









 , Bin Sun 1,†, Jing Luo 1, Guanhua Hu 1, Nan Shao 1, Wenwen Si 1,*
, Bin Sun 1,†, Jing Luo 1, Guanhua Hu 1, Nan Shao 1, Wenwen Si 1,*
1 School of Integrated Chinese and Western Medicine, Anhui University of Chinese Medicine, 230012 Hefei, Anhui, China
†These authors contributed equally.
Abstract
Acute ischemic stroke (AIS) is one of the leading critical neurological conditions globally, resulting in significant adult mortality and disability. Previous studies have demonstrated a close relationship between AIS and the ferroptosis signaling pathway. Muscone, the primary active small-molecule component of musk, is a traditional Chinese medicine that exhibits significant pharmacological effects in reducing stroke injury. However, there is still only limited research on whether muscone can modulate ferroptosis-related injury in AIS, and on the underlying regulatory molecular mechanisms.
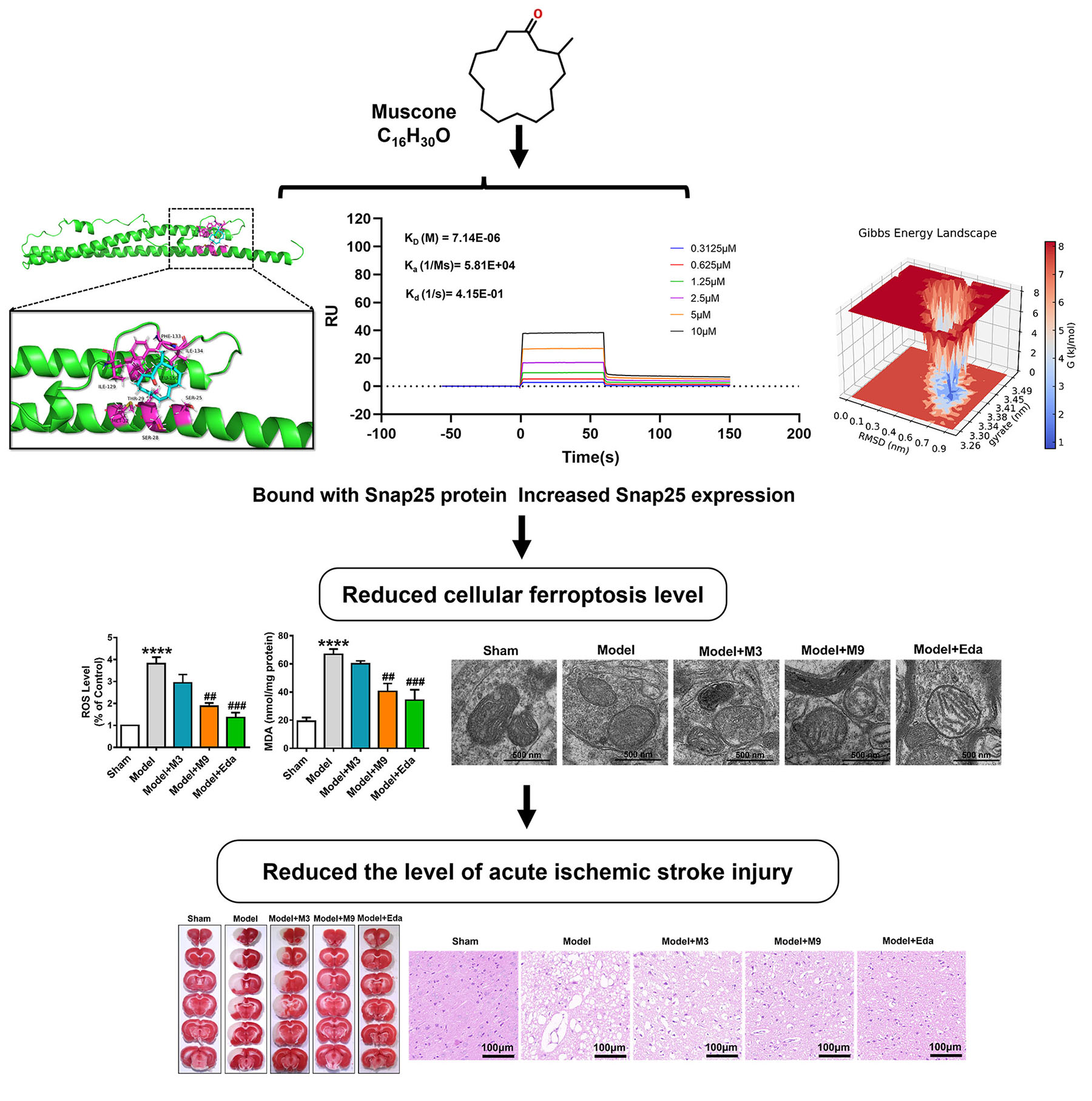
We utilized a transmission electron microscope and concurrently performed assays for glutathione peroxidase 4 (GPX4) activity, glutathione (GSH), reactive oxygen species (ROS), lipid peroxides, as well as cell viability and live/dead cell staining to investigate alterations in ferroptosis levels. RNA sequencing, bioinformatics analysis, and western blot (WB) assays were employed to evaluate the changes in synaptosome-associated protein 25 kDa (Snap25) expression levels. Furthermore, molecular docking, surface plasmon resonance (SPR) detection, and molecular dynamics (MD) simulation were implemented to examine the binding affinity and interaction between muscone and Snap25.
RNA sequencing technology, bioinformatics analysis, and WB assays revealed that Snap25 was specifically downregulated under simulated AIS conditions. Snap25 knockdown and overexpression experiments were also conducted to elucidate the molecular mechanism by which muscone modulates Snap25 expression, thereby mitigating ferroptosis injury in AIS. Additionally, the results of molecular docking, SPR detection, and MD simulations indicate that muscone has multiple binding sites that allow it to bind directly to the Snap25 protein, thereby stabilizing the protein structure.
Our findings suggest that muscone produces an anti-AIS effect in the context of AIS injury by increasing Snap25 protein expression, thus reducing ferroptosis. This investigation offers insight into the anti-stroke mechanism of muscone and introduces a promising new treatment option for clinical AIS management.
Graphical Abstract
Keywords
- muscone
- ischemic stroke
- ferroptosis
- Snap25
- MCAO/R
Stroke is still one of the primary causes of death and disability in the adult population globally [1, 2, 3]. The majority of cases are due to acute ischemic stroke (AIS), distinguished by sudden onset, fast progression, and complex clinical manifestations [4, 5]. AIS results in disrupted blood supply, inadequate nutrition, and the accumulation of toxic substances which leads to severely impaired nerve function [6, 7, 8]. The most frequently used treatment for AIS is revascularization therapy. However, since the time window for revascularization treatment is narrow, a substantial proportion of AIS patients do not derive any benefit [9, 10]. Therefore, developing safer and more effective drugs for treating AIS has major clinical significance.
Ischemic stroke represents a complex and highly heterogeneous condition. The primary subtypes of ischemic stroke include atherosclerotic thrombotic infarction, cardiogenic embolic stroke, and lacunar infarction, each of which is characterized by distinct etiologies and risk factors [11]. Atherosclerotic thrombotic infarction is predominantly associated with large artery atherosclerosis, and shows a higher prevalence in adult males. Patients with this subtype typically experience prolonged hospital stays and face an elevated short-term risk of recurrence. Cardioembolic stroke is primarily attributable to cardiac pathologies, and occurs more frequently in elderly women. Individuals affected by this subtype often present with severe symptoms at onset, encounter numerous complications, and have a poor prognosis [12]. Lacunar infarction is commonly related to small vessel disease and is prevalent among hypertensive patients. This subtype generally manifests with milder symptoms and has a more favorable prognosis [13].
AIS involves various mechanisms of disease progression, including excitotoxicity, oxidative stress, inflammatory response, and ferroptosis [14, 15, 16]. Current research has shown that functional and structural damage to AIS neurons is closely associated with the level of oxidative stress. Specifically, the elevated concentration of reactive oxygen species (ROS) has been identified as the primary factor contributing to the substantial increase in oxidative stress [17, 18]. ROS-mediated lipid peroxide (LPO) damage is a primary mechanism of cellular injury, resulting in altered physiological properties of the cell membrane and increased membrane permeability during AIS, ultimately leading to neuronal cell death [19, 20]. Abnormally elevated levels of ROS and lipid peroxidation are critical indicators of the ferroptosis pathway [21, 22]. Ferroptosis is a type of cell death that relies on iron and is initiated by excessive damage from lipid peroxidation, resulting in elevated oxidative stress and eventual cell death [23, 24, 25]. Recent investigations have underscored the pivotal function of ferroptosis in the pathogenesis of neurological disorders [26, 27]. Nevertheless, the regulatory mechanisms underlying ferroptosis in AIS-induced injury remain incompletely understood.
Musk is a traditional Chinese medicine (TCM) utilized extensively in treating cerebrovascular disorders such as ischemic stroke. Muscone is a crucial constituent of musk and the primary physiologically active substance. It has been demonstrated to decrease cerebral infarct volume and neurological dysfunction [28, 29]. Synaptosome-associated protein 25 (Snap25) is a widely expressed neurotransmitter transport protein in neuronal cells that is pivotal in neurotransmission, primarily facilitating synaptic vesicle fusion and neurotransmitter release [30]. Snap25 has been linked to the development of multiple neurological conditions, such as attention deficit hyperactivity disorder [31], Alzheimer’s disease [32], and developmental and epileptic encephalopathy [33]. Recent study has demonstrated that Snap25 can mitigate postoperative cognitive dysfunction by promoting mitochondrial autophagy and reducing pyroptosis-induced damage in hippocampal neurons [34]. In the present study, RNA sequencing and western blot (WB) experiments revealed that muscone significantly upregulates Snap25 expression under AIS conditions. This prompted us to investigate whether muscone reduces ferroptosis levels in AIS by modulating Snap25, an area that to our knowledge has yet to be explored.
Our study utilized 2,3,5-Triphenyl-Tetrazolium Chloride (TTC) and live/dead cell staining to assess the damage level in a rat model of middle cerebral artery occlusion/reperfusion (MCAO/R), and in oxygen-glucose deprivation/reperfusion (OGD/R) injury of PC12 cells. The ferroptosis level was determined by evaluating the activity of glutathione peroxidase 4 (GPX4) and the concentrations of glutathione (GSH), ROS and LPO, as well as by observing changes in mitochondrial morphology. We employed a combination of experimental approaches, including molecular docking, surface plasmon resonance (SPR) detection, molecular dynamics (MD) simulation, RNA interference (RNAi), and protein overexpression to explore the mechanism by which muscone regulates ferroptosis in AIS. The findings of this study should increase our comprehension of the underlying mechanisms of AIS injury and the therapeutic effects of muscone, thus offering promising therapeutic approaches for AIS clinical management.
Animal trials received approval from the Animal Ethics Committee, Anhui University of Chinese Medicine (Hefei, China; ethical clearance number AHUCM-rats-2023126) and were conducted as per the ARRIVE instructions. Twenty-four male Sprague-Dawley rats (specific pathogen-free, SPF, Anhui Qingyuan Biotechnology Co., Ltd, Hefei, China) with a weight of 230–260 g and aged 8–9 weeks were equally assigned to four groups in a random manner: sham control, M3 (muscone 3 mg/kg), M9 (muscone 9 mg/kg), and M18 (muscone 18 mg/kg). Pharmaceutical-grade muscone (S25595, Yuanye Biotechnology, Shanghai, China) was delivered intravenously via the tail vein. Following a 24-h post-administration period, anesthetization of rats was conducted with 5% isoflurane (R510-22-10, RWD Life Science Co., Ltd, Shenzhen, China) and 2.5% isoflurane was used to maintain them. Terminal blood collection (5 mL) was performed via abdominal aortic puncture prior to euthanasia by cervical dislocation under sustained anesthesia. The blood specimens were spun at 1800 g for 15 min to isolate serum components. Hepatic and renal biomarkers (ALT, alanine aminotransferase; AST, aspartate aminotransferase; BUN, blood urea nitrogen; Cr, creatinine) were quantified using a HITACHI 7600-020 Automatic Analyzer (Hitachi High-Technologies Corporation, Tokyo, Japan) and following standardized protocols.
Sprague-Dawley rats (n = 60) with the age of 8–9 weeks (230–260 g body weight)
were classified into five experimental cohorts in a random manner: sham-operated
controls, MCAO/R model group, MCAO/R with low-dose muscone (3 mg/kg), MCAO/R with
high-dose muscone (9 mg/kg), and MCAO/R with edaravone (3 mg/kg) treatment. The
MCAO/R model was prepared as illustrated in previous work [35, 36]. A median
cervical incision was made to expose the left common (CCA), internal (ICA), and
external carotid arteries (ECA). The CCA was clamped, the distal ECA ligated, and
a micro-incision made at the proximal ECA. A silicone-coated thread embolus
(907-00029-01, RWD Life Science Co., Ltd) was inserted via the
ECA, advanced into the ICA through the CCA bifurcation, and stopped upon
resistance. After securing the embolus, ischemia was sustained for 1 h, followed
by thread withdrawal. The proximal ECA was ligated, the CCA clamp removed, and
reperfusion initiated. A laser Doppler flowmeter (PF5010, Perimed Co., Ltd,
Stockholm, Sweden) was utilized for monitoring the regional cerebral blood flow (rCBF). A
The Longa score scale was utilized to evaluate the neurological deficit in MCAO/R rats as described previously [36]. The neurological deficit evaluation was performed at the 24th hour after blood perfusion was restored in MCAO/R rats.
Following the establishment of the AIS model, MCAO/R rats were euthanized by rapid decapitation, and their whole brains extracted, and then cutting them was conducted into slices of 2 mm thickness on ice. Staining the slices was then conducted (37 ℃, 20 min) with 2% TTC dye solution (BB-44549, Bestbio Co., Ltd, Shanghai, China) and photographed. Infarct areas were identified as pale regions in the cerebral hemisphere, and the percentage of infarct size was calculated using Image J software (v1.52a; NIH, Bethesda, MD, USA).
Total RNA extraction was conducted from both healthy and infarcted cortical
tissues via Trizol reagent (15596026CN, Invitrogen, Carlsbad, CA, USA). Following
library preparation and alignment, genes were classified as differentially
expressed genes (DEGs) if they exhibited a
After adequate fixation, the tissues/cells were stained with HE staining kits (C0105S, Beyotime Biotechnology, Shanghai, China). The degree of damage in the cortex of rats was then assessed via an inverted microscope (IX81, Olympus, Tokyo, Japan).
In ice-cold RIPA buffer (89900, Thermo Fisher Scientific, Waltham, MA, USA), cortical tissue and PC12 cells were lysed. Beyotime Biotechnology’s BCA kit (P0012, Shanghai, China) was used to measure total protein. After being separated using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), protein samples weighing 30 µg were transferred to polyvinylidene fluoride (PVDF) membranes with a thickness of 0.45 µm. Before incubating with primary and secondary antibodies, a 1-h blockage of membranes was conducted with 5% bovine serum albumin at room temperature. Located in Wuhan, China, ProteinTech is the source of the anti-Snap25 antibody (1:5000, 14903-1-AP). Abcam (Cambridge, UK) provided the following antibodies: GAPDH (1:5000, ab181602), horseradish peroxidase (HRP, 1:5000, ab6721), and ab6789, which are goat anti-rabbit and mouse IgG H&L. The enhanced chemiluminescence (ECL) substrate (34580, Pierce, Thermo Fisher Scientific) was used for band visualization. The gray value of the protein band was calculated by using the Image J software (v1.52a; NIH, Bethesda, MD, USA). The original western blots can be found in the Supplementary Material.
The PC12 cell viability was strictly assessed with the CCK-8 assay kit (C0037,
Beyotime Biotechnology) as per the manufacturer’s guidelines.
Subsequently, cell viability was evaluated via an enzymatic reader at 450 nm. The
formula used to determine cell viability was: % cell viability = OD
(treatment)/OD (control)
PC12 cells were brought from the American Type Culture Collection (Catalogue no.
CRL-1721.1TM, ATCC, Manassas, VA, USA) and cultivated in RPMI 1640 medium
(Catalogue no. 11875093, Invitrogen) with 5% fetal bovine
serum (Catalogue no. A5670701, Invitrogen). The cells underwent regular
authentication through morphological examination and short tandem repeat (STR)
authentication and were checked for Mycoplasma contamination. The mycoplasma
contamination detection kit (Beyotime Biotechnology, Cat. No. C0297S) was
used to test cell lines for mycoplasma, and they were found to be negative. PC12
cells were cultivated into 6-well plates (2
ROS concentrations in cortical tissue were assessed via a ROS detection kit
(BB-470515, BestBio Co., Ltd.) as per the guidelines of the
manufacturer. Fluorescence was detected at 510 nm (excitation) and 610 nm
(emission) via a SpectraMax iD3 microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Intracellular ROS in PC12 cells was evaluated with the Cell ROS Assay Kit
(S0033S, Beyotime Biotechnology). Cells (3
The TBARS Assay Kit (S0131M, Beyotime Biotechnology, Shanghai, China) was
utilized to measure lipid peroxidation in cortical tissue and PC12 cells. For
this analysis, cortex tissue samples (25 mg) and cell suspensions (1
The brain cortex tissue of rats, as well as cultured PC12 cells, were stabilized at 4 ℃ for 2 h with 2.5% glutaraldehyde (Catalogue no. G5882, Sigma-Aldrich, St. Louis, MO, USA). Subsequently, the mitochondrial morphology was examined by TEM (JEM-1400 PLUS, Tokyo, Japan).
The LIVE/DEADTM imaging kit from Thermo Fisher Scientific
(L3224, Invitrogen) was used as recommended by the
manufacturer to assess cell mortality across all experimental groups. PC12 cells
were photographed at 100
GSH content and GPX4 activity were measured in homogenized rat cortex and PC12 cell supernatants using the GSH test kit (ab138881, Abcam) and GPX4 Activity Assay Kit (E-BC-K883-M, ElabScience Biotechnology Co., Ltd, Wuhan, China), respectively, as recommended by the guidelines of the manufacturer.
An iron content detection kit (I291, Dojindo Laboratories, Kumamoto, Japan) was used to measure the concentrations of ferrous and total iron ions in rat cortical tissue and PC12 cells, as recommended by the manufacturer. Microplate readers from Thermo Fisher Scientific were employed to calculate absorbance at 593 nm, which was subsequently used to calculate the concentrations of iron ions.
Autodock Vina 1.1.2 software (Scripps Research Institute, San Diego, CA, USA) was employed to conduct molecular docking simulation analysis. The crystal structure of Snap25 protein (Protein Data Bank (PDB) ID: 1N7S) was gained from the RCSB Protein Data Bank (https://www.rcsb.org/, USA) in PDB format. The molecular structure of muscone (ID 10947) was retrieved from the PubChem database (https://pubchem.ncbi.nlm.nih.gov) and transformed into PDB format with the PyMOL Molecular Graphics System (v 2.5.0, Schrödinger, Inc., New York, NY, USA). Finally, Autodock Vina software was employed to perform the docking study.
SPR detection was employed to assess the interaction between Snap25 protein
(HY-P73634, MedChemExpress, Monmouth Junctio, NJ, USA) and muscone.
Immobilization of the Snap25 protein was conducted on a Series S CM5 Sensor Chip
(GE Healthcare, Washington, DC, USA) via amino coupling. Muscone solutions were
prepared at numerous doses (0.3125, 0.625, 1.25, 2.5, 5, and 10 µM) using
1
MD simulations were conducted via GROMACS (v 5.1.4, Uppsala University, Uppsala, Sweden). Two systems were constructed: with ligand (Snap25 protein + muscone), and without ligand (Snap25 protein only). Topological files for the molecular structures were generated via the CHARMM-GUI (http://www.charmm-gui.org) web interface. Both systems were positioned in a cubic simulation box with applying periodic boundary circumstances, using the gmx solvate tool in GROMACS (https://www.gromacs.org, GROMACS 2023.3, KTH Royal Institute of Technology, Stockholm, Sweden). Upon completion of the simulation, various metrics were computed and analyzed.
Synthetic siRNA was used to perform RNAi targeting Snap25. Three Snap25-specific siRNAs (Snap25-siRNA-1, Snap25-siRNA-2, Snap25-siRNA-3) and a negative control (NC) siRNA were synthesized by Hanbio Corporation (JY20220810WY-SI02, Shanghai, China). The siRNAs and NC sequences were as follows: Snap25-siRNA-1, UUGCUCAUCCAACAUAACCTT; Snap25-siRNA-2, UUAUUGAUUUGGUCCAUCCTT; Snap25-siRNA-3, UUCUGCUUCUUUCAUAUCCTT; NC, ACGUGACACGUUCGGAGAATT. Transfection was carried out via Lipofectamine™ 3000 (L3000015, Invitrogen, Carlsbad) with Snap25-siRNA-1, Snap25-siRNA-2, Snap25-siRNA-3, or NC siRNA (100 nM for each). PC12 cells were transfected for 24-h prior to further experiments.
Snap25 overexpression plasmids were prepared by Hanbio Corporation
(JY20220811WY-PC01). The
pcDNA3.1-cytomegalovirus (CMV)-Snap25 plasmids were introduced
into PC12 cells via Lipofectamine™ 3000 (L3000015, Invitrogen). A transfection of 1
SPSS software (v 20.0) (IBM Corp., Armonk, NY, USA) was employed for the statistical
analysis of data. GraphPad Prism (v 8.0, GraphPad Software, Inc., San Diego, CA,
USA) was utilized to create all graphical images. The Kruskal-Wallis
non-parametric test or one-way ANOVA compares two or more groups, and then
post-hoc analysis with Tukey’s multiple comparisons test was conducted. A
p-value of
To select the appropriate dose of muscone for experimentation,
the liver and kidney functions in rats were first evaluated by serum biochemical
indicators. Liver and kidney functions showed no significant damage in the M3 and
M9 groups, but substantial liver and kidney toxicity were found in the M18 group,
as shown in Fig. 1A (ALT: M18 vs. sham, p
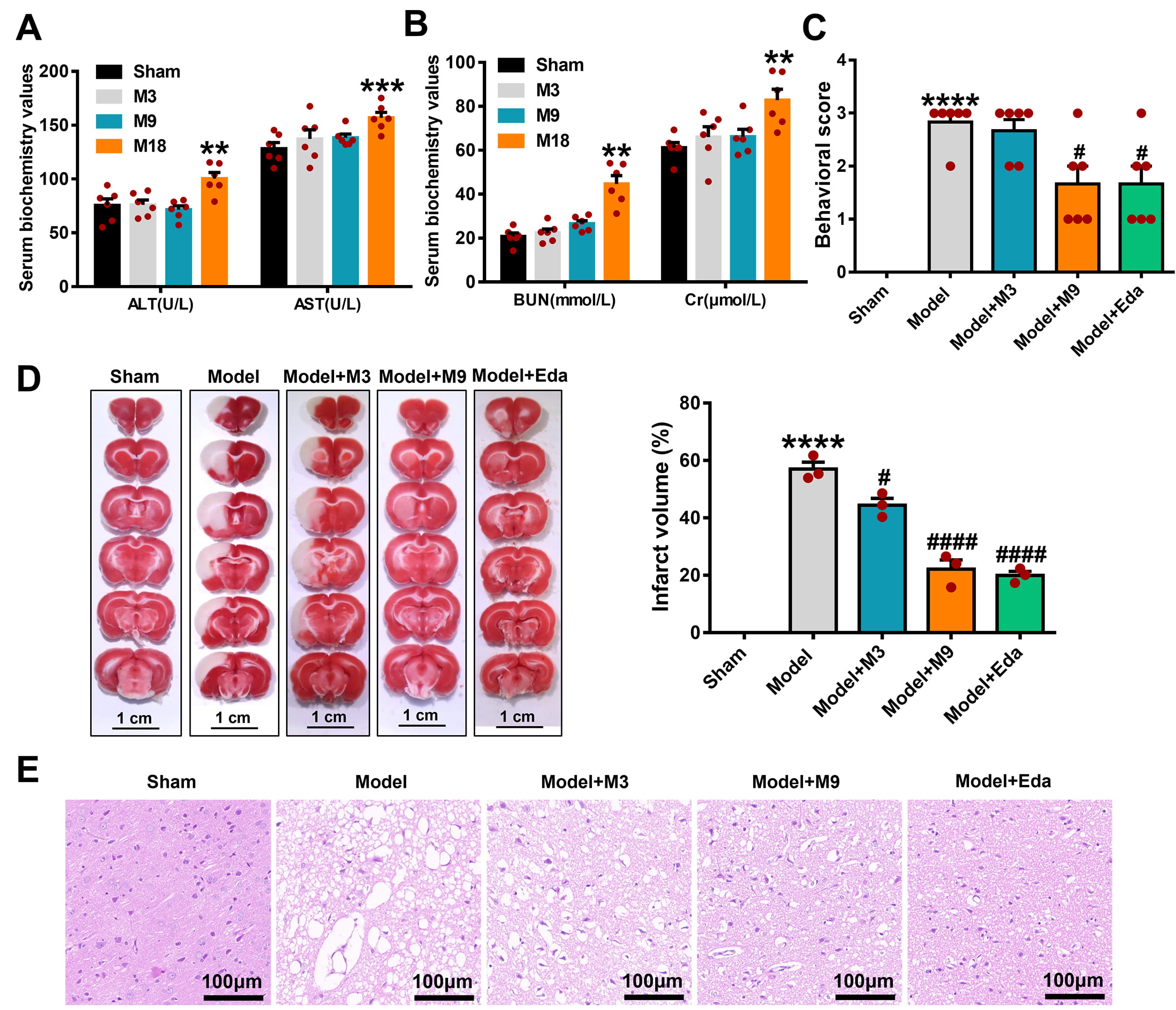 Fig. 1.
Fig. 1.
Muscone mitigates acute cerebral ischemic injury in
middle cerebral artery occlusion/reperfusion (MCAO/R) rats. (A) The impact of
different concentrations of muscone (M3 group, 3 mg/kg; M9 group, 9 mg/kg; M18
group, 18 mg/kg) on liver function in rats, as detected by the serum biochemical
index. (B) The impact of different concentrations of muscone on kidney function
in rats, as detected by the serum biochemical index. (C) Muscone enhanced the
neurological deficit scores in MCAO/R rats. The protective influence of muscone
on the ischemic cortex in MCAO/R rats was observed by 2,3,5-Triphenyl-Tetrazolium
Chloride (TTC) staining (Scale bar = 1 cm) (D) and hematoxylin-eosin (HE) staining (Scale bar = 100 µm) (E). Data are
signified as the mean
TEM illustrated the disrupted mitochondrial cristae and membranes in the
cortical tissue of model rats, which were markedly preserved in the M3, M9, and
Eda groups (Fig. 2A). GPX4 activity (Fig. 2B) and GSH levels (Fig. 2C) were
significantly higher in these groups vs. the model (M3: p
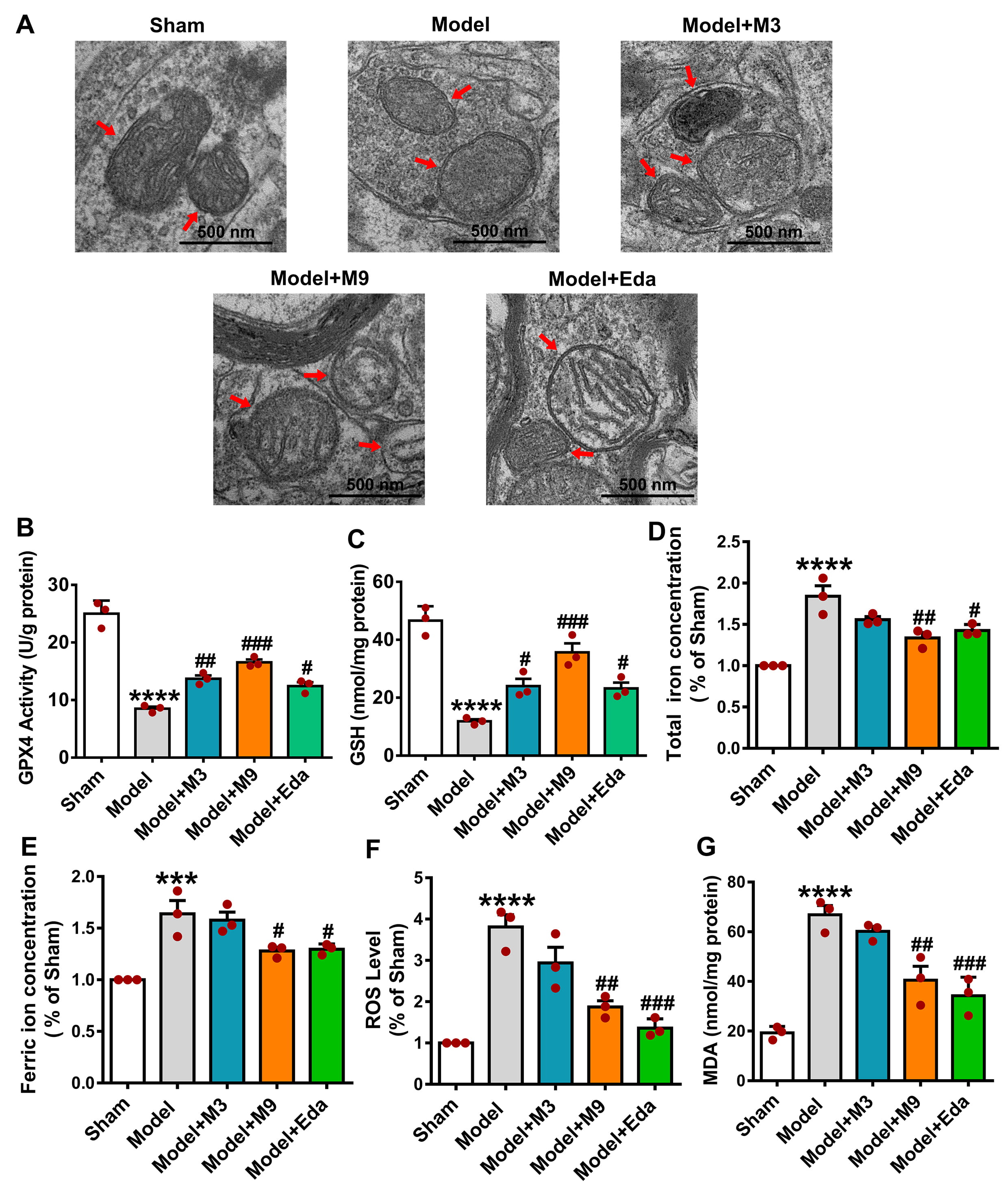 Fig. 2.
Fig. 2.
Muscone attenuates ferroptosis in the ischemic cortex of MCAO/R
rats. (A) Transmission electron microscopy (TEM) shows reduced mitochondrial
damage in the cortex after muscone treatment. The red arrows indicate
mitochondria. Muscone increased glutathione peroxidase 4 (GPX4) activity (B) and
glutathione (GSH) levels (C), and decreased total iron (D), ferric iron (E),
reactive oxygen species (ROS) (F), and malondialdehyde (MDA) concentrations (G)
in MCAO/R rats. Mean
Cortical tissues from the sham and model groups (n = 3 per group) were subjected
to RNA sequencing. DEGs were subsequently identified and analyzed using
bioinformatics approaches. The results of GO analysis revealed that Snap25 was
significantly enriched in multiple signaling pathways associated with BP (Fig. 3A), CC (Fig. 3B), and MF (Fig. 3C). These results suggest that differential
expression of Snap25 may have a crucial function in cortical damage in MCAO/R
rats. Heatmaps (Fig. 3D) show a significant downregulation of Snap25 expression
in the model group compared to the sham group. WB results shown in Fig. 3E
confirm that Snap25 protein expression in the cortex illustrated a significant
decrease in the model group (sham vs. model, p
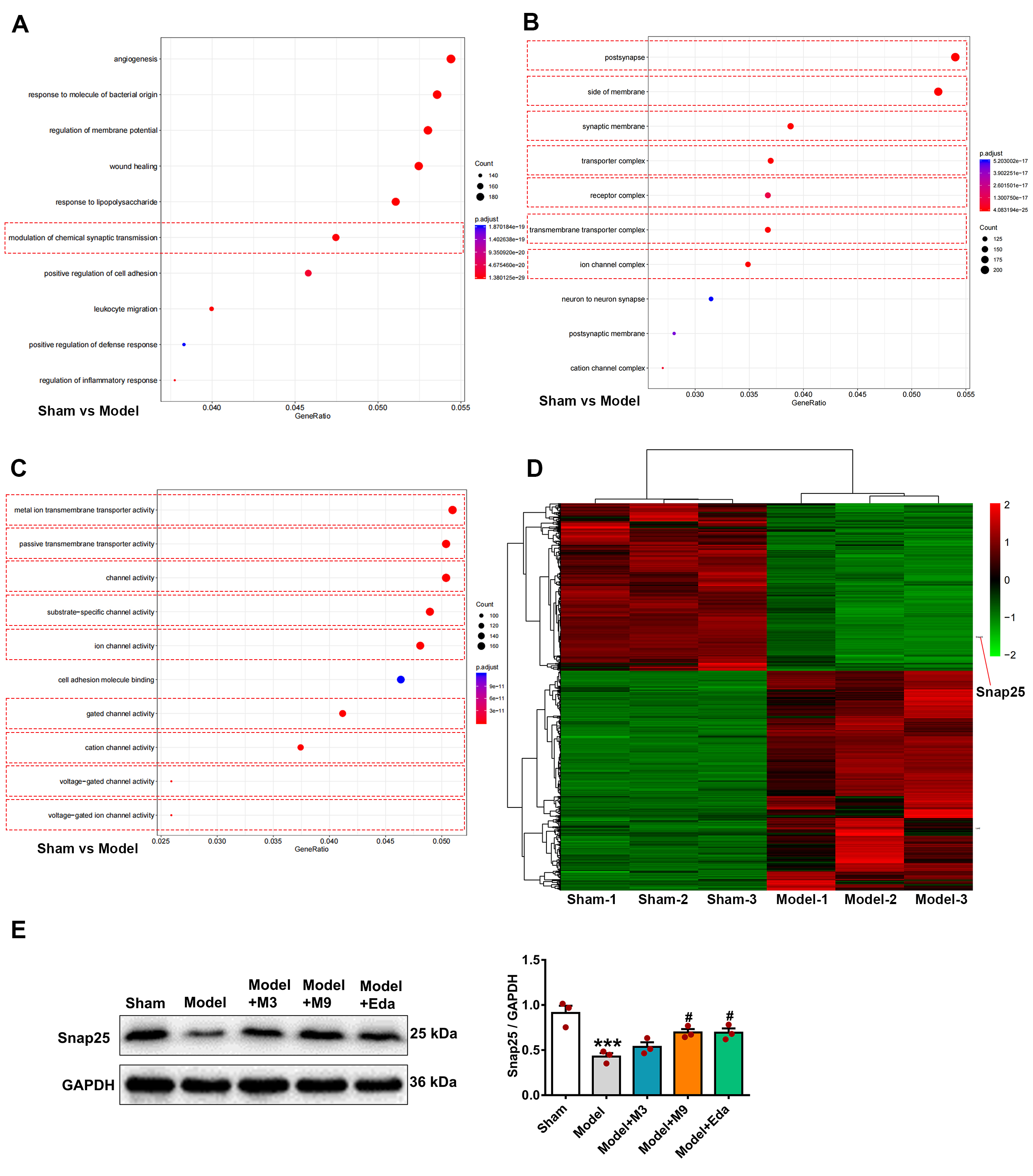 Fig. 3.
Fig. 3.
Decreased Synaptosome-associated protein 25 kDa (Snap25)
expression in the ischemic cortical tissue of MCAO/R rats. Cortical tissues from
the sham and model groups (n = 3 per group) underwent RNA sequencing to identify
differentially expressed genes (DEGs). These were subsequently analyzed using
Gene Ontology (GO) enrichment analysis, with the Snap25 gene found to be
prominently involved in several signaling pathways associated with biological
process (BP)s (A), cellular component (CC)s (B), and molecular function (MF)s
(C). Heatmap visualization (D) revealed a noticeable reduction in Snap25
expression in the model group relative to the sham group. Additionally, western
blotting (WB) analysis (E) revealed that the Snap25 protein level in the rat
cortex showed a significant lessening in the model group compared to the sham
group, but elevated markedly following muscone treatment. One-way ANOVA compared
the means of several groups. Data are shown as the mean
The impact of muscone on PC12 cells exposed to OGD/R was examined using CCK8
assay and live/dead cell staining. CCK8 analysis revealed the M300 group (300
ng/mL muscone) had substantially better cell viability compared to the control
group (Fig. 4A; control vs. M300, p
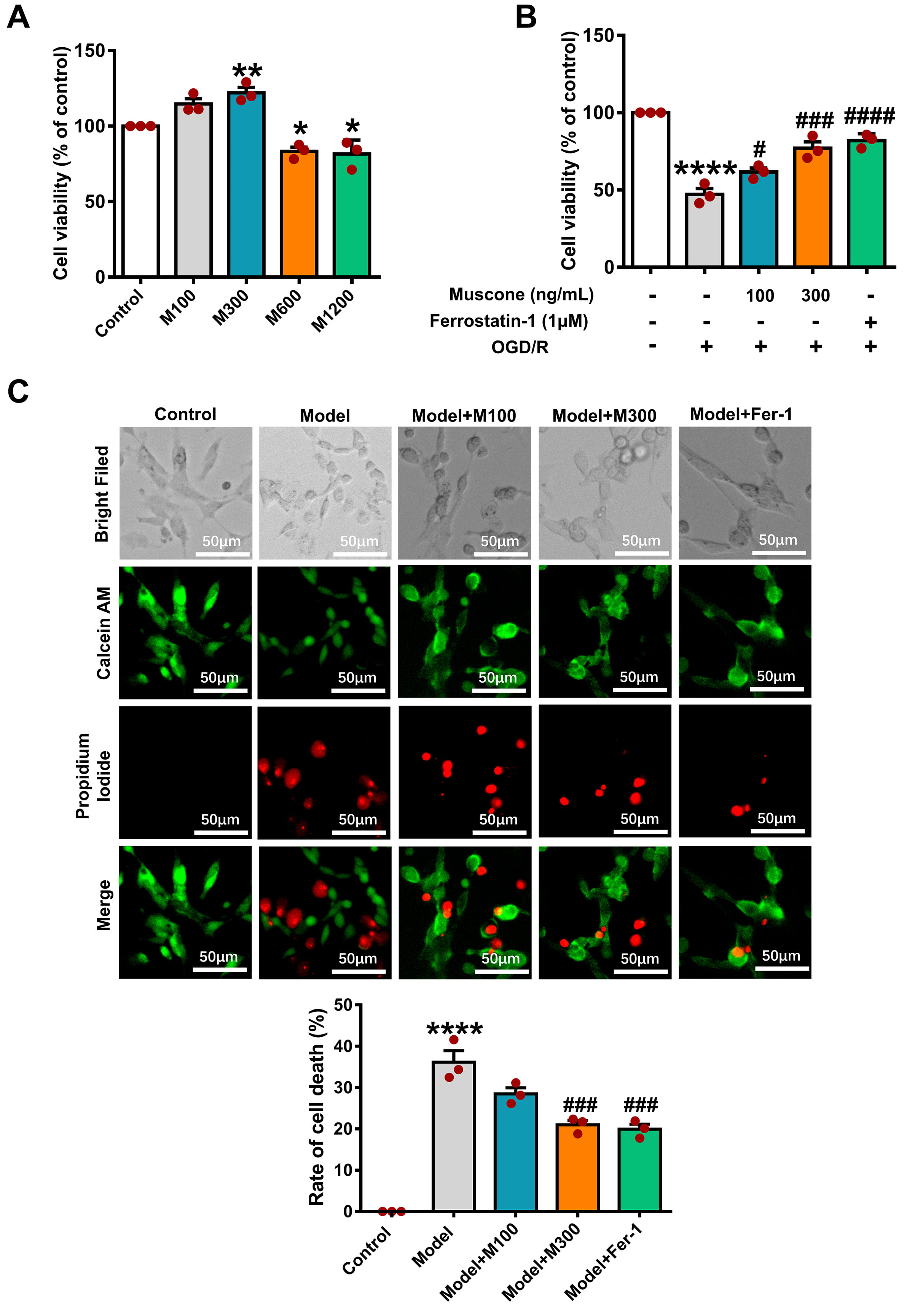 Fig. 4.
Fig. 4.
Muscone reduces mortality in PC12 cells following oxygen-glucose
deprivation/reperfusion (OGD/R) damage. (A) CCK-8 assay shows the effects of
muscone (100–1200 ng/mL) on PC12 cell viability. (B) Muscone increased viability
in OGD/R-injured cells; Ferrostatin-1 served as a positive control. (C) Live/dead
staining indicates muscone (100 and 300 ng/mL) reduced cell mortality after
OGD/R. Data are mean
TEM analysis revealed substantial mitochondrial damage in the model group, such
as decreased cristae and disrupted membranes (Fig. 5A). Damage was significantly
reduced in the M300 and Fer-1 groups. GPX4 activity (Fig. 5B) and GSH levels
(Fig. 5C) were significantly higher in the M100, M300, and Fer-1 groups vs. the
model (M100: p
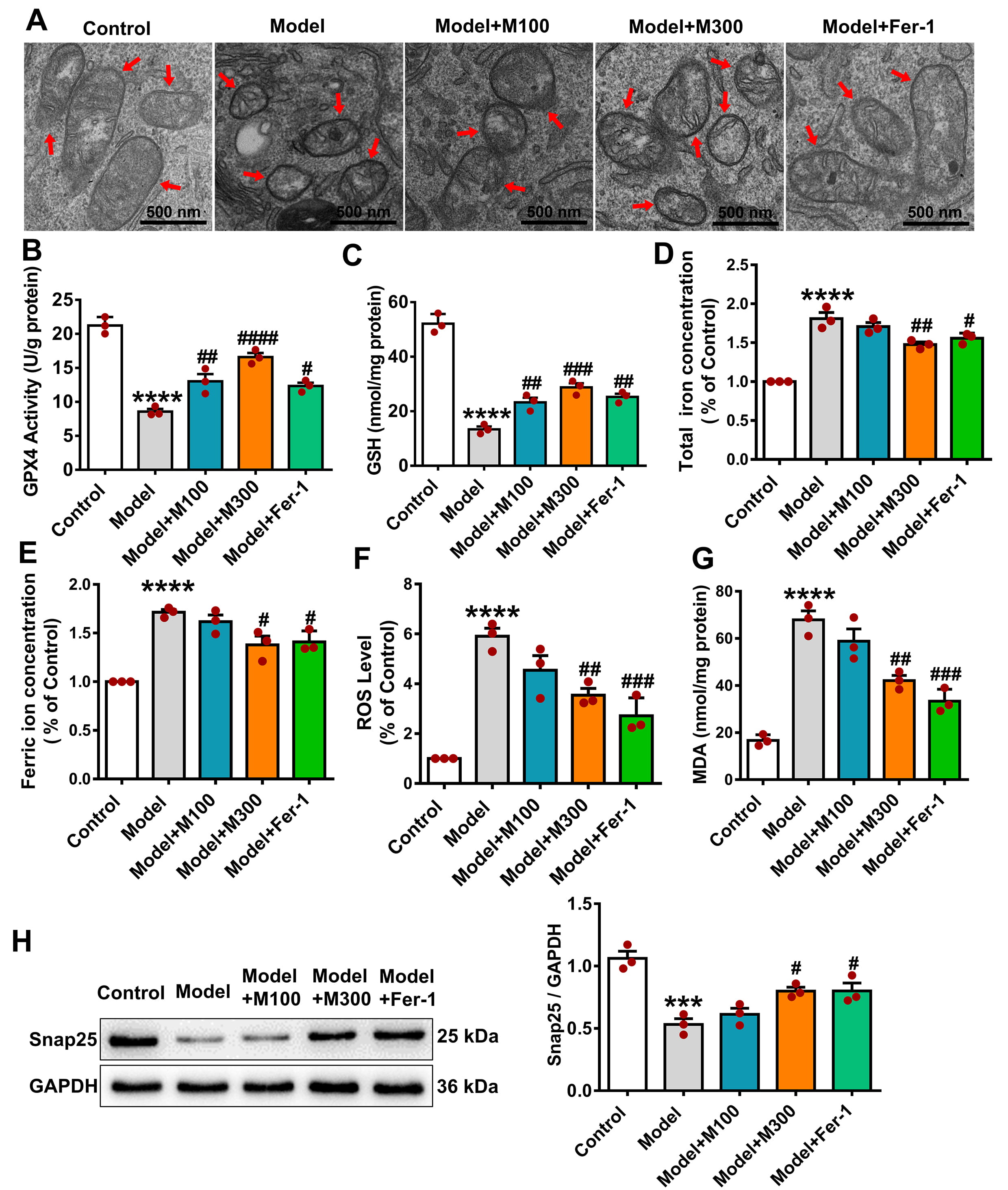 Fig. 5.
Fig. 5.
Muscone decreases ferroptosis in PC12 cells exposed to OGD/R.
(A) Muscone attenuates mitochondrial damage in OGD/R-injured PC12 cells. The red
arrows indicate mitochondria. Scale bar: 500 nm. Muscone increased GPX4 activity
(B) and GSH levels (C), while decreasing total iron (D), ferric iron (E), ROS
(F), and MDA levels (G). (H) Snap25 protein expression, reduced by OGD/R, was
restored by muscone. Data are mean
We next investigated the Snap25 function in the ferroptosis
pathway during OGD/R injury by using Snap25-specific siRNAs and NC siRNA. WB
results established effective silencing of Snap25 in the Snap25-siRNA-1
group compared to the NC group (Fig. 6A; Snap25-siRNA-1 vs. NC, p
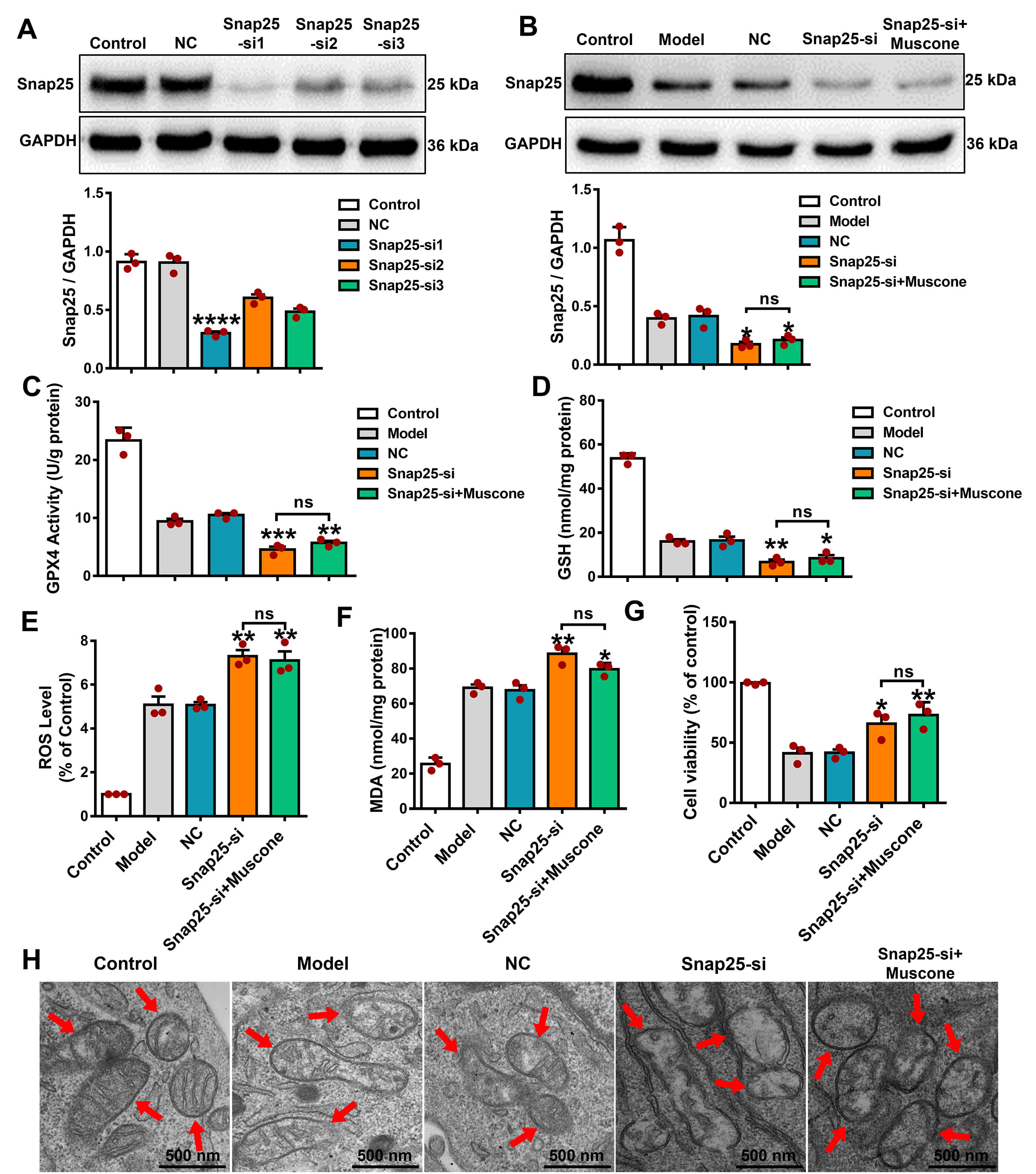 Fig. 6.
Fig. 6.
Snap25 silencing increased the level of ferroptosis and
eliminated the anti-ferroptosis impact of muscone in OGD/R-damaged PC12 cells.
(A) Effective knockdown of Snap25 in PC12 cells. WB analysis established the
effective reduction of Snap25 expression in PC12 cells. (B) Snap25 expression
following knockdown and muscone treatment of PC12 cells exposed to OGD/R. GPX4
activity (C) and GSH (D) levels after Snap25 knockdown and muscone administration
in OGD/R-injured PC12 cells. In addition, the ROS (E), MDA levels (F), and cell
viability (G) were assessed after Snap25 silencing and muscone treatment in PC12
cells subjected to OGD/R injury. (H) Representative TEM images after Snap25
silencing and muscone treatment of PC12 cells exposed to OGD/R. The red arrows
indicate mitochondria. Scale bar, 500 nm. One-way ANOVA compares the means of
multiple groups. Data shown are the mean
WB analysis established the effective Snap25 upregulation in the
Snap25-over expression (OE) group in comparison to the NC group (Fig. 7A; p
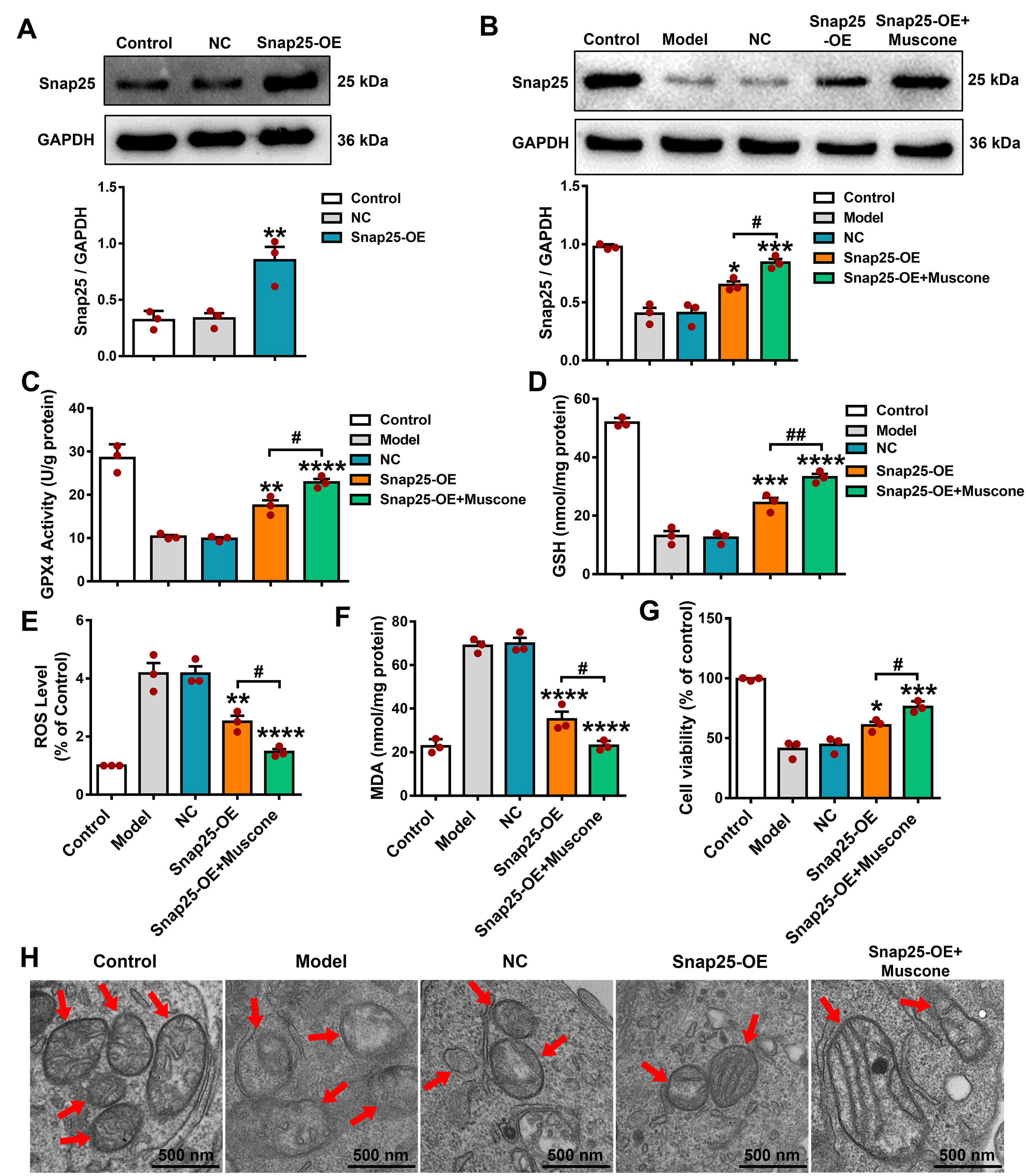 Fig. 7.
Fig. 7.
Overexpression of Snap25 reduced the ferroptosis level and
strengthened the anti-ferroptosis effects of muscone in OGD/R-injured PC12
cells. (A) WB analysis confirmed the Snap25 overexpression in PC12 cells. (B)
Following Snap25 overexpression and muscone administration in PC12 cells exposed
to OGD/R damage, significant increases were observed in GPX4 activity (C) and GSH
levels (D). Furthermore, Snap25 overexpression combined with muscone treatment
led to decreases in ROS (E) and MDA levels (F), increased cell viability (G), and
attenuation of mitochondrial injury (H) in PC12 cells subjected to OGD/R. The red
arrows indicate mitochondria. Scale bar, 500 nm. One-way ANOVA compared the means
of multiple groups. Data shown are the mean
Molecular docking experiments predicted the binding sites of muscone and Snap25.
This revealed 8 potential amino acid residues (Ser-25, Ser-28, Thr-29, Met-32,
Ile-129, Phe-133, and Ile-134) involved in the interaction between muscone and
Snap25 protein (Fig. 8A,
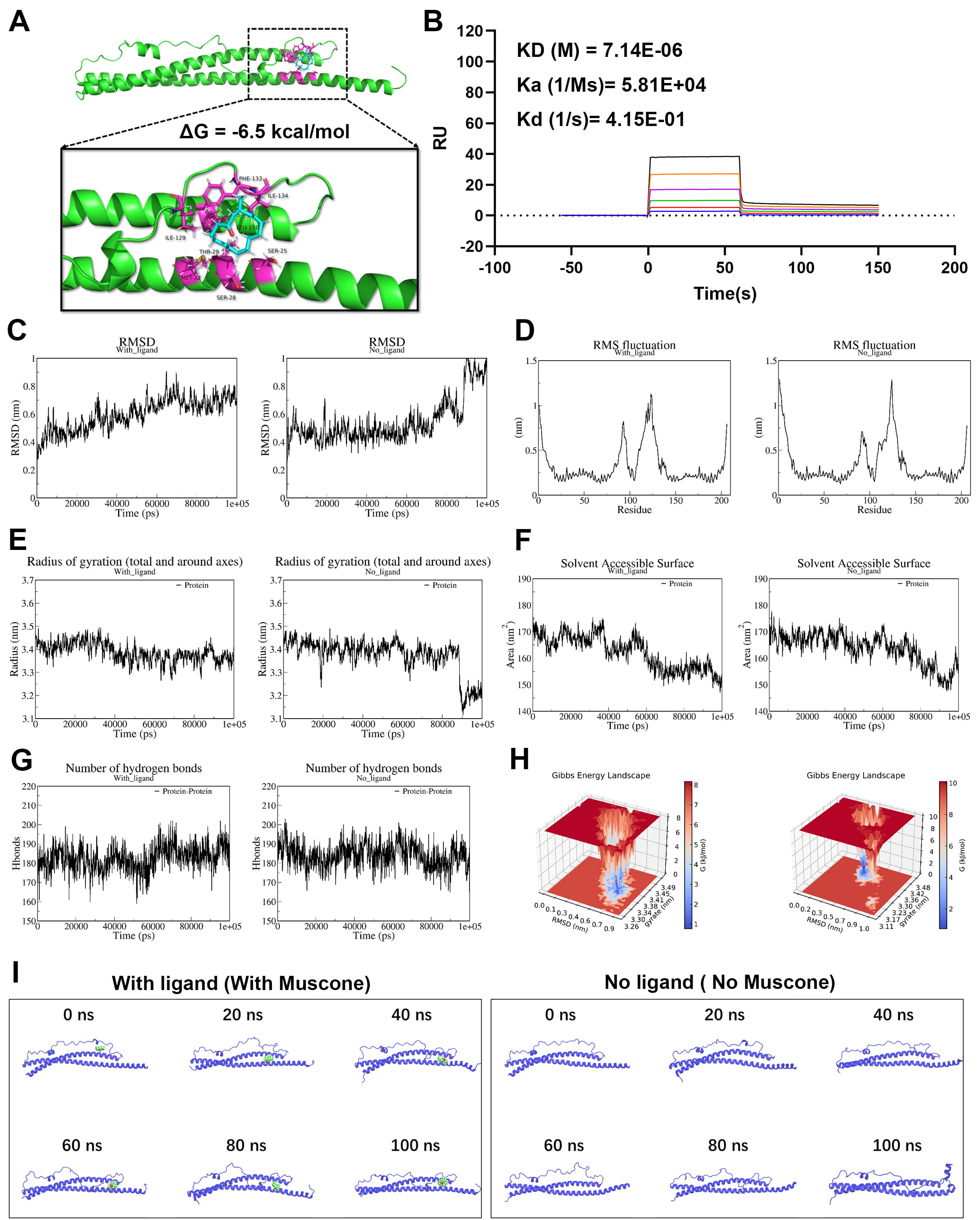 Fig. 8.
Fig. 8.
Muscone binds to Snap25 protein and enhances its structural
stability. Molecular docking tests were conducted to predict the binding sites
between muscone and Snap25. (A) Stereoview of the binding mode for Snap25 with
muscone. (B) Surface plasmon resonance (SPR) analysis showing the direct binding
effect between muscone and Snap25 protein (KD = 7.14
The multifaceted pathogenesis of AIS means the therapeutic window is narrow, resulting in severely compromised neurological function. Drug therapy for AIS has so far failed to produce satisfactory clinical outcomes. Therefore, it is important to elucidate the specific pathogenic mechanism of AIS to facilitate the development of effective drugs. The key findings of this investigation can be summarized as follows: (1) markedly elevated ferroptosis levels were observed in both the MCAO/R model and OGD/R-treated PC12 cells; (2) muscone significantly alleviated AIS-triggered damage both in vivo and in vitro by effectively decreasing ferroptosis levels; (3) binding of muscone to the Snap25 protein enhances its expression and structural stability, thereby reducing the ferroptosis level in AIS damage.
The role of ferroptosis in stroke pathology has recently gained significant attention. Multiple studies have suggested that AIS is closely related to ferroptosis [39, 40, 41]. AIS-damaged rat models with excessive iron intake were found to have severe neuronal damage. Moreover, iron chelating agents can decrease reperfusion damage in AIS animal models, suggesting that neuron injury in AIS may be closely related to iron accumulation [42]. An excessive increase in the ROS level is the main cause of ferroptosis [43]. Intracellular lipid molecules, such as cell membranes, are the main target affinity of ROS [44]. Therefore, excessive ROS levels result in a higher level of lipid peroxidation [45, 46]. Herein, we detected substantial increases in ROS and lipid peroxidation levels, as well as significant mitochondrial ridge breakage, in in vivo and in vitro AIS models. These findings further confirm the pivotal participation of ferroptosis in the pathological progression of AIS and offer a mechanistic foundation for potential therapeutic interventions that target AIS injury.
For centuries, the TCM compound musk has been employed in treating stroke injuries. The potential anti-stroke properties of muscone, the principal active constituent of musk, have been widely recognized. Muscone is a small active molecule that can enter the blood-brain barrier and exert neuroprotective effects. Previous studies have suggested that muscone may be protective against AIS injury by promoting microglia polarization or by inhibiting cell apoptosis [47, 48]. Herein, we detected a significant reduction in infarct volume following muscone treatment of MCAO/R rats, as well as increased survival of OGD/R cells. Furthermore, muscone treatment in the AIS model resulted in notable improvements in mitochondrial morphology, increased GPX4 activity and GSH level, and decreased ROS and MDA levels, thus indicating an anti-ferroptosis effect. However, the precise mechanism by which muscone regulates ferroptosis in AIS remains unclear. Snap25 has an essential function in various neurological diseases and pathological conditions, and is significantly correlated with AIS injury. Research has demonstrated that Snap25 expression in mouse models of ischemic stroke is significantly decreased in the injured brain tissue. However, upon expression of Snap25 there is marked improvement in the synaptic structure of ischemia-injured neurons [49]. RNA sequencing and bioinformatics analysis in this investigation revealed a noticeable decrease in Snap25 expression in the in vitro and in vivo AIS models, thus corroborating previous findings. A deficiency in Snap25 leads to neurotransmitter transmission disorders, increased ROS levels, and neuroinflammation, which may be key factors in Snap25-induced ferroptosis [50]. Our previous studies and current research indicate that Snap25 regulates the level of ferroptosis in AIS. Through a series of experiments, including Snap25 silencing and overexpression, this investigation detected Snap25 as a target of muscone. Specifically, muscone mitigated ferroptosis during AIS injury by upregulating the expression of Snap25 protein. We conducted molecular docking, SPR detection, and MD simulation experiments to illuminate the mechanism by which muscone increases the Snap25 protein expression. These revealed multiple potential binding sites and a direct binding effect between muscone and the Snap25 protein. Moreover, muscone can significantly enhance the structural stability of Snap25 and prevent its degradation under conditions of AIS injury. Our results provide a molecular basis for the substantial upregulation of Snap25 expression following muscone treatment, leading to mitigation of ferroptosis injury in AIS.
This research study has certain limitations. Although we examined the mechanism by which muscone controls the expression and stability of Snap25 protein and protects against ferroptosis-induced injury through in vitro silencing and overexpression experiments, further in vivo validation experiments are necessary. In subsequent studies, we intend to utilize adeno-associated, virus-mediated infection techniques to conduct Snap25 silencing and overexpression experiments at the animal level, thereby further confirming the specific interaction between muscone and Snap25 protein. Future research should also investigate the therapeutic potential of muscone in various subtypes of ischemic stroke injuries, including its capacity to alleviate small deep brain infarctions associated with lacunar syndrome [51].
This study confirmed the correlation between AIS and ferroptosis, while specifically investigating the protective mechanism of muscone in AIS. Our research findings indicate that muscone can attenuate ferroptosis during AIS injury by increasing the expression of Snap25. Consequently, this research provides additional evidence supporting a close association between AIS injury and ferroptosis. This could potentially lead to the development of more effective therapeutic medications for clinical application in AIS.
Datasets generated for this study are available upon request from the corresponding author.
RJY conducted the animal and cell experiments and drafted the manuscript. BS and JL performed the animal and cell experiments and analyzed the experimental data. GHH carried out the GSH and GPX4 activity analyses. NS conducted the ROS and CCK-8 assays. BS, JL, GHH and NS revised the manuscript critically for important intellectual content. WWS performed molecular docking and molecular dynamics simulations, provided technical guidance on the experiments, and reviewed and edited the manuscript. All authors have read and approved the final version of the manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
The Ethical Committee of Animal Experiments of the Anhui University of Chinese Medicine (Hefei, China; approval no. AHUCM-rats-2023126) specifically approved animal experiments. The study was carried out in compliance with ARRIVE guidelines.
Not applicable.
This research was supported by the National Natural Science Foundation of China (grant no. 82374209 and 82104570), the Anhui Provincial Natural Science Foundation (grant no. 2208085Y33), the Talent Project of Anhui University of Chinese Medicine (grant no. 2022rczd007), and the Natural Science Research Projects at Higher Institutions in Anhui Province (grant no. 2023AH050748).
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/JIN39116.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.