



 , Bo Ning 2,†, Shengjie Wang 3, Wenshuo Chen 1, Lianqin Zhang 1, Chunxiu Chen 1, Junfeng Qiu 1,3,*
, Bo Ning 2,†, Shengjie Wang 3, Wenshuo Chen 1, Lianqin Zhang 1, Chunxiu Chen 1, Junfeng Qiu 1,3,* , Zhiming Hong 1,3,*
, Zhiming Hong 1,3,*1 The Fourth Clinical Medical College, Guangzhou University of Chinese Medicine, 518033 Shenzhen, Guangdong, China
2 The Second Clinical College, Guangzhou University of Chinese Medicine, 510006 Guangzhou, Guangdong, China
3 Department of Andrology, Shenzhen Traditional Chinese Medicine Hospital, 518033 Shenzhen, Guangdong, China
†These authors contributed equally.
Abstract
Premature ejaculation (PE) accompanied by anxiety or depression is a complex clinical condition at the intersection of male reproductive dysfunction and emotional disorders. Increasing evidence suggests that serotonin (5-HT) and brain-derived neurotrophic factor (BDNF) play central and interrelated roles in its pathogenesis. In this review we examine the bidirectional functions of 5-HT and BDNF in both the reproductive and nervous systems, highlighting their importance in regulating ejaculation, emotional stability, and synaptic plasticity. A comprehensive literature search (2010–2025) was conducted across multiple databases using relevant Medical Subject Headings (MeSH) terms, including pertinent original research and review articles, to synthesize the roles and regulatory pathways of 5-HT and BDNF in PE with comorbid anxiety or depression. We summarize the shared and distinct roles of 5-HT and BDNF in maintaining physiological balance across these systems and focus on their involvement in the major pathological processes underlying PE with anxiety or depression, including neurotransmitter imbalance, neuroendocrine dysregulation, inflammation, and oxidative stress. Furthermore, we outline the related signaling pathways through which 5-HT and BDNF exert their effects and interact. We also evaluate current pharmacological and non-pharmacological interventions targeting these molecules, demonstrating their potential to improve both ejaculatory control and emotional symptoms, and critically appraise selective serotonin reuptake inhibitor (SSRI)-related risks and highlighted the need for individualized dosing and monitoring. Emerging evidence suggests that Traditional Chinese Medicine formulations can extend intravaginal ejaculatory latency and mitigate mood symptoms and may serve as stand-alone or adjunctive options to reduce reliance on selective serotonin reuptake inhibitors (SSRIs). Overall, 5-HT and BDNF are not only deeply involved in the biological mechanisms of PE with comorbid psychological disorders, but also represent promising biomarkers and therapeutic targets, and their integrative neuro-reproductive regulatory functions provide new insights into the diagnosis and treatment of this multifaceted condition.
Keywords
- anxiety
- biomarkers
- brain-derived neurotrophic factor
- depression
- premature ejaculation
- serotonin
Premature ejaculation (PE) is one of the most common sexual dysfunctions in men.
According to the 2014 edition of the International Society for Sexual Medicine
(ISSM) guidelines for the diagnosis and treatment of PE, the definition of PE
includes three elements: (a) lifelong premature ejaculation (LPE), which occurs
often or always before or within about 1 minute after the initiation of sexual
activity. A significantly shortened intravaginal ejaculatory latency (IELT),
usually less than 3 minutes, is called acquired premature ejaculation (APE); (b)
these two types of PE patients cannot control and delay ejaculation almost every
time they had sex; (c) PE brings negative physical and mental effects to
patients, such as distress, anxiety, depression, interpersonal barriers, or
avoidance of sexual life [1]. The American Urologist Association/Sexual Medicine
Association of North America 2022 guideline further distinguishes between LPE and
APE. LPE is defined as poor ejaculation control with distress, and ejaculation
within approximately 2 minutes after initial sexual activity; APE is defined as
persistent poor ejaculatory control, associated distress, and significantly
reduced IELT from prior sexual experience during insertion [2]. The 2021 clinical
management guidelines for PE issued by the Italian Society of Andrology and
Sexual Medicine (SIAMS) further delineated two important subtypes: subclinical
premature ejaculation (SPE) and loss of control over erection and ejaculation
(LCEE) [3]. SPE is characterized by subjectively inadequate ejaculatory control
despite a normal objective IELT, accounting for approximately 15–20% of
patients with suspected PE [4]. LCEE refers to the coexistence of PE with
erectile dysfunction (ED) or subclinical ED; it occurs in up to 50% of patients
with PE, and the two conditions exacerbate one another, creating a
pathophysiological cycle [4, 5]. The 2025 European Association of Urology (EAU)
Guidelines did not propose their own definition of PE; instead, they reported the
ICD-11 definition based on the ISSM criteria and acknowledge variable PE and
subjective PE as potential subtypes [6]. However, an additional and often
overlooked presentation exists in clinical practice: some men have normal sexual
performance at times, yet on other occasions experience a sudden, uncontrollable
rapid ejaculation that can catch their partners off guard—this is a new concept
introduced in 2025, termed intermittent premature ejaculation (IPE) [7]. A survey
of 1976 Chinese men aged 18 to 50 years found a prevalence of PE of 2.3% [8]. In
a study of 62,675 Danes aged 15 to 89, 10.0% of men had experienced PE in the
previous year [9]. A health study of 2500 men in Bavaria, Germany, showed that
the prevalence of PE was 5.2% [10]. In two surveys of samples in five countries
(Turkey, the Netherlands, Spain, the United Kingdom, and the United States), only
2.95% of men had an IELT
Modern medicine has many views on the pathogenesis of PE. Initially, Jannini et al. [20] believed that PE had psychological factors or an interpersonal basis, and it was caused by anxiety or lack of early sexual life experience. Recent evidence has suggested that it may be related to a disorder of serotonin (5-HT) neurotransmitter in the central nervous system (CNS), reduced synthesis of brain-derived neurotrophic factor (BDNF), hypersensitivity of the glans penis, psychology, and other factors [21]. Among these factors, 5-HT is closely related to mental and psychological disorders such as anxiety or depression [22]. It is also the most widely studied inhibitory neurotransmitter in the regulation of ejaculation, and an increase in central 5-HT concentration can lead to a delay in ejaculation [23]. Selective serotonin reuptake inhibitors (SSRIs) have been used in the treatment of mental disorders, and have the side effect of delayed ejaculation. The mechanism of their action is to inhibit the reuptake of 5-HT by 5-hydroxytryptamine transporter (5-HTT), to increase the concentration of 5-HT in the synaptic cleft, and to delay ejaculation. Therefore, this class of drugs is also used in the treatment of PE [24].
BDNF is the most important member of the neurotrophic factor family. It is widely distributed in the CNS of mammals. It plays a very important role in the development and maintenance of the CNS, and can supply a variety of neurons such as 5-HT, dopamine (DA), and cholinergic neurons, and multiple brain areas such as raphe nuclei, prefrontal cortex, and hippocampus [25, 26]. A clinical study conducted by Atik et al. [27] showed that serum BDNF levels in PE patients were significantly lower than those in healthy subjects, and with the decrease of serum BDNF levels, PE diagnostic scale scores significantly rose, suggesting that BDNF deficiency may be a potential predisposing factor for PE. In addition to its neurotrophic functions, BDNF is critically involved in the regulation of emotional states such as anxiety and depression by modulating neuronal growth, survival, apoptosis, neuroinflammatory responses, and synaptic plasticity [28]. The relationship between BDNF and 5-HT is complex and reciprocal, and Guiard et al. [29] demonstrated that a decrease in BDNF levels leads to an increase in baseline exogenous 5-HT levels in the ventral hippocampus of mice. However, Deltheil et al. [30] found that local injection of BDNF reduced exogenous 5-HT levels, and this effect was abolished by K252a, a tropomycin-receptor-kinase B (TrkB) inhibitor. In addition, BDNF may cooperate with 5-HT homeostasis through TrkB signaling to promote the neurogenic effects of SSRIs [31]. The above studies suggest some degree of interaction between BDNF and 5-HT. The frequent coexistence of PE with anxiety or depression suggests a common pathological basis in which 5-HT and BDNF serve as key molecular mediators. They may lead to PE accompanied by anxiety or depression through a series of complex physiological and pathological processes.
In the present review, first introduced is the correlation of 5-HT and BDNF with male reproductive system and nervous system function. In the male reproductive system, 5-HT plays a pivotal role in regulating ejaculation, penile erection, and sperm quality through the modulation of intracellular messengers and hormones, including cAMP, testosterone, endothelin-1 (ET-1), and norepinephrine (NE). BDNF regulates sperm production, erectile function, and the ejaculation mechanism by activating multiple signaling pathways such as phosphatidylinositol-3-kinase (PI3K)/Akt and mitogen-activated protein kinase (MAPK). In the nervous system, 5-HT regulates emotion and cognitive function by regulating the 5-hydroxytryptamine 1A (5-HT1A) receptor and the 5-hydroxytryptamine 2A (5-HT2A) receptor. By influencing key brain regions such as the hippocampus and prefrontal cortex, BDNF contributes significantly to the neural mechanisms underlying anxiety and depression. Second, the review focuses on the latest research progress of the role of 5-HT and BDNF in PE with anxiety or depression, including the specific mechanisms of 5-HT and BDNF in regulating serotonergic system abnormalities, regulating dopaminergic system abnormalities, regulating inflammatory response, improving oxidative stress damage, and mediating multiple signaling pathways. Last, the review summarized the potential of 5-HT and BDNF as potential biomarkers for the treatment of PE with anxiety or depression.
This review adopted a narrative synthesis approach to systematically identify, screen, and integrate research evidence published since 2010 on the mechanistic roles, biomarker value, and therapeutic potential of 5-HT and BDNF in PE with comorbid anxiety or depression. We also considered established theories and emerging directions in the field to ensure scientific rigor and currency.
We conducted comprehensive searches of PubMed (https://pubmed.ncbi.nlm.nih.gov), Embase (https://www.embase.com), the Web of Science Core Collection (https://www.webofscience.com), and ScienceDirect (https://www.sciencedirect.com). In addition, we hand-searched the reference lists of included articles to capture seminal pre-2010 studies on 5-HT and BDNF regulation of male reproductive or nervous system function. Database searches covered January 2010 to August 2025; the hand search had no time restriction to ensure completeness of foundational work.
We combined keywords and MeSH and optimized the queries using Boolean operators (AND/OR). Representative terms included: disease-related: “premature ejaculation”, “PE”, “anxiety”, “depression”; biomarker-related: “serotonin”, “5-hydroxytryptamine”, “5-HT”, “brain-derived neurotrophic factor”, “BDNF”, “biomarker”; mechanism and therapy: “pathogenesis”, “mechanism”, “signaling pathway”, “treatment”, “selective serotonin reuptake inhibitors”, “SSRIs”, “traditional Chinese medicine”. To improve precision, we also included relevant modifiers such as “dopaminergic”, “oxidative stress”, and “inflammatory response”.
Study type: English-language original research (clinical studies and basic/preclinical experiments) and systematic reviews/meta-analyses;
Study population/models: clinical studies involving patients with PE, anxiety, or depression; preclinical studies employing animal models of PE, anxiety, or depression;
Relevance: studies explicitly examining associations of 5-HT or BDNF with PE, anxiety, or depression, including pathophysiology, diagnostic biomarker utility, or effects of targeted interventions;
Quality: published in peer-reviewed journals with complete data and clear methodological descriptions.
Study type: case reports, conference abstracts, editorials, letters, and records without full text;
Irrelevance: studies unrelated to PE or anxiety/depression or primarily focused on other mechanisms;
Quality issues: incomplete data, unclear methodology, or conclusions lacking statistical support;
Duplicates: when multiple publications from the same dataset were identified, only the most recent or most comprehensive report was included.
After deduplication, two reviewers independently screened titles and abstracts against the inclusion/exclusion criteria; disagreements were resolved through discussion with a third reviewer. Records deemed potentially eligible proceeded to full-text evaluation. Studies that merely described phenomena without mechanistic interrogation, or reported ambiguous efficacy data, were excluded to preserve the scientific robustness and persuasiveness of the synthesis.
Serotonin (5-HT) is a monoamine neurotransmitter that is synthesized and released by serotonergic neurons in the CNS and chromaffin cells in the gastrointestinal tract [32]. 5-HT is thought to affect a range of behavioral, physiological, and cognitive functions in the human body, such as memory, mental health, mood, sleep, appetite, sexual function, and temperature regulation [33]. The serotonergic nervous system is divided into the peripheral serotonergic system and the central serotonergic system. Peripheral 5-HT is thought to be involved in the regulation of physiological functions such as platelet aggregation, vasodilation, hypertension, and intestinal peristalsis [34]. In the CNS, 5-hydroxytryptophan (5-HTP) is generated from tryptophan (Trp) via tryptophan hydroxylase 2 (TPH2), and then 5-HT is generated by 5-hydroxytryptophan decarboxylase (5-HTPDC) [35]. The vesicular monoamine transporter (VMAT) is responsible for the transfer of 5-HT synthesized by presynaptic neurons from the cytoplasm to the vesicles, and then it is excreted to the synaptic cleft where it binds to presynaptic or postsynaptic receptors to play a physiological role [36]. 5-HT in the synaptic cleft is mainly recycled from 5-HTT to the presynaptic membrane, and then catabolized by monoamine oxidase (MAO) to form inactive 5-hydroxyindoleacetic acid (5-HIAA) [37]. 5-HT receptors are mainly divided into seven families (5-HT1 through 7), each of which is divided into different subtypes, for a total of 14 receptor subtypes. Except for 5-HT3 receptor, which is an ion-type gated channel protein, the rest are G-protein-coupled receptors (GPCRs) [38]. 5-HT receptors are highly distributed in areas related to emotion regulation and memory (such as hippocampus, amygdala, and cortex). By coupling with GPCRs, 5-HT receptors increase the expression of downstream cAM-response-element binding protein (CREB) and BDNF, thereby participating in the regulation of physiological activities [39].
BDNF is a member of the family of neurotrophin-secreting proteins that supports
neuronal survival, synaptic plasticity, and neurogenesis [40]. BDNF protein is
synthesized in neurons and glial cells, and then widely expressed in the CNS. It
is enriched in the hippocampus and cortex, especially in the CA3 region of the
hippocampus [41]. BDNF is synthesized in multiple forms—pro-pro-BDNF, pro-BDNF,
and m-BDNF—with m-BDNF serving as the principal effector in promoting neuronal
survival and differentiation [42]. The BDNF mRNA is first synthesized in the
endoplasmic reticulum and then transported to the Golgi apparatus, where it is
cleaved to remove the pro-region sequence to form pro-BDNF with certain
functional activity [43]. Then, pro-BDNF can then be converted into m-BDNF in the
cell through two different pathways. One, pro-BDNF is cleaved by furin, a member
of the Bacillus subtilis endonuclease family, to produce m-BDNF in the Golgi
network. Two, pro-BDNF can also form m-BDNF in intracellular vesicles through the
action of intracellular converting enzymes such as protein convertase. The m-BDNF
produced by the above two pathways is released to the outside of the cell through
the presynaptic membrane [44]. BDNF, secreted into synapses, activates the p75
neurotrophin receptor [p75(NTR)], TrkB, and their downstream signaling cascades.
Among them, p75(NTR)-mediated signaling pathways mainly include the nuclear
factor-
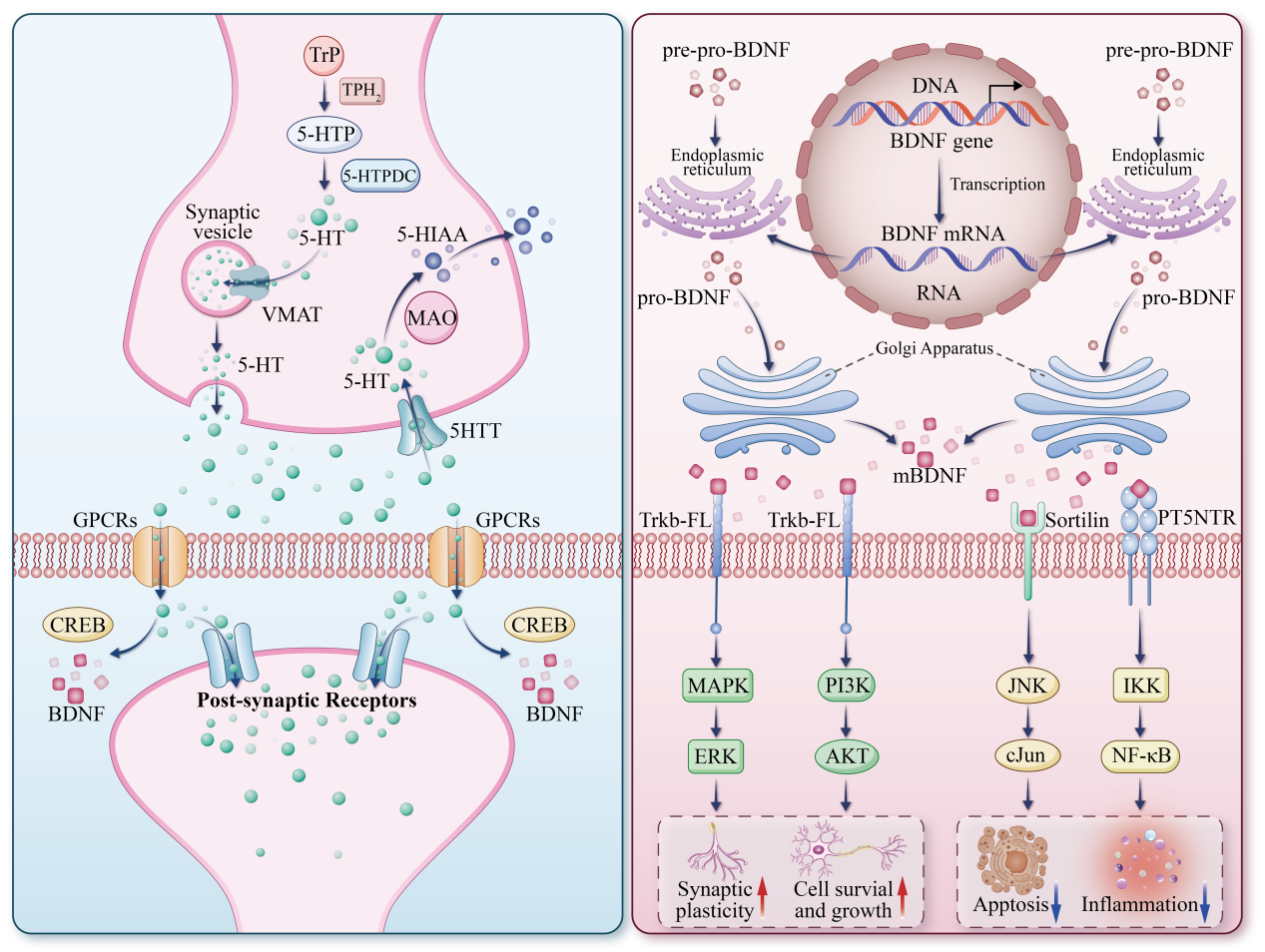 Fig. 1.
Fig. 1.
Physiological functions of 5-HT and BDNF. The downward arrow
indicates a decrease (downregulation) and the upward arrow represents an increase
(upregulation). 5-HT, 5-hydroxytryptamine; BDNF, brain-derived neurotrophic factor; Trp, tryptophan; TPH2, tryptophan hydroxylase 2; 5-HTP,
5-hydroxytryptophan; 5-HTPDC, 5-hydroxytryptophan decarboxylase; 5-HIAA,
5-hydroxyindoleacetic acid; MAO, monoamine oxidase; VMAT, vesicular monoamine
transporter; 5-HTT, 5-hydroxytryptamine transporter; GPCRs, G protein-coupled
receptors; CREB, cAMP response element-binding protein; TrkB, tropomycin receptor
kinase B; p75NTR, p75 neurotrophin receptor; PI3K, phosphatidylinositol-3-kinase;
MAPK, mitogen-activated protein kinase; JNK, c-Jun N-terminal Kinase;
NF-
5-HT and BDNF are key molecular regulators involved in maintaining normal physiological function within the male reproductive system. 5-HT affects male reproductive health by regulating ejaculation and erectile function. The disturbance of 5-HT in the CNS can lead to ejaculation disorders, and different receptors of 5-HT, such as 5-HT1A and 5-HT2A, play different roles in ejaculation and erection. For example, 5-HT2A receptor activation is associated with ED because it leads to blocked penile blood flow. In addition, 5-HT affects sperm motility and fertilization rate by regulating cAMP and testosterone levels. On the other hand, BDNF is equally important in the development and function of the male reproductive system. By binding to TrkB and p75(NTR), BDNF activates multiple signaling pathways (such as PI3K/Akt and MAPK) to promote the proliferation of Leydig cells, thereby increasing sex hormone levels and enhancing sperm motility. In addition, BDNF may be involved in the regulation of ejaculation by affecting 5-HT synthesis. Therefore, the interaction between 5-HT and BDNF, as well as their effects on male reproductive function, becomes an important direction of current research.
The effect of central 5-HT neurotransmitter disorder on ejaculation has been a research hotspot in recent years [48]. The neural areas involved in ejaculation include the spinal ejaculatory center, the ascending fibers of the spinal ejaculatory center and the descending fibers of some brain regions. Ejaculation is regulated by a variety of neurotransmitters including 5-HT [49]. The 5-HT system consists of 5-HT, 5-HTT and the 5-HT receptors. In the hypothalamic synaptic cleft, 5-HT is regulated by 5-HTT and regulates ejaculation by binding to the 5-HT receptor. The greater the transport activity of 5-HTT, the lower the 5-HT concentration in the synaptic cleft and the shorter IELT [50]. Different 5-HT receptors have different biological functions: 5-HT2 receptors inhibit ejaculation, whereas 5-HT1A receptors promote ejaculation [51]. This suggests that 5-HT, 5-HTT, and the 5-HT receptor are closely related to the occurrence of PE. It should be noted that the widespread distribution of the 5-HT system renders interventions prone to nonspecific effects. With clinical use of SSRIs, peripheral adverse effects such as hyperhidrosis and gastrointestinal discomfort are common, and excessive activation of central 5-HT receptors may precipitate a risk of serotonin syndrome, including symptoms such as anxiety or confusion [52]. In addition, 5-HT is also associated with ED. A related study has shown that diabetes can cause a significant increase in the serum concentration of 5-HT, which enhances the vasoconstriction effect of 5-HT through the 5-HT2A receptor, and this process leads to the blockage of penile blood flow, resulting in ED [53]. Studies have shown that 5-HT and the 5-HT2A receptor can indirectly enhance the vasoconstrictor effect of vasoconstrictor substances such as ET-1 and NE, and change the normal vasoconstriction activity of blood vessels [54, 55]. 5-HT elevates intracellular calcium levels in penile smooth muscle cells, thereby inducing muscle contraction. This effect can be suppressed by 5-HT2A receptor antagonists, implicating 5-HT and its 5-HT2A receptor in the development of ED [56]. Furthermore, 5-HT acts as a neural signal carrier capable of triggering the acrosome reaction in sperm, which in turn facilitates reproductive system maturation and enhances sperm functionality [57, 58]. In humans, 5-HT also controls sexual desire and regulates sexual behavior through hormonal action, and improves sperm motility and fertilization rate by regulating cAMP, testicular blood flow and testosterone in the reproductive system [59].
BDNF has a wide range of effects on the mammalian nervous system and gonadal development. It can enhance sperm motility, increase sex hormone levels, and promote the regeneration of blood vessels and nerve fibers. Robinson et al. [60] demonstrated the expression and cellular localization of BDNF and its receptors in fetal Leydig cells and adult testis by immunohistochemical methods.
Spermatogenic dysfunction is one of the important causes of male infertility by leading to decreased sperm quantity, decreased quality, or insufficient motility [61]. Safari et al. [62] showed that exogenous BDNF supplementation significantly enhanced sperm motility. Gao et al. [63] demonstrated that BDNF induced the proliferation of TM3 Leydig cells by upregulating proliferating cell nuclear antigen and enhancing the phosphorylation of extracellular signal-regulated protein kinase 1/2, thereby promoting sperm production by increasing sex hormone levels. The cavernous nerve (CN), originating from the pelvic plexus, is the main autonomic nerve that regulates penile erection. ED caused by cavernous nerve injury is a common complication of prostate cancer surgery or other pelvic surgery, and has a negative impact on the prognosis and quality of life of patients [64, 65]. During ED repair and regeneration, stem cells promote angiogenesis and nerve fiber regeneration by releasing active factors such as vascular endothelial growth factor (VEGF) and BDNF. Chen et al. [66] injected BDNF into the cavernous body of CN-injured rats, and the erectile function of the treated group was improved over that of the control group. Lin et al. [67] used a combination of different molecular pathway inhibitors in the presence of BDNF and showed that the Janus kinase/signal transducer and activator of transcription (JAK/STAT) signaling pathway was the main signal transduction pathway for BDNF to promote CN fiber growth. In addition, BDNF expression may also be involved in the regulation of the male ejaculation mechanism. Huang et al. [68] observed that rapid ejaculation in rats was associated with reduced TPH2 expression, decreased 5-HT synthesis, and diminished endogenous BDNF levels in the raphe nuclei, indicating that BDNF may modulate ejaculatory function by enhancing central 5-HT synthesis.
Within the nervous system, 5-HT and BDNF contribute significantly to the regulation of mood and cognition. 5-HT neurons are mainly located in the raphe nucleus of the brain stem and project to a variety of areas such as substantia nigra, striatum, and prefrontal cortex, which are involved in the regulation of cognition and emotion. Depression and anxiety are closely related to changes in 5-HT levels in the brain, especially to the dysfunction of 5-HT1A and 5-HT2A receptors. The role of the 5-HT1A receptor in anxiety regulation is prominent, and its reduction may lead to mood disorders, whereas the 5-HT2A receptor is closely linked to the symptoms of schizophrenia. Relatively high levels of BDNF expression in the hippocampus and prefrontal cortex have also been associated with emotional stability. The decrease of BDNF has been significantly associated with the pathogenesis of various psychological disorders such as depression and anxiety disorders. In addition, BDNF genetic polymorphisms such as Val66Met have been significantly associated with the development of anxiety disorders, and low serum BDNF levels may be a biomarker for anxiety disorders. In summary, emotional and cognitive processes within the nervous system are profoundly influenced by 5-HT and BDNF, whose dysregulation has been implicated in the etiology of multiple psychiatric conditions.
In the brain, the cell bodies of 5-HT neurons are primarily distributed in the raphe nuclei of the brainstem, with efferent projections extending to the substantia nigra, striatum, globus pallidus, hypothalamus, thalamus, and cortex, which are regions critically involved in cognitive and emotional processing [69]. The effects of 5-HT on cognition and emotion regulation are achieved through a number of different 5-HT receptors, and almost all 5-HT receptor subtypes are involved in the regulation of depression or anxiety [70]. The 5-HT1A receptor is the receptor subtype that has been studied more in anxiety. Patients with panic disorder have been found to have reduced 5-HT1A receptors in the forebrain and raphe nucleus [71]. Mice with anxiety-like behaviors were also found to have reduced 5-HT1A receptors in the amygdala, suggesting that 5-HT1A receptors are involved in the regulation of anxiety-like behaviors [72]. The decrease of 5-HT level in the brain is considered to be the main pathogenic factor of depression [73]. 5-HT1A receptors exert inhibitory effects on serotonergic neurons, so these autoreceptors may limit the response of serotonergic neurons to afferent excitation. Chen et al. [74] showed that mice develop depressive-like behaviors in response to uncontrollable stress, which is associated with up-regulation of 5-HT1A autoreceptors. However, when mice face repetitive, predictable stress, they adapt to the stress and down-regulate 5-HT1A, resulting in increased extracellular 5-HT concentration and increased transmission of monoamine neurotransmitters.
Nevertheless, the extensive projection patterns of serotonergic neurons make pharmacological interventions susceptible to off-target effects on non-target brain regions; in addition to therapeutic actions, they may provoke central adverse effects—such as insomnia and appetite loss—by disrupting sleep-wake cycles or appetite-regulatory pathways [75]. In addition, 5-HT dysfunction is associated with schizophrenia. Lysergic acid diethylamide (LSD) causes schizophrenia-like hallucination and delusional symptoms by activating 5-HT receptors, particularly 5-HT2A receptors [76]. Diez-Alarcia et al. [77] used three radiotracers with different pharmacological properties to study the conformation of the 5-HT2A receptor in the prefrontal cortex (PFC) of patients with schizophrenia. They found that under normal physiological conditions, the active conformation of the 5-HT2A receptor is in dynamic equilibrium with the inactive conformation. However, the proportion of 5-HT2A receptors in the active conformation is higher in the brain of schizophrenia patients, and the dynamic balance is broken, causing 5-HT2A receptors to be in a state of excessive activation.
Under normal circumstances, BDNF content is high in hippocampus, prefrontal
cortex, and other brain regions related to emotion, and is closely related to the
regulation of a variety of emotions. Stress can lead to a decrease in BDNF
levels. In rats experiencing unpredictable, chronic mild stress, proBDNF is
upregulated in the neocortex and hippocampus, thereby inducing depression-like
behaviors [78]. Injection of anti-probDNF antibody reversed depression-like
behaviors in chronic-stress rats [79]. BDNF has a pathway with a number of
molecules such as TrkB, p75(NTR), peroxisome proliferator-activated receptor
gamma coactivator-1
At present, the relationship of the pathological mechanism of PE with anxiety or
depression is believed to be closely related to abnormalities in the serotonergic
system, imbalance of dopaminergic system, neuroinflammatory response, and
oxidative stress injury. The abnormality in the serotonergic system mainly leads
to the disorder of ejaculation control and to anxiety and depression through
insufficient synthesis or a metabolic disorder of 5-HT. The imbalance of the
dopaminergic system is mainly manifested as excessive activation of the DA signal
to accelerate the ejaculatory reflex, which interacts with 5-HT to regulate
emotion. The neuroinflammatory response is mainly caused by the interference of
proinflammatory factors (IL-6, TNF-
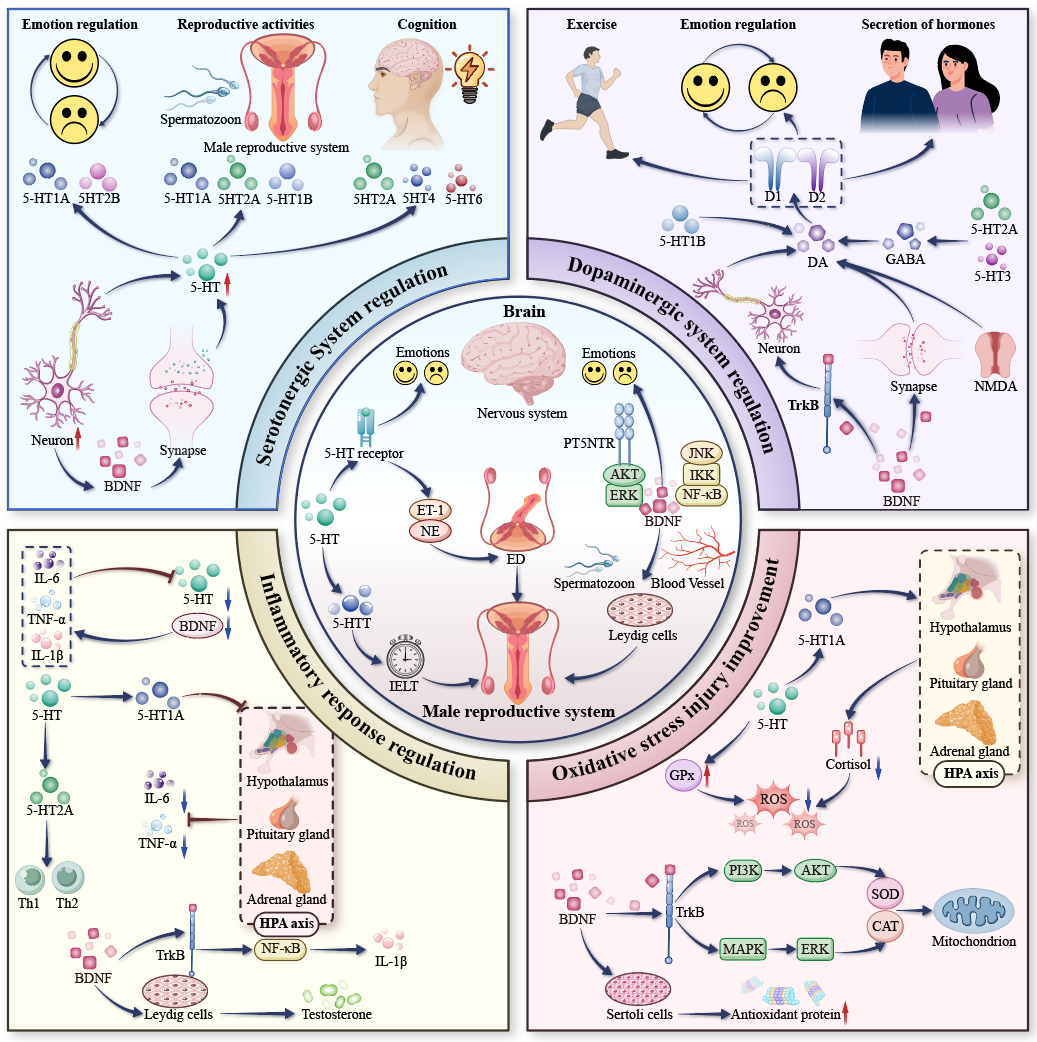 Fig. 2.
Fig. 2.
The mechanistic role of 5-HT and BDNF in PE with anxiety or
depression. The downward arrow indicates a decrease (downregulation), and the
upward arrow represents an increase (upregulation). DA, dopamine; GABA,
gamma-aminobutyric acid; TrkB, tropomycin receptor kinase B; NMDA,
N-methyl-D-aspartate; HPA, hypothalamic-pituitary-adrenal; ED, erectile dysfunction; IELT, intravaginal
ejaculatory latency time; ET-1, endothelin-1; NE, norepinephrine; IL-6, Interleukin-6; TNF-
The serotonergic system is an important neurotransmitter system in the human body. With serotonin as the core, it is widely involved in the regulation of emotion, cognition, sleep, and other physiological and psychological functions [88]. The cell bodies in the raphe nuclei neurons are the main source of 5-HT in the brain. They are divided into nine nuclei, named B1-B9. Most of these nuclei contain 5-HT neurons. However, ascending fibers from more medullary clusters (B5-B9) are connected with cerebral cortex, hypothalamus, thalamus, basal ganglia, hippocampus, etc. [89]. Because of this extensive neuronal network, 5-HT may be viewed as one of the major regulators of the CNS. 5-HT directly regulates the serotonergic system by activating different receptors, these include reproductive activities (5-HT1A, 5-HT2A, 5-HT1B, etc.), emotional regulation (5-HT1A, 5-HT2B, etc.), appetite and digestion (5-HT2B, 5-HT4, etc.), and pain perception (5-HT1A, 5-HT2B, 5-HT1F, etc.), cognition and decision making (5-HT2A, 5-HT4, 5-HT6, etc.) [90]. Although BDNF has an indirect regulatory effect on the serotonergic system, BDNF can support the survival of raphe nucleus neurons and axonal growth by promoting the development of neurons [91]. In addition, BDNF is involved in the regulation of synaptic plasticity, which affects emotional and memory functions by enhancing synaptic connectivity between the hippocampus and prefrontal cortex and improving the efficiency of serotonergic signaling [47]. Therefore, there is a close interaction between 5-HT and BDNF in the regulation of serotonergic system function, and an abnormality of 5-HT or BDNF may lead to male sexual dysfunction, mood disorders, cognitive deficits, and other pathological problems. Maillet et al. [92] showed that the degree of depression and anxiety was mainly related to the degree of serotonergic disruption in the bilateral subgenual portion or the right dorsal aspect of the anterior cingulate cortex. Huang et al. [93] found that aerobic exercise can significantly increase the expression of 5-HT and BDNF and prolong IELT by ameliorating abnormalities of the serotonergic system, thus improving the symptoms of PE in rats, which was used as an auxiliary treatment for PE along with the SSRIs dapoxetine. Liu et al. [94] confirmed through experiments that high-frequency repetitive transcranial magnetic stimulation (rTMS) could regulate the abnormal regulatory effect of 5-HT and BDNF on the serotonergic system in rats, thereby significantly prolonging IELT and reducing the symptoms of PE in rats. da Fonseca et al. [95] found that probiotics significantly increased the expression of serotonergic-system proteins in the hippocampus and prefrontal cortex of food-deprived rats by correcting abnormal serotonergic regulation—through restoring 5-HT and BDNF signaling—and finally improved the anxiety- and depression-related behavior of those rats. Diniz et al. [96] found that BDNF overexpression in the ventral hippocampus could significantly reduce depression and anxiety behaviors in serotonin-transporter-deficient rats, which revealed the potential of BDNF in the treatment of depression and anxiety in the regulation of serotonergic system abnormalities.
The dopaminergic system is a neurotransmitter system that regulates motor function, emotion regulation, and stress responses. With DA as the core, it is widely involved in the regulation of movement, emotion, hormone secretion, and other important physiological and psychological activities in the CNS [97]. DA mainly acts through four pathways, among which the mesocortical channels are involved in regulating cognitive decision-making, the mesolimbic channels drive motivational behavior, the nigrostriatal pathway controls motor coordination, and the nodular-infundibular pathway regulates endocrine homeostasis [98]. DA acts by binding to receptors on the cell membrane. DA exerts a variety of physiological effects by activating D1-like (DRD1, DRD5) and D2-like (DRD2, DRD3, DAD4) receptor families [99]. Among them, D1-like receptors enhance neuronal excitability through the cAMP-PKA signaling pathway, whereas D2-like receptors inhibit this pathway by coupling Gi/o proteins [100, 101]. 5-HT exerts bidirectional influence on the dopaminergic system via distinct receptor-mediated mechanisms. For example, 5-HT1B receptors, located at the axon terminals of striatal dopaminergic neurons, suppress dopamine release and thereby attenuate dopaminergic signaling efficiency [102]. In contrast, 5-HT2A and 5-HT3 receptors indirectly promote the release of dopamine by activating striatal gamma-aminobutyric acid (GABA) ergic interneurons [103]. The development, function, and synaptic remodeling of dopaminergic circuits are strongly influenced by BDNF-mediated signaling mechanisms. BDNF promotes the survival of dopaminergic neurons and axonal outgrowth by activating the TrkB receptor [31, 104]. BDNF regulates the synaptic plasticity of dopaminergic neurons by enhancing synaptic connectivity and N-methyl-D-aspartate (NMDA) receptor function [47, 105]. Gao et al. [106] demonstrated that the SSRI dapoxetine upregulates the expression of DRD4 to delay ejaculation by increasing the level of 5-HT and mediating the synergistic effect of histone serotoninization, and myeloid zinc finger protein 1, on the promoter region of the DRD4 receptor. By establishing a gradient-overpressure-explosion-trauma model, Ma et al. [107] showed that high-pressure shock waves could induce sequential fluctuations in serum 5-HT, BDNF, and DA levels in rats (the levels of 5-HT and DA decreased in the acute phase, and the level of BDNF decreased continuously in the chronic phase), which were significantly correlated with the bipolar recurrence of anxiety-like behavior. Chen et al. [108] demonstrated that saffron essential oil significantly alleviated depressive symptoms and partially reduced hippocampal neuronal damage by activating the MAPK-CREB1-BDNF signaling axis and upregulating serum 5-HT, DA, and BDNF levels, in mice that had been exposed to chronic, unpredictable, mild stress.
Inflammation is a complex and highly coordinated defense response of the body to
tissue injury, infection, or harmful stimuli, in order to clear pathogens, repair
damaged tissues, and restore body homeostasis [109]. In the male reproductive
system, inflammatory activity contributes to homeostatic regulation by
eliminating pathogens and promoting tissue repair via cytokine-mediated signaling
under normal physiological conditions [110]. Under pathological conditions,
chronic inflammation can trigger oxidative stress and apoptosis, leading to
reproductive-system dysfunction [111]. In the male reproductive system, glandular
tissue such as that in the prostate and seminal vesicle is particularly sensitive
to inflammation; inflammatory diseases such as chronic prostatitis are closely
related to male conditions such as PE and ED [112, 113]. As an important
neurotransmitter in men, 5-HT can be involved in the regulation of male
inflammatory response. In the CNS, 5-HT inhibits the hyperactivation of the
hypothalamic-pituitary-adrenal (HPA) axis by activating the 5-HT1A receptor,
thereby reducing the release of pro-inflammatory factors IL-6 and TNF-
There are interactions among 5-HT, BDNF, and the inflammatory response. On the
one hand, inflammatory microenvironment can lead to the damage of central 5-HT
neurons and the decrease of BDNF expression [68]. On the other hand, decreased
5-HT levels aggravate inflammation and decrease the availability of BDNF [120].
Zhang et al. [121] showed that experimental autoimmune prostatitis
induces disorders of 5-HT metabolism in the CNS by increasing serum levels of
proinflammatory cytokines IL-1
Oxidative stress is a pathological state caused by excessive production of oxides or imbalance of the antioxidant defense system. Its core mechanism is lipid, protein, and DNA damage caused by excessive accumulation of ROS [124]. In the male reproductive system, oxidative stress is an important cause of male diseases such as decreased sperm quality and chronic prostatitis [125, 126]. Testicular tissue is particularly sensitive to oxidative damage due to its active metabolism and low levels of antioxidant enzymes. Chronic oxidative stress can damage the function of the hypothalamic-pituitary-testis axis, reduce the synthesis of testosterone, and induce apoptosis of germ cells; this has become the potential pathological basis of male infertility, ED, and PE [127, 128]. 5-HT exerts antioxidant effects in both CNS and peripheral tissue. 5-HT inhibits excessive activation of the HPA axis by activating the 5-HT1A receptor and reducing cortisol release, thereby decreasing ROS generation [129]. In addition, 5-HT enhances glutathione peroxidase (GPx) activity, directly scrubbing free radicals and protecting Leydig cells from oxidative damage [130].
BDNF ameliorates oxidative stress by regulating neuroplasticity and mitochondrial function. The binding of BDNF to the TrkB receptor can activate PI3K/Akt and MAPK/ERK pathways, up-regulate the expression of SOD and CAT, and enhance the antioxidant-defense ability of cells [131, 132]. In the male reproductive system, BDNF can maintain the synaptic function between hippocampus and prefrontal cortex and alleviate oxidative stress induced by anxiety or depression [133]. It also directly acts on the Sertoli cells of the testis to promote the synthesis of antioxidant proteins [119]. However, the decreased BDNF level caused by chronic stress may impair the ability of the reproductive system to remove ROS, resulting in inflammation and neuronal apoptosis [134]. Jiang et al. [135] found that plasma luteinizing hormone levels were significantly increased in male patients with hypersexual disorder, suggesting dysfunction of the hypothalamic-pituitary-gonadal axis, and suggesting that 5-HT and BDNF may participate in abnormal regulation of sexual behavior including PE through oxidative-stress-mediated neuroendocrine disorders. Xia et al. [136] showed that sesamol significantly restored hippocampal 5-HT and NE levels and alleviated depression and anxiety-like behaviors by activating the Nrf2 antioxidant pathway and up-regulating the BDNF/TrkB/CREB signaling axis. The results suggested that mood disorders associated with gut-brain interaction may be alleviated through the combined actions of 5-HT and BDNF in regulating oxidative stress and neuroinflammatory pathways. Keshavarzi et al. [137] established a methamphetamine-induced neurotoxicity model in rats and confirmed that metformin significantly improved oxidative stress and neuroinflammation by activating the CREB/BDNF and Akt/GSK3 signaling axes, and reversed methamphetamine-induced anxiety, depression, and cognitive impairment. Those findings indicate that neurotransmitter metabolism and mood regulation may be modulated through BDNF-associated signaling pathways.
Emerging evidence has suggested that 5-HT and BDNF exert their physiological and pathological effects through complex intracellular signaling cascades that govern neurotransmission, synaptic plasticity, inflammation, and oxidative stress. Table 1 (Ref. [138, 139, 140, 141, 142, 143, 144, 145, 146, 147]) summarizes key studies that elucidate the mechanisms by which 5-HT-related pathways contribute to PE with comorbid anxiety or depression. Pharmacological and non-pharmacological interventions, including SSRIs (e.g., dapoxetine, fluoxetine), 5-HT receptor agonists/antagonists, acupuncture, and pulsed radiofrequency, have been shown to modulate serotonergic signaling via diverse pathways such as TPH2-mediated 5-HT synthesis, 5-HT1A/1B/2A/2BR/6 receptor activity, SERT modulation, and downstream cascades involving ERK1/2, CREB, and BDNF expression. Those findings have demonstrated the ability of serotonergic interventions to regulate both ejaculatory control and emotional behaviors through central and peripheral mechanisms. Table 2 (Ref. [68, 94, 126, 148, 149, 150, 151, 152, 153, 154]) focuses on BDNF-mediated signaling pathways involved in the neurobiological regulation of PE accompanied by anxiety or depression. Various treatment modalities—including rTMS, herbal compounds, antidepressants, acupuncture, and neurostimulation—activate canonical BDNF signaling via TrkB, CREB, ERK1/2, PI3K/Akt, mTOR, and Nrf2 axes. These pathways contribute to the restoration of hippocampal and prefrontal synaptic plasticity, dendritic integrity, neurogenesis, and antioxidative defense, thereby alleviating ejaculatory dysfunction and affective symptoms. Additionally, synergistic regulation of 5-HT and BDNF—such as BDNF-enhanced TPH2 expression—further underscores their coordinated role in modulating serotonergic tone.
| Signaling pathways/mechanisms | Study drugs/treatments | Subjects | Indicators of observation | Ref. |
| Trp |
Dapoxetine (SSRI) | LPE patients and healthy controls | Trp/LNAAs ratio |
[138] |
| Tph2-mediated signaling pathways for 5-HT synthesis | Dapoxetine (SSRI) | Rats | TPH2-mediated 5-HT synthesis pathway |
[139] |
| The excitability of 5-HT1B receptor was enhanced | Acupuncture needle | PE patients and controls | 5-HT1B receptor excitability |
[140] |
| Inhibition mediated by peripheral 5-HT1A receptors | Selective 5-HT receptor agonists | Wistar rats | Peripheral 5-HT1A receptor activation |
[141] |
| Enhanced 5-HT1A receptor-mediated neurotransmission | Imipramine (tricyclic), fluoxetine (SSRI) | Wistar rats | 5-HT1A receptor-mediated neurotransmission in dPAG |
[142] |
| ERK1/2 phosphorylation mediated by 5-HT2A receptor hyperactivity was enhanced | Fluoxetine (SSRI), doxorubicin combined with cyclophosphamide | Rats | 5-HT2A receptor activation |
[143] |
| 5-HT2BR/ |
Fluoxetine (SSRI) | CMS model mice | 5-HT2BR/ |
[144] |
| 5-HT/SERT | Pulsed radio frequency (PRF) | Wistar rats | SERT density in descending 5-HTergic pathway |
[145] |
| 5-HT6/AC-cAMP-PKA-MEK-CREB-BDNF | EMD-386088; SB-258585; SQ-22536; H89; U0126 | Rats | VLO 5-HT6 receptor activation |
[146] |
| 5-HT3R/NK-1/NK-2 | Plant alkaloids (Berberine) | Patients with neuropathic pain | 5-HT3 receptor activity |
[147] |
LPE, lifelong premature ejaculation; IELT, intravaginal ejaculatory
latency time; dPAG, dorsal periaqueductal gray; CMS, chronic mild stress; SERT,
5-hydroxytryptamine transporter; AC, Adenylyl cyclase; cAMP, Cyclic adenosine monophosphate; PKA, Protein kinase A; MEK, Mitogen-activated protein kinase; VLO, ventrolateral orbital cortex; NK, neurokinin;
| Signaling pathways/mechanisms | Study drugs/treatments | Subjects | Indicators of observation | Ref. |
| BDNF/TrkB | High frequency rTMS (10 Hz) | PE rats and sham stimulation control group | Hippocampal 5-HT concentration |
[94] |
| BDNF/TPH2/5-HT | Observation of natural behavior | Male rats | Dorsal raphe BDNF level |
[68] |
| BDNF/TrkB/CREB | 2-phenyethylamine (PEA) combined with/against CORT | Cort-induced hippocampal neurons/depression model mice | Dendritic spine damage |
[148] |
| BDNF/MAPK | Bladder Meridian Acupressure (ABM) | Chronic stress-induced anxiety in rats | MAPK signaling in hippocampus |
[149] |
| BDNF/ERK1/2/CREB | Spinosin | C57BL/6J male mice | ERK1/2/CREB/BDNF signaling |
[150] |
| BDNF-TrkB-Akt | Imipramine | Female SERT KO rats | BDNF–TrkB–Akt signaling in subcortical limbic cortex |
[151] |
| BDNF/TrkB-ERK1/2 | Bilateral vmPFC DBS | Male CUS model rats and non-stressed rats | BDNF/TrkB signaling |
[152] |
| BDNF/AKT/mTOR | Esketamine (ESK) | Maternal rats with postpartum depression | BDNF/AKT/mTOR pathway integrity in prefrontal cortex |
[126] |
| eIF4E/BDNF | Ketamine | Cort-induced depression in male mice | eIF4E phosphorylation |
[153] |
| Nrf2-BDNF | Chlorphenamine (CPA) | Male Wistar rats | Nrf2–BDNF signaling in hippocampus |
[154] |
SYN, synapsin; PSD95,
postsynaptic density protein 95; TPH2, tryptophan hydroxylase 2; PEA,
2-phenyethylamine; CORT, corticosterone; ABM, bladder Meridian Acupressure; CREB, cAMP response element-binding protein; SERT KO, serotonin transporter
knockout; vmPFC DBS, ventromedial prefrontal cortex deep brain stimulation; CUS,
chronic unpredictable stress; ESK, esketamine; mTOR, mammalian target of
rapamycin; eIF4E, eukaryotic translation initiation factor 4E; MNK1, MAP
kinase-interacting serine/threonine-protein kinase 1; 7,8-DHF,
7,8-dihydroxyflavone; Nrf2, nuclear factor erythroid 2-related factor 2; CPA,
chlorphenamine; MDA, malondialdehyde;
Although no standardized therapy directly targeting 5-HT and BDNF has been established for PE with comorbid anxiety or depression, accumulating evidence has indicated that several existing pharmacological and non-pharmacological approaches may exert therapeutic effects by modulating these pathways. The following summarizes representative treatments, their mechanisms, and related experimental or clinical findings (Fig. 3).
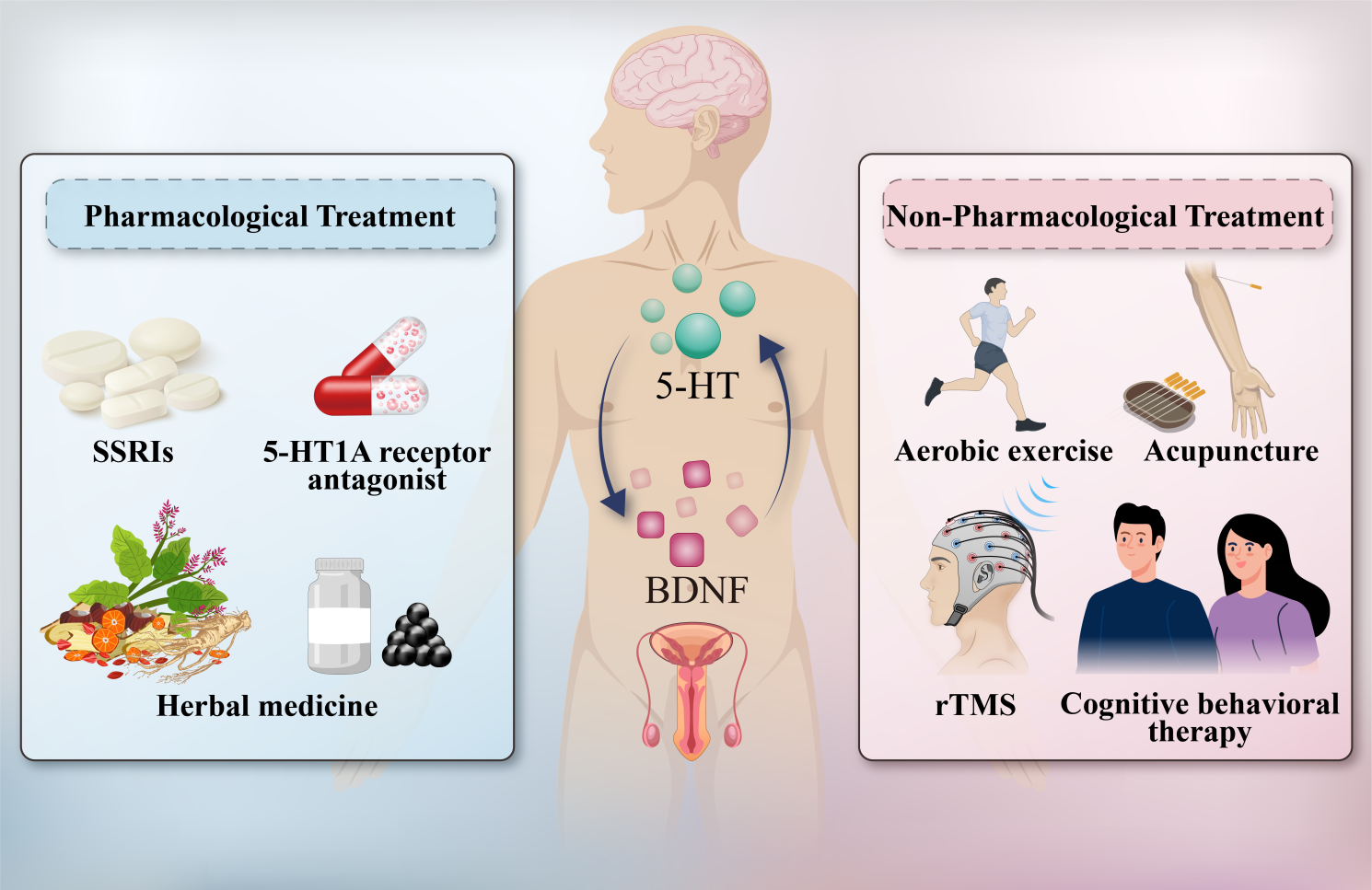 Fig. 3.
Fig. 3.
Clinical application of 5-HT and BDNF in the treatment of PE with anxiety or depression. SSRIs, selective serotonin reuptake inhibitors; rTMS, repetitive transcranial magnetic stimulation. The figure was drawn using Adobe Illustrator.
In a randomized clinical trial, Siroosbakht et al. [155] compared four SSRIs (sertraline, fluoxetine, paroxetine, and citalopram) for the treatment of PE and found that each significantly prolonged IELT via inhibition of 5-HT reuptake. Although adverse effects such as somnolence and dyspepsia were observed, no serious complications occurred and no treatment discontinuations were reported; nonetheless, interindividual differences in tolerability warrant clinical attention. Animal experiments showed that the SSRI fluoxetine reduced anxiety and depression behavior in mice by activating the 5-HT1A receptor (5-HT1AR) to disrupt the nNOS-CAPON interaction, up-regulating the BDNF/ERK/CREB signaling pathway in the hippocampus, and enhancing neural plasticity [156]. Migliorini et al. [157] demonstrated in a double-blind controlled trial that GSK958108, a 5-HT1A receptor antagonist, significantly prolonged IELT in PE patients by 16% in the 3-mg group and 77% in the 7-mg group by blocking the central 5-HT1A autoreceptor and enhancing 5-HT neurotransmission. A study on rats demonstrated that combining the SSRI paroxetine with the 5-HT1A receptor antagonist Atlas987 synergistically modulated the 5-HT system to acutely, on-demand, prolong IELT, suggesting a novel, potentially on-demand strategy for treating LPE [158]. Although SSRIs—by elevating 5-HT concentrations in the synaptic cleft—are first-line agents for PE, recent studies underscored multiple clinical risks, including off-label use, adverse effects, long-term safety concerns, and contraindications in special populations [159]. In addition to their ejaculatory-delay efficacy, SSRIs may induce multiple adverse events during treatment, chiefly manifesting as neurologic symptoms (e.g., dizziness, headache), gastrointestinal disturbances (e.g., nausea, diarrhea), and sexual adverse effects such as decreased libido and secondary sexual dysfunction [160, 161]. Although SSRIs can modulate the 5-HT system to improve both mood and ejaculatory function, clinicians should be aware that excessive serotonergic activity may exacerbate anxiety; therapy should be initiated at low doses with close monitoring of mood changes [162]. Moreover, studies have indicated a strong association between SSRIs and severe hyponatremia, with the highest risk occurring within the first two weeks after initiation; the risk also increases with age, warranting particular vigilance for early-treatment hyponatremia in older patients [163].
In recent years, Traditional Chinese Medicine (TCM)-based therapies have played
a significant and increasingly recognized role in managing PE with comorbid
anxiety or depression [164, 165]. Xie et al. [166] reported that
commonly used TCM monomers—such as Cistanche (Roucongrong), Epimedium
(Yinyanghuo), and Lycium barbarum L. fruits (Gouqizi)—can enhance sexual
function, partly by increasing testosterone and exerting antioxidant effects,
thereby indirectly alleviating anxiety and depression; meanwhile, multi-component
formulas including Shugan Yiyang Capsules, Bazhen Decoction, and Chaihu Shugan
San have been shown to improve PE symptoms and accompanying anxiety/depressive
states via multi-target actions that modulate the NOS–cGMP pathway and
neurotransmitter balance. An animal study demonstrated that a five-herb TCM
formula—Lycium barbarum L. fruits (Gouqizi), Epimedium koreanum Nakai leaves
(Yinyanghuo), Morinda officinalis How roots (Bajitian), Cinnamomum cassia bark
(Rougui), and Eugenia caryophyllata flower buds (Dingxiang)—significantly
improved IELT and ejaculatory control in male rats with sexual dysfunction by
activating the dopaminergic system and increasing serum testosterone levels
[167]. Han et al. [168], in a PE rat model, demonstrated that the TCM
formula Shugan Yidan Fang improved ejaculatory behavior, with its therapeutic
effects likely mediated by modulation of the central dopaminergic system. Cui
et al. [169], in a randomized controlled trial, reported that an
eight-herb topical spray—comprising, among others, Radix Aconiti (Chuanwu),
Herba Asari (Xixin), and Pericarpium Zanthoxyli (Huajiao)—combined with PE
desensitization therapy, significantly prolonged IELT and improved sexual
function indices, and outperformed monotherapy, in patients with lifelong PE.
Feng et al. [170] demonstrated that Xiao-Chai-Hu-Tang can promote nerve
regeneration, regulate neurotransmitter metabolism, and inhibit neuroinflammation
by activating the BDNF/TrkB/CREB and PI3K/AKT signaling pathways, and reduce
depression and anxiety-like behaviors in rats. A study using a chronic,
stress-induced-depression rat model showed that the TCM formula Xiaoyao San
significantly improved depressive-like behavior and locomotor activity, exerting
multi-pathway antidepressant effects by modulating hepatic mitochondrial
metabolomics, energy metabolism, and inflammation-related targets such as IL-6
and TNF-
Huang et al. [93] showed that aerobic exercise enhanced ejaculatory control and prolonged ejaculatory latency in rats with PE by up-regulation of BDNF and 5-HT in the raphe nucleus, and the effect of aerobic exercise on delayed ejaculation was almost equivalent to that of dapoxetine. Seo et al. [172] showed that acupuncture at certain acupoints could alleviate anxiety and stress responses in rats by activating the BDNF-TrkB signaling pathway in the amygdala, and reducing corticotropin-releasing hormone (CRH) expression and inhibiting HPA-axis over-activation, suggesting that acupuncture that targets BDNF may provide a new avenue for the treatment of anxiety and depression by improving neuroplasticity and the stress-emotion pathway. A randomized controlled trial demonstrated that both acupuncture and the SSRI paroxetine significantly prolonged IELT in patients with PE, with each factor performing significantly better than placebo; although paroxetine was superior to acupuncture in extending IELT, acupuncture still exhibited clear therapeutic potential, suggesting benefits mediated via modulation of the serotonergic system [173]. Liu et al. [94] demonstrated that rTMS enhanced the 5-HT level in the hippocampus and up-regulated BDNF, SYN, and PSD95 expression by activating the BDNF-TrkB-CREB pathway, thereby regulating neuroplasticity and significantly prolonging IELT in PE rats. Schosser et al. [174] demonstrated that cognitive behavioral therapy (CBT) regulates neuroplasticity through the BDNF gene polymorphisms; depression patients with specific BDNF variants have more significant responses to CBT treatment, suggesting that CBT may improve depressive symptoms by activating BDNF-dependent synaptic remodeling and hippocampal neurogenesis.
This article reviewed the interrelationship between 5-HT and BDNF in the context of PE comorbid with anxiety or depression. The pathogenesis of PE involves disrupted central 5-HT signaling and impaired BDNF-mediated neuroplasticity, which compromise neuronal survival and synaptic integrity, thereby contributing to the emergence of negative emotional states. Both 5-HT and BDNF appear to be key molecular mediators in the neurobiological mechanisms linking PE with mood disorders.
In both the male reproductive and nervous systems, 5-HT and BDNF play essential roles. In the male reproductive system, 5-HT modulates the ejaculatory threshold via 5-HT1B receptors in the spinal ejaculatory center and promotes sex hormone synthesis, and BDNF enhances Leydig cell proliferation and sperm motility through the PI3K/Akt pathway. In the nervous system, 5-HT inhibits anxiety by regulating the HPA axis and GABAergic transmission in the amygdala via 5-HT1A receptors, whereas BDNF improves cognitive and emotional function by promoting hippocampal neurogenesis and prefrontal synaptic plasticity.
Mechanistically, their synergistic interaction is reflected in four major ways.
First, 5-HT synthesis is reduced due to TPH2 inhibition in the raphe nucleus,
thereby weakening inhibitory input to the spinal ejaculatory center, and BDNF
deficiency disrupts emotional regulation in the limbic system by impairing
TrkB-mediated ERK/CREB phosphorylation. Second, dopaminergic
dysregulation—characterized by D2 receptor overactivation—accelerates
ejaculation, whereas 5-HT2A antagonism and BDNF/TrkB signaling contribute to
DA/5-HT balance. Third, inflammatory cytokines such as IL-6 and TNF-
With respect to clinical applications, management of PE with comorbid anxiety or depression is currently dominated by SSRIs, and no disease-specific targeted pharmacotherapies have yet been developed. Non-pharmacological interventions—including aerobic exercise, acupuncture, repetitive transcranial magnetic stimulation (rTMS), and CBT—have also been reported in clinical studies of PE with anxiety/depression. From the perspectives of physiological and pathological relevance and current therapeutic practice, 5-HT and BDNF emerge as potential biomarkers—and mechanistic nodes—for this condition, acting through multiple pathways in vivo. TCM shows distinctive value: multi-herb formulas such as Shugan Yidan Fang and Xiao-Chai-Hu-Tang may concurrently improve ejaculatory control and mood by rebalancing the 5-HT and dopaminergic systems and activating the BDNF/TrkB signaling pathway; moreover, an herbal spray combined with desensitization therapy can further enhance outcomes in LPE, underscoring the holistic advantage of TCM based on syndrome differentiation (bianzheng lunzhi). Additionally, BDNF- and 5-HT-oriented gene therapies, exogenous BDNF protein administration, BDNF enhancers, and TCM interventions show preliminary promise; however, their precise therapeutic windows, target engagement, and mechanisms of action remain to be fully elucidated.
It is important to note that the central pleiotropy of the 5-HT and BDNF systems makes therapeutic applications unavoidably accompanied by adverse-effect risks. 5-HT is broadly involved in regulating sleep, appetite, and cognition. Although SSRIs improve PE and mood symptoms by inhibiting 5-HT reuptake, they commonly elicit neurologic and gastrointestinal adverse events (e.g., dizziness, nausea), and with long-term use, excessive activation of 5-HT receptors may lead to decreased libido and secondary sexual dysfunction; older patients warrant particular vigilance for hyponatremia. Moreover, supraphysiologic serotonergic tone can destabilize mood regulation, paradoxically exacerbating anxiety and, in severe cases, precipitating serotonin syndrome. Therefore, we propose that special attention should be paid to individual dosage, timely monitoring of mood changes, close attention to drug complications, and careful selection of drug users when using SSRIs for the treatment of PE. In addition, due to the high expression of BDNF in the hippocampus, prefrontal lobe and other key brain regions for emotion and cognition, excessive exogenous supplement of BDNF or excessive activation of TrkB signaling pathway may interfere with normal neuroplasticity, and there is a potential risk of inducing emotional fluctuations or cognitive dysfunction. Fundamentally, these adverse effects reflect the difficulty of restricting current interventions to the pathological targets specific to PE with anxiety/depression. Developing strategies that preferentially engage specific receptor subtypes or modulate signaling within defined brain regions to reduce systemic side effects remains a key challenge for clinical translation.
As PE with anxiety or depression represents a cross-domain disorder involving both neural and male reproductive systems, investigating the roles of 5-HT and BDNF in its pathophysiology remains a central focus of current research. Moving forward, optimizing the application of these molecular targets in clinical treatment—while ensuring both efficacy and safety—will be a key objective.
PE, premature ejaculation; LPE, lifelong premature ejaculation; IELT, intravaginal ejaculatory latency time; APE, acquired premature ejaculation; 5-HT, 5-hydroxytryptamine; CNS, central nervous system; BDNF, brain-derived neurotrophic factor; SSRIs, selective serotonin reuptake inhibitors; 5-HTT, 5-hydroxytryptamine transporter; DA, dopamine; TrkB, tropomycin receptor kinase B; ET-1, endothelin-1; NE, norepinephrine; PI3K, phosphatidylinositol-3-kinase; MAPK, mitogen-activated protein kinase; JNK, c-Jun N-terminal Kinase; NF-
JFQ and ZMH designed the research study and revised the manuscript critically for important intellectual content. YW, BN, and SJW conceptualized the study, performed literature searches, drafted the manuscript, and designed the figures. WSC, LQZ, and CXC collected and sorted references, assisted in data collation, and provided technical support for figure optimization. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This study was supported by the National Natural Science Foundation of China (No. 82305229) and the Shenzhen Science and Technology Program (No. JCYJ20230807094759010).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.