




 , Haihua Xie 1, Yingke Ouyang 1, Ting Zhu 2, Qianyan Liu 1, Weiai Liu 3, Shulin Xiong 2,*
, Haihua Xie 1, Yingke Ouyang 1, Ting Zhu 2, Qianyan Liu 1, Weiai Liu 3, Shulin Xiong 2,* , Mi Liu 1,*
, Mi Liu 1,*1 College of Acupuncture & Tuina and Rehabilitation, Hunan University of Chinese Medicine, 410208 Changsha, Hunan, China
2 Department of Preventive Center, The Second Affiliated Hospital of Hunan University of Chinese Medicine, 410010 Changsha, Hunan, China
3 Acupuncture Department, The Second Affiliated Hospital of Hunan University of Chinese Medicine, 410010 Changsha, Hunan, China
Abstract
Mild cognitive impairment (MCI) represents a critical prodromal stage of Alzheimer’s disease. This review synthesizes current evidence to present a coherent pathological cascade driving MCI progression: gut microbiota dysbiosis (e.g., enrichment of Prevotella and depletion of Akkermansia) triggers a butyrate deficit and compromise of intestinal integrity, leading to systemic inflammation. This inflammatory milieu upregulates indoleamine 2,3-dioxygenase 1 (IDO1), shifting tryptophan metabolism toward the kynurenine pathway and resulting in the dominance of neurotoxic branches (3-hydroxykynurenine [3-HK], quinolinic acid [QUIN]) over neuroprotective kynurenic acid (KYNA). This metabolic imbalance promotes N-methyl-D-aspartate (NMDA) receptor-mediated excitotoxicity, oxidative stress, and neuroinflammation, which collectively precipitate synaptic dysfunction and cognitive decline. We explicitly highlight this “gut-immune-metabolic” vicious cycle as the core framework of MCI pathology. Targeting this cycle through a dual strategy—restoring microbial diversity and pharmacologically inhibiting the IDO1/kynurenine 3-monooxygenase (KMO) enzymes—represents a promising therapeutic approach to delay the transition from MCI to dementia.
Graphical Abstract
Keywords
- kynurenine
- mild cognitive impairment
- gut-brain axis
- gut microbiota
- review
Mild cognitive impairment (MCI) represents a critical clinical window between normal ageing and dementia and is associated with a rapidly increasing global burden. A 2025 systematic review involving more than 280,000 participants estimated the global prevalence of MCI in the older population to be 23.7% [1]. The principal challenge underlying this high prevalence is the instability of its clinical prognosis. A large longitudinal cohort study of 3553 individuals with MCI reported an annual progression rate to dementia of 15.7%, with 92.8% of patients eventually converting to dementia over a 17-year follow-up period [2]. Although epidemiological evidence highlights the urgency of early intervention, the core molecular mechanisms driving this transition—particularly the specific cascade linking peripheral metabolism to central pathology—remain incompletely understood. The aetiology of MCI is multifactorial; however, accumulating evidence indicates a strong association between gut microbiota and cognitive [3, 4]. Through the gut-brain axis, intestinal microorganisms are proposed to modulate brain structure and physiological function, thereby influencing host immunity and metabolism that are essential for neurological homeostasis [4]. Multiple studies have demonstrated significant differences in bacterial composition between individuals with MCI and healthy controls [5, 6, 7]. Taxa such as Prevotella, Akkermansia, and members of the Enterobacteriaceae family have been significantly associated with cognitive decline [6, 7]. In addition, microbial metabolites, including short-chain fatty acids (SCFAs) and kynurenine derivatives, have been implicated in the regulation of brain health and cognitive function [8, 9]. This study explores the interplay between gut microbiota and kynurenine metabolism in MCI, examining how microbial dysbiosis may contribute to disease progression through alterations in metabolic pathways.
Recent research has increasingly highlighted a strong association between the gut microbiome and MCI. Multiple microbial strains have been identified that show significant correlations with cognitive function, suggesting a potential role in the onset and progression of cognitive decline and the development of neurodegenerative diseases. As summarised in Table 1 (Ref. [10, 11, 12, 13, 14, 15]), these findings underscore the critical importance of specific microbial compositions in modulating cognitive outcomes in individuals with MCI. This section provides a comprehensive overview of current studies examining the gut microbiome and its mechanistic involvement in MCI, based on evidence synthesised from multiple research sources.
| Reference | Sample | Bacteria strain | Taxonomic level | Strain abundance direction | Impact on MCI/AD |
| Fan et al., 2023 [10] | MCI vs healthy controls | Flavonifractor | Genus | Increased | Associated with executive function decline and inflammation |
| Fan et al., 2023 [10] | MCI vs healthy controls | Lactococcus | Genus | Increased | Potentially compensatory; involved in neurotransmitter synthesis |
| Pan et al., 2021 [11] | MCI vs healthy controls | Staphylococcus intermedius | Species | Increased | Enriched in MCI; possibly neurotoxic oral-derived strain |
| Pan et al., 2021 [11] | MCI vs healthy controls | Leptotrichia buccalis | Species | Decreased | Reduced abundance may reflect mucosal immune dysfunction |
| Gallo et al., 2024 [12] | MCI vs healthy controls | Faecalibacterium prausnitzii | Species | Decreased | Cited from multiple sources; SCFA producer; anti-inflammatory |
| Lu et al., 2025 [13] | MCI vs healthy controls | Allisonella | Genus | Increased | Histamine-related metabolism linked with cognitive dysfunction |
| Lu et al., 2025 [13] | MCI vs healthy controls | Oscillibacter | Genus | Decreased | Potential anti-inflammatory effect; less abundant in MCI |
| Vogt et al., 2017 [14] | AD vs Control | Bacteroidetes | Phylum | Decreased | Associated with increased inflammation and metabolic disruption in AD |
| Vogt et al., 2017 [14] | AD vs Control | Firmicutes | Phylum | Increased | Increased Firmicutes/Bacteroidetes ratio, indicating gut dysbiosis |
| Zhuang et al., 2018 [15] | AD vs Control | Enterococcaceae | Family | Increased | A potential pathobiont may influence inflammation |
MCI, mild cognitive impairment; AD, Alzheimer’s disease; SCFA, short-chain fatty acid.
Zhu and colleagues [16] reported that faecal 16S rRNA gene sequencing revealed significant differences in gut microbiota composition among individuals with MCI, Alzheimer’s disease (AD), and normal controls (NC). The relative abundance of taxa from the Erysipelatoclostridiaceae family, as well as Erysipelotrichales and Saccharimonadales, was significantly increased in patients with MCI, and these taxa were significantly associated with poorer cognitive performance. In addition, patients with MCI exhibited higher levels of Prevotella, which showed a negative correlation with cognitive scores on the Montreal Cognitive Assessment (MoCA) and the Mini-Mental State Examination (MMSE) [16].
Similarly, Khedr et al. [17] demonstrated significant differences between
the gut microbiota of patients with MCI and that of healthy individuals. Patients
with MCI showed increased abundances of Prevotella, Akkermansia, and
Enterobacteria compared with healthy controls. In contrast, levels of beneficial
probiotic microorganisms, including Bifidobacterium and Firmicutes, were reduced
in MCI. Notably, elevated Prevotella abundance was significantly negatively
correlated with cognitive performance (e.g., MoCA scores), suggesting that a high
relative abundance of Prevotella may serve as a potential biomarker for
identifying cognitive deterioration in MCI [17]. Evidence from animal models
further supports the involvement of gut microbiota in cognitive impairment.
Harach and colleagues [18] investigated gut microbial alterations in amyloid
precursor protein/presenilin 1 (APPPS1) transgenic mice and demonstrated marked
disruptions in microbial composition compared with non-transgenic controls.
Significant changes were observed in the abundance of Bacteroidetes, Firmicutes,
and Akkermansia. These microbial alterations were directly correlated with
amyloid-
Collectively, these findings indicate a robust association between alterations in gut microbiota composition and the cognitive deficits observed in MCI. Experimental studies in MCI animal models further demonstrate that modulation or reduction of gut microbiota, through antibiotic treatment or probiotic intervention, can substantially influence cognitive function and neuropathological outcomes.
SCFAs are fatty acids with fewer than six carbon atoms that are primarily produced through the colonic bacterial fermentation of dietary fibre; acetate (C2), propionate (C3), and butyrate (C4) constitute the majority of SCFAs [20]. Butyrate serves as the principal energy source for colonocytes and, together with other SCFAs, exerts biological effects via G-protein-coupled receptors (e.g., free fatty acid receptor 2 [FFAR2], free fatty acid receptor 3 [FFAR3], and G protein-coupled receptor 109A [GPR109A]) and histone deacetylase inhibition, thereby reinforcing epithelial tight junctions and modulating mucosal and systemic immune responses [20, 21]. Through the microbiota–gut–brain axis, microbial metabolites such as SCFAs and tryptophan derivatives influence vagal, endocrine, and immune pathways, regulate blood-brain barrier (BBB) integrity and microglial activity, and affect neurotransmitter systems relevant to cognitive function [21]. Structural alterations in the gut microbiota are accompanied by marked changes in its metabolic activity in patients with MCI. SCFA production is essential for maintaining host immune function, preserving intestinal barrier integrity, and supporting normal nervous system health. Research by Putignani et al. [22] demonstrated that patients with MCI exhibit pronounced alterations in gut microbiota metabolite profiles, particularly in SCFA production. In these patients, overall SCFA metabolism was reduced, with butyric acid showing the most substantial decline compared with healthy controls. Reduced butyrate production may impair intestinal barrier integrity, a condition frequently associated with increased systemic inflammation [22]. Dietary patterns also play a critical role in shaping SCFA production. Studies by Maciejewska et al. [23] indicated that diets high in fat and cholesterol induce marked alterations in gut bacterial metabolic activity. In MCI mouse models, a high-fat and high-cholesterol diet led to abnormal SCFA profiles, characterised by decreased butyric acid levels and increased propionic acid levels [23]. These findings suggest that gut microbial dysbiosis and associated metabolic disturbances may contribute to the progression of MCI pathology.
Further evidence indicates that gut microbial metabolites, including SCFAs, tryptophan derivatives, and lipopolysaccharides (LPS), influence MCI pathogenesis through their effects on neural and immune functions. Research by Kelly et al. [24] showed that the metabolic activity of the gut microbiota differs between individuals with MCI and healthy controls. These microbial metabolites may affect MCI progression by modulating neurotransmitter synthesis and inflammatory responses [24].
Peredo-Lovillo et al. [25] investigated the effects of dietary fibre and prebiotics on the metabolic function of intestinal microbiota. Their findings demonstrated that dietary fibre intake promotes the growth of beneficial intestinal bacteria, enhances microbial metabolic activity, and increases SCFA production. These changes are associated with improved intestinal barrier function and immune responses. Modulation of dietary components may therefore help restore gut microbiota balance and alleviate metabolic dysfunction in patients with MCI [25].
In summary, the metabolic signature of MCI is characterised by a “butyrate gap”, reflecting a critical deficiency in energy substrates for colonocytes that precedes the barrier dysfunction described in the following section.
Metabolic dysfunction and structural imbalance of the gut microbiota not only
compromise intestinal barrier integrity but also profoundly affect key signalling
pathways of the gut–brain axis. Evidence indicates that, when intestinal barrier
function is impaired, pro-inflammatory molecules such as LPS can translocate into
the circulation and activate systemic immune responses [26]. These
pro-inflammatory factors may subsequently access the central nervous system (CNS)
via a compromised BBB, leading to microglial activation. Activated microglia
transition from a resting to a reactive state, releasing substantial quantities
of pro-inflammatory cytokines, including interleukin-1
The vagus nerve acts as a two-way bridge between the gut and the brain.
Microbial products like SCFAs normally activate this nerve to send healthy
signals to the brain. In MCI patients, a lack of SCFAs means the vagus nerve
receives fewer triggers. This drop in signalling leads to lower levels of
protective chemicals like acetylcholine. As a result, the brain loses its
stability and memory gets worse [30]. This disruption impairs central nervous
system stability and may accelerate cognitive decline [26]. The impairment of
this signalling pathway leads to CNS instability, resulting in further cognitive
deterioration [31]. At a molecular level, these disruptions involve endothelial
junctional components that maintain BBB integrity. Tight-junction
proteins—including claudin-5, occludin, and junctional adhesion molecules
(JAM-A/B), scaffolded by zonula occludens-1 and -2 (ZO-1/ZO-2)—and
adherens-junction proteins such as vascular endothelial cadherin (VE-cadherin)
with
The kynurenine pathway (KP) is the principal route for tryptophan catabolism, generating a series of neuroactive metabolites, among which kynurenic acid (KYNA), quinolinic acid (QUIN), and 3-hydroxykynurenine (3-HK) are of particular research interest. These metabolites exert distinct effects on the CNS and contribute to the progression from MCI to AD by modulating neuronal and glial cell functions as well as synaptic plasticity.
KYNA is a neuroprotective metabolite of the KP, primarily synthesised by
astrocytes via kynurenine aminotransferases. Its mechanisms of action in the CNS
are largely defined by its broad antagonism of ionotropic glutamate receptors [38].
KYNA blocks the glutamate/glycine co-agonist site on N-methyl-D-aspartate (NMDA)
receptors, thereby reducing glutamate-mediated excitotoxicity [8]. At elevated
concentrations, KYNA is also reported to antagonise
3-HK is an intermediate metabolite of the KP, synthesised from kynurenine via
kynurenine 3-monooxygenase (KMO) in microglia [44]. It does not act as a
conventional receptor ligand but functions as a redox-active pro-oxidant capable
of spontaneously generating free radicals, such as superoxide anion and hydrogen
peroxide, earning it the designation of a “radical donor”. In the presence of
transition metals, 3-HK participates in Fenton-like reactions to produce hydroxyl
radicals, which can damage proteins and DNA. Its downstream metabolites,
including 3-hydroxyanthranilic acid and QUIN, further propagate oxidative chain
reactions. By inducing oxidative stress, 3-HK damages mitochondria and cell
membranes, triggers cytochrome c release, and promotes apoptosis, establishing it
as a major driver of KP-related neurodegeneration. Beyond direct neuronal injury,
3-HK indirectly affects glial cells and synaptic plasticity. Oxidative
environments can activate microglia into inflammatory phenotypes and suppress
astrocytic support for synapses, ultimately leading to synaptic degradation and
decreased levels of neurotrophic factors, such as BDNF. In vitro studies
have shown that IL-1
In summary, KP metabolites exert differential effects within the CNS and collectively influence the progression from MCI to AD. KYNA is neuroprotective, antagonising NMDA receptors and buffering excitotoxicity and inflammation, particularly in early AD. QUIN promotes neurotoxicity, tau hyperphosphorylation, and inflammatory amplification, often in regions adjacent to amyloid plaques. 3-HK induces oxidative stress and suppresses neurogenesis. These mechanisms indicate that KP dysregulation is not merely a late-stage consequence of AD but contributes throughout the disease course [48]. In conclusion, chronic inflammation in MCI triggers a “metabolic switch” that tends to upregulate the neurotoxic branch (3-HK and QUIN). However, this switch is not absolute across all clinical contexts. The observed variability in 3-HK and QUIN levels underscores the need for a more integrated framework that accounts for individual metabolic differences and disease stages, rather than relying on a simplified model of constant neurotoxic dominance. Concurrently, this toxic shift suppresses hippocampal neurogenesis and downregulates BDNF signalling. The disruption of the neuroprotective balance (e.g., KYNA/QUIN ratio) represents a critical potential mechanism accelerating the transition from MCI to AD. However, future frameworks must integrate the divergent findings reported in systemic vs. central compartments, ensuring that the ‘gut-immune-metabolic’ axis is interpreted as a complex regulatory network rather than a unidirectional pathological pathway.
QUIN is a neurotoxic end-product of the KP, formed downstream of
3-hydroxykynurenine and produced predominantly by activated microglia and
infiltrating macrophages during inflammatory states [39, 50, 51]. Functionally,
QUIN acts as an NMDA-receptor agonist and, at elevated concentrations,
overactivates N-methyl-D-aspartate receptors (NMDARs), provoking excessive
Ca2+ influx, mitochondrial dysfunction, ROS generation, and apoptotic
neuronal death (excitotoxicity) [39, 50]. QUIN also promotes oxidative stress and
lipid peroxidation; formation of Fe(II)-QUIN complexes can catalyse
hydroxyl-radical production, amplifying damage to membranes and proteins [50].
Beyond direct neurotoxicity, QUIN exerts pro-inflammatory actions, activating and
recruiting microglia and astrocytes. Experimentally, QUIN can impair BBB
integrity via nitric-oxide–dependent cytoskeletal perturbations, thereby
increasing BBB permeability [50, 52]. In Alzheimer-related contexts, KP
dysregulation with excess QUIN and upregulated IDO1 has been repeatedly reported.
A
Although endogenous KYNA levels rise in a compensatory attempt to protect neurons, this elevation is often insufficient to counteract the toxic burden and may inadvertently inhibit memory-related synaptic plasticity. Ultimately, the imbalance between escalating neurotoxicity and failing neuroprotection precipitates synaptic loss and cognitive decline, as illustrated in Fig. 1.
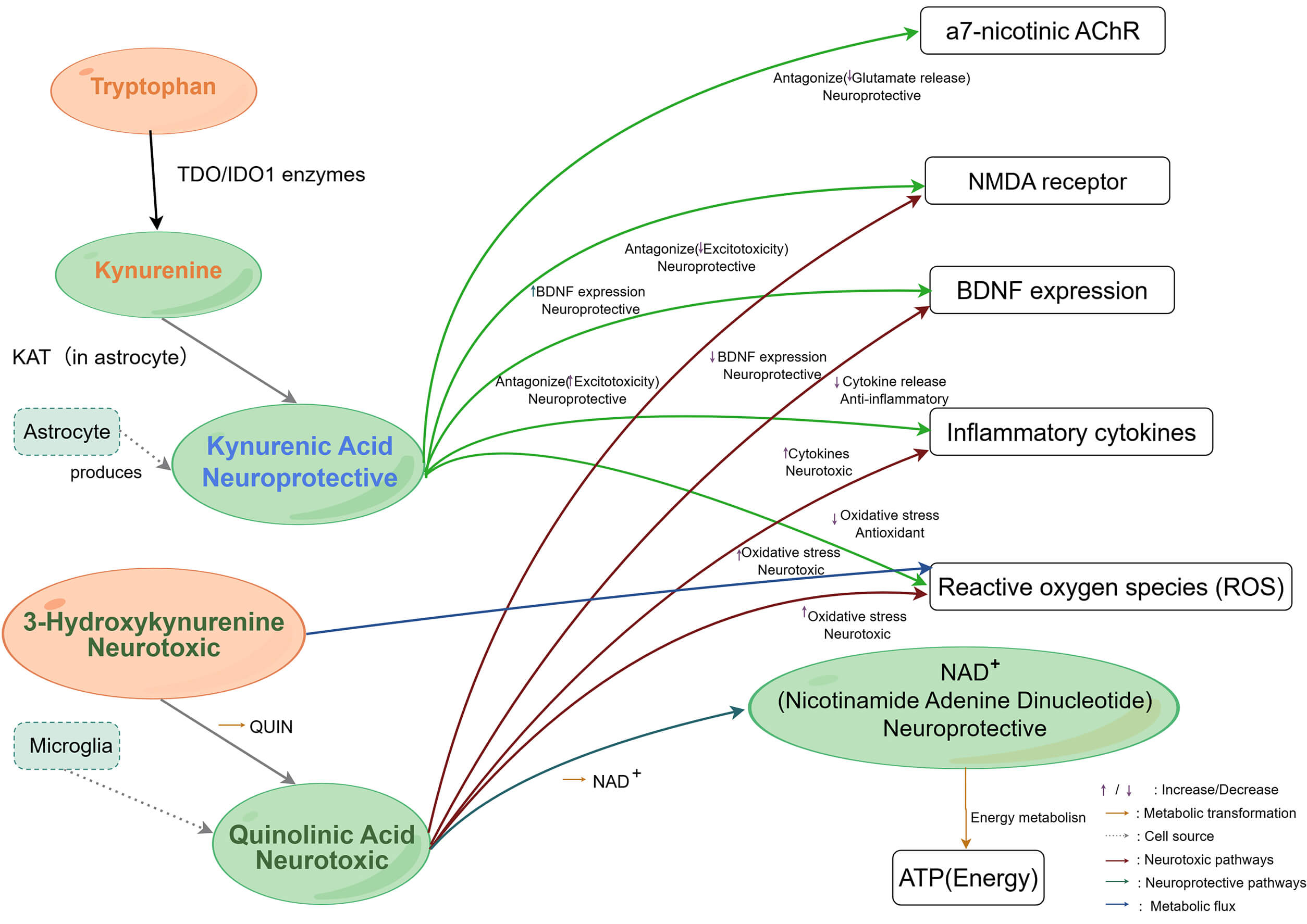 Fig. 1.
Fig. 1.
Illustrates the role of the kynurenine pathway in regulating the
nervous system, immune response, and cellular metabolic homeostasis. Schematic
representation of the kynurenine pathway (KP) metabolism and its downstream
targets in the CNS. Tryptophan is metabolized into kynurenine by IDO/TDO enzymes.
Kynurenine is further metabolized into two distinct branches: The formation of
kynurenic acid (KYNA) in astrocytes, which targets NMDA receptors,
Pharmacological interventions targeting key KP enzymes have progressed from preclinical studies to clinical translation, highlighting their considerable potential as disease-modifying therapies (DMTs) for neurodegenerative diseases. Regarding IDO1 inhibition, a breakthrough study by Minhas et al. [55] in 2024 demonstrated that the highly selective IDO1 inhibitor PF-06840003 penetrates the BBB and significantly reverses memory deficits in AD models by alleviating astrocytic metabolic inhibition and restoring neuronal lactate supply. Furthermore, the IDO pathway modulator Indoximod has exhibited favourable CNS tolerability in clinical trials for paediatric brain tumours, thereby providing a safety rationale for metabolic reprogramming therapies in AD [56].
In the domain of KMO inhibitors, the novel compound KNS366 recently completed its first-in-human Phase I trial in healthy volunteers, successfully validating its ability to reduce levels of the neurotoxic metabolite 3-HK in vivo [57]. This achievement represents a pivotal step toward the clinical application of specific KMO inhibitors. Concurrently, drug repurposing strategies have demonstrated promise; the anti-inflammatory agent Diclofenac, identified as possessing KMO inhibitory activity, is currently undergoing clinical evaluation (NCT06636227) to assess its efficacy in modulating kynurenine metabolism [58]. These clinical advances underscore the therapeutic significance of targeting KP enzymes: precise inhibition of IDO1 or KMO not only halts the production of neurotoxic species but also restores cerebral immune-metabolic homeostasis. This dual mechanistic benefit—simultaneously mitigating neuroinflammation and reinstating energy metabolism—provides robust theoretical support for clinical translation and offers a promising avenue to disrupt the vicious cycle of AD pathology.
This is evidenced by a notable increase in QUIN levels and a decrease in KYNA, an imbalance that is hypothesised to further exacerbate nervous system compromise. Clinical data have demonstrated that higher plasma QUIN and lower KYNA levels in patients with MCI correlate with reduced cognitive function scores. Furthermore, blood levels of kynurenine, rather than tryptophan (1/6), associated with higher kynurenine-to-tryptophan ratios (1/3), are considered early biomarkers of MCI [59, 60]. These metabolic alterations are closely related to the pathological progression of MCI.
As detailed in Section 3.2, the dysregulation of KYNA and QUIN disrupts synaptic homeostasis and NMDA receptor signalling, leading to the following pathological consequences in MCI. Crucially, QUIN amplifies the microglial activation cascade initiated by LPS (as detailed in Section 2.3). Unlike the initial immune response, QUIN-induced microglial activation is self-perpetuating, locking microglia in a neurotoxic phenotype that further impairs synaptic transmission [61].
This mechanism has been validated in animal models. IDO1 induction by inflammatory factors in mice increased QUIN levels, leading to cognitive impairment and hippocampal neuronal damage [39]. However, significant amelioration of kynurenine metabolic dysregulation, attenuation of neuroinflammation, neuronal protection, and substantial enhancement of cognitive function can be achieved through probiotic and short-chain fatty acid supplementation, which suppresses IDO1 overexpression [62]. These experimental results have enhanced our understanding of the role of kynurenine metabolism in the transition from MCI.
Together, decreased KYNA and increased QUIN are hypothesised to promote a self-perpetuating pathological cycle. The loss of KYNA’s protective effects, combined with QUIN’s neurotoxicity, disrupts neurotransmitter balance, calcium ion homeostasis, and synaptic function, resulting in further cognitive decline [63]. Clinical and experimental evidence indicates that dysregulation of the kynurenine pathway contributes substantially to the pathological basis of MCI and that excessive metabolic abnormalities accelerate cognitive deterioration [64].
In conclusion, the balance of the kynurenine metabolism pathway can be considered a core feature of the pathological progression in patients with MCI. This imbalance is characterised by decreased KYNA levels, which weaken neuroprotective functions, and increased QUIN levels, which exacerbate neurotoxicity and inflammatory responses. As illustrated in Fig. 2.
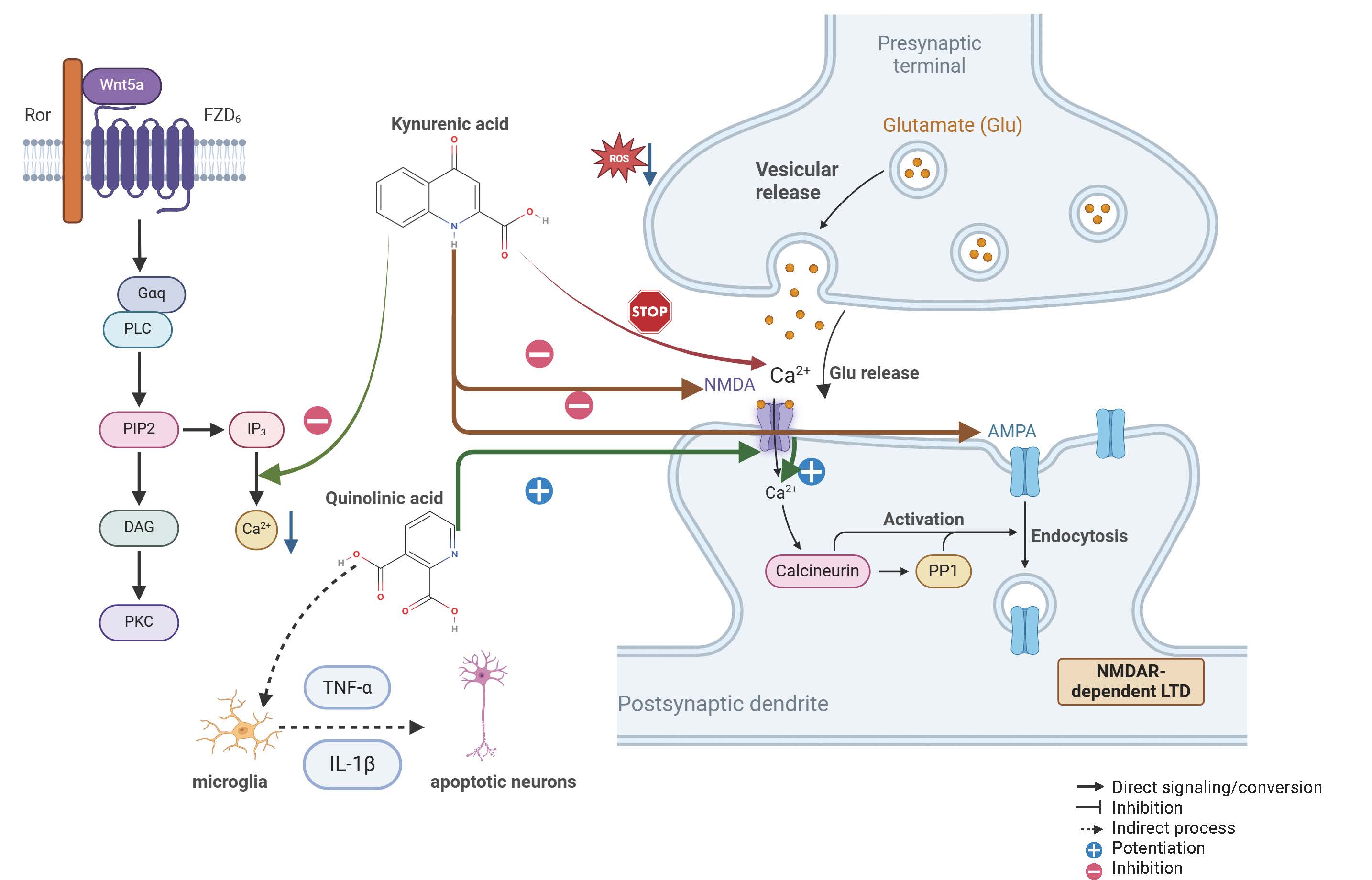 Fig. 2.
Fig. 2.
Mechanisms of kynurenine metabolites in the nervous system:
neuroprotective and neurotoxic effects. KYNA functions as a crucial maintainer
of synaptic homeostasis. Mechanistically, it acts as a broad-spectrum antagonist
at ionotropic glutamate receptors, specifically blocking the glycine site of NMDA
receptors and antagonizing AMPA receptors. This blockade effectively inhibits
excessive calcium influx (Ca2+) and attenuates glutamate-mediated
excitotoxicity. Furthermore, as depicted in the signalling pathway on the left
side of Fig. 2, the Wnt5a-FZD6-Ror (Wnt5a stands for Wingless-type MMTV integration site family, member 5A. FZD6 stands for Frizzled Class Receptor 6. Ror refers to Receptor Tyrosine Kinase Like Orphan Receptor) axis plays a regulatory role in stabilizing
intracellular calcium levels and maintaining synaptic plasticity (LTD). Under
normal conditions, these pathways collectively suppress ROS generation and
protect neurons from oxidative stress. Gaq, G protein subunit alpha q; PLC,
Phospholipase C; PIP2, Phosphatidylinositol 4,5-bisphosphate; IP3, Inositol
trisphosphate; DAG, Diacylglycerol; PKC, Protein Kinase C; TNF-
Building on the KP biochemistry described in Section 3.1, the metabolic fate of tryptophan is further governed by the composition of the intestinal microbiota. Under healthy conditions, beneficial bacteria expressing tryptophanase (TnaA), such as certain Lactobacillus and Bacteroides strains, metabolise a proportion of tryptophan into indole and indolepropionic acid (IPA). These metabolites exhibit antioxidant and anti-inflammatory properties, which reduce the entry of tryptophan into the kynurenine pathway, thereby decreasing QUIN production and mitigating its neurotoxicity [65]. However, under conditions of intestinal dysbiosis, the abundance of pro-inflammatory bacteria, such as Escherichia coli, increases. This shift promotes IDO1 expression, driving a greater influx of tryptophan into the kynurenine pathway [66]. Consequently, QUIN levels rise substantially, while KYNA levels decline markedly, further disrupting the dynamic equilibrium of tryptophan metabolism [67].
In addition to direct metabolic effects, intestinal dysbiosis indirectly
regulates tryptophan pathway selection through inflammatory signalling. When
intestinal barrier integrity is compromised, inflammatory mediators such as LPS
enter the circulation and induce the release of pro-inflammatory cytokines,
including IL-6 and Interferon-gamma (IFN-
Building on the metabolic deficits described in Section 2.3, depletion of SCFAs
leads to a specific loss of regulatory control over the kynurenine pathway.
Beyond their general anti-inflammatory effects, SCFAs act as histone deacetylase
(HDAC) inhibitors that directly suppress the transcriptional activation of IDO1.
SCFAs do more than just fight inflammation directly. They also help the brain
control the gut through the vagus nerve. This process is called the “cholinergic
anti-inflammatory reflex”. The brain sends signals back down the vagus nerve to
stop gut cells from making too many inflammatory toxins. This reflex keeps the
gut wall strong and prevents leaks. When this system fails in MCI patients, gut
inflammation gets out of control and harms the brain further [70].
Mechanistically, SCFAs, particularly butyrate, function as potent HDAC
inhibitors. By modulating the epigenetic landscape of immune cells, they suppress
the signal transducer and activator of transcription 1 (STAT1) signalling cascade
required for IDO1 transcription. Consequently, SCFAs not only attenuate
inflammation but also directly inhibit the genomic mechanisms that drive
excessive tryptophan flux into the kynurenine pathway, maintaining IDO1 in a
downregulated state. In addition, butyric acid reduces the expression of
inflammatory genes by inhibiting the nuclear factor kappa-light-chain-enhancer of
activated B cells (NF-
SCFAs also regulate peripheral immune responses and directly influence the central nervous system by crossing the BBB. Through these mechanisms, BDNF activation, neuronal repair, and synaptic plasticity are promoted, while QUIN production is reduced and neurotoxicity is alleviated [71]. Furthermore, SCFAs enhance KYNA synthesis, restoring its antagonistic effects on NMDA receptors, reducing excitotoxic neuronal damage, and preserving synaptic function [39]. Through these combined actions, SCFAs mediate communication between the gastrointestinal tract and the central nervous system. Ultimately, their coordinated roles in suppressing immune activation and protecting neuronal survival support the maintenance of kynurenine metabolic homeostasis [61].
The systemic LPS burden resulting from intestinal barrier dysfunction (as established in Section 2.3) acts as a potent upstream regulator of tryptophan metabolism. Specifically, circulating LPS stimulates extrahepatic IDO1 expression [62]. Consistently, in 2,4,6-trinitrobenzenesulfonic acid (TNBS)-induced colitis, acupuncture and moxibustion restored colonic architecture and reduced mucosal inflammatory infiltration while rebalancing gut microbiota and associated metabolites, suggesting a protective effect on the intestinal barrier [72]. Furthermore, LPS translocation is capable of stimulating IDO1 expression, shifting tryptophan catabolism towards the kynurenine pathway and markedly increasing QUIN production [73]. Evidence indicates that intestinal barrier dysfunction is widespread in individuals with MCI, where it both exacerbates systemic inflammation and further amplifies inflammatory responses within the central nervous system [71].
Under this pathological condition, beneficial bacteria such as Akkermansia can restore intestinal epithelial integrity by producing mucin, thereby reducing inflammatory infiltration and excessive IDO1 activation [74]. In addition, improvement of intestinal barrier function may decrease the release of LPS-induced pro-inflammatory factors, thereby fundamentally alleviating metabolic disturbances associated with the kynurenine pathway [61].
A dynamic competitive relationship exists between the kynurenine metabolic pathway and neurotransmitter balance, with the gut microbiota playing a critical regulatory role. When kynurenine pathway activity is excessively elevated, a substantial proportion of tryptophan is diverted towards the production of QUIN and KYNA, leading to a marked reduction in 5-hydroxytryptamine (5-HT) synthesis [75]. Reduced 5-HT levels directly affect emotional regulation, sleep quality, and cognitive function, which are characteristic features of MCI [76].
Indole metabolites and SCFAs generated by intestinal microbial metabolism
promote serotonin (5-HT) synthesis by stimulating enterochromaffin cells, thereby
alleviating excessive tryptophan consumption by the kynurenine pathway [77].
Serum metabolomics from an electroacupuncture clinical study further indicate
normalisation of amino acid metabolism with rebalancing of glutamatergic and
This dynamic competitive interaction indicates that gut microbiota regulation of tryptophan metabolism not only influences kynurenine pathway activity but also substantially affects central nervous system function by altering neurotransmitter synthesis and metabolism [81]. The gut microbiota appears to modulate the activity and branch selection of the kynurenine metabolic pathway by influencing tryptophan metabolism, inflammatory signalling, SCFA production, and intestinal barrier integrity [82]. Under physiological conditions, these regulatory mechanisms maintain a balance between KYNA and QUIN, thereby preserving nervous system function. However, gut microbiota dysbiosis may contribute to excessive activation of the kynurenine pathway, disrupt neurotransmitter balance, and exacerbate neurotoxicity and inflammatory responses, ultimately promoting the pathological progression of MCI [83].
The gut microbiota, as a core regulator of host metabolism and immunity, interacts with the central nervous system through the gut–brain axis and plays a crucial role in cognitive impairment disorders. In recent years, accumulating evidence has demonstrated that gut microbiota imbalance can significantly influence the pathological progression of MCI through the regulation of tryptophan metabolism, inflammatory responses, neurotransmitter synthesis, and related pathways. The following analysis focuses on three aspects: the molecular mechanisms by which gut microbiota imbalance drives MCI, the dynamic balance of endogenous regulatory networks, and the potential for clinical translation.
MCI pathology emerges not as a localized brain event but as a systemic failure
of regulatory containment, where the KP serves as a transductive hub converting
peripheral microbial signals into central neurotoxic cascades [84]. This systemic
integration is highly dependent on clinical subtypes: Amnestic MCI (aMCI)
exhibits a more severe microbial and metabolic dysregulation profile than
non-amnestic MCI (naMCI), with aMCI specifically linked to the enrichment of
Alistipes indistinctus and the depletion of protective Bacteroides eggerthii
[85]. These aMCI-specific taxonomic shifts correlate directly with neurovascular
dysfunction, including decreased cerebral blood flow (CBF) and arterial transit
time (ATT) [86]. At the molecular level, the resulting accumulation of QUIN acts
as a potent NMDA receptor agonist, triggering excessive intracellular calcium
Ca2+ influx and activating the protein kinase glycogen synthase kinase-3
beta (GSK-3
Intestinal flora imbalance is a prominent feature observed in the early stages of MCI and frequently represents the initiation point of a detrimental pathological cycle. External factors can alter the relative abundance of symbiotic and pathogenic bacteria within the intestinal microbiota, thereby predisposing individuals to pathological states. Endogenous regulatory mechanisms may mitigate the adverse effects of dysbiosis and limit the amplification of inflammatory cascades, potentially slowing the pathological progression of MCI.
Evidence indicates that symbiotic bacteria actively inhibit the proliferation of
conditionally pathogenic bacteria and enhance intestinal microenvironmental
stability through metabolic by-products and competitive metabolic mechanisms
[90]. SCFAs enhance the competitive advantage of probiotics within their
ecological niche by lowering intestinal pH, thereby promoting restoration of
microbiota diversity and reducing intestinal inflammation [91]. In addition,
SCFAs directly inhibit NF-
Concurrently, the host immune system plays a pivotal role in this regulatory
process. Protective immune regulation mediated by gut-associated lymphoid tissue
(GALT), including the secretion of anti-inflammatory cytokines such as IL-10 and
transforming growth factor-beta (TGF-
At this stage, recovery of intestinal microbiota diversity and local suppression of inflammatory responses form a mutually reinforcing virtuous cycle. Restoration of SCFA production enhances anti-inflammatory immune responses, which further restrict systemic inflammatory dissemination and provide a stable microenvironment conducive to intestinal flora recovery [95].
As inflammatory responses progressively subside, the body further restricts the expansion of the vicious cycle through coordinated metabolic and neuroprotective mechanisms. Within the kynurenine metabolic pathway, although IDO1 activation promotes increased quinolinic acid production, KYNA is concurrently generated. As a neuroprotective metabolite, KYNA effectively mitigates neurotoxicity through antagonism of NMDA receptor activation [96, 97]. In addition, KYNA suppresses excessive activation of microglia and astrocytes, thereby attenuating cumulative neuroinflammatory effects [98].
With declining levels of pro-inflammatory mediators, tryptophan metabolism gradually shifts from the kynurenine pathway towards serotonin (5-HT) synthesis. Restoration of 5-HT not only alleviates emotional disturbances and sleep disorders in patients with MCI but also directly enhances cognitive function by improving synaptic plasticity and neural network connectivity [61]. This metabolic rebalancing signifies a transition from a vicious pathological cycle to a beneficial regulatory cycle.
At the level of the gut-brain axis, BBB integrity begins to recover. Following attenuation of systemic inflammation, epithelial and endothelial cells reactivate tight junction protein synthesis, thereby reducing the translocation of toxic metabolites and pro-inflammatory factors across the blood–brain barrier [99, 100].
In summary, the pathological cascade of “intestinal flora imbalance
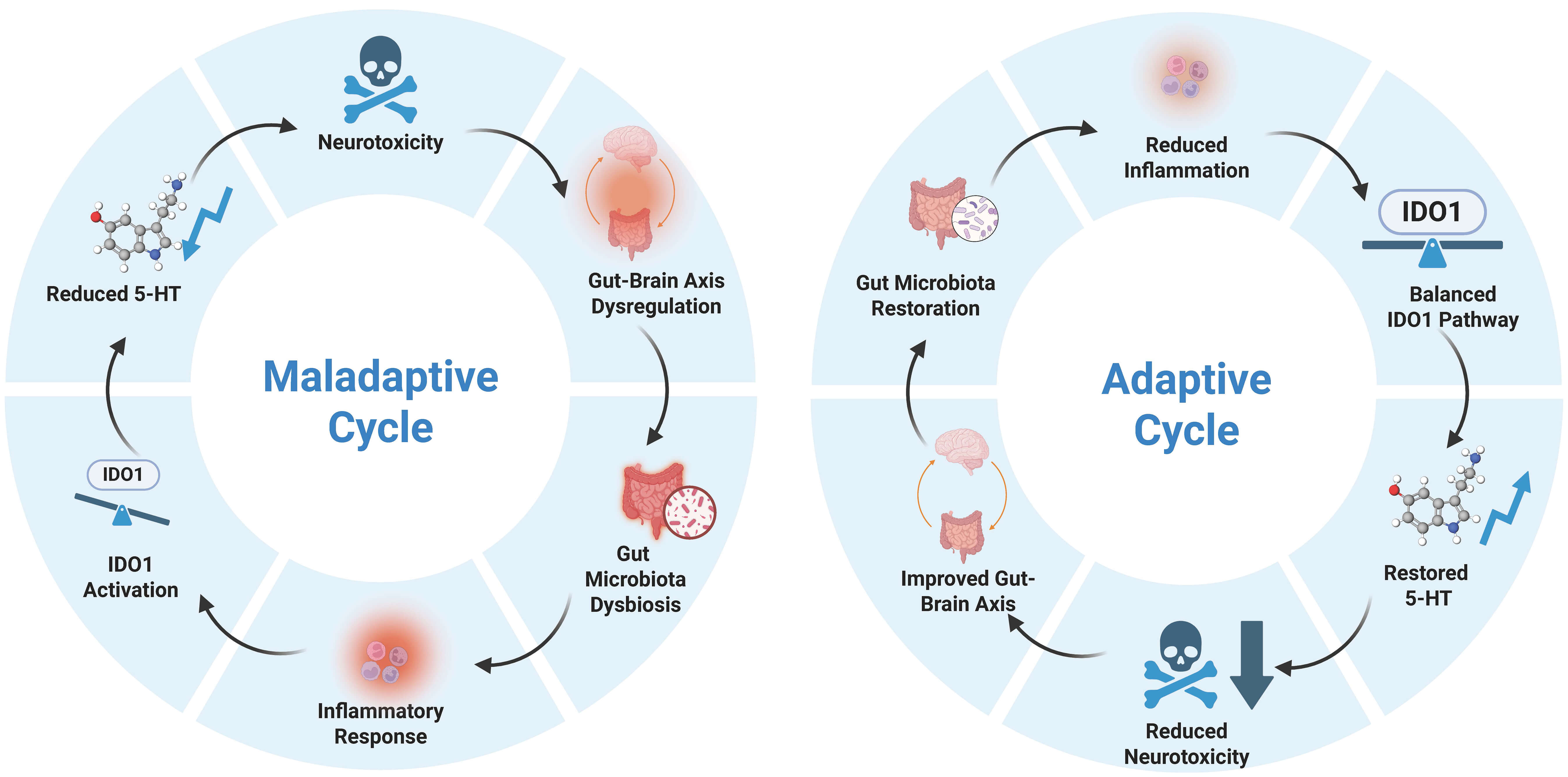 Fig. 3.
Fig. 3.
Self-regulatory mechanisms of the gut-brain axis.Transition from vicious to beneficial cycles and their impact on MCI pathology. The arrows in the figure illustrate the causal progression and dynamic feedback loops within the gut-brain axis. Specifically, the clockwise arrows represent a self-perpetuating cycle where each pathological or physiological component reinforces the next, ultimately maintaining either a maladaptive (left) or adaptive (right) state. 5-HT, 5-hydroxytryptamine. Created with Biorender (https://www.biorender.com/).
This review highlights the significant association between gut microbiota and MCI, particularly the link between gut microbiota alterations and cognitive deterioration mediated through kynurenine metabolism. First, a distinct structural signature of gut microbiota alteration is evident in patients with MCI, characterised by an increased abundance of pro-inflammatory bacteria (Prevotella, Enterobacteriaceae) and a reduction in beneficial probiotics (Bifidobacterium). Concurrently, the metabolic functions of the gut microbiota undergo substantial changes, most notably in the production of SCFAs. These metabolites play a critical role in maintaining intestinal barrier integrity, immune homeostasis, and nervous system health.
Patients with MCI exhibit marked metabolic disturbances in kynurenine metabolism. In particular, a functional imbalance characterised by an increased QUIN/KYNA ratio—reflecting a shift towards neurotoxicity irrespective of absolute metabolite fluctuations—may exacerbate nervous system damage. QUIN excessively activates NMDA receptors, thereby inducing neuroinflammation and oxidative stress, which further accelerates cognitive decline. Conversely, reduced KYNA levels weaken neuroprotective mechanisms and diminish the nervous system’s capacity to counteract excitotoxicity.
Accordingly, our perspective on future therapeutic strategies extends beyond single-agent interventions and instead proposes the establishment of a multi-level, multi-target “virtuous cycle” intervention model. Central to this approach is the restoration of gut microbiota diversity and SCFA production capacity (e.g., through supplementation with Akkermansia and Faecalibacterium) to repair the intestinal barrier and suppress upstream inflammatory drivers. In parallel, pharmacological interventions are applied to precisely modulate key KP enzymes (specifically targeting IDO1 and KMO), thereby correcting downstream metabolic imbalances and re-establishing a neuroprotective KYNA/QUIN ratio.
To inform future experimental design and facilitate clinical translation, this review systematically synthesises key therapeutic targets, their underlying pathological mechanisms, and specific recommendations for future research priorities. Table 2 comprehensively outlines a spectrum of potential intervention strategies, ranging from ecological remodelling of the gut microbiota to the precise modulation of metabolic enzymes. This integrated summary aims to provide a clear operational framework for addressing the pathological imbalances of MCI through synergistic multi-target interventions, thereby delaying or reversing disease progression and ultimately establishing a neuroprotection-centred “virtuous cycle”.
| Therapeutic target | Mechanistic basis | Key recommendations for future research |
| Gut Microbiota | Dysbiosis (Prevotella, Akkermansia) leads to barrier impairment. | Develop next-generation probiotics targeting Akkermansia and Faecalibacterium; explore precision FMT donor screening based on metabolic phenotypes. |
| SCFAs | “Butyrate Gap” leads to insufficient HDAC inhibition and IDO1 upregulation. | Validate epigenetic inhibition mechanisms of butyrate on IDO1/KMO; develop colon-targeted SCFA formulations to repair the blood-brain barrier. |
| IDO1 Enzyme | Inflammation (LPS/Cytokines) induces overexpression, hijacking tryptophan metabolism. | Develop peripheral/gut-restricted IDO1 modulators; investigate synergistic effects of tryptophan dietary intervention and enzyme inhibitors. |
| KMO Enzyme | Key “switch” directing metabolic flow to the toxic branch (QUIN). | Develop BBB-permeable KMO inhibitors to reshape the QUIN/KYNA ratio and enhance endogenous neuroprotection. |
| Comprehensive Intervention | Multi-link interactions of the gut-brain axis. | Conduct clinical trials for “Microbiota Regulation + Enzyme Inhibition” combination therapy; establish MCI early warning and efficacy evaluation systems based on metabolomics. |
KMO, kynurenine 3-monooxygenase; LPS, lipopolysaccharides; FMT, fecal microbiota transplantation; BBB, blood-brain barrier; HDAC, histone deacetylase.
Step 1: Restoring Gut Homeostasis. Intervention must begin at the source in the gut. By supplementing probiotics and dietary fiber, we can fill the “butyrate gap”. Butyrate does more than just repair the intestinal barrier. It also acts as a key regulator to inhibit IDO1 enzyme activity at the genetic level. This reduces the production of harmful metabolites from the start.
Step 2: Maintaining Blood-Brain Barrier Function. SCFAs produced by gut bacteria are vital for brain health. These metabolites strengthen the integrity of the blood-brain barrier. This protection effectively blocks peripheral inflammatory factors from entering the central nervous system. It keeps neurons safe from external disturbances.
Step 3: Correcting Cerebral Metabolic Pathways. Inside the brain, we need to precisely downregulate IDO1 or KMO enzymes. Doing this redistributes the tryptophan metabolic flux. This not only reduces the buildup of neurotoxins (QUIN) but also boosts levels of protective substances (KYNA). This keeps the normal functions of neural synapses stable.
Formulation: JZ, HX, SX, ML, QL, and WL; Literature management: JZ, HX, and YO; Supervision: HX, ML, and SX; Drawing Figures: JZ, HX, and TZ; Initial draft: JZ, YO, WL and HX; Revision and proofreading: JZ, HX, QL, WL, ML, and YO. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
We sincerely thank BioRender and FigDraw, two powerful graphic design software programs, for their important support in creating the illustrations and charts for my thesis.
This work was supported by the projects of the science and technology innovation Program of Hunan Province (No. 2024JK2132, No. 2024RC1061), the Scientific Research Project of Hunan Provincial Health Commission (No. 20257637), the State Administration of Traditional Chinese Medicine 2022 Youth Qihuang Scholars Training Program (National Letter of Traditional Chinese Medicine Education [2022] 256), the Hunan Provincial Graduate Joint Cultivation Base for Acupuncture-Moxibustion and Tuina of Hunan University of Chinese Medicine (No. [2022] 357, Hunan Provincial Department of Education Notice), the Acupuncture Bioinformation and Smart Wellness Innovation and Entrepreneurship Education Center of Hunan University of Chinese Medicine (No. [2021] 356, Hunan Provincial Department of Education Notice), and the Innovative Graduate Cultivation Base for Chinese Medicine Sub-health of Hunan University of Chinese Medicine (No. [2020] 19, University Administrative Research Document).
The authors declare no conflicts of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.