

 , Jing Li 3,*
, Jing Li 3,*1 Department of Preventive Medicine, School of Public Health, Wenzhou Medical University, 325027 Wenzhou, Zhejiang, China
2 Department of Neurology, The Second Affiliated Hospital of Wenzhou Medical University, 325000 Wenzhou, Zhejiang, China
3 Department of Neurology, The First Affiliated Hospital of Wenzhou Medical University, 325027 Wenzhou, Zhejiang, China
†These authors contributed equally.
Abstract
The probiotic Lactobacillus mucosae has been widely shown to have many positive effects. However, its neuroprotective effects and underlying mechanism in Alzheimer’s disease (AD) remain elusive.
Male APP/PS1 mice were treated for 4 weeks with L. mucosae WMU007, followed by the evaluation of cognitive function, neuronal damage, amyloid-β (Aβ) deposition, and Tau phosphorylation. RNA-seq coupled with Gene Ontology (GO) enrichment analysis implicated L. mucosae WMU007 in modulating oxidative stress in this AD model. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis and qPCR were performed to identify the specific mechanism by which this probiotic suppresses oxidative stress in the pathogenesis of AD. In addition, we quantified the levels of classical oxidative stress markers, such as superoxide dismutase 2 (SOD2) and glutathione peroxidase 4 (GPX4). We also examined the expression of cannabinoid receptor type 2 (CB2) and its key downstream regulators in the redox pathway, namely nuclear factor erythroid 2-related factor 2 (Nrf2) and heme oxygenase 1 (HO-1), in both animal and cellular models.
Our results showed that treatment with L. mucosae WMU007 significantly decreased cognitive impairment, neuronal damage, Aβ deposits, and Tau phosphorylation in APP/PS1 mice. Activation of CB2 was identified as the key mechanism by which L. mucosae WMU007 reduces oxidative stress in AD. In addition, L. mucosae WMU007 reduced oxidative stress and increased the levels of CB2 pathway-related proteins in vivo and in vitro.
These results indicate that L. mucosae WMU007 confers neuroprotection in AD by targeting CB2-mediated oxidative pathways, highlighting its therapeutic potential as a novel probiotic intervention.
Keywords
- Alzheimer’s disease
- cannabinoid receptor type 2
- Lactobacillus mucosae
- neuroprotective effects
- oxidative stress
Alzheimer’s disease (AD) is a progressive neurodegenerative disorder and the
leading cause of dementia. AD is primarily characterized by cognitive impairment,
and its incidence is rising with the aging of the global population [1, 2, 3]. The
pathogenesis of AD is multifactorial and involves processes such as
amyloid-
Probiotics are live bacteria that interact with the intestinal environment and
influence brain function through the bidirectional gut-brain communication axis
[13]. Lactobacillus mucosae WMU007, a probiotic bacterium from the
healthy human gut, has recently attracted increasing attention [14]. Oral
administration of Lactobacillus mucosae NK41 alleviated cognitive
deficits and A
In this study, we examined the impact of L. mucosae WMU007 on cognitive impairment and pathological changes in the APP/PS1 mouse model. We elucidated the role of L. mucosae WMU007 in modulating oxidative stress in this AD model. The levels of oxidative stress-related proteins (SOD2 and GPX4) following treatment with L. mucosae WMU007 were also evaluated, allowing us to identify the potential mechanism by which L. mucosae attenuates oxidative stress. The expression of CB2 and downstream redox regulators, including Nrf2 and heme oxygenase 1 (HO-1), was also assessed. We demonstrated that L. mucosae WMU007 could inhibit oxidative stress via CB2-mediated signaling in APP/PS1 mice. This comprehensive study of how the specific microbiota L. mucosae influences oxidative stress and AD progression offers new insights into its therapeutic potential for AD.
L. mucosae WMU007 was deposited in the China General Microbiological
Culture Collection Centre (CGMCC). The strain was cultured in MRS broth
(HB0384-1, HopeBio, Qingdao, China) under aerobic conditions at 37 °C for 24 h.
Bacterial cells were collected by centrifugation (5000 rpm) and subsequently
suspended in phosphate-buffered saline (PBS, ST477,
Beyotime, Shanghai, China) to obtain a final density of 1
All experimental animals, including 6-month-old male APP/PS1 transgenic mice and C57BL/6J mice, were obtained from the Hangzhou Ziyuan Laboratory Animal Technology Co., Ltd. (Hangzhou, Zhejiang, China). The mice were housed under controlled environmental conditions, including an ambient temperature of 20–22 °C and a relative humidity maintained at approximately 50%. A 12-hour light/dark cycle was applied, with illumination from 08:00 to 20:00, and animals had ad libitum access to standard chow and drinking water. The mice were randomly divided into three groups: the WT group, the APP/PS1 group, and the L. mucosae WMU007-treated group (APP/PS1 + WMU007 group). Mice in the APP/PS1 + WMU007 group were intragastrically administered L. mucosae WMU007 for 4 weeks, while mice in the WT and APP/PS1 groups received an equal volume of PBS as a control. All mice were gavaged with 8 mg arachidonic acid (A131025, Aladdin, Shanghai, China) before L. mucosae WMU007 or PBS treatment. Then, all mice underwent behavioral evaluation, such as the Nesting behavior test, novel object recognition test (NORT), and open field test (OFT).
The NORT was applied to assess the memory and cognitive abilities of mice. Two
identical cuboids were symmetrically placed (5 cm from the edges of the field) in
an open field with a bottom side length of 25 cm, a height of 25 cm, and each
mouse was permitted to explore the open field without restriction for 5 min. The
next day, one of the cuboids was replaced with a cylinder of the same material
with a bottom diameter of 3 cm, a height of 6 cm, and each mouse was allowed to
explore freely for 5 min. After the mouse completed the test, the equipment was
wiped with 75% alcohol to eliminate odor interference. Then, the time of the
mouse exploring the cylinder (defined as TN) and the cuboid (defined as TF) were
calculated. Mice sniffing and lying on objects were defined as exploration; the
discriminant index (DI) = (TN – TF) / (TN + TF)
Nesting behavior, which is closely associated with hippocampal function in
rodents, was assessed using a nesting test. Thirty squares of paper (each
measuring 5 cm
The OFT was carried out to measure the exploratory behavior and curiosity of mice towards the new environment. Each mouse was placed in an open field with a side length of 20 cm at the bottom and a height of 30 cm, and permitted to explore freely for 5 min. The active time and rest time of mice were quantified using the Digbehv Animal Behavior Analysis System (version 2.0, Shanghai Jiliang Software Technology Co., Ltd., Shanghai, China). At the end of each test, the open-field apparatus was cleaned with 75% ethanol and allowed to air-dry to prevent potential olfactory cues from influencing subsequent tests.
After behavioral testing, mice were deeply anesthetized by intraperitoneal injection of pentobarbital sodium (P3761, Sigma-Aldrich, St. Louis, MO, USA) (50 mg/kg). Adequate anesthesia was confirmed by the absence of the pedal withdrawal reflex. The mice were then euthanized by cervical dislocation, and brain tissues were quickly collected. The brain tissue was quickly removed and
thoroughly homogenized in a RIPA Lysis Buffer (P0013B, Beyotime, Shanghai, China)
mixed with PMSF and Protein Phosphatase Inhibitor (P1260, Solarbio, Beijing,
China), centrifuged at 4 °C at 12,000 rpm for 30 min, and the supernatant was
removed. The total protein concentration was measured by the Enhanced BCA Protein
Assay Kit (P0010, Beyotime, Shanghai, China), and the sample was denatured at 95
°C for 5 min with the SDS-PAGE Sample Loading Buffer (P0015L, Beyotime, Shanghai,
China). The 40 µg of protein from each sample were added to 10% SDS-PAGE
gel system for electrophoretic separation and transferred to the PVDF membrane.
Then, the membrane was sealed in 5% skim milk at room temperature for 1 h, and
incubated with the following primary antibodies at 4 °C overnight:
A
The brain tissue of mice was fixed in 4% PFA (P0099, Beyotime), dehydrated by gradient alcohol, cleared in xylene (Changshu Hongsheng
Fine Chemical Co., Ltd., Changshu, Jiangsu, China), and embedded after wax
immersion. The wax block was cut into pieces of 5 µm, heated at 65 °C for 2
h, dewaxed in xylene for 30 min, and rehydrated through a graded ethanol series.
Then, the sections were repaired with the antigen via the Hotfix method, soaked
in sodium citrate buffer, heated in a pressure cooker for 5 min, left for 10 min,
and applied for subsequent staining until cooled to room temperature. Next, the
sections were permeabilized with 0.3% Triton for 15 min, blocked in 10% BSA for
1 h, and incubated overnight at 4 °C with primary antibodies, including NeuN
(1:200, HA601111, HUABIO, Hangzhou, China), A
N2a cells were obtained from the National Collection of Authenticated Cell Cultures (Shanghai, China), which provides STR-authenticated cell lines. All cell lines were routinely tested for mycoplasma contamination using a PCR-based detection kit, and all results were negative. APP/SWE cells were generated by transient transfection of N2a cells at 60% confluency with 0.5 µg APPswe695 plasmid using PEI Transfection Reagent (HY-W250110, MCE, Princeton, NJ, USA) and were pretreated with DMEM (C0891, Beyotime) medium containing 20% L. mucosae WMU007 fermentation for 8 h.
The N2a cells were cultured in serum-free medium and exposed to the DCFH-DA probe (S1105S, Beyotime) at 37 °C for 20 min. Subsequently, nuclear counterstaining was carried out using Hoechst dye (C1011, Beyotime), and intracellular fluorescence signals were captured and quantified under a fluorescence microscope.
Total RNA was isolated from mouse cerebral cortex samples with TRIzol reagent
(Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) following
standard protocols. The RNA’s concentration, purity, and integrity were then
assessed. Procedures including mRNA enrichment, RNA fragmentation, reverse
transcription, library construction, and high-throughput sequencing were
completed by Shanghai Majorbio Bio-pharm Biotechnology Co., Ltd. (Shanghai,
China). To ensure the reliability of downstream bioinformatics analyses, raw
sequencing reads were subjected to quality control using fastp (OpenGene, GitHub,
San Francisco, CA, USA). Clean reads with high quality were then
de novo assembled using the Trinity platform. The preliminary assembled
sequences underwent refinement and filtering via TransRate
(http://hibberdlab.com/transrate/). The completeness of the assembled transcripts
was further evaluated with BUSCO (Benchmarking Universal Single-Copy Orthologs).
Differential gene expression analysis was performed using the DESeq2 package
(Bioconductor, Seattle, Washington, USA), and genes meeting the criteria of
Real-time quantitative PCR was performed using ChamQ Universal SYBR qPCR Master Mix (Q711, Vazyme Biotech Co., Ltd., Nanjing, Jiangsu, China) according to the manufacturer’s instructions. Relative gene expression was normalized to Actin and calculated using the 2-ΔΔCT method. The primer sequences were as follows: CB2-F, 5′-ACGGTGGCTTGGAGTTCAAC-3′; CB2-R, 5′-GCCGGGAGGACAGGATAAT-3′.
Data were represented as mean
To investigate the effects of L. mucosae WMU007 treatment on cognitive
dysfunction in APP/PS1 mice, a series of behavioral assessments was conducted. In
NORT, the discrimination index of APP/PS1 mice was significantly lower than that
of WT mice, while WMU007 treatment significantly increased the discrimination
index compared with the APP/PS1 group (p
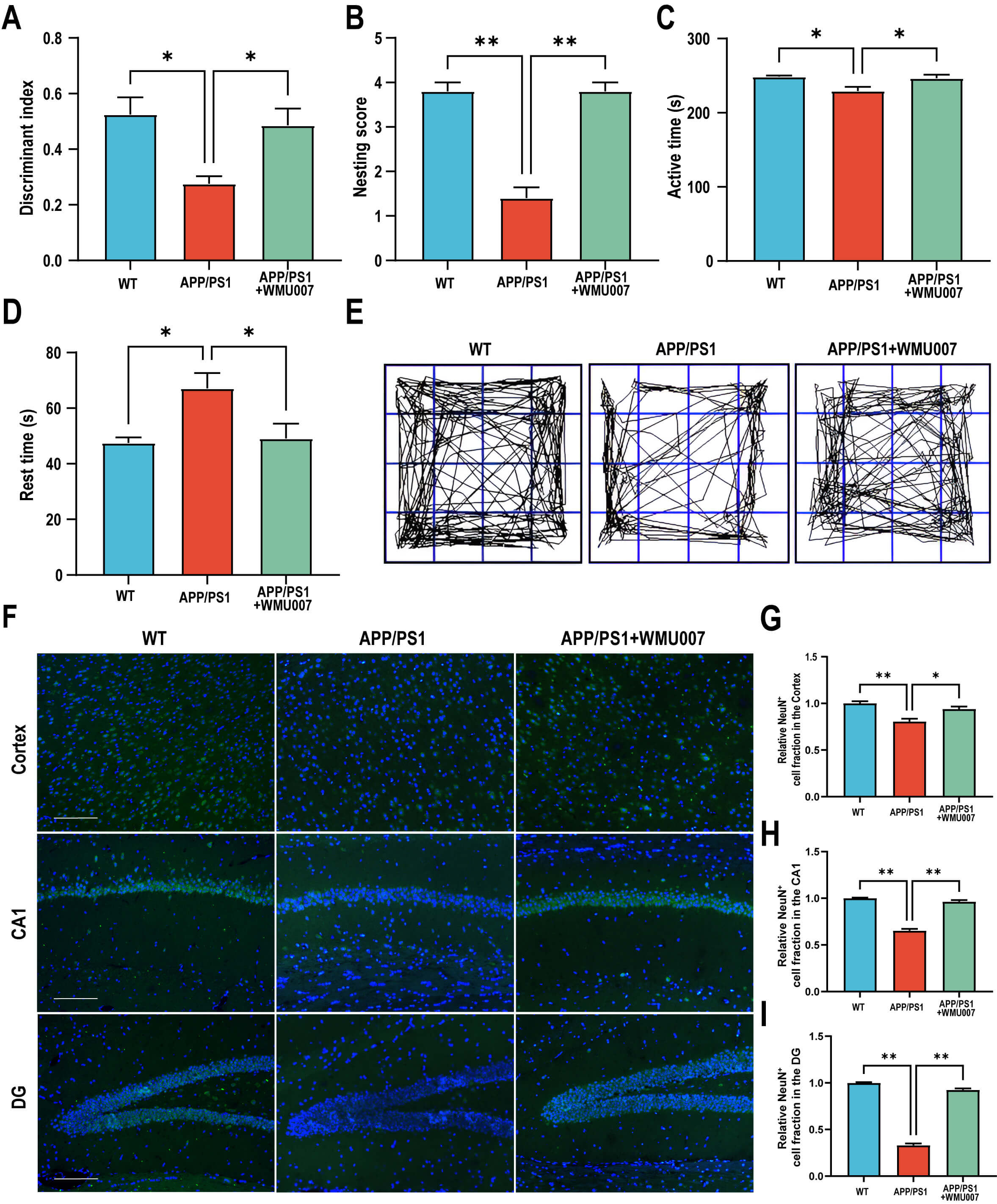
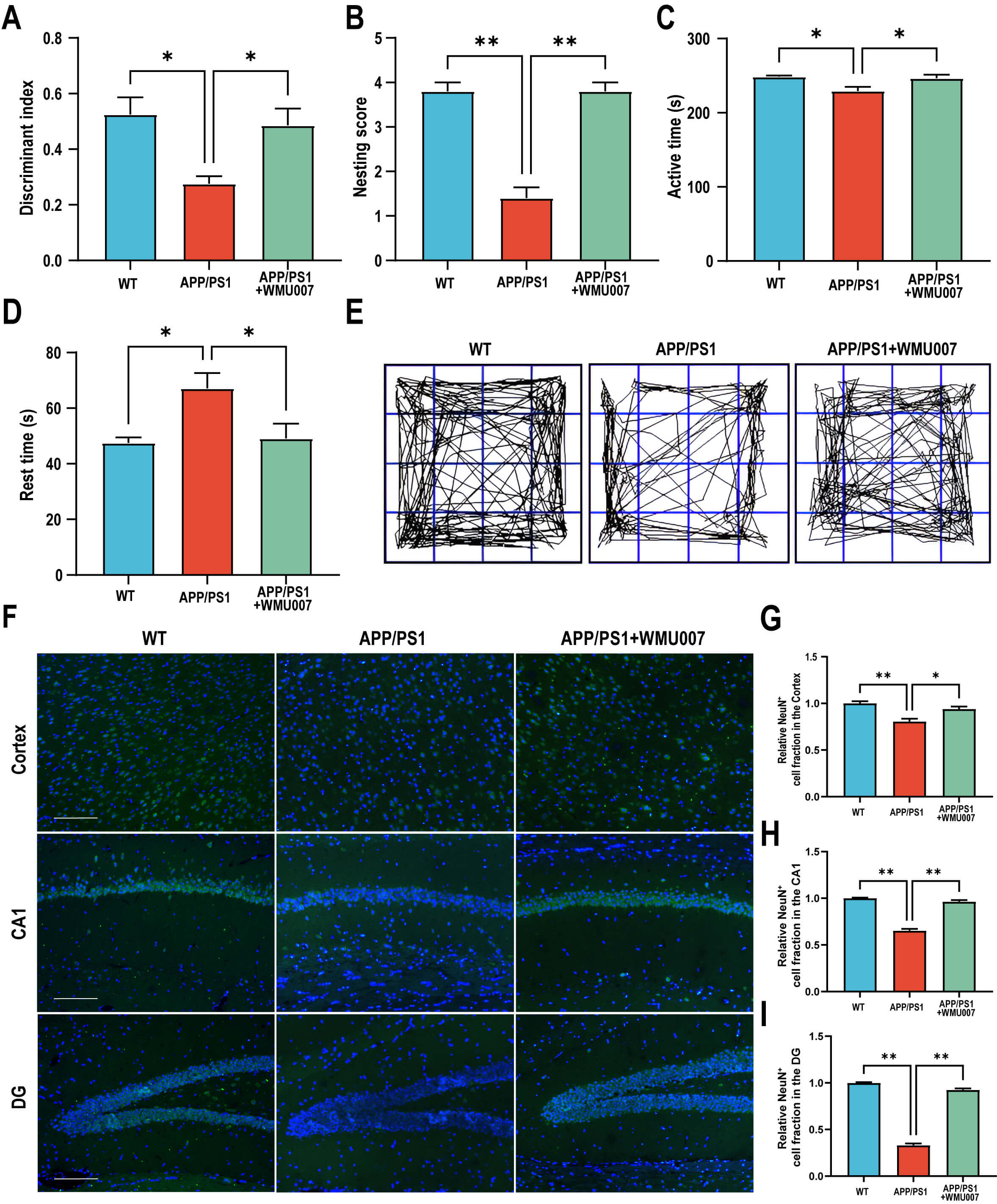 Fig. 1.
Fig. 1.
L. mucosae WMU007 treatment improved cognitive
dysfunction in APP/PS1 mice (n = 5 mice per group). (A) The discriminant index in NORT. (B) The nesting
score in the Nesting Behavior Test. (C) The active time in the open field test.
(D) The rest time in the open field test. (E) Representative images of activity
trajectory in the open field test. (F) Representative images of NeuN staining
(green) in the cortex and hippocampus (CA1, DG), counterstained with DAPI (blue).
Magnification, 200
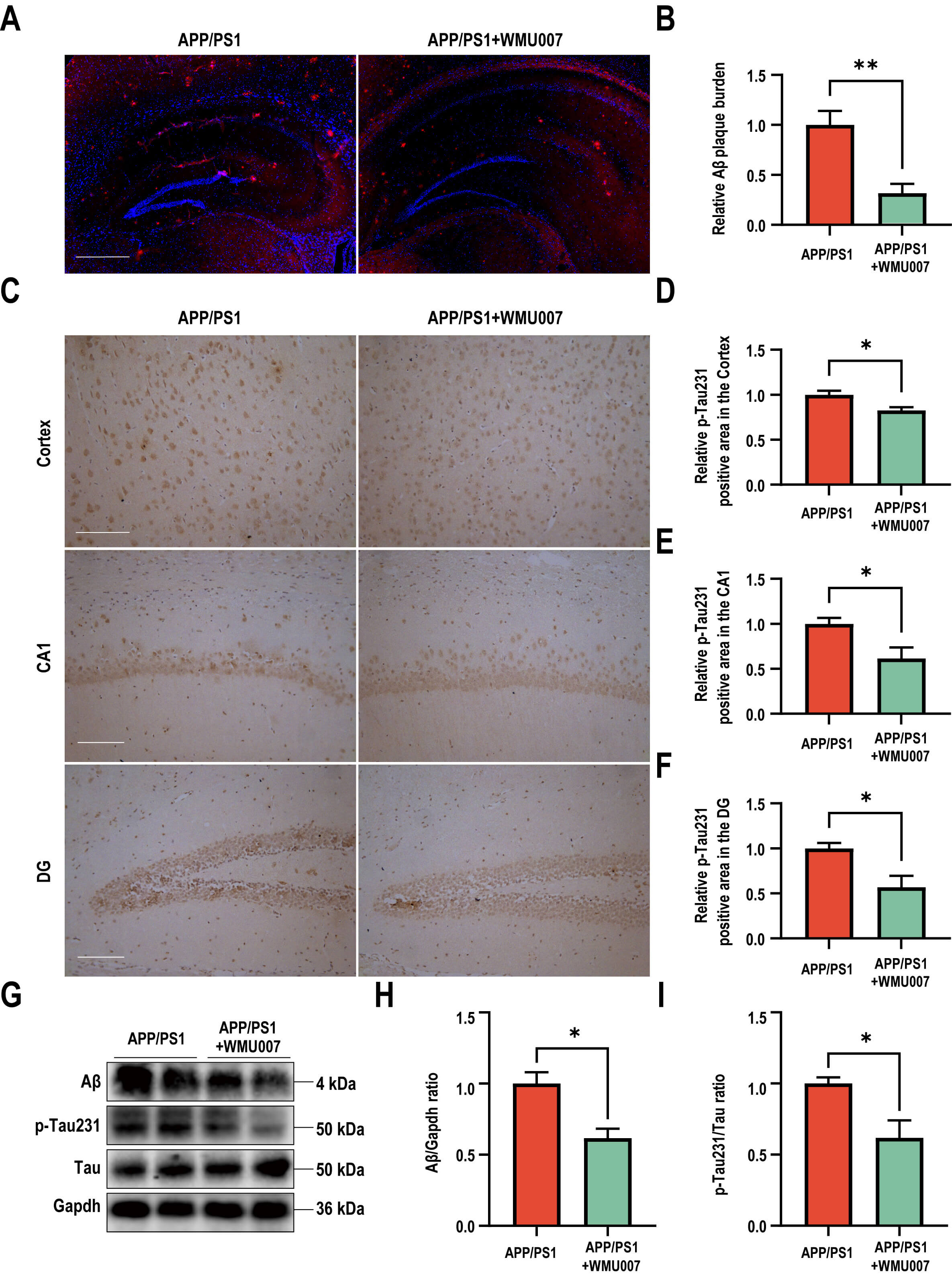
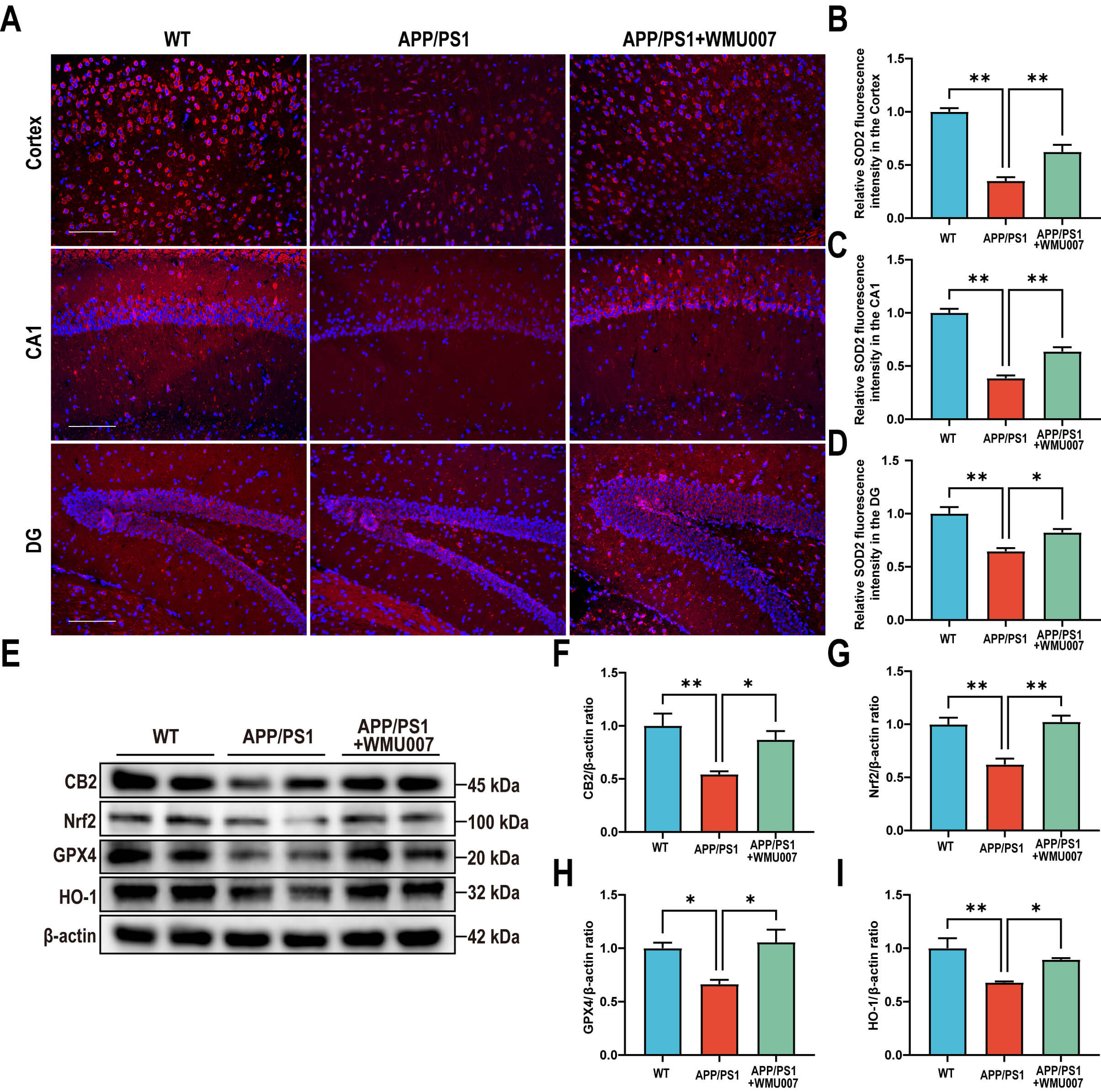
To assess the effects of L. mucosae WMU007 on neuropathological
changes, NeuN staining, A
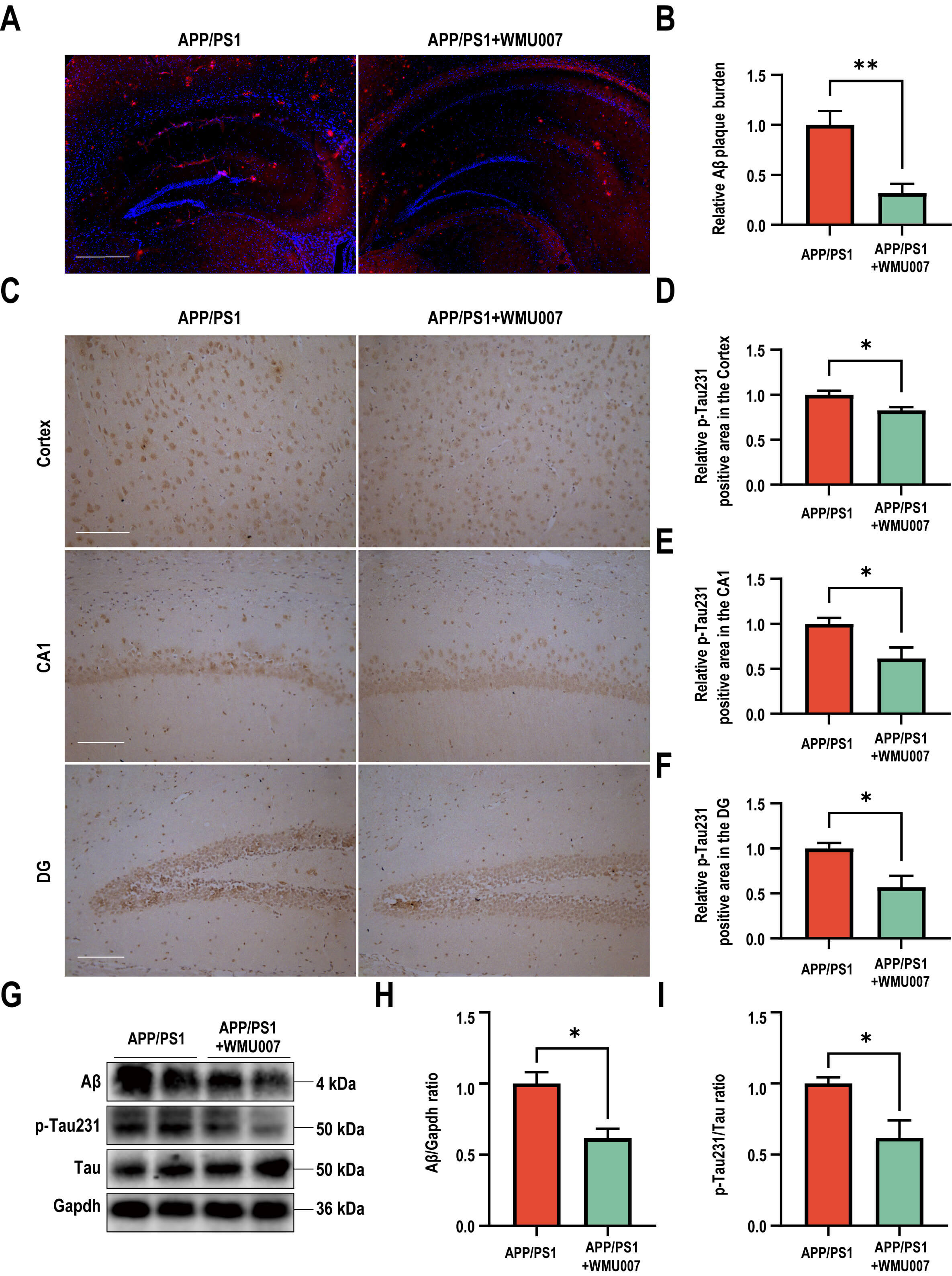 Fig. 2.
Fig. 2.
L. mucosae WMU007 treatment improved neuronal damage
and decreased A
To elucidate the molecular mechanism of L. mucosae WMU007 underlying
the decreased A
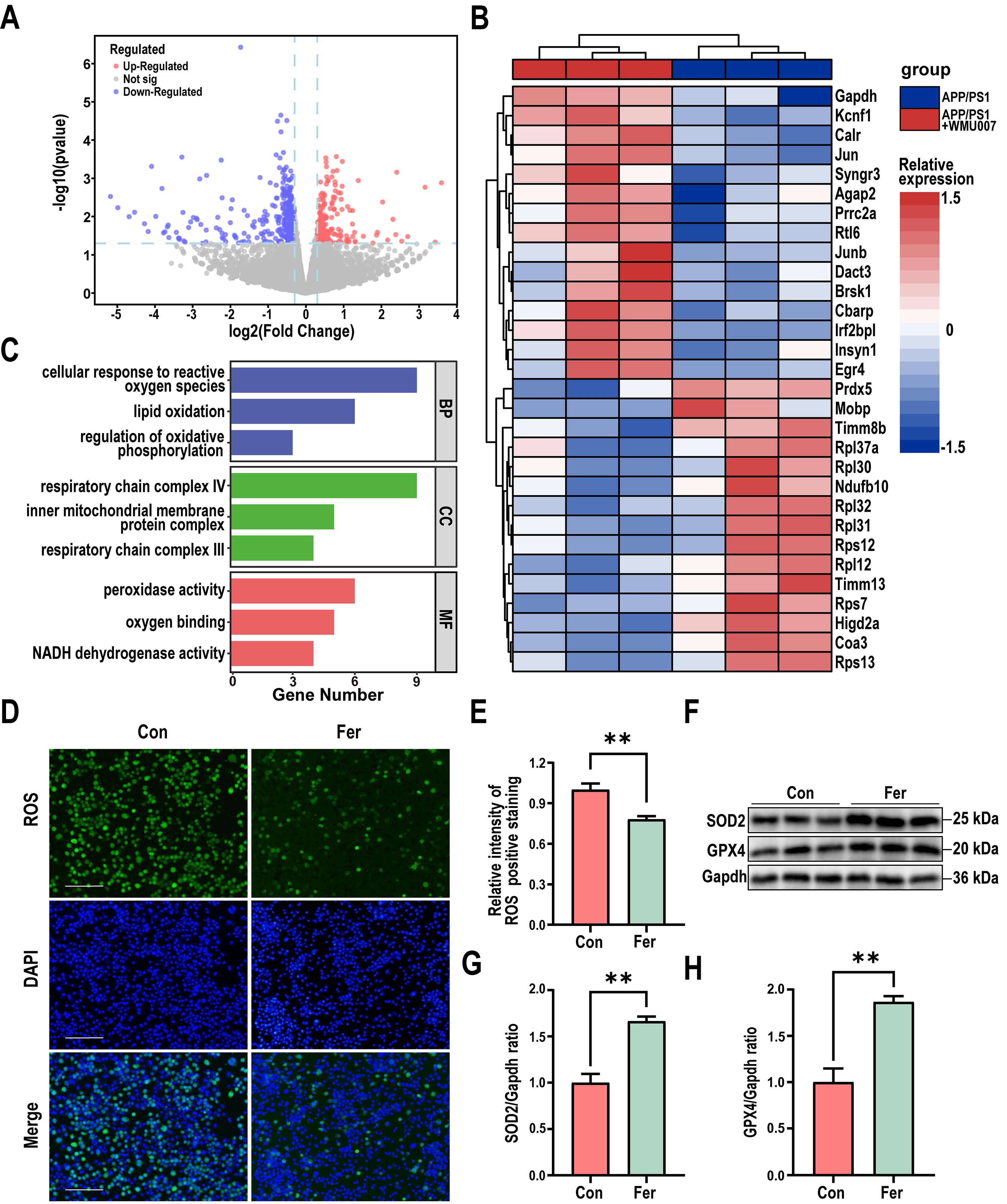
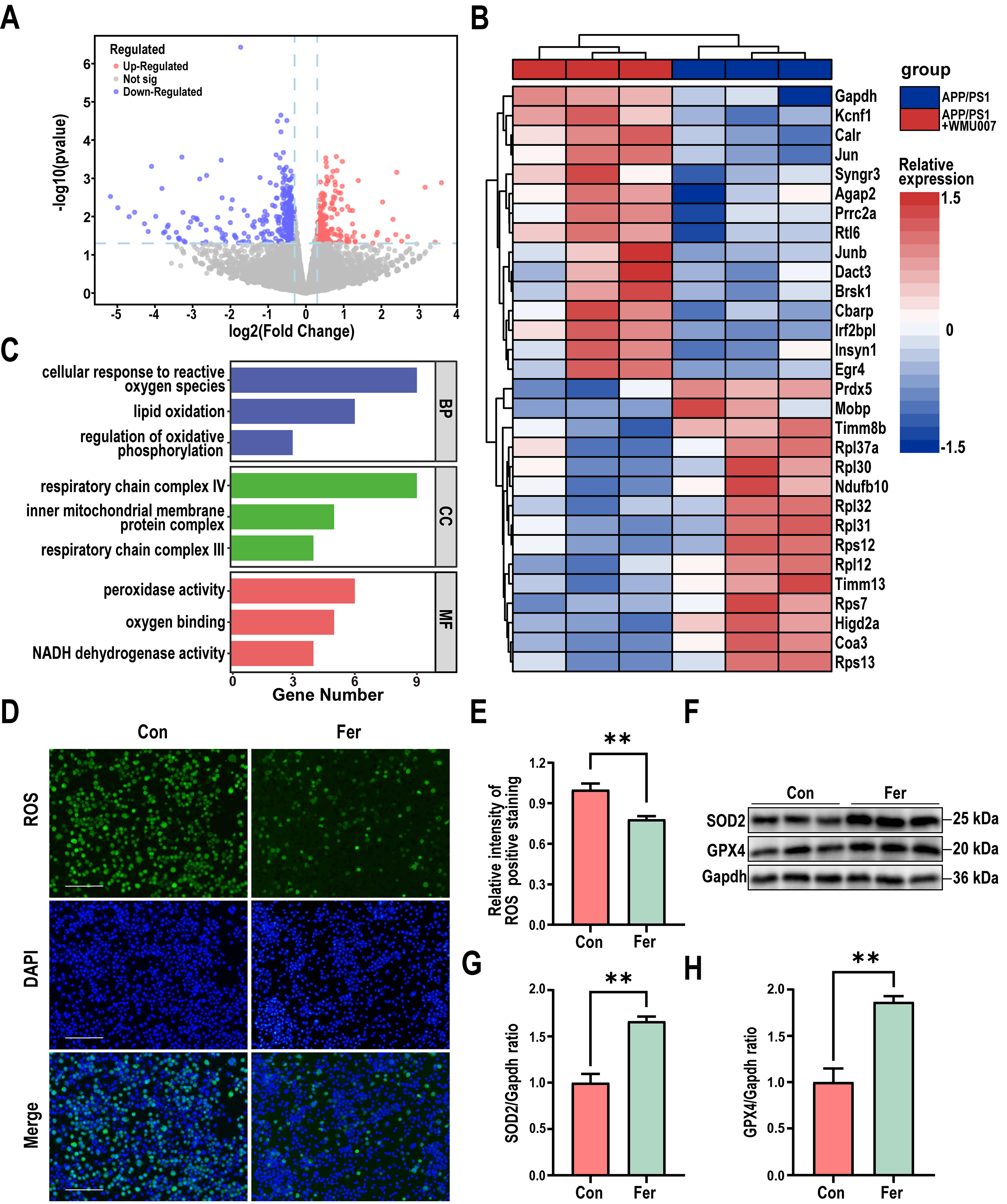 Fig. 3.
Fig. 3.
L. mucosae WMU007 treatment decreased oxidative stress. (A) Volcano plot showing the distribution of gene expression
level differences. p-value
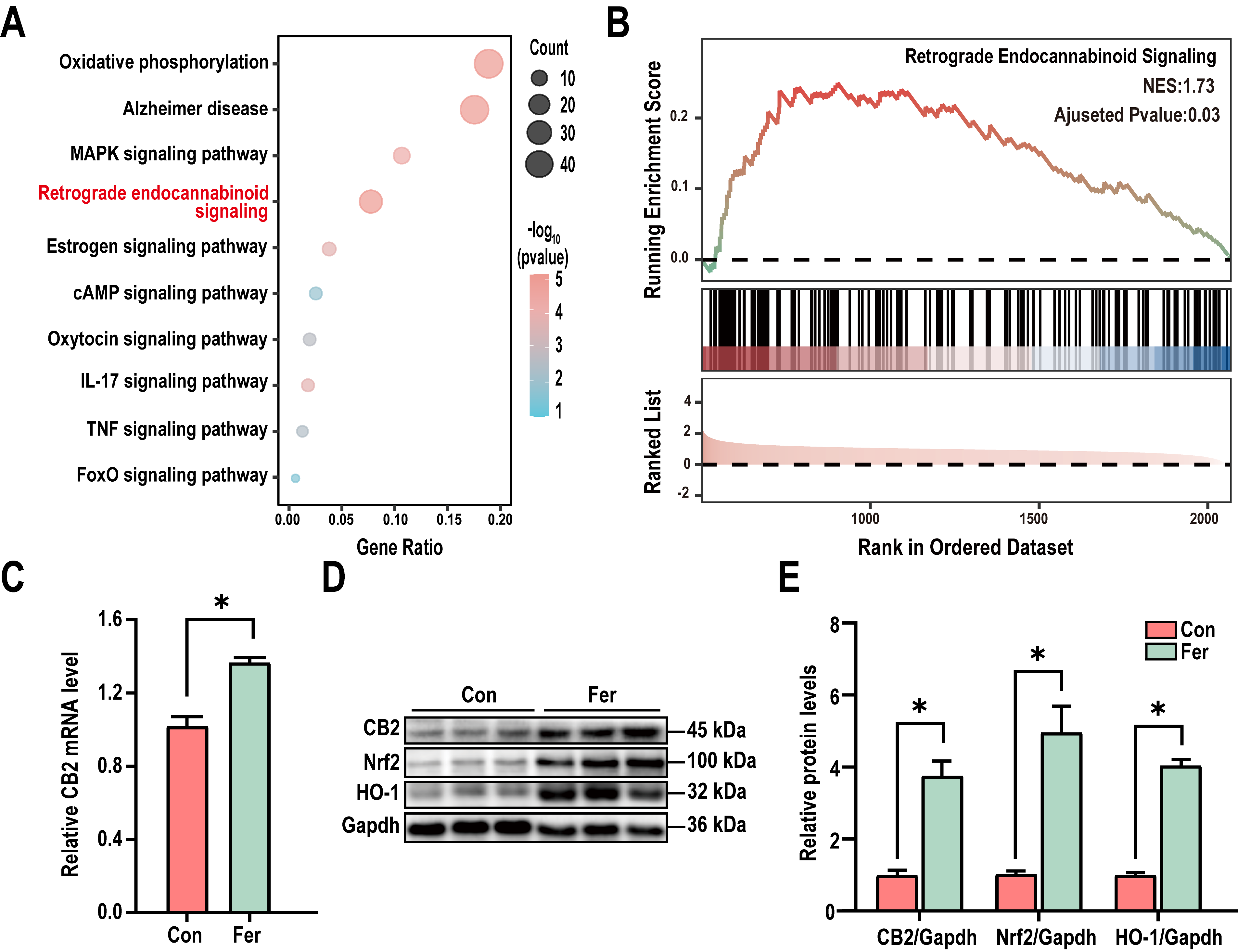
To further investigate the mechanism of L. mucosae WMU007 on inhibiting
oxidative stress, KEGG enrichment analysis was conducted based on genes
exhibiting altered expression in the RNA-Seq results. KEGG analysis revealed
significant differences in the Retrograde endocannabinoid signaling among the
DEGs (Fig. 4A). To gain a more comprehensive understanding of the changes in the
Retrograde endocannabinoid signaling, we conducted GSEA enrichment analysis. A
normalized enrichment score (NES) of 1.73 indicates that L. mucosae
treatment can significantly activate the aforementioned pathway (Fig. 4B). As
shown in Fig. 4C, the qPCR analysis revealed a significant elevation of CB2transcript abundance in mice receiving L. mucosae WMU007 relative to
controls (p
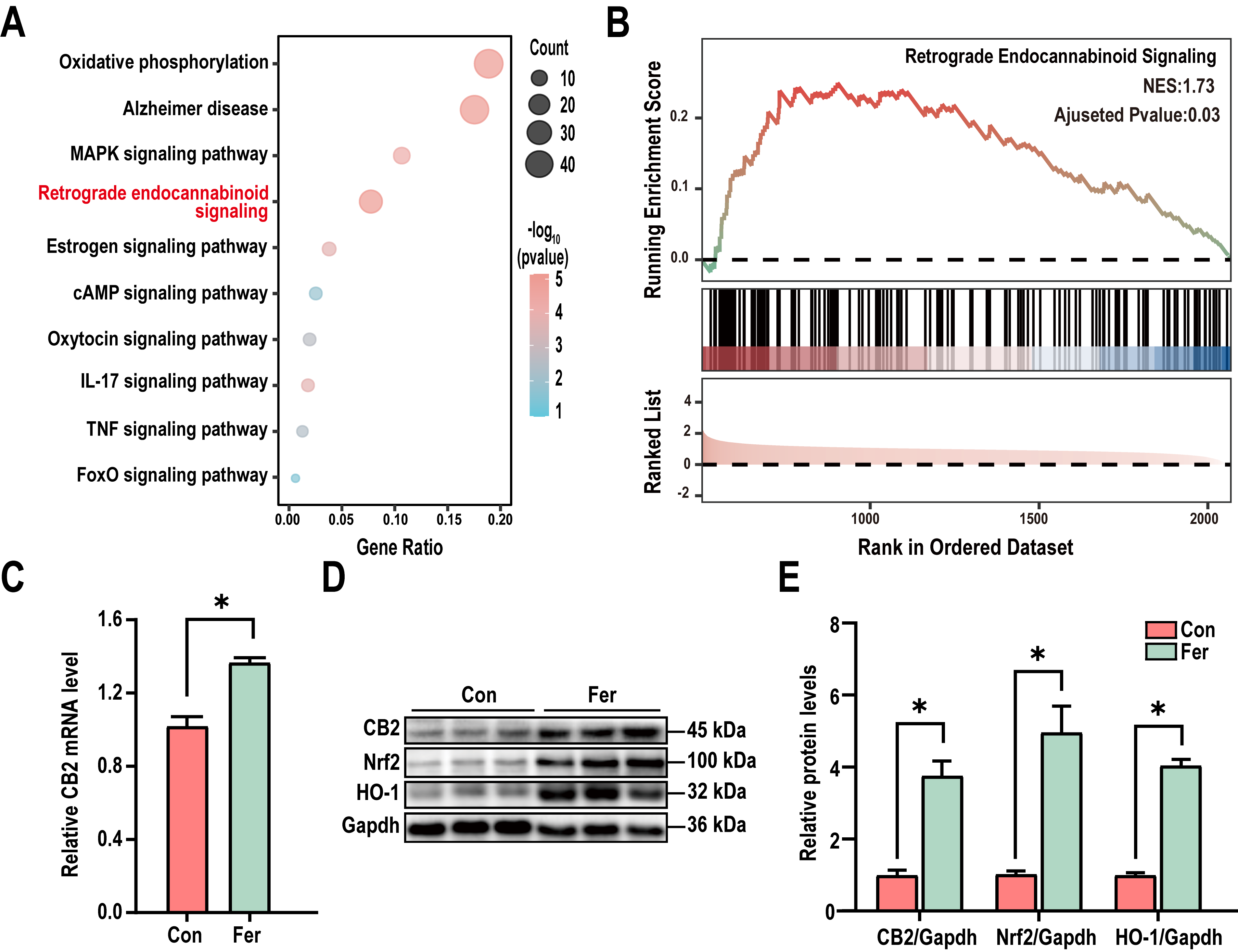 Fig. 4.
Fig. 4.
L. mucosae WMU007 inhibited oxidative stress via
activating the CB2 pathway. (A) KEGG pathway enrichment showing the Retrograde
endocannabinoid signaling was significantly enriched in DEGs. p-value
To confirm the role of L. mucosae WMU007 in inhibiting oxidative stress
through CB2 receptor activation, APP/PS1 mice were treated with L.
mucosae WMU007. Immunofluorescence staining revealed a significant increase in
SOD2-positive fluorescence intensity in the cortex, CA1, and DG of the L.
mucosae WMU007-treated group compared to the APP/PS1 group (Fig. 5A–D). To
further assess antioxidant responses and CB2 signaling, we analyzed protein
expression by Western blot. Our results showed that L. mucosae WMU007
treatment significantly increased the levels of CB2, Nrf2, GPX4 and HO-1 compared
to the APP/PS1 group (CB2: p
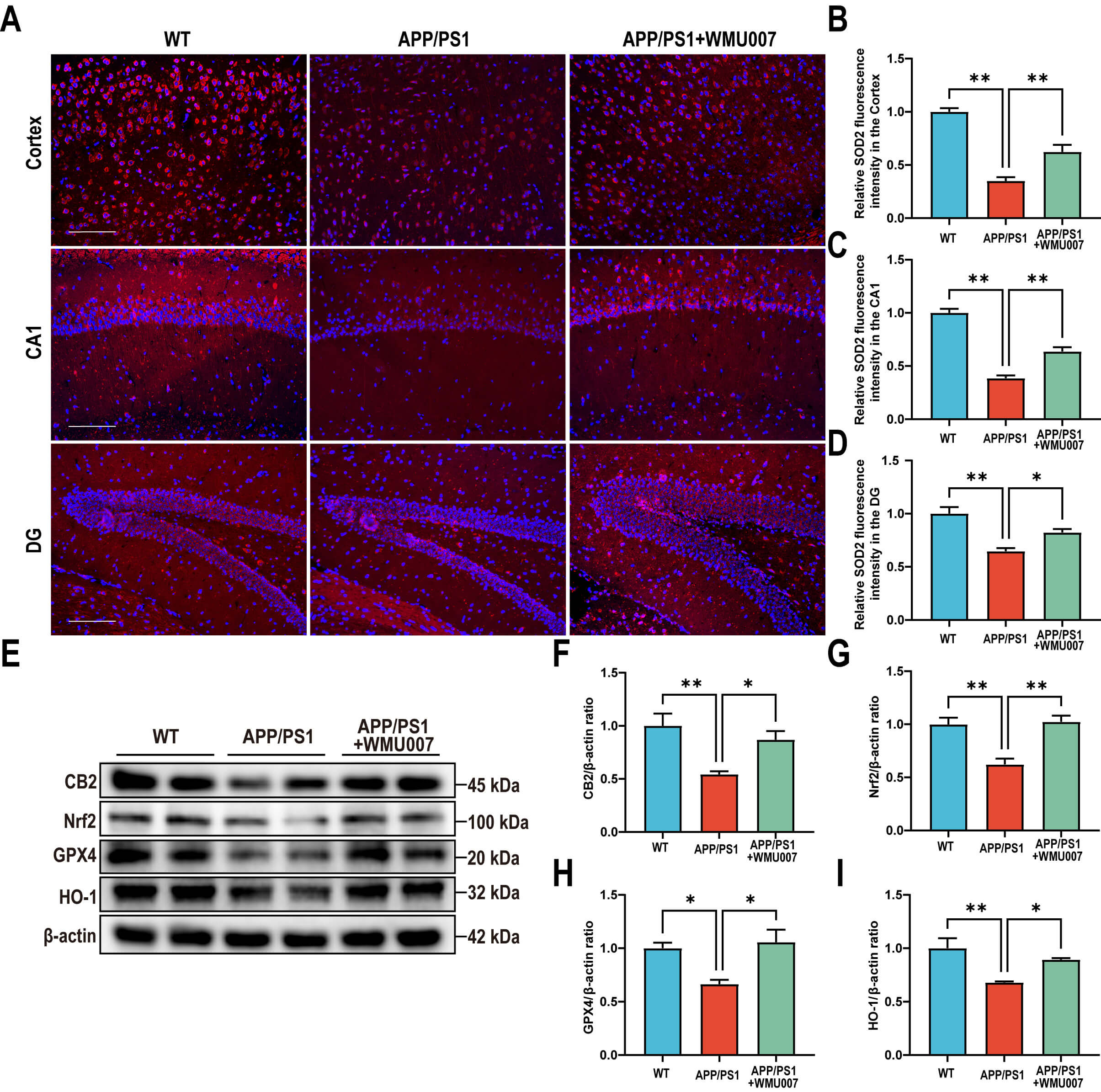 Fig. 5.
Fig. 5.
L. mucosae WMU007 treatment reduces oxidative stress in
APP/PS1 mice via activating the CB2 pathway. (A) Representative
immunofluorescence images of SOD2 (red) in the cortex and hippocampal (CA1 and
DG) regions, counterstained with DAPI (blue). Magnification, 200
This study revealed that L. mucosae WMU007 confers protective effects against oxidative stress in AD. Treatment with L. mucosae WMU007 significantly alleviated cognitive impairment, neuronal damage, and AD-related pathological changes in APP/PS1 mice. We found that L. mucosae WMU007 was involved in regulating oxidative stress, a key factor in the progression of AD. Furthermore, activation of the CB2 receptor was identified as the key mechanism by which L. mucosae WMU007 inhibits oxidative stress in AD. Supplementation with L. mucosae WMU007 was confirmed to inhibit oxidative stress and increase CB2-associated protein levels, both in vivo and in vitro. The above findings suggest that L. mucosae WMU007 may inhibit oxidative stress in AD through the CB2-mediated pathway.
Our study found that L. mucosae WMU007 could ameliorate cognitive
impairment, reduce neuronal damage, and decrease A
We identified CB2 activation as a key mechanism for inhibiting oxidative stress
in AD. As members of the G-protein coupled receptor family, cannabinoid receptors
were shown to participate extensively in central nervous system diseases [27].
Importantly, the expression level of CB2 has been associated with the
A
Although the results supported protective effects of L. mucosae WMU007 in the AD models and suggested that CB2 activation might be a key mechanism, several limitations still need to be addressed. This study only used male mice; future research should include female animals to assess potential differences. Further long-term observations should be evaluated potential adverse effects.
In conclusion, this study revealed that L. mucosae WMU007 might be involved in the inhibition of oxidative stress by activating the CB2, ultimately improving the pathogenesis of AD. Our study provided insights into the association between L. mucosae and oxidative stress in AD, suggesting that specific probiotics may become a promising therapeutic target for AD.
The datasets used during the current study are available from the corresponding author on reasonable request.
JLiu and JLi conceived and designed the experiments. YK, XLv, YY, QL, CD and XLin performed the experiments and conducted the statistical analyses. All authors contributed to editorial changes in the manuscript. All authors have read and agreed to the published version of the manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All experiments were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and under the guidance of the Animal Experiment Ethics Committee of First Affiliated Hospital of Wenzhou Medical University (No. WYYY-AEC-YS-2024-0378).
Not applicable.
This work was supported by Science and Technology Funds of Wenzhou (Y20240038).
The authors declare no conflicts of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/JIN48598.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.