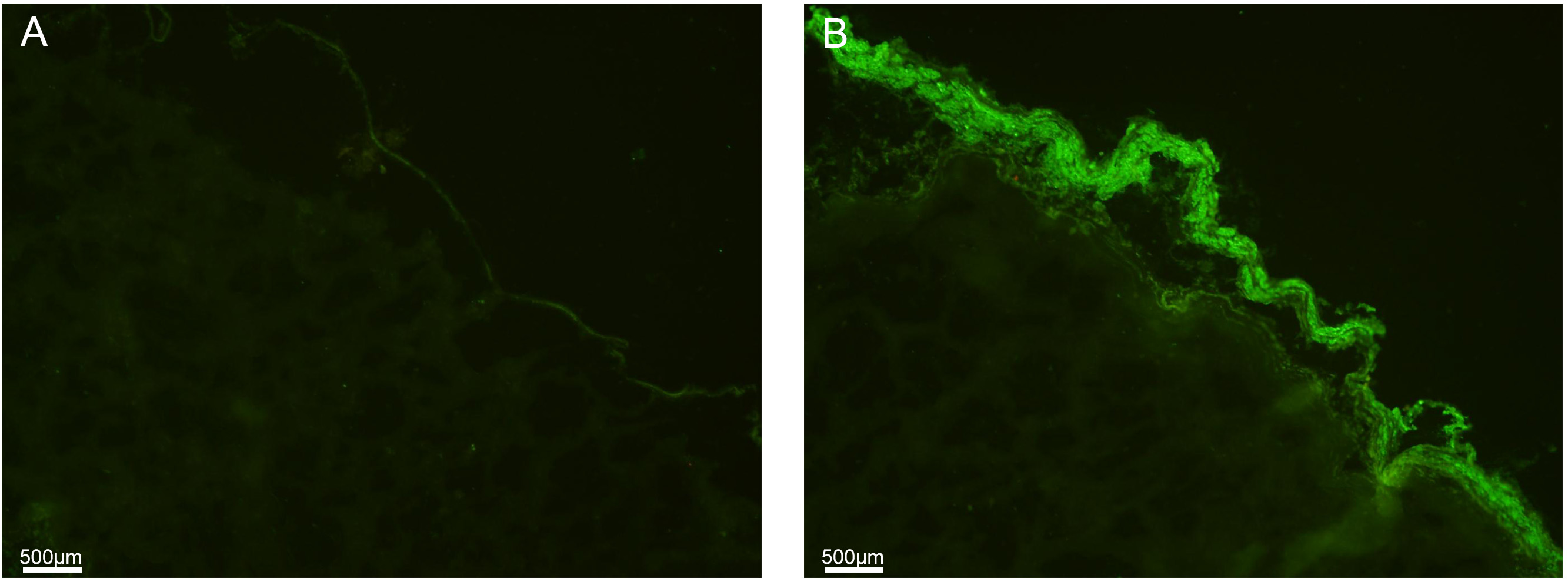 Fig. 2.
Fig. 2.Immunofluorescent staining for
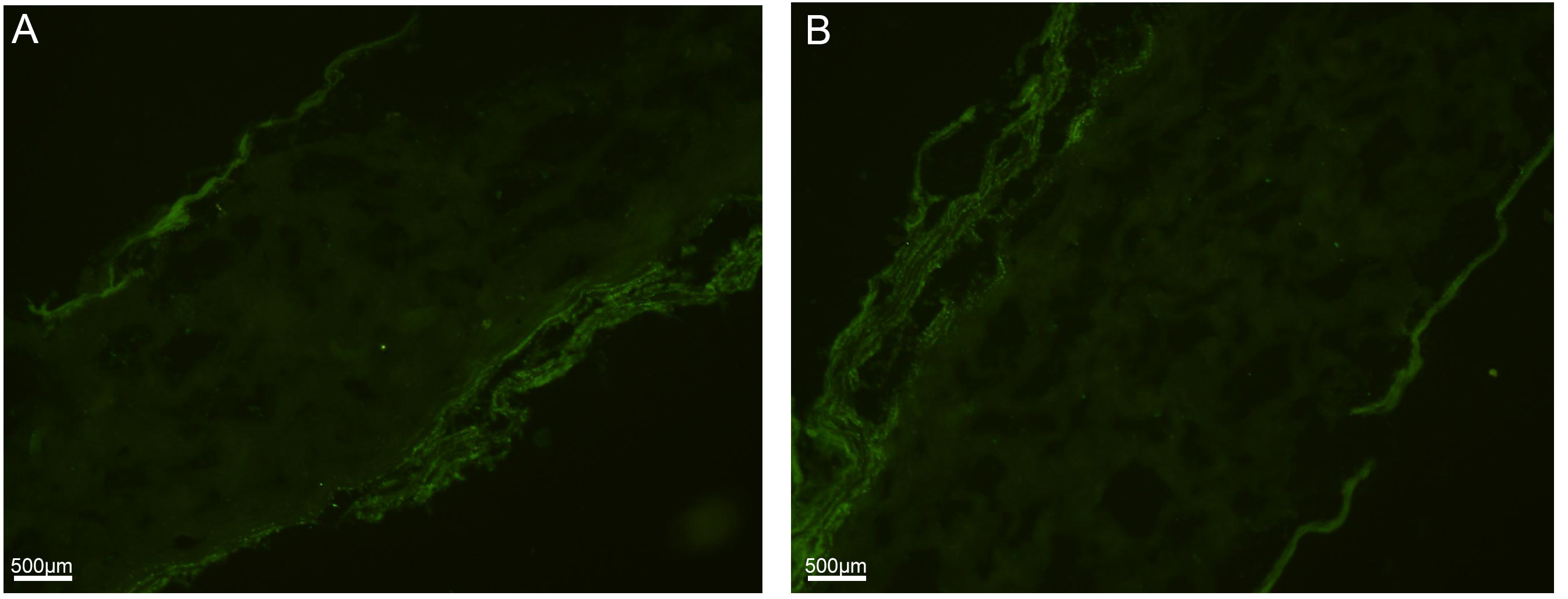 Fig. 3.
Fig. 3.Immunofluorescent staining for Vimentin in the (A) control group and in the (B) diseased group.
A comparison between groups showed that the DNA content of the diseased group was higher than that of the control group, and the hydroxyproline content was lower than that of the control group (Table 3).
| DNA content (µg/mg) | Hydroxyproline content (µg/mg) | |
| Diseased | 0.120 |
4.20 |
| Control | 0.094 |
5.21 |
Compared with the control group, the maximum load and maximum stress of the diseased group decreased, while the maximum strain and elastic modulus increased (Table 4).
| Maximum load ( N ) | Maximum stress (N/mm |
Maximum strain (%) | Modulus of elasticity (N/mm | |
| Diseased | 8.01 |
1.84 |
63.21 |
13.43 |
| Control | 12.92 |
2.55 |
42.55 |
9.49 |
The relative content of the target genes—SMemb, MMP-13, MMP-1 and TIMP-1—and the internal reference in the diseased group and the control group are shown in Table 5. The expression of SMemb and MMP-13 are indicators that reflect the cell phenotype; they were hardly detected in the control group, but highly expressed in the diseased group. The expression of MMP-1 and TIMP-1 mRNA reflect general metabolism in the ECM. Compared with the control group, the expression of MMP-1 mRNA in the diseased group was increased, but the expression of TIMP-1 mRNA decreased. The gel images of the final products of SMemb, MMP-13, MMP-1 and TIMP-1 mRNA detected by RT-PCR are shown in Fig. 4.
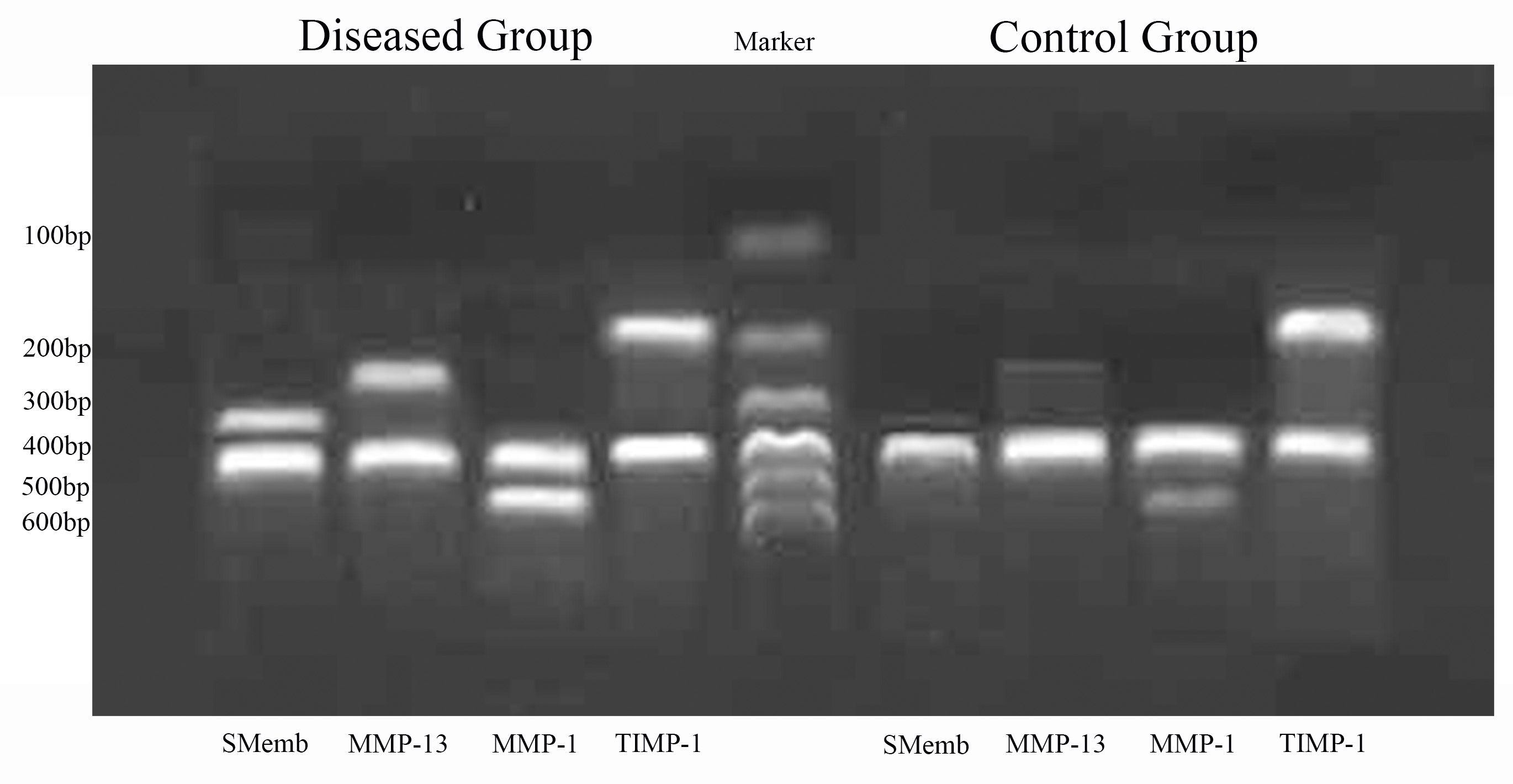 Fig. 4.
Fig. 4.The gel images of the final products of SMemb, MMP-13, MMP-1 and TIMP-1 mRNA detected by reverse transcription-polymerase chain reaction.
| SMemb | MMP-13 | MMP-1 | TIMP-1 | |
| Diseased | 0.55 |
0.47 |
0.71 |
0.70 |
| Control | 0.03 |
0.05 |
0.27 |
0.88 |
VICs are the predominant cells responsible for the synthesis of extracellular
matrix and expression of matrix-degrading enzymes and their inhibitors, which
mediate matrix remodeling [4, 5, 6, 7]. VICs have been demonstrated to exhibit cell
phenotypes in two different states under different conditions, namely
fibroblast-like phenotype and myofibroblast-like phenotype [3, 8]. In normal
adult valves, VICs mainly manifest as a relatively quiescent fibroblast-like
phenotype, while in infancy they exhibit an activated myofibroblast-like
phenotype, both of which express Vimentin, but the latter is characterised by
expression of
Cardiac valves require cells with differing mechanical properties to work together in concert; their strength is closely related to valve structure and chemical composition [20]. The normal heart valve includes a three-tier structure consisting of ventricular layer, spongiosa and fibrous layer, each having a different major constituent (in order: elastin, glycosaminoglycan, and collagen). Different components play differing mechanical roles in the movement of the valve, ensuring normal opening and closing of the valve throughout the cardiac cycle. Elastin permits radial stretch of the leaflets, which allows the valves to extend during diastole and shorten when the heart contracts; glycosaminoglycan shears and cushions impact transmitted from the ventricular layer and the fibrous layer; collagen fibers impart strength and stiffness to the valve, which maintains closure during systole. Clearly, the close relationship of composition-structure-function dictates that changes in the microscopic composition of the valve can lead to changes in the macrostructure, which in turn affects the mechanical properties of the valve and the maintenance of normal function.
The results of this study show that the VICs of non-myxomatous valves
demonstrate a relatively static fibroblast-like phenotype, which may be related
to the low metabolic rate of relatively normal valve tissue. Positive expression
of
In summary, systemic blood flow impacts the valve leading to degeneration over
time, which institutes mechanical stress changes in the valves. VICs are
activated and then proliferate, increasing the expression of
Vimentin,
Compared to the quiescent interstitial cells in non-myxomatous valves, interstitial cells in myxomatous valves exhibit myofibroblast activation and express excessive levels of matrix metalloproteinases. The homeostatic balance between MMP/TIMP is disrupted. We conclude that overactivation of VICs and the imbalance of MMP/TIMP could be important features of the pathomechanism of myxomatous mitral valve degeneration.
ND and MZ designed the research study. CH and QW performed the research. HX and HH provided help and advice on the mechanical and RT-PCR experiments. JS analyzed the data. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript.
All patients involved in this study gave their informed consent. Institutional review board approval of our hospital was obtained for this study (IRB file number: 2020-36).
We appreciate the kind help from Daniel Edward Porter and Whitney Annie Long in polishing this manuscript. Thanks to all reviewers for their opinions and suggestions.
This work was supported by the National Key Research and Development Program of China (2016YFA0101100), National Natural Science Foundation of China (81930052), Tsinghua University Initiative Scientific Research Program (20161080069) and Pilot Fund of the The First Hospital of Tsinghua University (LH-03).
The authors declare no conflict of interest.