




1 Chair of Zoology, TUM School of Life Sciences, Technical University of Munich, 85354 Freising, Germany
Abstract
How do neural circuits change to incorporate newly learned events? After decades of research, we now have a good understanding of the diversity and duration of memory types, the context-specifics of learning regimes, and the brain areas that are likely to be involved. However, we are still far from a mechanistic understanding of the neural activity required to transform new behavioral experiences into long term memory accessed during recall events. What kind of network activity is required to affect these wide-spread changes in neuronal circuitries? Among current theories of memory in mammals, one of the most intriguing is concerned with the role of large-scale synchronous neural activity, which is thought to enable the transfer of information from the hippocampus to numerous regions of the neocortex. In this contribution, we first review sleep, learning, and memory in birds before highlighting evidence that large-scale synchronous neural activity also exists in the avian brain, making the case that by examining these questions from a comparative research perspective, we gain important insight into the canonical features of memory consolidation.
Keywords
- sleep
- birds
- memory
- memory consolidation
- learning
- hippocampus
Given the importance of sleep to animal life, it is surprising how little we know about its function. Although the scientific study of sleep began in the mid-19th century [1], research was limited to humans and other mammals for over one hundred years [2, 3]. Only in the 1960s did studies of sleep in non-mammalian animals start appearing [4, 5, 6], and through this comparative approach, we have learned that sleep serves a vital function. Across the animal kingdom, a lack of sleep can detrimentally affect development [7], cognitive abilities [8], and life span [9].
Recent innovations in recording techniques [10] and animal tracking software [11, 12] have revitalized the study of sleep in non-mammalian animals, and recent publications have reported sleep in animals as diverse as octopus [13] and bearded dragons [14]. What is sleep, exactly, and what can we learn by studying sleep (and its functions) in non-mammalian brains? This is the topic of this short review.
While we all know what it feels like to be asleep, how do we go about determining whether an animal is asleep? In the field of comparative sleep research, sleep is usually defined using two frameworks. One framework involves a behavioral definition of sleep. This definition was first described by Piéron in 1913 [15] and later extended by Flanigan and Tobler in the 70s and 80s into a set of operational criteria including: (1) the presence of a specific sleeping site; (2) a typical body posture; (3) physical quiescence; (4) elevated arousal threshold; (5) rapid state reversibility; and (6) regulatory capacity, i.e., (homeostatic) compensation after loss [16, 17, 18, 19]. Using this behavioral framework, it became possible to investigate sleep-like behavior in a number of non-mammalian vertebrates and invertebrates.
In particular, the measurement of arousal thresholds has been instrumental in a large body of comparative sleep research, especially for work in invertebrates like fruit flies, nematodes, and even jellyfish [20, 21, 22]. Typical responses show that when an animal is engaged in sleep, more intense stimuli are needed to elicit a behavioral reaction than when it is engaged in drowsiness or in quiet waking. However, the use of arousal thresholds to identify sleep is not without its pitfalls: stimulus modality, salience, and contextual relevance are highly variable across different animal species, highlighting the problem with assigning a universal “high arousal threshold” metric across species. Furthermore, while behavioral definitions of sleep offer important guidelines that are appropriate in most cases, they may not be suitable for describing complex sleep patterns such as those found in aquatic mammals or migrating birds engaged in unihemispheric sleep.
The electrophysiological state of the brain provides another framework to describe sleep. Despite the (often) reduced external behavior during sleep, the sleeping brain remains highly active. The first electrophysiological studies of brain activity during sleep in humans revealed clear electroencephalographic (EEG) correlates of brain states [3], and researchers now divide sleep EEG patterns into two main types: (i) rapid eye movement (REM) sleep and (ii) non-REM (NREM) sleep. These two states of sleep have been documented in numerous non-mammalian animals, including birds [23, 24, 25, 26, 27], reptiles [14, 28, 29], zebra fish [30], and cephalopods [13]. Similar to mammals, REM sleep states in these animals are characterized by low amplitude and aperiodic electrical activity that is highly similar to the awake brain state, whereas NREM sleep is characterized by the emergence of periodic ‘slow-wave activity’ (SWA; 1–4 Hz oscillations) during slow-wave sleep (SWS).
Although there are more than 18,000 species of birds by some counts [31] electrophysiological signs of sleep have been investigated in only a handful of these animals: chickens and pigeons [5]; geese [32]; multiple species of songbirds, including the white-crowned sparrow [33], zebra finch [25, 26], and blackbird [34]; parrots (budgerigars) [23]; sandpipers [35]; burrowing owls [36], and ostriches [37] to name a few.
Sleep in birds is generally similar to mammalian sleep due to the largely uncontested presence of both SWS sleep and REM sleep, although the duration of REM sleep epochs are usually much shorter, the differences between sleep states are much smaller than in mammals [38, 39, 40], and sleep spindles, a hallmark of mammalian NREM, have not been observed during avian sleep [23, 25].
Given the electrophysiological similarity between REM sleep and the awake state (as is also true for mammals), it is important to differentiate between true REM sleep and short bouts of wakefulness in birds. Decreased skeletal muscle tone, concomitant eye movement, and irregular heart rate are all features that accompany REM sleep in mammals. These mammalian co-indicators of REM sleep are present to varying degrees in non-mammalian species [41]. For example, while eye movements are present during REM-like sleep in reptiles [14] and many bird species, they are generally absent in burrowing owls [36]. Many birds remain vigilant during sleep, opening one or both eyes during SWS, and only closing both eyes during REM sleep [36, 42, 43]. Similarly, REM-related atonia in birds is often a function of the specific sleeping posture, and large variability is present in sleep-related electromyography (EMG) readings across avian species [23, 37]. Such findings have led to an observable technology shift in the quantification of avian sleep, in which contemporary studies have replaced classical EMG and electrooculography (EOG) recordings of the eye with non-invasive accelerometry or video recording analysis to assess body movement and posture, and/or eye state (see Table 1 for a review (Ref. [5, 23, 24, 25, 32, 33, 35, 36, 37, 44, 45, 46, 47, 48, 49, 50])).
| Source | Bird species used | SWS presence | REM presence | EMG usage | EOG usage | Movement or accelerometer |
| Klein et al. (1964) [5] | Chicken and Pigeon | Present (as sommeil lent) | Present (as sommeil hypotonique) | Used (Neck muscles) | Used | Visual behavioral observation |
| Rojas-Ramírez & Tauber (1970) [45] | Hawk (Buteo jamaicensis); Falcon (Herpetotheres cachinnans) | Present (as NREM) | Present (as REM) | Used (Neck muscles) | Used (Periorbital screws) | Scanning behavior and motor activity |
| Van Twyver & Allison (1972) [46] | Pigeon (Columba livia) | Present | Present (as Paradoxical Sleep) | Used (Neck muscles) | Used | CCTV monitoring |
| Berger & Walker (1972) [36] | Burrowing Owl (Speotyto cunicularia) | Present | Present (as Desynchronized Sleep) | Used (Neck and eye muscles) | Used (Orbital screws) to confirm absence of REM | Strain gauge and infrared CCTV |
| Walker & Berger (1972) [47] | Domestic Pigeon (Columba livia) | Present | Present (as REM) | Used (Identical to owl study) | Used | Head movements recorded |
| Dewasmes et al. (1985) [32] | Goose (landaise strain) | Present | Present | Used (Neck muscles) | Used | CCTV/Behavioral observation |
| Szymczak (1989) [48] | Rook (Corvus frugilegus); Magpie (Pica pica) | Present | Present (as Paradoxical Sleep) | Used (Electrographic) | Used (Electrographic) | Behavioral observation |
| Rattenborg et al. (2004) [33] | White-crowned sparrow (Zonotrichia leucophrys gambelii) | Present | Present | Used (Neck muscles) | Not Mentioned | Infrared beam and video |
| Low et al. (2008) [25] | Zebra Finch (Taeniopygia guttata) | Present | Present | Not Mentioned | Not Used | Video monitoring |
| Lesku et al. (2011) [37] | Ostrich (Struthio camelus) | Present | Present (Mixed state) | Used (Neck muscles) | Used | 3-axis head accelerometer and GPS |
| Lesku et al. (2012) [35] | Pectoral Sandpiper (Calidris melanotos) | Present (as NREM) | Rarely Observed | Used (Neck muscles) | Not Used | Video and EMG-based activity |
| Rattenborg et al. (2016) [44] | Great frigatebird (F. minor) | Present | Present | Not Used | Not Used | 3-axis head accelerometer and GPS |
| Tisdale et al. (2017) [49] | Elegant crested tinamou (Eudromia elegans) | Present | Present | Used (Neck muscles) | Used | 3-axis head accelerometer and video |
| van Hasselt et al. (2020) [50] | European Starling (Sturnus vulgaris) | Present (as NREM) | Present | Not Used | Not Used | 3-axis head accelerometer |
| Canavan & Margoliash (2020) [23] | Budgerigar (Melopsittacus undulatus) | Present | Present | Not Used (Atonia considered unreliable) | Used | Video and automated motion detection |
| Libourel et al. (2023) [24] | Chinstrap penguin (Pygoscelis antarcticus) | Present | Present | Used (Neck muscles) | Not Used | 3-axis head accelerometer, GPS, and pressure sensors |
Atonia and EMG: Earlier studies (1960s–1980s) almost exclusively used neck EMG to identify the reduction of muscle tone during REM. More recent studies have omitted EMG leads because REM atonia is considered an unreliable indicator in birds. SWS, slow-wave sleep; EMG, electromyography; EOG, electrooculography; REM, rapid eye movement; NREM, non-REM; CCTV, closed-circuit television; GPS, Global Positioning System.
Further complicating the issue of avian sleep is that in addition to bilateral phases of SWS and REM, where both hemispheres exhibit the same sleep state, many species of birds show bouts of unilateral SWS (USWS) - the condition where one hemisphere shows unambiguous waking EEG activity while the other shows unambiguous SWS activity. Birds engaged in long migratory flights demonstrate both bilateral sleep as well as USWS [44]. USWS activity has been linked to unilateral eye opening, such that the awake EEG activity accompanies contralateral eye opening, and some authors have posited that USWS is under facultative control in birds and may have a role in heightening vigilance against predators [42, 51].
The widespread activity that occurs in the brain during sleep has a purpose; however, there is still no consensus on what that might be. Non-mutually exclusive hypotheses range from energy conservation [52], brain thermoregulation [53], detoxification [54], and tissue restoration [55]. One view, which is well supported by data, is that sleep benefits memory [56].
In psychological terms, memory is defined as the ability to encode, store, and subsequently retrieve information and past experiences [57]. Memory is the sum total of what we remember, and it endows us with the capability to learn and adapt from previous experiences. Over the years, psychologists have established three main categories of memory: short-term memory, which can only hold limited information; long-term memory, which can store an indefinite amount of information; and sensory memory, which is not consciously controlled [58].
There are also different types of memories: declarative memories are consciously acquired and include memories of experiences (i.e., episodic memory) and facts (i.e., semantic memory). In contrast, non-declarative memories are implicitly acquired and include procedural skill learning and behavioral conditioning (i.e., classical or operant conditioning [59]. While evidence suggests that non-declarative memories involve the basal ganglia and motor cortex, declarative memories involve the hippocampus [60].
Newly formed declarative memories require consolidation, a process by which they are transformed and stabilized into long-term memory [61, 62]. While the initial encoding of a memory is a rapid (milliseconds) process, its long-term maintenance requires processes that continue to modify it over hours to years, as long-term memory representations are distributed over brain circuits. Compelling evidence in humans [63, 64, 65] and rodents [66, 67, 68] has shown that these processes occur during sleep and are collectively known as memory consolidation.
Research suggests that the consolidation of declarative and non-declarative memory differentially depends on the sleep state. For example, studies have shown that humans perform better in declarative memory-related tasks after sleep compared to wakefulness [63, 64, 69] and that this improvement is linked to SWS [70, 71, 72]. On the other hand, REM sleep has been shown to be important for perceptual tasks [73] and motor/visuomotor tasks in humans [74, 75], and for procedural memories acquired through behavioral training in rodents [66, 67, 68]. These results demonstrate that memory consolidation during sleep may require different modes of brain activity. In this review, we will focus on memory consolidation involving the hippocampus and the brain oscillations known as sharp-wave ripples.
During phases of SWS, large-scale synchronous neural activity is thought to enable the transfer of information from the hippocampus to numerous regions of the neocortex [76]. These sharp wave-ripple (SWR) events result from the coordinated firing of many neurons, and in the rodent hippocampus, the neural activity that initiates a SWR is well understood: massive excitation of neurons in the Cornu Ammonis 1 (CA1) subfield by the pyramidal neurons in the CA3 subfield causes a synchronization of inhibitory interneurons, which generates a ripple in the pyramidal layer [77].
Importantly, the spiking activity associated with SWRs is highly structured across neurons. In mammals, this activity reflects a temporally compressed version of sequential neuronal firing patterns experienced by the awake animal [78]. Specifically, in the rodent hippocampus, firing patterns in place cell assemblies are “replayed” in the same temporal order as that which occurred during the awake task performance [79, 80], and many studies have documented replay events during SWRs, where sets of hippocampal place cells that fired together during exploration are more likely to fire together afterwards during sleep or awake rest [78, 81, 82].
Notably, studies in rodents have demonstrated that SWRs have a causal role in memory consolidation [83, 84, 85, 86]. In these experiments, animals that underwent SWR suppression during sleep performed significantly worse in spatial memory tasks compared to control animals. These results highlight the functional role of SWRs in the consolidation of spatial memories.
The local network activity within the hippocampus is also coordinated with distant cortical activity [87], and neural coordination between brain areas provides a likely substrate for the network activation and reactivation required for learning and memory. Through its large-scale synchronous activity, SWR output brings about sequential activations in the prefrontal cortex, leading to a systems-level transformation of new experiences into stable memory traces. SWRs appear to be conserved throughout mammalian evolution: they have been found in every mammal investigated so far, including humans [79].
How do the avian brain and the mammalian brain compare? The “sameness” of neural circuits or brain areas that is due to common evolutionary origin across animals is known as “homology” [88]. The identification of homologies across brain areas in different animals remains an active area of research that has been revitalized by recent advances in transcriptomic methods [89]. The goal is that by studying homologies across neural circuits, we can gain considerable insight into their canonical function across animals.
During embryonic development, a remarkably similar “bauplan” exists across all vertebrate brains [90]. First, the primordial telencephalon develops into the pallium and the subpallium. Then the pallium further differentiates into a number of subregions that give rise to different brain structures in different animals [91]. Sometimes, these brain areas are similar across structure and function; for example, in both birds and mammals, the medial pallium gives rise to the hippocampal formation. In birds, the hippocampal formation receives visual input from the Wulst [92], and general characteristics such as connectivity, neuron types, and the presence of adult neurogenesis seem to be conserved between birds and mammals [93]. Similar to mammals, spatial learning in birds (see next section, “Models of memory and learning in birds”) also seems to rely on the function of the hippocampus.
Sometimes, however, embryonic development gives rise to very different neural architectures in mammals and birds. In mammals, for example, the dorsal pallium gives rise to the 6-layered neocortex, whereas in birds, the dorsal pallium becomes a structure known as the hyperpallium. Although the mammalian neocortex and avian hyperpallium derive from the same dorsal pallial structure, there is very little resemblance between the two structures in terms of morphology or function [94]. Indeed, the cognitive functions that are associated with the 6-layer mammalian neocortex seem to be seated in another brain area altogether in birds known as the “dorsal ventricular ridge” (DVR).
In mammals, parts of the ventral pallium give rise to the amygdalar complex [95]. In birds and reptiles, the ventral pallium gives rise to the DVR. This large pallial region dorsal to the basal ganglia is unique to reptiles and birds. In general, the anterior DVR consists of spatially segregated neuronal types specialized in processing visual, auditory, or somatosensory stimuli [96], whereas the posterior DVR is thought to be homologous with the mammalian claustrum and amygdala [97]. In birds, the DVR is subdivided into the mesopallium, nidopallium, entopallium, and arcopallium (Fig. 1A).
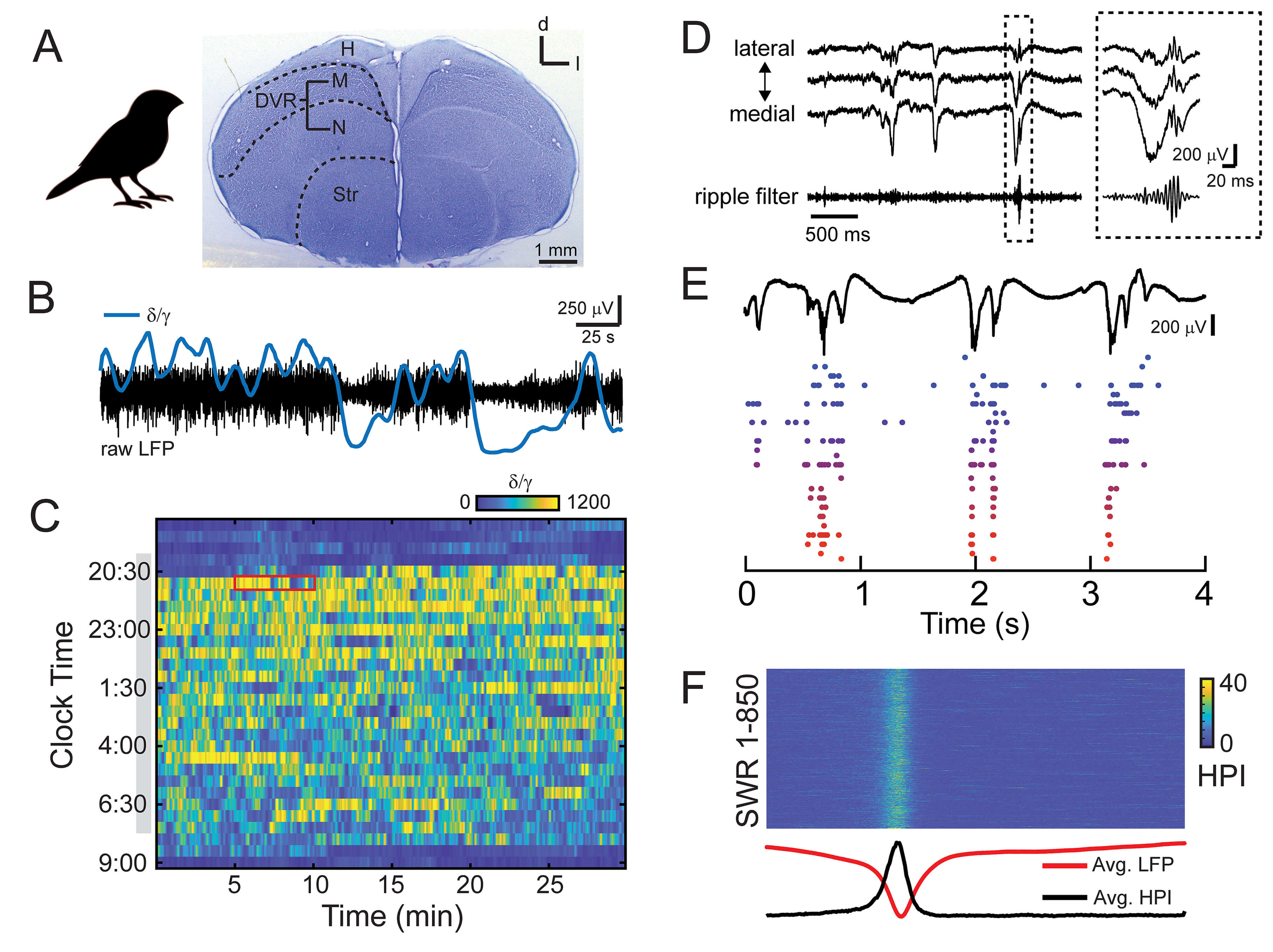 Fig. 1.
Fig. 1.
Sleep characteristics and sharp wave-ripple (SWR) activity in
the avian brain. (A) The zebra finch brain. Histological section stained with
cresyl violet indicates the subdivision of the avian pallium. H, hyperpallium; M,
mesopallium; N, nidopallium; Str, striatum; DVR, dorsal ventricular ridge.
Entopallium and arcopallium are not pictured. Scale bar indicates 1 mm. (B) 5
minutes of raw local field potential (LFP) (black) recorded during natural sleep.
Data corresponds to red box in (C). The LFP is composed of distinct states, as
evident in the
Although the regions of the DVR have no direct mammalian counterpart, recent advances using imaging technology, viral tools, and single-cell transcriptomics seek to better understand the relationship between the avian DVR and mammalian brain structures. The emerging story is complicated.
For example, although the avian DVR appears to lack a laminar structure, a recent imaging study showed that the sensory DVR displays radially and tangentially organized fibers, similar to the canonical microcircuit of the mammalian neocortex [98]. On the other hand, recent experiments that used single-cell transcriptomics to characterize gene expression patterns in the avian DVR revealed that the avian DVR and the mammalian neocortex derive from fundamentally different neurodevelopmental regions and are therefore not homologous structures [99].
The story is also complicated at the cellular level. A recent study used optogenetic tools to specifically target excitatory and inhibitory neurons in the avian DVR and found physiological and computational similarities with mammalian excitatory and inhibitory neocortical neurons [100]. However, experiments using single-cell sequencing found that the entire songbird pallium is populated by a type of interneuron that is largely absent in the neocortex [99], suggesting that “cortical-like” microcircuits described in the avian DVR may engage an entire class of interneurons that has no counterpart in the mammalian neocortex [101]. These contrasting results highlight the dearth of knowledge about the cell types, anatomical connections, and computations of the avian DVR.
Although the anatomy of avian DVR is fundamentally different from the mammalian networks that underlie memory consolidation, its placement as a nexus of sensory input within the avian telencephalon might make DVR networks well-suited for a role in learning and memory, allowing avian intelligence to arise from neural circuitry fundamentally similar to that of the neocortex. And indeed, many of the impressive learning abilities of birds rely on networks within the DVR.
The avian DVR is often compared to the mammalian neocortex [102], and indeed, the avian DVR has received renewed attention in recent years because of its role in multiple avian learning paradigms. For example, high-level cognition in corvids has been linked to a nidopallial area of the DVR (nidopallium caudolaterale [NCL]; Table 2, Ref. [33, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139]). NCL has been compared to mammalian neocortex [102, 103], and neurons in NCL have been shown to encode information ranging from numerosity [104] to abstract behavioral rules [105] to sensory consciousness [106].
| Learning modality | Bird | Abbreviation | Brain region | Brain area | References |
| Vocal Learning | Zebra finch | NCM | nidopallium caudomedial | DVR - nidopallium | [107, 108, 109, 110] |
| CM | caudal mesopallium | DVR - mesopallium | [111] | ||
| CLM | caudal lateral mesopallium | DVR - mesopallium | [112] | ||
| Av | Avalanche, caudal mesopallium | DVR - mesopallium | [113] | ||
| RA | robust nucleus of the arcopallium | DVR - arcopallium | [114, 115, 116] | ||
| Avian Cognition | Carrion crow | NCL | nidopallium caudolaterale | DVR - nidopallium | [103, 104, 105, 106] |
| Visual Imprinting | Chicken | IMM | intermediate and medial mesopallium | DVR - mesopallium | [117, 118, 119, 120, 121] |
| Spatial memory | Zebra finch | HF | Hippocampal formation | Hippocampus | [122] |
| Chickadee | HF | Hippocampal formation | Hippocampus | [127, 128, 129] | |
| Homing pigeon | HF | Hippocampal formation | Hippocampus | [125, 126] | |
| Quail | HF | Hippocampal formation | Hippocampus | [123, 124] | |
| Episodic memory | Chickadee | HF | Hippocampal formation | Hippocampus | [133] |
| Jays | [130, 131, 132, 137] | ||||
| Blue tit, great tit | [134] | ||||
| Operant learning | Zebra finch | [138, 139] | |||
| Starlings | [135, 136] | ||||
| White-crowned sparrows | [33] |
DVR, dorsal ventricular ridge.
Similarly, key elements of vocal learning in songbirds rely on brain areas in the DVR. During vocal learning, juvenile zebra finches use auditory feedback and motor learning to transition from acoustically simple songs to complex and stereotypical adult songs [140], a complex memory task that involves the formation of auditory memories (the “template”), sequences of motor output, and associative higher-order representations of learned vocalizations [141, 142, 143]. Vocal learning requires a set of interconnected brain nuclei that share analogies with brain areas involved in human language learning [144], and several areas within the DVR have been implicated in vocal learning. For example, immediate early gene studies have suggested that neurons in a nidopallial area of the DVR (nidopallium caudomedial [NCM]) may encode the memory of the “tutor” song during vocal learning [107, 108, 109, 110]. Similarly, neurons in mesopallial parts of the DVR have been shown to have a role in auditory object recognition [111] (the caudal mesopallium [CM]) and to encode stimulus surprise [112] (caudal lateral mesopallium [CLM]), whereas neurons in another small cluster of neurons embedded in the CM (Avalanche) have been shown to have a role in juvenile song copying [113]. Finally, neurons in the arcopallial part of DVR, in a nucleus known as the robust nucleus of the arcopallium (RA) have been shown to spontaneously reactivate during sleep [114, 115, 116], highly similar to the replay observed in mammalian hippocampus.
Filial (visual) imprinting also involves brain areas in the mesopallial part of the DVR. Young, visually naive domestic chicks quickly learn the characteristics of certain visually conspicuous objects upon being exposed to them. This imprinting memory critically relies on the left intermediate and medial mesopallium (IMM; formerly known as the intermediate and medial hyperstriatum ventral [IMHV] [117, 118, 145]): destruction of this region before training impaired imprinting [119, 120]; destruction after training eliminated an acquired preference [121]. Furthermore, biochemical studies have shown that several features of the IMM undergo changes as a result of visual imprinting, including an increase in the size of post-synaptic densities percentage in IMM neurons and upregulation of N-methyl-D-aspartate (NMDA)-type glutamate receptors [117], highlighting a molecular mechanism for visual imprinting in the DVR.
Spatial learning refers to the process by which an animal acquires a mental representation of its environment, and in humans, it is considered to be a form of declarative memory [146]. Tasks that query spatial learning have long been the preferred method for investigating memory consolidation in rodents, and decades of research have culminated in an understanding that the hippocampus is essential for memory consolidation. However, it is clear that non-mammals such as birds are also capable of learning and forming memories, and a significant amount of memory research has focused on the avian hippocampus, known as the “hippocampal formation”.
Similar to the spatial learning tasks commonly performed in rodents and which rely on the hippocampus, experiments investigating spatial learning in birds typically involve measuring the effect of hippocampal lesions on spatial memory tasks. For example, zebra finches that received ibotenic acid lesions in the hippocampus were impaired in their ability to remember the location of hidden seeds [122]. Similar studies conducted in Japanese quail [123, 124], homing pigeons [125, 126], as well as a seed-caching songbird, the black-capped chickadee [127, 128, 129], have all implicated the avian hippocampus in spatial learning.
Episodic memory also relies on the hippocampus, and it is often considered the “pinnacle” of the different types of memory [147]. Episodic memories are memories of experiences, and they provide the “what-when-where” content for an experience [148]. Although spatial memories provide a structural framework for episodic memories by encoding information about the location where an event may have occurred, episodic memories must also have an association of what happened and when it occurred in time.
Experiments that leveraged the seed caching abilities of some bird species highlight the evidence that episodic memory is not unique to humans [147, 148]. For example, scrub jays were able to remember the locations of perishable and non-perishable food items and recover the food before it became inedible [130, 131, 132]. Recent work in another seed-caching specialist, the chickadee, was able to show that seed-caching involved event-specific “barcodes” of neural activity in the hippocampus, which encoded unique identifiers to episodic memory events [133]. Finally, evidence of episodic memory also appears in non-seed caching generalist foragers (like blue tits and great tits [134]) and hummingbirds [149].
A growing body of evidence has shown that sleep is an essential requirement for several learning and memory tasks in birds. During vocal learning, anecdotal evidence reports that juvenile birds tend to fall asleep after first exposure to the singing of an adult tutor [150] and furthermore, that the initial tutor song exposure causes a circadian pattern of vocalization deterioration which continues into adulthood, but does not exist prior to tutor song exposure [151]. Immediate early gene expression increases during sleep in learning juveniles in a brain area thought to house the auditory memory of the tutor song, and this activation is related to the amount of tutor song exposure and accuracy of song performance during prior wakefulness [152]. Finally, neurons in a premotor brain nucleus involved in song learning (RA) showed an increase in high-frequency bursting activity during sleep in juvenile zebra finches [116], and spontaneous activity patterns recorded during sleep in adult zebra finches were highly similar to patterns recorded during daytime singing [114, 153, 154]. Interestingly, recordings from the muscles of the syrinx, the vocal organ of songbirds, also show song rehearsals during sleep [155].
Similarly, studies investigating visual imprinting have also highlighted a role for sleep. Specifically, neurons in the IMM become selectively responsive to a trained imprinting stimulus [117]. This stimulus selectivity is critically dependent on a period of SWS after exposure to the visual imprinting stimulus for the memorization of the stimulus to be successful [118].
Operant learning has also shown a requirement for sleep in birds. One study, in which starlings were trained to discriminate between different auditory stimuli, showed that sleep supported the consolidation of recent auditory memories [135], and furthermore, that sleep could improve learning for interference tasks as well [136]. Similarly, white-crowned sparrows which were trained on a pecking task performed less accurately after their sleep was restricted to 3 hours [33].
Although sleep has been shown to be required for spatial memory task performance in rats [156] and humans [157], scientific reports of its effects on spatial memory learning in birds remain rare. In one study which focused on mono-ocular sleep (sleep with one eye open), chicks were trained in a spatial learning task to find food in one of 4 containers, and chicks reportedly spent more time sleeping during learning compared to non-learning control chicks [158, 159]. Altogether, this evidence highlights a critical role for sleep in a wide range of avian learning paradigms. However, the exact mechanisms of this sleep requirement remain unclear: how does sleep support learning in a brain that lacks the complex laminar neural-architecture that defines the mammalian brain?
Exactly how diverse memories are consolidated across the networks of the avian brain is not currently understood. In mammals, the process of memory consolidation involves specific interactions between two main brain areas: the hippocampus and the cortex. Is the specific neuronal hardware (e.g., neural circuits, physiological properties) a critical component of this process? Or alternatively, are there specific population patterns (i.e., SWRs) that are vital to memory consolidation and can arise from various neural-architectures? The latter theory is appealing, as it suggests that other brain areas that participate in the encoding of the animal’s experience during behavior may use similar schemes to transfer and consolidate memories and experiences. In the following sections, I will review some of our recent findings to highlight the importance of studying sleep, learning, and memory in alternative animal models like birds.
Through its large-scale synchronous activity, SWR output brings about sequential activations in the prefrontal cortex, leading to a systems-level transformation of labile experiences into stable memory traces [160]. We found evidence of SWR-like activity in the avian brain that is remarkably similar to mammalian hippocampal SWR activity [27, 161] during SWS in birds. Importantly, we did not record SWR activity in the avian hippocampus [162] but rather in the avian DVR. Avian SWRs are characterized by a large, sharp wave in the local field potential (LFP), a fast ripple oscillation, and a burst in gamma-band activity [27]. Based on these results, I propose that avian SWRs have several attributes that likely facilitate memory consolidation in the avian brain.
In birds, it is possible to track the transitions between different phases of
sleep by calculating the ratio of delta activity (
As is true in mammals, SWS phases in birds correspond to an increase in delta activity (Fig. 1C). Avian SWRs appear during phases of SWS in zebra finches, paralleling the situation in the rodent hippocampus, where SWR activity specifically occurs during bouts of rest and SWS [77]. Similar to mammals, sleep in birds likely provides an optimal “offline” period for memory consolidation due to the greatly reduced processing of external information [165].
Avian SWRs can be recorded throughout a large expanse of the avian brain. Using 4-shank silicon probes in isoflurane-anesthetized zebra finches, we measured avian SWRs throughout the medial-lateral extent of the avian DVR. Avian SWRs were apparent at recording locations throughout this area; however, the shape of the sharp wave and the presence or absence of the ripple component appear to change as a function of spatial position (Fig. 1D). These findings were also replicated in our recent paper using Neuropixels 1.0 probes (IMEC, Leuven, Belgium) chronically implanted in the brains of male zebra finches [27], which revealed avian SWR activity that spanned more than 3 mm. By affecting neural activity over such a large part of the brain, avian SWRs have the capacity to synchronize the firing of many neurons in a powerful, population-wide event that can affect both local and distal neural activity.
Avian SWRs are associated with increased neural activity, and across neurons, these firing patterns form distinct sequences of activity that could represent neural reactivation of events experienced during awake periods (Fig. 1E). These patterns are highly similar to the reactivation of neuronal firing patterns, or “replay” events, observed in the rodent hippocampus [77, 79] and drive the population in a temporally locked spiking pattern (Fig. 1F).
In birds, the relationship between patterns of neural firing during SWRs and awake behavior remains unresolved. Furthermore, it remains unclear which behavioral experiences might be replayed during sleep in birds. Further work is required to specify how neural activity patterns that occur during awake behavior are recapitulated during SWRs in offline sleep phases.
Could it be that SWRs in avian brains are used for memory consolidation or memory transfer? If true, SWRs may constitute a basic mechanism for memory transfer that is shared across vertebrate brains. Importantly, similar SWR-like LFP patterns have also been recorded during sleep in the reptilian DVR [14, 28, 29]. The fact that SWR-like activity patterns arise from both reptilian and avian DVR during sleep suggests that some version of the neural circuit that gives rise to mammalian hippocampal SWRs may also be present in the avian DVR – despite the absence of clear anatomical lamination in the DVR (although see [98]). However, the function of avian SWR-like activity is still a mystery. As proposed by Sherry and Schacter in their landmark paper [166], it is possible that an existing memory system may be co-opted for use in other situations than for which it had been evolutionarily selected. Further work is required to determine whether the SWR-like activity observed in the avian DVR is an “epiphenomenon” related to the ongoing dynamics of the sleeping brain, or whether it has a true function in learning and memory. Experiments that build upon the recent approaches in rodents to block replay in birds would more clearly link the dynamics of replay and memory consolidation in avian models.
As others have highlighted [167], comparative approaches serve as powerful tools in assessing the validity of universal principles in neuroscience research. This is especially true when we consider that for the past decades, the field of neuroscience has been slowly converging on a few selected model organisms. By diversifying the field of memory research to include alternative animal models and behaviors, we will deepen our existing knowledge about sleep, learning, and memory, and may thereby reveal foundational principles that underlie learning and memory for all animals.
A previous German version of this paper was published as “Schlaf und Gedächtnis in neuronalen Nicht-Säugetier-Architekturen” in Neuroforum (e-ISSN 2363-7013), 2024, Volume 30, Issue 2, https://nwg-info.de/sites/nwg-info.de/files/media/pdf/neuroforum/Neuroforum_02-2024.pdf.
JMO prepared the figure and conducted the literature search for the manuscript and wrote the paper. JMO read and approved the final manuscript. JMO has participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
The author would like to thank the two anonymous reviewers for their comments on a previous version of this manuscript as well as Harald Luksch for his support of her independent research at his Chair of Zoology at the Technical University of Munich.
This research was funded by Deutsche Forschungsgemeinschaft (ON 151/1-1).
The author declares no conflicts of interest.
The author used Google Notebook LM to assemble some of the information in Table 1. After using this tool, the author reviewed the information in the table and takes full responsibility for its contents.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.