

 , Emmanuel Rodriguez-Nava 1,2, Gabriel Roldan-Roldan 1, Donald B. Katz 3,4, Jean-Pascal Morin 1,*
, Emmanuel Rodriguez-Nava 1,2, Gabriel Roldan-Roldan 1, Donald B. Katz 3,4, Jean-Pascal Morin 1,*
1 Department of Physiology, Faculty of Medicine, National Autonomous University of Mexico (UNAM), 04510 Coyoacan, Mexico City, Mexico
2 Section of Postgraduate Studies and Research, School of Medicine, National Polytechnic Institute, 11340 Miguel Hidalgo, Mexico City, Mexico
3 Department of Psychology, Brandeis University, Waltham, MA 02453, USA
4 Program in Neuroscience, Brandeis University, Waltham, MA 02453, USA
Abstract
The basolateral amygdala (BLA) is crucial for assigning emotional valence to sensory experiences, driving approach or avoidance behaviors during subsequent encounters. Particularly, the BLA plays a critical role in the coding, storage and retrieval of emotional learning. While traditionally viewed through the lens of memory consolidation, cholinergic signaling—mediated by dense inputs from the basal forebrain and abundant muscarinic receptors (mAChRs) in the BLA—plays a far more dynamic role. Acetylcholine, often described as a “memory molecule”, is central to this process, with scopolamine induced amnesia models underscoring its importance. Recent evidence suggests that cholinergic activity not only supports memory formation but also imparts emotional valence under specific conditions. This review examines the molecular and cellular mechanisms by which mAChR-mediated cholinergic signaling modulates BLA processing and the storage of emotional memories. We integrate psychopharmacological insights with loss and gain-of-function studies to demonstrate how cholinergic signaling in the BLA shapes approach and avoidance behaviors. Based on this evidence, we propose that acetylcholine’s influence in the BLA is highly context-dependent, reflecting its versatile role in emotional processing beyond mere memory consolidation.
Keywords
- basolateral amygdala
- acetylcholine
- muscarinic receptor
- learning
- avoidance learning
- appetitive behavior
The self has been suggested to be composed of two parts: the experimenting self, which lives in the experience of the present moment; and the remembering self, which lives in the stories we later tell ourselves about such experiences [1]. The emotional tone attached to stimuli experienced by the experimenting self, as well as the thinking about those stimuli that is performed by the remembering self, has profound evolutionary roots and is biased towards the negative in most psychiatric disorders [2]. Considerable progress has been made in the last few years toward understanding how the brain codes, stores and retrieves emotionally relevant experiences, and about the brain structures involved in these processes. Among these structures, the basolateral amygdala is thought to play a crucial role [2, 3]. In that regard, recent studies have revealed genetically and projection-specific neuronal populations within the basolateral amygdala (BLA) that appear to assign positive or negative valence to experiences in both rodents [4, 5] and humans [6]. These same circuits have been shown to be disrupted in mice models of depression and similar disruptions have been shown to underlie negative bias in patients with depression [7].
Experience-dependent neuroplasticity mechanisms within BLA circuits have been shown to involve neuromodulatory inputs from monoamine, and neuropeptide-releasing neurons, and from neurons releasing acetylcholine (ACh) [8, 9, 10]. Slight disturbances in these inputs have been linked to maladaptive conditions including to conditions involving maladaptive learning such as post-traumatic stress disorder (PTSD) and addiction [11, 12, 13]. Therefore, a crucial goal of modern neuroscience research is to deepen our understanding of how experience-dependent neuromodulatory events in the BLA and other regions of the limbic system may, under certain conditions such as prolonged stress or genetic vulnerability, may produce attractor states that favor the activation of negative valence neural ensembles [14]. In recent years, the central cholinergic system, and its association with BLA, has drawn attention for its role in valence processing, affect and mood [15, 16, 17].
In this review article, we discuss the role of acetylcholine in the codification and storage of emotional memory within BLA networks, with a special emphasis on recent psychopharmacological, cellular and molecular findings. We start with an overview of the BLA as a structure that is central to the formation of emotional memories of both positive and negative valence. Next, we discuss how cholinergic modulation affects its dynamics at the cellular/molecular level and finally, we delve into recent psychopharmacological findings involving the cholinergic system. We mainly focus on muscarinic signaling; for recent reviews on the role of BLA nicotinic receptors on emotional learning and memory, the reader is kindly referred to recent reviews on the subject [18, 19]. We start with a brief overview of valence coding in BLA circuits. For more detailed reviews the reader is referred to previous published work [2, 5, 20].
The BLA, which is located within the medial temporal lobe of mammals, contains cytoarchitecture resembling that of the cerebral cortex [21]. Although not organized in layers, the vast majority (~80–90%) of its neurons are pyramidal, with magnocellular types mostly present in its anterior part (BLAa) and intermediate and parvocellular types in its posterior part (BLAp) [17, 22, 23] and the rest are parvalbumin-positive (PV), somatostatin (SOM)-positive, cholecystokinin-γ-aminobutyric acid-positive (CCK-GABA) and calretinin/vasoactive intestinal peptide (VIP)-expressing GABAergic interneurons (INs) [17, 24, 25].
Perhaps the most well-studied function of the BLA has to do with coding and storing emotional valence, both positive and negative. The participation of the BLA in these tasks has long been established [26] and findings in the last decade or so have greatly improved our ability to identify positive and negative valence-specific neural networks within the BLA that can be modified by experience. This endeavor has benefitted from the development of techniques that enable activated-neuron targeting and manipulation via the tagging of cells expressing immediate early genes (such as c-fos) that mark neuronal activity [27, 28]. One such study revealed that rostral magnocellular neurons expressing roof plate-specific spondin-2 (Rspo2) mRNA drive avoidance behavior, whereas posterior parvocellular neurons expressing protein phosphatase 1 regulatory subunit 1B (Ppp1r1b) drive approach behaviors [22]. A follow-up study by the same group further revealed that fear extinction memories, in which the omission of an aversive stimulus is rewarding [29], are stored in an engram including BLA Ppp1r1b+ appetitive cells [30], providing evidence that valence-specific neurons within the BLA provide emotional tone to specific memories [26].
Findings from in vivo single-unit recordings however did not observe a segregation of valence encoding along the anteroposterior axis but did so along the dorsoventral axis where most cues related to the innately aversive taste quinine were found in the dorsal BLA, while sucrose-related cues were mostly present in the ventral part [31]. Meanwhile, using neuronal activity monitoring through in vivo calcium imaging, another study found that BLA neurons expressing transcription factor-coding mRNA Fezf2, which consist of a subpopulation of Rspo2+ neurons, were more heterogeneous in their activation pattern, with some responding to rewarding stimuli and others responding to aversive ones. They further established that whether these neurons were positive or negative valence-driven depended on their specific projection targets in the ventral stratum [32], that is, BLA pyramidal neurons (PN) projecting to the Olfactory Tubercule promoted the encoding, learning and execution of reward-based tasks whereas those projecting to the Nucleus Accumbens were involved in avoidance behavior.
A still more recent study using holographic stimulation did report both antero-posterior and dorsoventral gradients of valence representation consistent with earlier studies [22, 31] but also suggested that negative and positive valence-specific neurons in the BLA are largely intermingled [33], consistent with earlier findings from single-unit recordings in primates [34]. In summary, these studies demonstrate the presence of both positive and negative valence-specific neurons in the BLA although their precise location and identity may be task-specific and appear to be highly sensitive to the methodology employed for their identification.
A crucial question remains however as to how these valence-specific engrams are “selected” through experience to be associated with specific memories or sensory experiences. As mentioned above, the BLA contains neurons that specifically code valence, including neurons that are specifically activated by innately aversive experiences such as pain [35]via sensory coding regions including the parabrachial nucleus, the mediodorsal thalamic nucleus and the insular cortex [36, 37].
In addition, research has shown that BLA output neurons undergo synaptic plasticity as the animal learns positive and negative associations. Furthermore, the direction of synaptic plasticity, that is, depression or potentiation, depends on the specific targets of these BLA PNs. Specifically, mouse BLA neurons projecting to the centromedial amygdala undergo an increase in the α-Amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) / N-methyl-D-aspartate (NMDA) receptor ratio (a proxy of long-term potentiation -LTP- expression) after mice are subjected to fear conditioning to a tone and this ratio decreases in reward conditioning in which a tone is paired with sucrose. A double dichotomy was established when the authors observed the opposite effects on BLA neurons projecting to the nucleus accumbens, in which reward learning produced synaptic potentiation and aversive learning produced synaptic depression [38]. This provided a mechanism explaining earlier psychopharmacological findings whereby NMDA receptor antagonists applied to this region blocked both appetitive and aversive associative memory [39, 40].
But although converging glutamatergic inputs to BLA PN may induce synaptic plasticity at BLA synapses, they do not perform this work alone. It appears, rather, that concomitant neuromodulator release contributes to that learning-related synaptic strengthening [41], facilitating associative memory formation, particularly when a long delay elapses between the cue and the aversive/appetitive consequence [10, 42, 43]. As the BLA is densely innervated by neuromodulatory inputs, including dopaminergic, serotoninergic, noradrenergic and especially cholinergic [44], it has taken a good deal of research to evaluate how these inputs could promote synaptic plasticity in the BLA during storage and retrieval of emotional experiences.
Salient stimuli, whether aversive or appetitive, increase dopamine release in the basal amygdala, priming neurons for plasticity by increasing intracellular cAMP levels [45]. Norepinephrine is also known to promote LTP at thalamo-amygdala synapses, possibly through GABAergic signaling suppression [46]. Norepinerphrine signaling in the BLA also increased stress-induced firing of BLA neurons and prevented extinction of a weak fear conditioning [47]. Conversely, dopaminergic signaling has been shown to promote long-term depression (LTD) at BLA-intercalated cells synapses by enhancing presynaptic GABAergic transmission, a mechanism that could prevent fear memories from being generalized as it is observed in PTSD [48]. Surprisingly, although the role of ACh signaling in the hippocampus and cortex has been extensively studied, very little was known until recently regarding its role in the BLA, even though this region receives the densest cholinergic projections from the basal forebrain [24, 49]. The next section will review recent evidence on the cellular and molecular impact of ACh signaling in the BLA and how it could promote emotional learning. We next link this evidence to recent findings from behavioral studies. As stated earlier, in the present text, we will mainly focus on muscarinic signaling.
Projection neurons from the basal forebrain cholinergic system (BFCS) are distributed across an extended region comprising the medial septum (MS), the vertical and horizontal limbs of the diagonal band of broca (DBB), the sustancia innominata (SI), the ventral pallidum (VP) and the nucleus basalis magnocelularis (NMB). Cholinergic neurons from these nuclei project to a wide array of cortical and subcortical targets including the medial prefrontal cortex (mPFC), the insular cortex (IC), the hippocampus and the BLA. Acetylcholine exerts its effect on these structures through activation of two main types of receptors: the nicotinic receptors, which are pentameric ionotropic receptors and muscarinic acetylcholine receptors (mAChRs), which are metabotropic, G-protein coupled receptors. The distribution and specific roles of these receptors has been extensively described elsewhere [24]; we will thus only provide a brief overview here.
Five subtypes of mAChRs have been identified, three Gq-coupled and mainly postsynaptic (M1, M3 and M5) and two, mainly presynaptic Gi-coupled receptors (M2, M4) [50]. All mAChRs subtypes (save M5) have been identified in the mammalian BLA through immunohistochemical and electrophysiological studies [51, 52, 53]. We will now summarize what is known about each in turn.
M1 receptors are almost exclusively present in PNs and are especially abundant
in the anterior two-thirds of the BLA [51]. Cholinergic projections from the
basal forebrain to the BLA are known to stimulate these receptors [43] at which
time the response of PNs appears to depend on their basal activity state. During
resting states, M1 stimulation inhibits PNs by a mechanism that depends on the
activation of G-protein-gated K+ inward rectifying channels (GIRK), leading to
postsynaptic neuron hyperpolarization. Stimulated at high-frequencies (
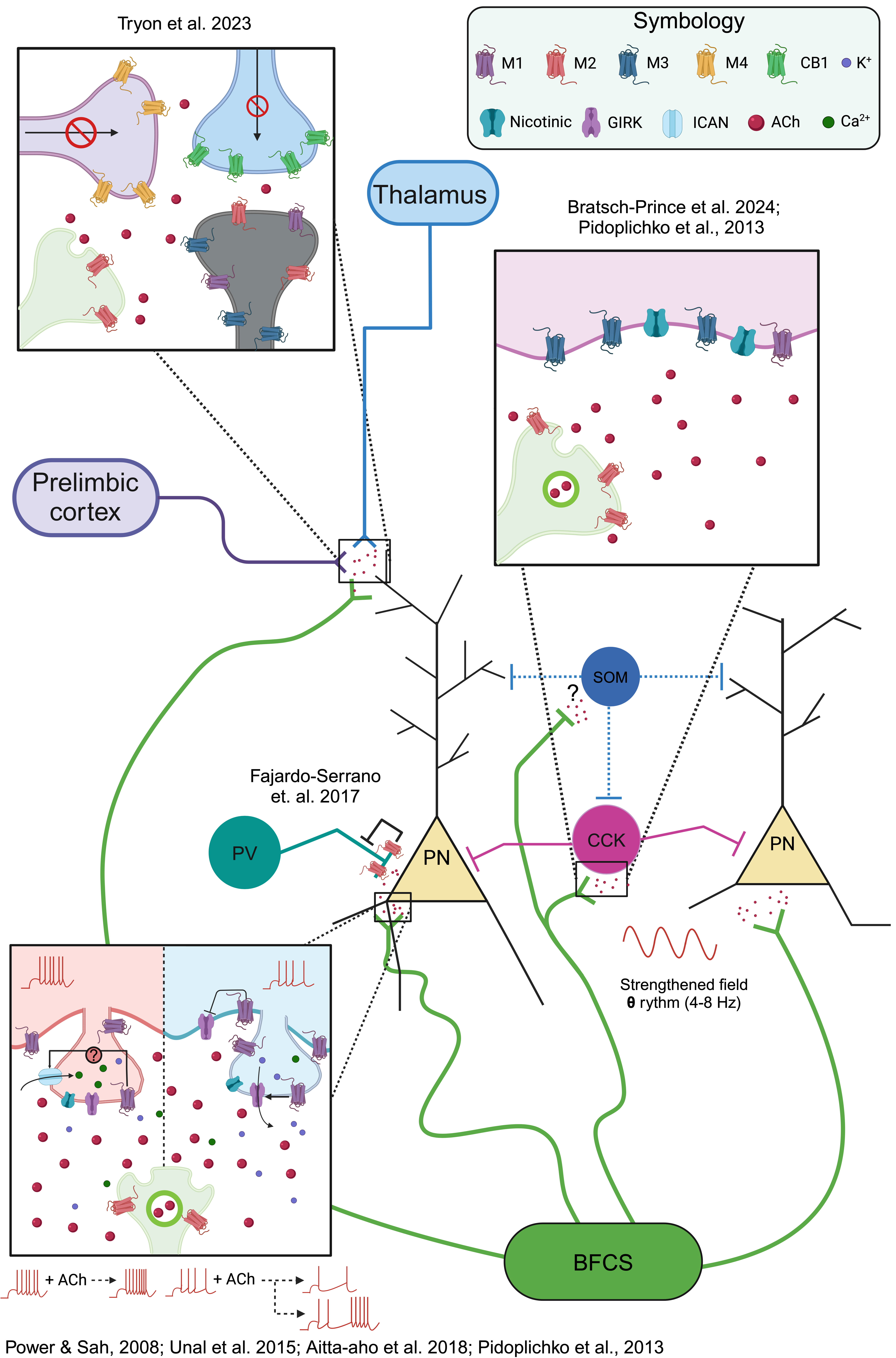
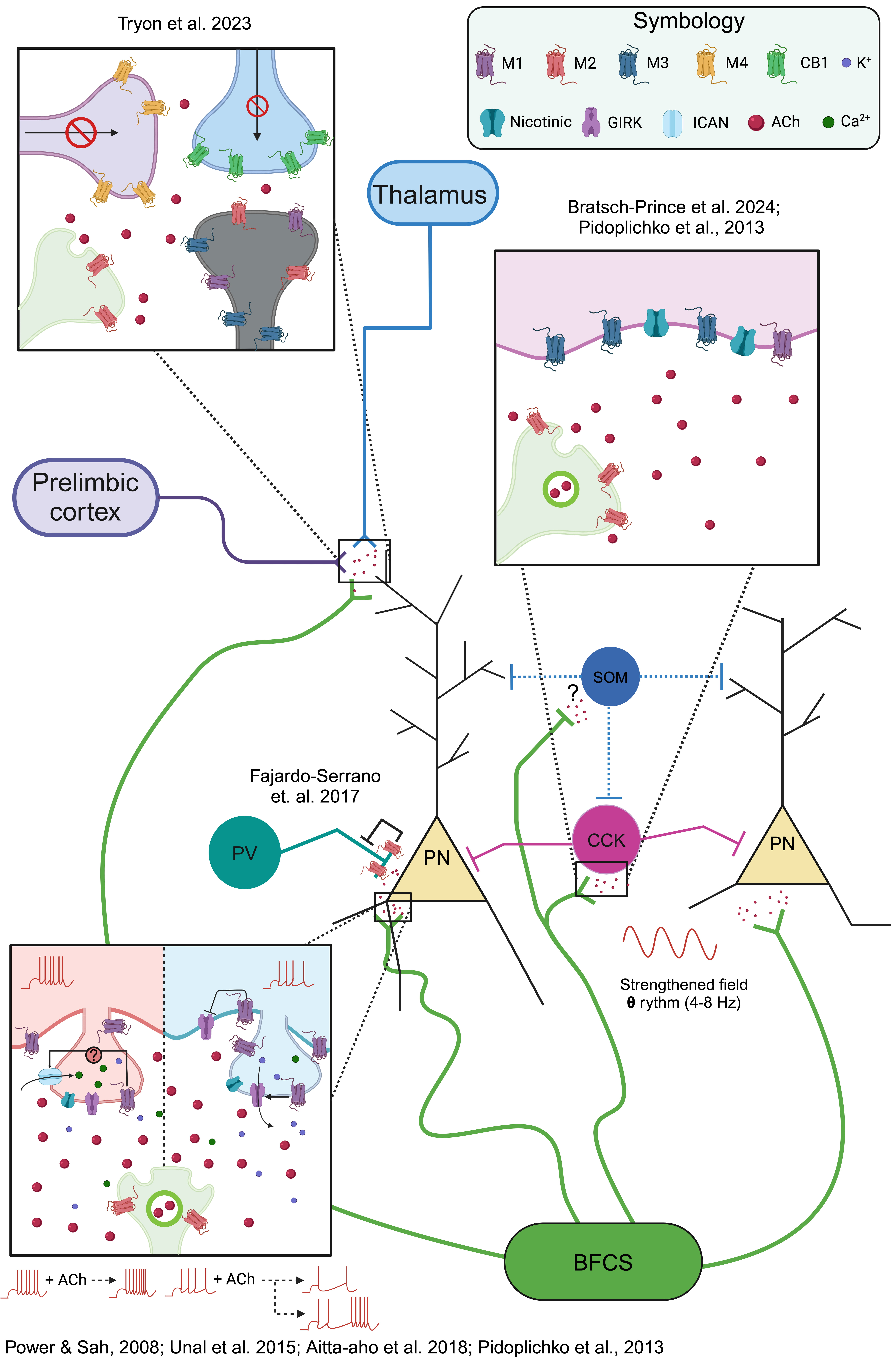 Fig. 1.
Fig. 1.
Schematic figure showing cellular mechanisms of cholinergic inputs in BLA through interactions mediated by mAChRs (M1-M4). M1 receptors activation of a PN with high depolarizing current, promotes excitation by the activation of ICAN through an unknown mechanism. In contrast, in PNs with low firing rate, M1 receptor stimulation produces inhibition by the activation of GIRK channels. Meanwhile, the activation of the Gi-coupled M2 receptor inhibits GABA release from PV INs axons. Gq-coupled M3 receptors stimulation on the other hand, increase the activity of CCK INs synchronizing BLA firing producing theta oscillations. Also, stimulation of M3 receptors produces inhibition of thalamic afferents to the BLA trough a mechanism dependent on retrograde endocannabinoid release followed by activation on Gi-coupled CB1 receptors at the thalamic terminals. Finally, M4 receptors activation suppresses glutamate release in Prelimbic Cortex inputs firing at low frequency, allowing high frequency signals to reach BLA. The inset provides a color key for the receptor types shown. BFCS, basal forebrain cholinergic system; PN, pyramidal neuron; PV, parvalbumin; CCK, cholecystokinin; ICAN, calcium-dependent non-selective cation channels; GIRK, G-protein-gated K+ inward rectifying channels; M1 and M4, muscarinic receptor types 1 and 4, respectively; CB1, cannabinoid receptor 1; ACh, acetylcholine; BLA, basolateral amygdala; GABA, γ-aminobutyric acid; INs, interneurons. Created in https://www.Biorender.com.
M2 receptors are present in the somata of INs, in the dendritic regions of both INs and PNs and in axon terminals of the anterior part of the BLA [24, 52], suggesting a role not only as autoreceptors, providing a negative feedback loop to further ACh release, but also at modulating glutamatergic inputs from the cortex [60], or GABAergic liberation form local INs [55]. In this regard, M4-type receptors at glutamatergic terminals from the prelimbic cortex were recently identified to have a similar function, wherein their activation inhibits glutamate release. Indeed, ACh in the BLA, likely by way of volume transmission, produces strong M4-dependent inhibition of cortical glutamatergic inputs firing at low frequency and a weaker and concomitant M3-receptor-mediated, endocannabinoid-dependent suppression of glutamatergic transmission at thalamo-amygdalar synapses. As the authors suggest, this mechanism could provide a mechanism by which attention to salient external cues is enhanced during learning [53]. Importantly, this M4-mediated glutamate release suppression is thought to be suppressed under conditions of strong membrane depolarization of the presynaptic membrane which would occur under conditions of high frequency stimulation from the cortex, again in case of salient information or perhaps under pathological conditions.
This, and another recent finding from the same group, has introduced the M3 receptor as an exciting new player in BLA physiology. Specifically, these researchers have shown ACh release to act through this receptor to activate GAD67/CCK INs, enhancing perisomatic inhibition of PN, in turn synchronizing BLA PN activity in the theta range [56]. That ACh release concomitantly induced distal dendritic spine disinhibition via SOM IN silencing, which could also contribute to long-term emotional memory formation by facilitating Hebbian plasticity at asymmetric synapses [56].
In addition to indirect acetylcholine-induced inhibition of BLA PNs through M3
activation, cholinergic signaling also inhibits PNs, specifically those
projecting to the mPFC, through direct monosynaptic stimulation of PN mAChRs
[61]. This is consistent with in vitro studies suggesting that more than
one type of muscarinic receptor is involved in the suppression of PN excitability
[62]. PNs may also undergo disynaptic inhibition through nicotinic receptor
dependent IN activation [61], possibly through activation of
Cholinergic signaling has long been understood to play a crucial role in the strengthening of synapses and other neuroplasticity mechanisms underlying memory formation [63, 64]. Although most behavioral literature evaluating the effects of acetylcholine on learning and memory have focused on the hippocampus and cortex [64, 65, 66, 67], a considerable number of studies have revealed the importance of cholinergic signaling in the BLA during memory formation (Table 1, Ref. [42, 43, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83]). In the case of the BLA however, we argue that overall, the data suggest that the requirements for cholinergic signaling are strikingly distinct for the learning of approach versus avoidance behavior. Relatedly, we suggest that the bulk of the evidence regarding the role of muscarinic signaling in the BLA strongly supports a view in which the brain’s default mechanism is to store an experience as aversive.
| Author | Receptor | Technique | Outcome | |
| Mechanism | Aitta-aho et al., 2018 [54] | Muscarinic | Optogenetic stimulation of basal forebrain cholinergic projections to BLA. | Basal forebrain cholinergic projections to the BLA synchronize local field potentials at theta frequency, muscarinic-dependent. |
| Bratsch-Prince et al., 2024 [56] | M3 | Electrophysiology recordings in BLA. | ACh induces field theta oscillations in BLA through M3 receptor-induced CCK interneurons activation. | |
| Fajardo-Serrano et al., 2017 [55] | M2 | Electron microscopy, immunohistochemistry and electrophysiology in BLA. | M2 receptor modulates inhibitory transmission in BLA by suppressing GABA release in interneurons. | |
| Kimchi et al., 2024 [61] | Muscarinic & Nicotinic | Optogenetics, fiber photometry, behavioral analysis, fluorescent imaging in BLA. | Basal forebrain cholinergic projections to BLA cause muscarinic-dependent IPSPs onto PNs and nicotinic receptor-dependent EPSPs onto INs. | |
| Lee & Kim, 2019 [68] | Muscarinic & nicotinic | Optogenetic stimulation of basal forebrain cholinergic inputs to BLA. | Cholinergic activation inhibits BLA pyramidal neurons via CCK interneurons. | |
| McDonald & Mascagni, 2010 [51] | M1 | Immunohistochemical localization of M1 receptors in BLA. | M1 receptors are localized in PNs of the BLA; more abundant in the rostral part. | |
| McDonald & Mascagni, 2011 [52] | M2 | Immunohistochemical localization of M2 receptors in BLA. | M2 receptors are localized in GABAergic SOM and NPY soma and axons. | |
| Muller et al., 2016 [60] | M2 | Electron microscopy in BLA dendrites, spines, and axon terminals. | M2 receptors in BLA located pre- and post-synaptically at glutamate, GABA, and cholinergic synapses. | |
| Pidoplichko et al., 2013 [57] | Muscarinic & nicotinic | Whole-cell recordings from BLA interneurons. | Cholinergic signaling balances inhibition and excitation in the BLA. | |
| Power & Sah, 2008 [58] | Muscarinic | Whole-cell patch-clamp in BLA projection neurons. | Muscarinic activation suppresses afterhyperpolarization, enhancing neuron excitability in the BLA. | |
| Tryon et al., 2023 [53] | M3, M4 | Electrophysiological recordings in BLA slices, optogenetic stimulation of cholinergic terminals. | ACh inhibits cortical and, to a lesser extent, thalamic inputs to the BLA in a pathway-specific and frequency-dependent manner. | |
| Unal et al., 2015 [43] | M1 | Optogenetic stimulation of basal forebrain and patch-clamp recordings of BLA neurons. | M1 receptors induce positive feedback on activity level of BLA PNs, enhancing contrast of excitability among these neurons. | |
| Yajeya et al., 2000 [62] | Muscarinic | Electrophysiological recordings in the BLA. | Carbachol reduces excitatory synaptic transmission through presynaptic inhibition of glutamate release. | |
| Womble & Moises, 1993 [69] | Muscarinic | Voltage-clamp and current-clamp recordings of pyramidal neurons in the BLA. | Muscarinic activation inhibits afterhyperpolarization, increasing excitability in BLA neurons. | |
| Avoidance Behaviors | Barros et al., 2002 [70] | Muscarinic | Inhibitory avoidance | Oxotremorine enhances, and scopolamine impairs, working and long-term memory. |
| Baysinger et al., 2012 [42] | Muscarinic | Trace fear conditioning | Acetylcholine is required for trace fear conditioning only when there is a long interval between the CS and US. | |
| Carballo-Márquez et al., 2011 [71] | Muscarinic | Two-way active avoidance | Scopolamine facilitates active avoidance learning. | |
| Crimmins et al., 2023 [72] | Nicotinic | Fear conditioning | Cholinergic signaling in the BLA during acquisition promotes fear memory strength and persistence. | |
| Jiang et al., 2016 [59] | Muscarinic & nicotinic | Fear conditioning | Concomitant muscarinic & nicotinic receptor antagonism, but not muscarinic antagonism alone, impairs retention. | |
| Malin & McGaugh, 2006 [73] | Muscarinic | Fear conditioning (context and tone) | Agonist oxotremorine enhances memory of both context and footshock. | |
| Morin et al., 2021 [74] | Muscarinic | Conditioned Taste Aversion (CTA) | Blocking mAChRs with scopolamine in the BLA impaired the acquisition of CTA and partially affected retrieval. | |
| Vazdarjanova & McGaugh, 1999 [75] | Muscarinic | Contextual fear conditioning | Post-training activation of mAChRs in the BLA with oxotremorine resulted in enhanced memory. | |
| Young & Thomas, 2014 [76] | M1 | Fear conditioning | The activation of M1 receptors in the BLA enhances fear memory consolidation. Loss-of-function experiments unveiled redundant neuromodulator systems involvement in fear memory consolidation. | |
| Approach Behaviors | Aitta-aho et al., 2018 [54] | Cholinergic | Place preference | Cholinergic activation correlates with and causes approach behavior. |
| Boccia et al., 2009 [77] | Muscarinic | Contextual fear conditioning and extinction | Muscarinic activation enhances extinction memory consolidation, reducing freezing behavior. | |
| Carballo-Márquez et al., 2009 [78] | Muscarinic | Social transmission of food preference | Muscarinic blockade in BLA impairs social transmission of food preference in rats. | |
| Crouse et al., 2020 [79] | Muscarinic | Cue-reward learning | Blocking mAChRs impairs cue-reward learning. | |
| Kimchi et al., 2024 [61] | Muscarinic & nicotinic | Windows of Opportunity Task with reward-seeking | Cholinergic efflux in BLA underlies appetitively conditioned behavior | |
| McIntyre et al., 1998 [80] | Muscarinic | Conditioned place preference | Scopolamine infusion intra-BLA impairs performance in conditioned place preference tasks. | |
| Ortiz-Guzman et al., 2024 [81] | Muscarinic & nicotinic | Feeding behavior | Cholinergic projections from the DBB to the BLA suppresses appetite by modulating BLA neuron activity. | |
| See et al., 2003 [82] | Muscarinic | Operant conditioning | Blocking mAChRs impairs cocaine-stimulus learning, reducing approach toward drug-paired cues. | |
| Torres-García et al., 2024 [83] | Muscarinic, M1, M3 | Incidental taste learning | Intra-BLA infusions of scopolamine after saccharin intake induces strong conditioned taste avoidance. |
IPSPs, inhibitory post-synaptic potentials; Ins, interneurons; EPSPs, excitatory post-synaptic potentials; SOM, somatostatin; NPY, neuropeptide Y; CS, conditioned stimulus; US, unconditioned stimulus; ACh, acetylcholine; DBB, diagonal band of broca; mAChRs, muscarinic receptors.
Studies focusing on the hippocampus have consistently reported that muscarinic
receptor activity in this region is necessary for (context) memory consolidation,
irrespective of whether it involves the learning of approach or avoidance
behavior [65]. In the case of the BLA however, data reporting a requirement for
muscarinic signaling in avoidance learning, especially in the post-acquisition
period, is rather scarce. At the turn of the century, the amygdala, which was
already known to receive dense cholinergic projections from the basal forebrain
[24, 84] was mostly viewed as specialized in processing aversive sensory stimuli
and storing fear memory [85]. This was at a time when the cholinergic theory of
memory dysfunction [86] was still gaining ground and effort was made to determine
the involvement of cholinergic signaling in the amygdala in aversive memory
formation. In this regard, while several psychopharmacological studies did report
a role of muscarinic signaling in the BLA in avoidance learning tasks, most of
the findings were limited to cases where muscarinic stimulation (such as with the
non-specific agonist oxotremorine) boosts memory [73, 75, 87] or where
pharmacological fear memory enhancement by glucocorticoid or
Some recent evidence however does indicate that under specific circumstances muscarinic signaling in the BLA is indeed necessary for long-term aversive memory formation. As alluded to in the previous section, several lines of evidence suggest that acetylcholine could act to facilitate the integration of a conditioned and an unconditioned stimulus when a longer time lapse occurs between the two. For instance, a study using observed that mAChRs in the amygdala are required trace fear conditioning only when a 17 s “silence” period occurred between the tone and the foot shock [42]; no effect is observed when the foot shock immediately follows the tone. Similar results hold for conditioned taste aversion (CTA) in which a taste is associated with post-ingestive malaise that is sometimes delayed by hours [67]: research from our group has shown that muscarinic receptor antagonism with scopolamine in the BLA during novel taste consumption abolished CTA formation but that the same treatment applied immediately after conditioning has no effect; only a partial reduction of CTA was observed when scopolamine was infused before test [73]. Importantly also, scopolamine infused either in the central amygdala or the BLA was without effect when administered after conditioning, in strike contrast with its effect in approach taste learning (see below). Therefore, in the case of associative aversive memory formation, it is possible that BLA mAChRs activation is “strictly” necessary only if a substantial delay occurs between the cue and the reinforcer and that the neurochemical mechanisms underlying more contingently paired conditioned and unconditioned stimuli are more redundant [75]. Overall, these findings showed that rather than being required for aversive memory consolidation, BLA muscarinic signaling modulates the strength and possibly duration of such memories.
Also, while the non-selective muscarinic receptor antagonist scopolamine has long been used and been recognized as an amnesia-inducing agent [88, 89, 90, 91], some investigations that tested the effects of infusing this drug in the BLA, however, suggest that mAChRs in this region may have distinct roles, beyond memory consolidation. For example, using an active avoidance learning paradigm, it was reported that intra-BLA scopolamine administered after a single 30-trial session, had no effect on retention when tested 24 h later. When administered after a weaker 10-trial session, however, scopolamine increased task retention [71]. These results contrast with those from another study by the same group showing that the learning of an approach behavior (socially transmitted food preference) was abolished by pre-training intra-BLA scopolamine infusion [77]. Taken together these findings also support a view in which fear learning is more “treatment resistant” compared to reward learning.
Although we have argued that involvement of BLA mAChRs in aversive memory formation is rather nuanced, recent data also shows that during the acquisition phase of aversive learning, cholinergic neurotransmission through ionotropic nicotinic receptors seems required, sometimes in combination with muscarinic receptors. One study, for example, examined the participation of BLA ACh on fear conditioning suggests the involvement of both muscarinic and nicotinic receptors in associative fear memory formation. Activation of BLA-projecting NBM axons during fear conditioning enhanced fear memory retention and delayed extinction, while inhibition of this pathway diminished fear learning and facilitated extinction; this effect was mimicked by intra-BLA infusions of muscarinic and nicotinic antagonists, but not a muscarinic antagonist alone [59]. More recently, the acquisition of a fear memory that was substantially weakened by the silencing of BLA-projecting NBM cholinergic fibers, was shown to involve nicotinic receptors as systemic administration of mecamylamine, a nicotinic receptor antagonist, mimicked the weakening effect of fear memory formation.
The advent of fluorescent ACh sensors suitable for in vivo studies has reshaped our understanding of cholinergic function in the brain [92, 93]. To this regards, recent research shows that ACh release in the BLA increases in a precisely time-locked fashion in response to salient stimuli and is involved in the learning of cue-reward contingency [78]. The swift burst of ACh release reported in this study contrasts with earlier models based on microdialysis studies with concomitant acetylcholinesterase inhibitors that proposed that the recruiting of learning-promoting mAChRs at target regions must occur slowly, through sustained ACh release over the course of minutes. However, another recent study, also using fiber photometry to study ACh release dynamics in vivo, found stress-induced alterations of ACh levels in the hippocampus over seconds to minute time range [93], consistent with microanalysis studies [94, 95]. This suggests that the dynamics of cholinergic flow may be region and task dependent.
Mounting evidence indicates a crucial role of BLA cholinergic signaling in reward learning. Early psychopharmacological studies using drugs of abuse as rewards consistently reported an involvement of amygdalar mAChRs activity in approach learning. For example, in a rat model of relapse to cocaine-seeking behavior, rats infused with a high dose of scopolamine (72 µg) in the BLA prior to conditioning made fewer cocaine-rewarded lever-presses than those receiving control infusions [81]. Also, in a conditioned place preference task using amphetamine or morphine as a positive reinforcer, post-training infusions of scopolamine or atropine into BLA blocked the consolidation [95, 96].
Similarly, blocking mAChRs with bilateral intra-BLA infusions of scopolamine impaired the learning of approach behavior in an eight-arm radial maze version of the conditioned place preference task in which food was used as a reward these rats utterly failed to show a preference for the reward-paired arm on test day, contrary to the vehicle-treated animals [97], and the extinction of contextual fear conditioning in which the absence of a negative reinforcer is perceived as a reward [30] was shown to be enhanced by intra-BLA infusion of the muscarinic agonist oxotremorine [76]. More recently it was reported that stimulation of cholinergic projections to the BLA promoted context-specific approach behavior, and that under specific conditions, this stimulation appeared to cause a pleasant experience [54]. Given that NBM neurons projecting to the BLA were also activated by a natural reward (sucrose) but not a naturally aversive stimulus (i.e., a foot shock) [54], it appears that activity of these BLA neurons bestow a positive valence to sweet tastes [98].
The sense of taste is inherently emotional [99] and the palatability or hedonic value of an edible can be modified by associative learning, interoceptive experience and/or familiarity [100, 101, 102], the latter having been suggested to be by itself rewarding [103]. In rodents, taste familiarization or incidental taste learning can be studied in the lab with the task of attenuation of neophobia (AN), which some authors argue to involve the learning of an approach behavior in addition to habituation [104]. In AN, the animal initially reluctant to consume an unknow taste, increases consumption upon subsequent exposures, identifying it as familiar and safe [101, 105]. Given that AN in freely drinking rats is dependent on BLA activity [106, 107], and also that cholinergic signaling in this region may favor approach learning [54], we recently formulated the hypothesis that mAChRs in this region should be required for this process. To test this hypothesis, we first administered scopolamine in the BLA 20 minutes before a novel taste exposure, showing that doing so does not affect taste preference (i.e., the neophobic response). While this finding isn’t in complete agreement with earlier observations reporting that stimulation of NBM cholinergic projections to the BLA per se has a rewarding effect [54]. This seeming contradiction makes sense when considered in the context of further results showing that scopolamine infused immediately after the 10-min exposure to novel taste caused a strong avoidance to the taste in a test done 2 days later [82].
Additional data further suggested that this effect was not dependent on taste novelty, did not generalize to another taste and is dependent on Gq protein-coupled (i.e., M1-type) mAChRs [82]. These findings contrast with the classic view of mAChRs being mainly involved in memory consolidation [65, 108] and scopolamine representing an amnesia inducing drug as a scopolamine-induced aversive memory to the taste was observed. Given that many studies have reported a redundancy in the BLA mechanisms involved in aversive learning and that muscarinic receptor inhibition alone in this region often spares various forms of fear conditioning [42, 75], the scopolamine-induced avoidance learning observed by our group and others [71, 82] may suggest that the assignment of an negative valence to a given sensory experience may be the “default” pathway and that sustained muscarinic activity may be required to “switch” to a positive valence.
Additional findings strongly point to a role for BLA cholinergic signaling in positively reinforced learning. Both liquid and solid food reward correlate with increased acetylcholine release in the BLA [78, 80, 109] and cholinergic stimulation in this region occludes feeding behavior [80]. Importantly also, ACh efflux in the BLA is also induced by predictors of rewards and is required for the learning of cue-reward contingency, in a muscarinic receptor- dependent fashion [61, 78]. Interestingly, feeding-induced cholinergic release in the BLA appears to be restricted to the posterior region [80], which has previously been implicated in approach behavior [22] and reward learning [30]. Further studies should determine whether mAChRs activation in this region could be causally linked to approach learning.
In the present text, we reviewed recent evidence on how cholinergic signaling, particularly through its action on mAChRs, promotes neuroplasticity mechanisms in BLA circuits that enable the storage of emotional memories. We first briefly discussed BLA circuits and their involvement in valence processing, then we provided a brief overview of the cellular and molecular mechanisms involved in ACh-induced plasticity events in the BLA. We finally linked this information to loss and gain of function studies on the role of muscarinic signaling in emotional learning, proposing that instead of promoting memory consolidation in a general way, cholinergic signaling in the BLA may contribute to modulate the saliency and/or valence assigned to a given experience along the hedonic spectrum. In addition to improving our understanding of how emotional memories are formed in the brain in both health and disease, it is reasonable to expect that our increasing knowledge in this field of study will help design more promising treatments for neuropsychiatric conditions.
Indeed, new pharmacological targets that focus on selective mAChRs in the BLA could modulate emotional memory consolidation or retrieval in a way that reduces maladaptive behaviors such as excessive or generalized fear responses and/or impaired reward processing seen in some neuropsychiatric conditions related to emotion processing, from post-traumatic stress disorder to binge eating and dysthymia. mAChRs are a promising target for new drug development, not least given that the expression of its distinct subtypes varies widely across brain regions [110]. In this regard, a recently approved combination drug for the treatment of schizophrenia that includes a brain-acting muscarinic agonist represents the first alternative to D2 antagonist in 70 years [111], with the added benefit of also improving negative symptoms, characterized by social withdrawal and emotional blunting [110, 111, 112]. The findings reviewed in the present text further support the rationale behind testing new muscarinic-acting compounds, perhaps helping to improve the negative emotional bias that characterizes most neuropsychiatric conditions [2].
At a more fundamental level, another crucial challenge will be to determine how distinct neuromodulator systems interact at BLA synapses to produce long-term changes in excitability in both principal and interneuron population. To this effect, some mechanisms such as convergent activation of Gq-protein signaling pathways in perisomatic CCK and PV-positive basket cells by different neuromodulator systems have been described [9]. With the improved spatiotemporal resolution permitted by newer technologies, future studies should unveil further valuable information on how different neuromodulatory systems in the BLA modulate the activity of the distinct subpopulations of BLA neurons, a critical step in our understanding of how emotional valence is assigned and maintained.
Conceptualization: JPM, VMTG, GRR, DBK; Funding Acquisition (biorender): JPM; Writing — original draft: VMTG, JPM, ERN; Supervision: JPM; Validation: JPM; Visualization: ERN, VMTG, JPM; Writing — review & editing: JPM, DBK, GRR. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This work benefitted from grants IN227123 and PA009-24 from the Dirección General de Asuntos del Personal Académico, UNAM.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.