








 , Liang-Xiao Ma 1,2,*
, Liang-Xiao Ma 1,2,* , Yu-Xin Zhuang 1, Jing-Si Wen 1, Ling-Hui Ma 1, Jing-Yun Xiu 1, Meng-Yu Chen 1
, Yu-Xin Zhuang 1, Jing-Si Wen 1, Ling-Hui Ma 1, Jing-Yun Xiu 1, Meng-Yu Chen 11 School of Acupuncture-Moxibustion and Tuina, Beijing University of Chinese Medicine, 100029 Beijing, China
2 The Key Unit of State Administration of Traditional Chinese Medicine, Evaluation of Characteristic Acupuncture Therapy, 100029 Beijing, China
Abstract
Post-stroke spastic hemiplegia (PSSH) frequently leads to severe motor dysfunction, with its primary pathology being spinal hyperexcitability arising from attenuated descending inhibition. We previously reported that acupuncture alleviated spastic hypertonia induced by middle cerebral artery occlusion (MCAO) via upregulating potassium-chloride cotransporter 2 (KCC2) expression. Cumulative evidence has indicated that N-methyl-D-aspartate receptor (NMDAR) can be a pivotal determinant of spinal excitability via modulating KCC2-mediated neuronal chloride homeostasis. The present study investigated whether acupuncture exerts its therapeutic effects through modulation of NMDAR-mediated activation of protein phosphatase 1 (PP1)/Calpain1-KCC2 pathway.
Multiple functional assessments, in vivo electrophysiological test, 2,3,5-triphenyl tetrazolium chloride (TTC) staining, immunofluorescence, quantitative real-time PCR (RT-qPCR), and Western blot were used.
In the male MCAO rat model, assessments using the neurological-function score, muscle-tone scale, and footprint analysis demonstrated that acupuncture significantly attenuated spasticity and improved motor performance. H-reflex recordings and immediate early gene c-Fos (c-Fos) immunofluorescence indicated that acupuncture reduced hyperexcitability in spinal ventral horn. These observed effects of acupuncture were associated with its downregulation of N-methyl-D-aspartate receptor 1 (NMDAR1) expression and restoration of both the expression and function of KCC2 in spinal cord. Pharmacological interventions using NMDAR agonist and antagonist demonstrated that acupuncture upregulated KCC2 by inhibiting NMDAR-mediated activation of PP1 and Calpain1.
Acupuncture modulated the NMDAR-PP1/Calpain1-KCC2 pathway in the spinal cord to suppress neuronal hyperexcitability, thereby relieving spasticity and promoting motor function in rats with PSSH.
Keywords
- stroke
- spastic hemiplegia
- NMDA receptor
- KCC2 cotransporter
- acupuncture
As a highly prevalent complication in stroke survivors, post-stroke spastic hemiplegia (PSSH) is distinguished by increased muscle tone and abnormal movement patterns dominated by spasticity, which has a profound effect on patients’ quality of life and places a considerable strain on healthcare systems [1, 2, 3]. After stroke, the obstruction of cerebral blood vessels leads to ischemic damage in the affected brain tissue, causing an impaired transmission of voluntary motor commands and inhibitory signals. It leads to hyperexcitability in the spinal ventral horn, causing involuntary muscle contractions and spastic hypertonia, which ultimately presents as spastic hemiplegia [4, 5]. Neuronal-excitability regulation is a complex process that involves the coordinated action of various neurotransmitters, receptors, and intracellular signaling pathways.
N-methyl-D-aspartate receptor (NMDAR) is an ionotropic glutamate receptor widely
distributed on the motor neurons in spinal ventral horn. It has high Ca2+permeability and slow activation and deactivation kinetics [6]. This enables NMDAR to serve as a key determinant in regulating neuronal excitability and
mediating excitotoxicity [7, 8]. Potassium-chloride cotransporter 2 (KCC2) is a
key transport protein located on the neuronal membrane. It extrudes intracellular
Cl–, thereby maintaining a low intracellular Cl– concentration and providing the
necessary conditions for
Commonly used drugs, such as baclofen, alleviate increased muscle tone by activating GABAB receptors to inhibit neuronal excitability [13]. Nevertheless, these drugs often show limited efficacy, with short-term effects and potential side effects. Acupuncture, as a non-pharmacological therapeutic approach, has garnered widespread attention in recent years for its enduring efficacy, simplicity, and cost-effectiveness in treating PSSH [14, 15, 16]. Previous research has observed that, in rat models of spinal cord injury resulting in motor dysfunction, NMDAR in the spinal cord was elevated, and is effectively reversed by electroacupuncture [17]. Additionally, in rats with middle cerebral artery occlusion (MCAO) exhibiting hypertonia, the expression levels of KCC2 and GABAA in the spinal cord were reduced; acupuncture restored their expression, alleviated spasticity, and promoted motor-function recovery [18]. However, how acupuncture modulates neuronal excitability through the coordinated action of NMDAR and KCC2 remains insufficiently understood. The present study investigated whether acupuncture exerts its therapeutic effects on PSSH through the regulation of the NMDAR-PP1/Calpain1-KCC2 pathway.
Male Sprague-Dawley rats (8 weeks), ranging in weight from 250 to 280 g, were
used for the study. All rats were kept in cages of 4 under SPF conditions at room
temperature (23
The experimental timeline and corresponding procedures are shown in Fig. 1A and Fig. 5A. To evaluate the effects of acupuncture on rats with PSSH, four groups were established: Sham-operated group (Sham), MCAO induced PSSH model group (Model), acupuncture at acupoint group (Acup), and acupuncture at control point (non-acupoint) group (CP). To investigate the role of NMDAR, the NMDAR agonist NMDA and antagonist D-(-)-2-Amino-5-phosphonopentanoic acid (D-AP5) were administered. Five groups were established: Model, Model + vehicle, Acup + vehicle, Model + D-AP5, and Acup + NMDA. All rats were randomly assigned to these groups. Excluding the 10 rats that underwent sham surgery, 112 rats were subjected to MCAO procedure for PSSH-model induction, of which 80 were successfully modeled and subsequently used in the experiments (Supplementary Table 1). The sample size was chosen according to previous research and our preliminary data, ensuring adequate statistical power to identify meaningful differences among groups.
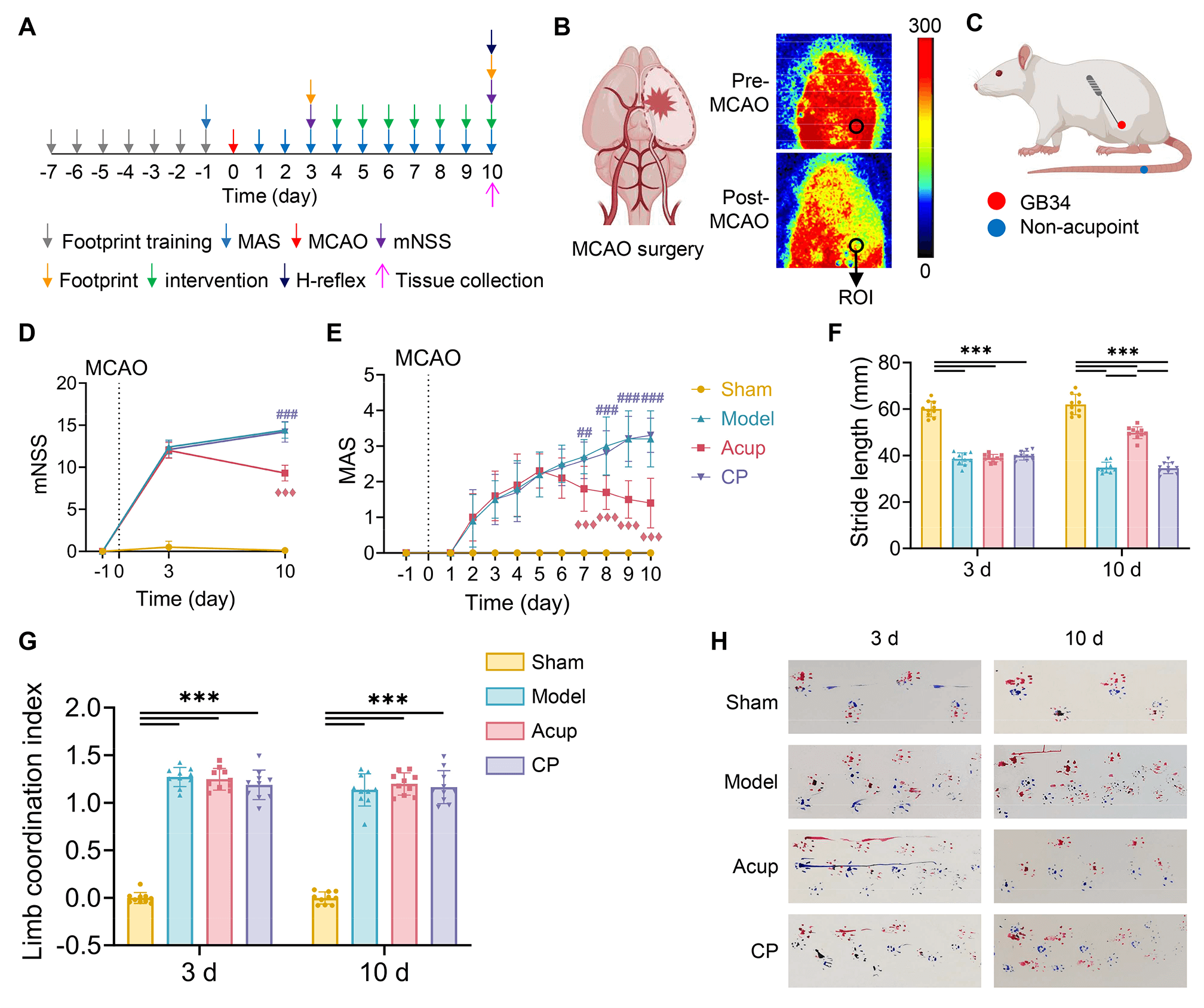 Fig. 1.
Fig. 1.
Acupuncture alleviates spasticity and enhances motor function in
MCAO rats with PSSH. (A) Timeline of experimental procedures. (B) Representative
laser speckle images (LSI) for monitoring cortical blood flow (CBF) after MCAO
surgery. (C) Acupuncture site location. (D) Neurological deficits assessed by the
mNSS. (E) Muscle tone assessed by the MAS. (F) Stride length of rats at 3 days
and 10 days. (G) Limb coordination index of rats at 3 days and 10 days. (H)
Footprints of rats in each group, with blue representing the forelimbs and red
representing the hind limbs. N = 10 per group. Data are presented as mean
The permanent MCAO procedure was performed to establish a PSSH model, following
the previously described method with slight modifications [19, 20]. After
anesthetization with isoflurane (3% in oxygen; flow rate, 1 L/min; cat. no. R510-22-10; RWD Life Science, Shenzhen, Guangdong, China), a midline skin
incision was performed on the ventral neck to expose the right common carotid
artery. A silicone-coated nylon monofilament with a diameter of 0.36 mm (Beijing
Cinontech Co., Ltd.; Beijing, China) was gently advanced from the right external
carotid artery to the internal carotid artery and further to the origin of the
middle cerebral artery (MCA). The filament was left in place for the remainder of
the experiment. A temperature-controlled heated pad was applied during the
ischemic period to maintain the body temperature constantly at
37
MCAO rats were included in the study if they exhibited a regional cerebral blood flow (CBF) reduction of greater than 70% of baseline levels, and a Zea-Longa score above 1. All behavioral evaluations were performed by an independent investigator who was blind to the interventions of the study.
CBF changes were assessed using a laser speckle imaging system (PeriCam PSI NR; Perimed AB; Stockholm, Sweden). Measurements were taken 5 min before and 5 min after MCAO surgery to determine relative perfusion in the MCA territory. After induction of anesthesia, a midline scalp incision was made to expose the skull. Circular regions of interest (ROIs) with a diameter of 2 mm were placed 3 mm lateral and 2 mm posterior to the bregma on the right cortical surface (Fig. 1B). CBF values were calculated as percentages relative to pre-occlusion baseline levels according to acquired microcirculation images.
Acupuncture treatment was administered once daily to rats in the Acup and CP
groups from postoperative day 4 to day 10. Sterile, disposable acupuncture
needles (0.25
NMDA (40 mM, cat. no. 0114; Tocris; Minneapolis, MN, USA) and D-AP5 (15 mM, cat. no. 0106; Tocris) were used for NMDAR modulation. Both compounds were freshly dissolved in sterile distilled water and administered via intrathecal injection at a volume of 30 µL once daily from postoperative day 4 to day 10.
The modified Neurological Severity Score (mNSS) was used to assess overall neurological function, including motor ability, sensory responses, reflexes, and balance [21]. In this scoring system, one point is assigned for the inability to perform a task or for the absence of a reflex. Neurological function was scored on a scale from 0 to 18, where 0 indicated normal function and 18 represented the maximum deficit.
The Modified Ashworth Scale (MAS) was used to assess muscle tone in the affected hind limb [22]. The scale ranges from 0 to 4 (normal muscle tone with free limb movement, 0; complete rigidity of limbs, 4) [18]. A higher score indicates higher muscle tone and greater limitation of joint movements. In this study, the maximum MAS score was set to 5, with scores of 1+ converted to 2 for analysis purposes [23].
Walking-track-footprint analysis was described previously [24]. Rats had their forepaws dipped in nontoxic blue dye and hind paws in red dye. Each rat was placed at the start of an 80-cm-long, 10-cm-wide wooden track leading to a darkened goal box containing food. Prior to surgery, each rat underwent one training session per day to ensure consistent gait. During each testing session, the rat traversed the track, leaving footprints on white paper. Trials were repeated if the rat hesitated, stopped, or failed to complete the walk smoothly. For each rat, 6 consecutive hind-paw prints were selected for analysis. Stride length was measured as the distance between two successive placements of the same hind paw. The coordination index was defined as the distance between the centers of the ipsilateral forepaw and hind-paw prints, representing interlimb coordination. The mean value of each parameter was calculated for statistical analysis.
A Bio-Signal Acquisition and Processing System (BL-420N, Chengdu Taimeng Technology Co., Ltd.; Chengdu, Sichuan, China) was used to perform Hoffmann-reflex (H-reflex) recording and analysis. After anesthetization with 3% isoflurane in 97% oxygen, the left sciatic nerve was exposed by blunt dissection and secured on a bipolar hook electrode. To prevent nerve desiccation during the procedure, oleovitamin was applied to the exposed nerve. Electromyographic (EMG) signals were collected using stainless-steel needle electrodes inserted into the interosseous muscles of the left hind paw, and a ground electrode was placed subcutaneously in the tail.
The H-reflex was elicited by applying single biphasic pulses (100 µs width) to the left sciatic nerve through an isolated pulse stimulator. Both M-waves and H-waves were recorded at varying stimulus intensities to determine the thresholds of motor (MT) and H-reflex. To assess frequency-dependent depression (FDD), stimuli of identical intensity (eliciting Hmax) were delivered at frequencies 0.3 Hz, 5 Hz, and 10 Hz [25]. Subsequently, the variations in the amplitude of the H-reflex at 5 Hz and 10 Hz were expressed as a percentage of the response at 0.3 Hz. A repeated 0.3 Hz stimulation was applied at the end of the recording to confirm that the M-wave remained within 95% of baseline; data were excluded if this criterion was not met. All H-reflex and M-wave measurements, including FDD, were obtained 15 min after the final treatment.
The rats were euthanized by cervical dislocation under deep anesthesia induced by intraperitoneal injection of pentobarbital sodium (100 mg/kg body weight, Sigma-Aldrich, St. Louis, MO, USA; Cat. No.: P3761) after the final interventions, and the whole brains were rapidly removed and cooled at –20 °C for approximately 30 min to facilitate sectioning. Each brain was sliced into 2-mm coronal sections and incubated in 0.3% TTC solution (cat. no. SL7140; Coolaber; Beijing, China) at 37 °C in the dark for 20–30 min. The stained slices were then stored in 4% paraformaldehyde (cat. no. SL1830; Coolaber) overnight. Digital images of the brain sections were captured, and infarct areas were analyzed and detected as previously described utilizing ImageJ software (Fiji; National Institutes of Health; Bethesda, MD, USA) [18].
The embedded lumbar-spinal-cord-enlargement tissue was sectioned into 10 µm slices using a motorized cryostat. Donkey serum was used to block first, and then the following primary antibodies were used: anti-c-Fos (1:500, cat. no. 2250T; Cell Signaling Technology; Danvers, MA, USA), anti-NMDAR1 (1:500, cat. no. ab109182; Abcam; Cambridge, Cambs, UK), and anti-KCC2 (1:500, cat. no. ab259969; Abcam) were incubated overnight at 4 °C. The next day, sections were incubated for 2 h at 37 °C in the dark with the fluorescence antibodies (1:500, cat. no. RGAR002, cat. no. RGAR004; Proteintech; Rosemont, IL, USA). After washing, the sections were mounted with anti-fade mounting medium containing 4′,6-diamidino-2-phenylindole (DAPI) (cat. no. S2110; Solarbio; Beijing, China) and then photographed.
Low-magnification images (4
Total RNA was isolated from the lumbar spinal ventral horn of each rat according
to the manufacturer’s instructions (cat. no. DP451; Tiangen; Beijing, China).
RT-qPCR was performed by mixing cDNA templates with gene-specific primers and
PowerUp SYBR Green Master Mix (cat. no. A25742; Thermo Fisher Scientific;
Waltham, MA, USA), followed by amplification using a StepOne Plus Real-Time PCR
Detection System (CFX Connect; Applied Biosystems; Foster City, CA, USA). The thermal cycling
conditions were as follows: initial denaturation at 95 °C for 3 min,
followed by 39 cycles of 95 °C for 10 s and 59 °C for 60 s.
Proteins were extracted from the lumbar spinal ventral horn and concentrations
were determined with a BCA kit (cat. no. AR0146; Boster; Wuhan, Hubei, China).
After SDS-PAGE (8 and 10%) and PVDF membrane transfer (cat. no. IPVH00010;
Millipore; Burlington, MA, USA), membranes were blocked and incubated with the
following primary antibodies: anti-NMDAR1 (1:6000, cat. no. ab109182; Abcam),
anti-KCC2 (1:10,000, cat. no. ab259969; Abcam), anti-phospho-KCC2 (Ser940)
(1:3000, cat. no. CAY-29291-100; Cayman Chemical; Ann Arbor, MI, USA),
anti-PP1C
The data analyses of this study were conducted using SPSS Statistics 25.0 (IBM;
Armonk, NY, USA) and GraphPad Prism 10 (GraphPad Software; San Diego, CA, USA).
One-way ANOVA, two-way ANOVA, and two-way repeated measures ANOVA were performed
as appropriate to assess the significance. Data are presented as mean
Neurobehavioral tests were conducted to assess the therapeutic effects of
acupuncture on PSSH. To evaluate the impact of acupuncture on neurological
deficits, the mNSS was assessed on day 3 and day 10 post-MCAO. All rats had an
mNSS score of 0 prior to surgery. On day 3 after surgery, compared to
sham-operated rats, MCAO surgery induced neurological impairments manifested by
significantly elevated mNSS scores (all p
To assess the role of acupuncture on spasticity, the MAS scores of the affected
hind limb were measured daily for 10 consecutive days after surgery. All rats had
a MAS score of 0 before surgery. Starting from postoperative day 2, the MAS
scores of rats in the three MCAO groups markedly increased, but did not in the
sham rats (p
Building on the results of subjective behavioral assessments, we subsequently
performed footprint analysis to objectively evaluate motor function in rats (Fig. 1H). On postoperative day 3, MCAO rats in the three surgery groups showed shorter
stride lengths of the hind limb than did the sham-operated rats (all p
We assessed the impact of acupuncture on spinal hyperexcitability and ischemic
brain damage. The H-reflex is widely used to assess
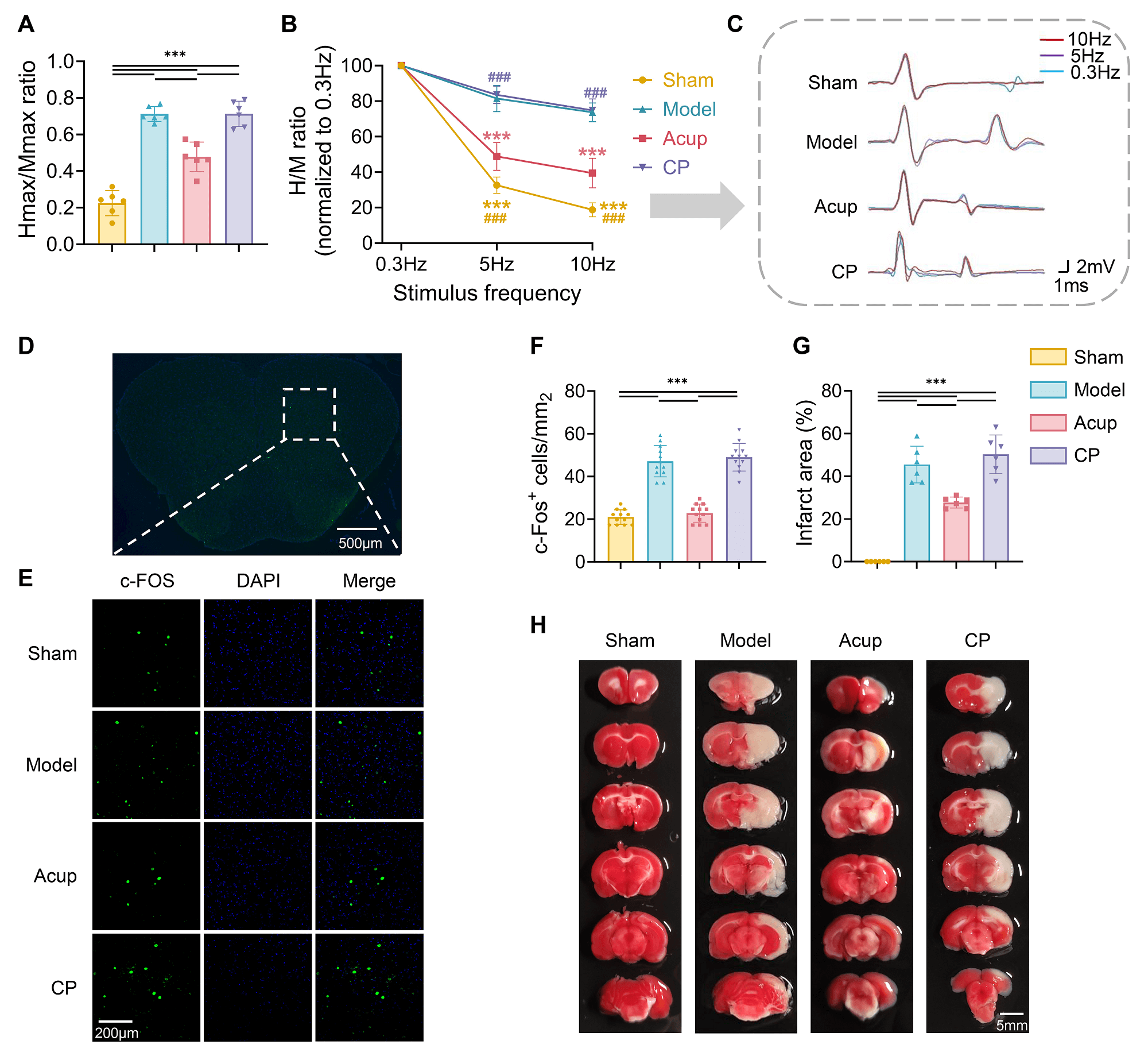 Fig. 2.
Fig. 2.
Acupuncture mitigates MCAO-induced spinal hyperexcitability and
ischemic brain injury. (A) Hmax/Mmax ratio was measured to assess spinal
excitability (N = 6). (B) FDD of the H-reflex (N = 6). (C) Representative
H-reflex traces recorded at 0.3 Hz (blue), 5 Hz (purple), and 10 Hz (red) to show
changes in the H-reflex (N = 6). (D) Distribution of c-Fos in the lumbar spinal
ventral horn neurons. Scale bar, 500 µm. (E) Representative images of c-Fos
expression in the lumbar spinal ventral horn of each group. Scale bar, 200
µm. (F) Analysis of c-Fos-positive cell density (cells per mm2) (12
spinal slices from 4 rats per group). (G,H) TTC staining and quantitative
analysis of infarction areas in the brain (N = 6). Scale bar, 5 mm. Data are presented as mean
Subsequently, FDD was measured to evaluate spinal reflex activity in MCAO rats.
Impaired FDD is widely recognized as a hallmark of spinal hyperreflexia [28]. We
evoked H-reflexes at stimulation frequencies of 0.3 Hz, 5 Hz, and 10 Hz to
evaluate the extent of FDD. The results showed a frequency-dependent decline in
H-reflex amplitude across all groups, indicating the presence of FDD to varying
degrees (Fig. 2B,C). Significant effects were observed for stimulation frequency
(F[2,60] = 588.7, p
As a cellular marker of neuronal activation, c-Fos expression in
spinal-ventral-horn neurons was analyzed to further verify the modulatory effect
of acupuncture on neuronal excitability (Fig. 2D,E). The c-Fos-positive neuron
count was higher in model rats than in sham-operated rats (p
TTC staining was used to evaluate ischemic brain injury. As depicted in Fig. 2G,
no obvious infarct area was observed in the Sham group, whereas all three surgery
groups exhibited clear ischemic regions, confirming successful induction of
cerebral infarction by MCAO. After intervention, the infarct area in the Acup
group was smaller than that in the Model group (p
NMDAR enhances neuronal excitability primarily by mediating Ca2+ influx,
whereas KCC2 maintains low intracellular Cl– levels to support GABAergic
inhibition. Both play critical roles in regulating neuronal excitability. We
investigated whether acupuncture treatment alleviated PSSH by modulating NMDAR
and KCC2 expression in the spinal ventral horn. NMDAR1 is involved in Ca2+
influx and regulating neuronal excitability [29]. We assessed NMDAR1 expression
in the spinal ventral horn. The results showed that NMDAR1 mRNA and protein
expression levels were higher in the Model group than in the Sham group (all
p
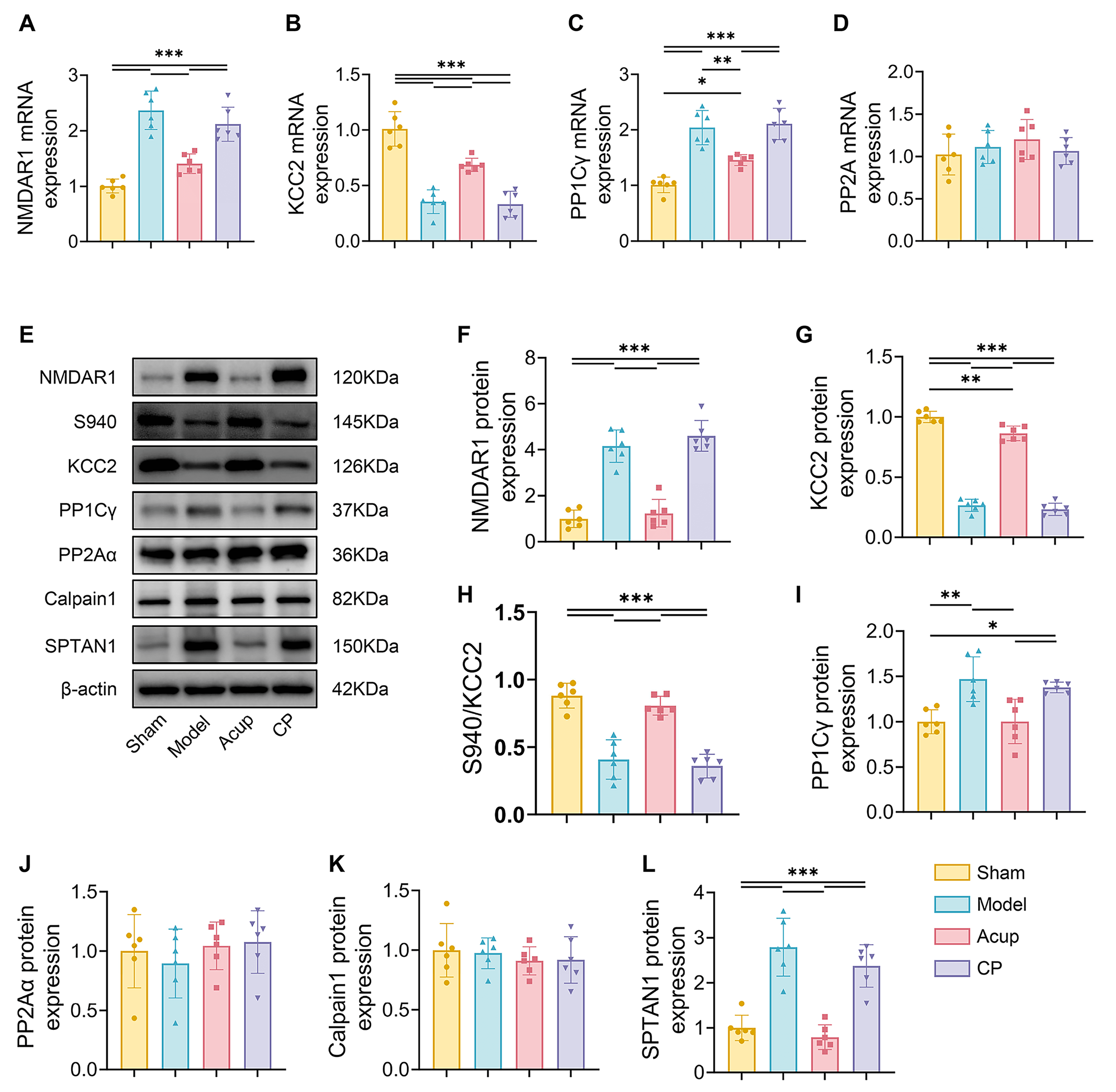 Fig. 3.
Fig. 3.
Acupuncture downregulates the expression of NMDAR1 and restores
KCC2 expression and activation in spinal cord of MCAO rats with PSSH. (A–D): The
mRNA expressions of NMDAR1 (A), KCC2 (B), PP1C
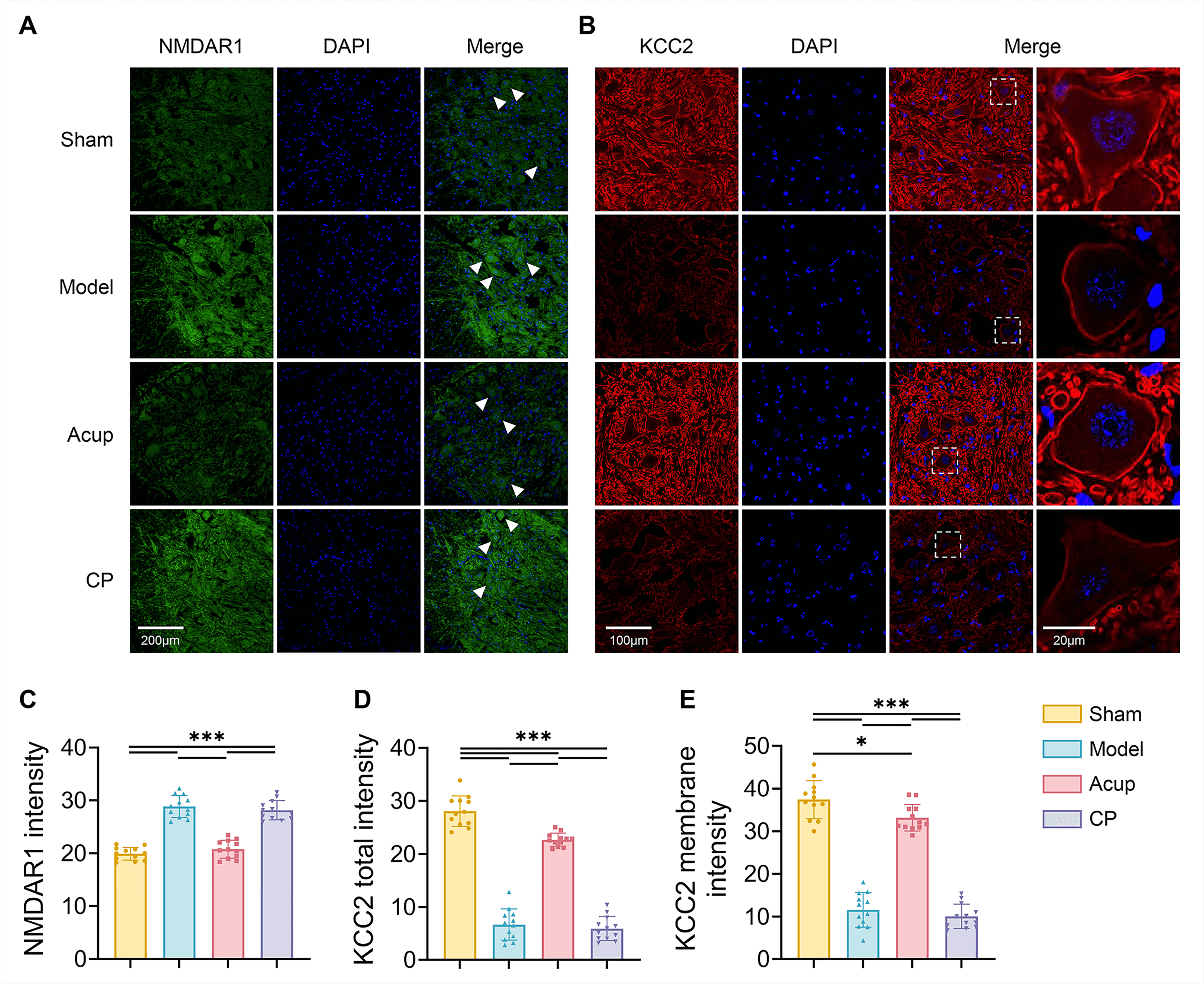 Fig. 4.
Fig. 4.
Acupuncture downregulates the expression of NMDAR1 in spinal
ventral horn and upregulates the total and membrane expression of KCC2 in spinal
motor neuros. (A,B) Representative immunofluorescence images indicating NMDAR1
(A, green), KCC2 (B, red) co-localization with all nuclei of cells stained by
DAPI (blue) in the lumbar spinal ventral horn. Scale bar, 200 µm, 100
µm, 20 µm. The white arrows indicate representative individual neurons, and the white boxes indicate regions of interest, which are enlarged and shown adjacent to the original image. (C–E) Mean fluorescence intensity of NMDAR1 (C), KCC2
total intensity (D) and KCC2 membrane intensity (E) were expressed as the ratio
of integrated density to the observation fields. 12 spinal slices from 4 rats per
group. Data are presented as mean
Then we evaluated KCC2 expression in the spinal ventral horn. The results
consistently indicated that both KCC2 mRNA levels and total protein expression
were lower in the Model group than in the Sham group (all p
We further explored the potential mechanisms by which NMDAR suppresses KCC2
expression and activity. Phosphorylation at the Ser940 site is primarily
dephosphorylated by PP1 and protein phosphatase 2A (PP2A). We assessed the
expression of PP1C
Dephosphorylation of KCC2 at the Ser940 site leads to its inactivation and
internalization, followed by degradation by calpain1 in the cytoplasm. The
activity of calpain1 is highly dependent on NMDAR-mediated Ca2+ influx [32, 33]. Nonerythroid spectrin
NMDAR directly enhances neuronal excitability by mediating Ca2+ influx and
downregulates the expression and activity of KCC2, thereby indirectly weakening
inhibitory regulation. Given that acupuncture has been shown to alleviate PSSH by
modulating KCC2, we further investigated whether this effect was associated with
its regulation of NMDAR1. To this end, the NMDAR agonist NMDA and antagonist
D-AP5 were administered intrathecally. We evaluated mNSS scores, MAS scores, and
H-reflex parameters in each group. After treatment, the Acup + NMDA group showed
significantly higher mNSS and MAS scores, a higher Hmax/Mmax ratio, and lower FDD
suppression than did the Acup + Vehicle group (all p
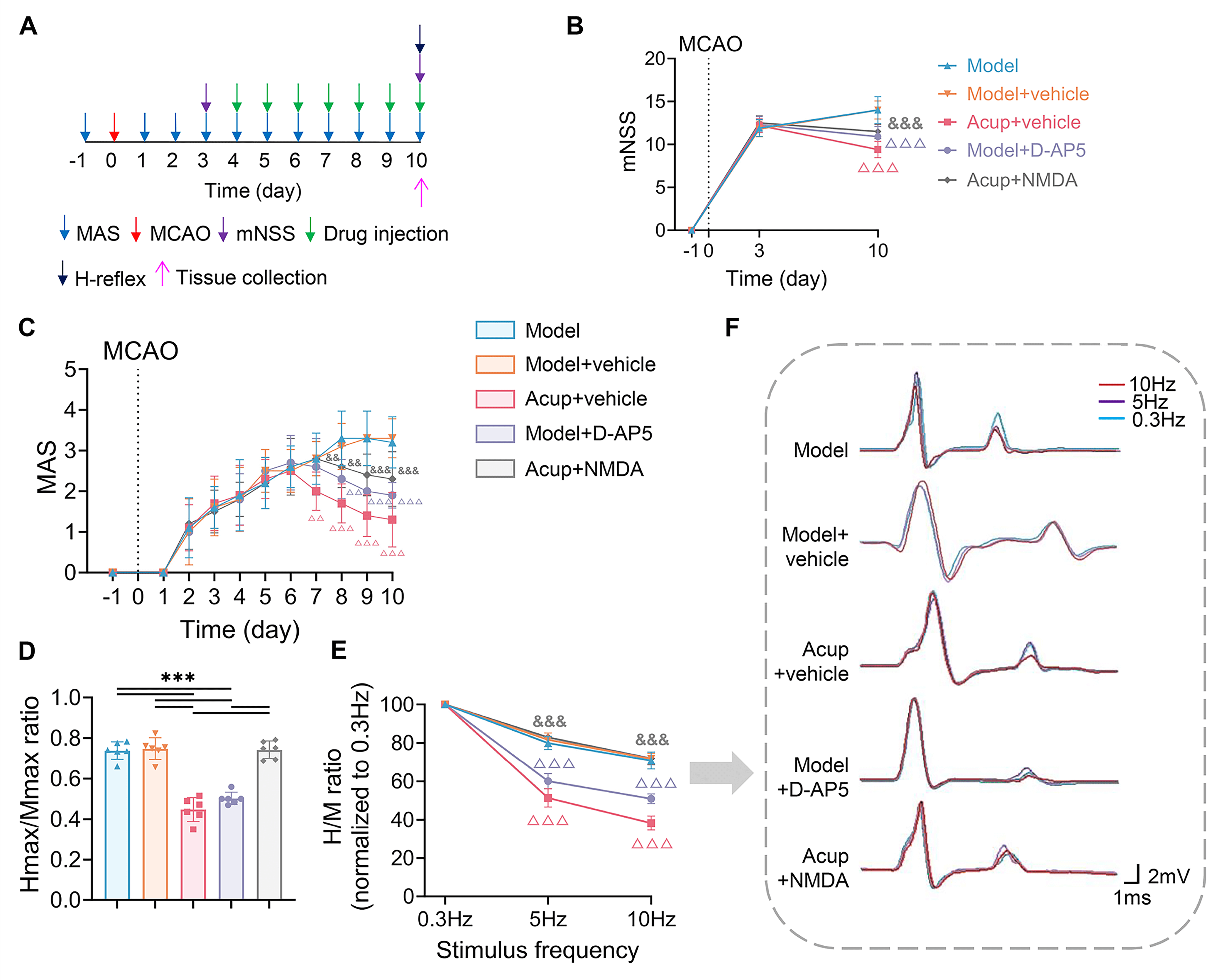 Fig. 5.
Fig. 5.
The impact of NMDAR antagonist and agonist on acupuncture
effects on neurological deficits, spasticity, and H-reflex in MCAO rats with
PSSH. (A) Timeline of experimental procedures. (B) Neurological deficits
assessed by the mNSS (N = 10). (C) Muscle tone assessed by the MAS (N = 10). (D)
Hmax/Mmax ratio was measured to assess spinal excitability (N = 6). (E) FDD of
the H-reflex (N = 6). (F) Representative H-reflex traces recorded at 0.3 Hz
(blue), 5 Hz (purple), and 10 Hz (red) to show changes in the H-reflex. Data are
presented as mean
To verify the effectiveness of the pharmacological intervention in modulating
NMDAR1, we examined NMDAR1 expression in the spinal ventral horn. Results showed
that NMDAR1 mRNA and protein levels were evidently lower in the Model + D-AP5
group than in the Model + Vehicle group (all p
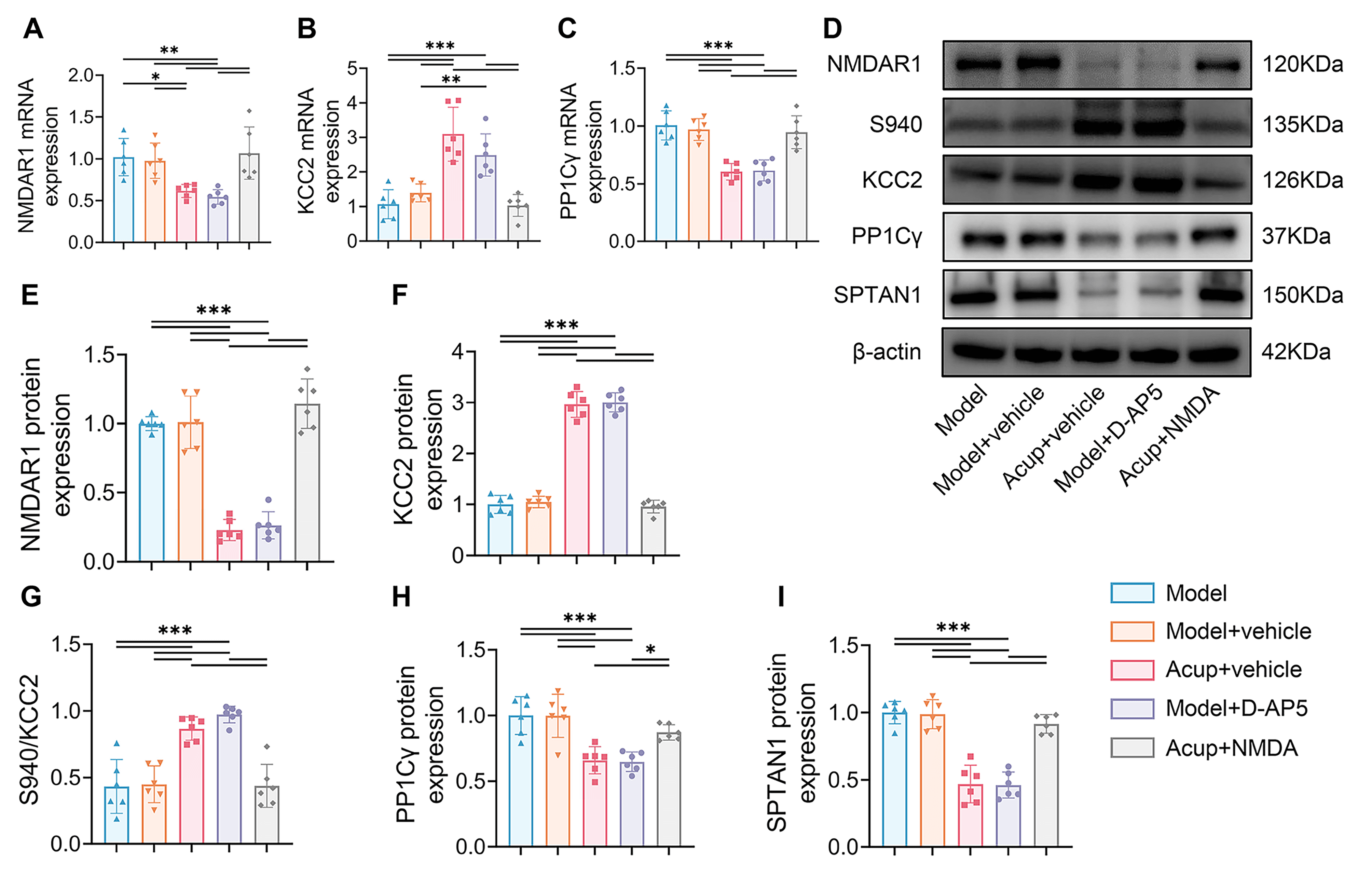 Fig. 6.
Fig. 6.
The effects of NMDAR antagonist and agonist on the
NMDAR-PP1/Calpain1-KCC2 pathway. (A–C) The mRNA expressions of NMDAR1 (A), KCC2
(B) and PP1C
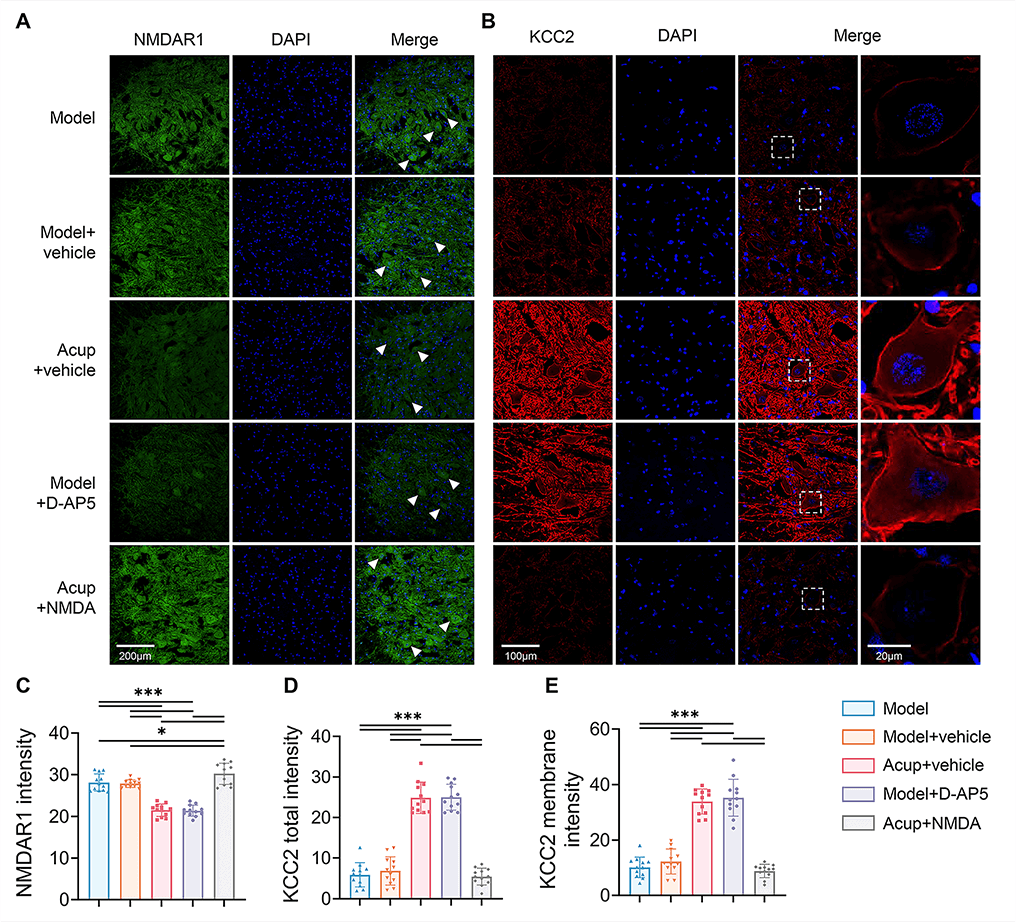 Fig. 7.
Fig. 7.
The effects of NMDAR antagonist and agonist on the expression of
NMDAR1 in spinal ventral horn and the total and membrane expression of KCC2 in
spinal motor neuros. (A,B) Representative immunofluorescence images indicating
NMDAR1 (A, green), KCC2 (B, red) co-localization with all nuclei of cells stained
by DAPI (blue) in the lumbar spinal ventral horn. Scale bar, 200 µm, 100
µm, 20 µm. The white arrows indicate representative individual neurons, and the white boxes indicate regions of interest, which are enlarged and shown adjacent to the original image. (C–E) Mean fluorescence intensity of NMDAR1 (C), KCC2
total intensity (D) and KCC2 membrane intensity (E) were expressed as the ratio
of integrated density to the observation fields. 12 spinal slices from 4 rats per
group. Data are presented as mean
We next determined whether the process by which acupuncture promoted KCC2
activity by inhibiting PP1-mediated Ser940 dephosphorylation was associated with
the regulation of NMDAR. The Acup + NMDA group exhibited higher PP1C
We further clarified whether the process by which acupuncture inhibited calpain1
to reduce KCC2 degradation was associated with the regulation of NMDAR1 by
acupuncture. SPTAN1 expression was substantially higher in the Acup + NMDA group
than in the Acup + Vehicle group (p
This study identified a potential mechanism underlying the therapeutic effects of acupuncture treatment on PSSH. Our results demonstrated that acupuncture treatment significantly alleviated spasticity and improved motor function in MCAO rats with PSSH. Acupuncture appears to exert its therapeutic effects by modulating the NMDAR-PP1/Calpain1-KCC2 pathway, which regulates the excitability of spinal ventral-horn neurons. The effects of pharmacological manipulation using the NMDAR agonist NMDA and antagonist D-AP5 further supported the critical involvement of NMDAR in the effects of acupuncture (Fig. 8).
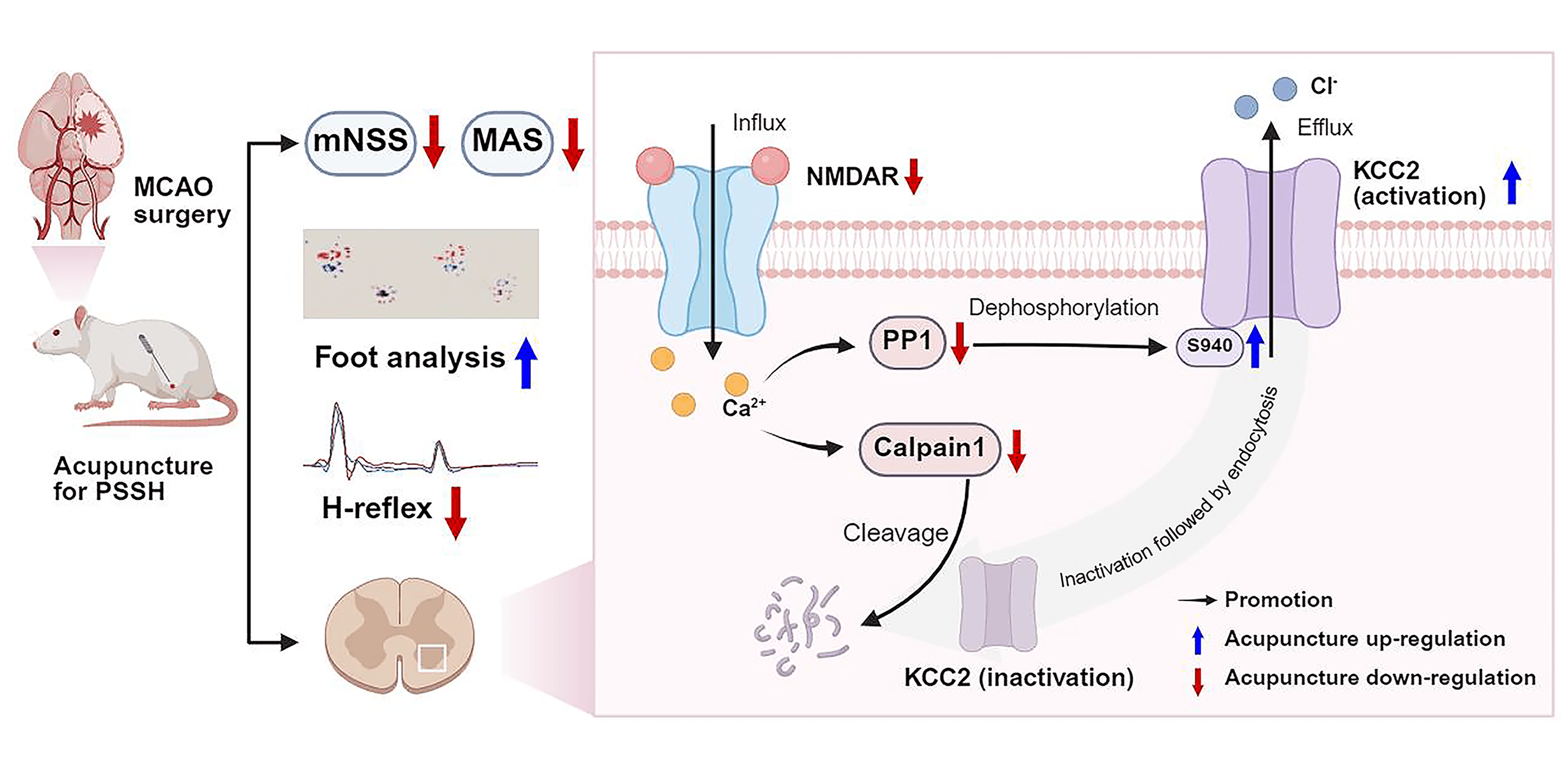 Fig. 8.
Fig. 8.
Acupuncture alleviates PSSH via modulating the NMDAR-PP1/Calpain1-KCC2 pathway in spinal cord (Created with BioRender.com).
PSSH is a common sequela of stroke, with a reported incidence of up to 39.5% [34]. It is characterized by increased muscle tone and spastic movement patterns, which impede motor recovery and reduce quality of life [35]. To replicate the pathological features of PSSH and evaluate the efficacy of acupuncture treatment, we used the well-established MCAO model in rats and selected GB34 as the intervention site. Located anteroinferior to the fibular head, GB34 corresponds segmentally to the lumbar-spinal-cord enlargement that governs lower limb motor control, and its efficacy in treating PSSH has been well documented [18, 36]. We assessed muscle tone in the affected hind limb using the MAS scores and conducted footprint analysis to evaluate motor performance quantitatively. Given that the regulation of movement also depends on sensory input and central integration, we used the mNSS scores to assess the overall degree of neurological impairment. Acupuncture at GB34 effectively reduced spasticity, improved motor function, and facilitated the recovery of overall neurological function in MCAO rats.
Spasticity after stroke is primarily attributed to disrupted supraspinal inhibition of spinal reflex circuits. Specifically, damage to upper motor neurons leads to a loss of descending inhibitory control, resulting in hyperexcitability of spinal stretch reflexes and consequent pathological increases in muscle tone [37]. As the convergence site of motor neurons, spinal ventral-horn hyperexcitability directly contributes to sustained muscle contractions, ultimately driving spasticity and motor impairment [38]. The H-reflex serves as a classical electrophysiological index of spinal excitability. In our study, MCAO rats exhibited a significantly increased Hmax/Mmax ratio and reduced FDD, indicating enhanced excitability of spinal reflexes. These results were consistent with previous findings in both spinal cord injury and MCAO models [18, 25]. Acupuncture intervention effectively reversed these abnormalities. Building on these observations, we further examined the expression of c-Fos in spinal ventral-horn neurons. The classical immediate early gene c-Fos is widely used as a molecular marker of neuronal activation. Elevated c-Fos expression in spasticity models induced by photothrombosis has been reported [39]. Our results revealed a marked increase in the number of c-Fos-positive neurons in the spinal ventral horn of MCAO rats; this number was significantly reduced by acupuncture. This result reinforced our electrophysiological findings, providing further evidence that acupuncture treatment mitigated spinal ventral-horn hyperexcitability.
We focused on the critical role of the NMDAR-PP1/Calpain1-KCC2 pathway in regulating neuronal excitability. NMDAR, an ionotropic glutamate receptor with high Ca2+ permeability, is widely expressed on motor neurons within the spinal ventral horn. Sustained activation of NMDAR can lead to neuronal hyperexcitability and trigger multiple downstream signaling cascades [8]. In addition to motor neurons, the ventral horn also contains GABAergic inhibitory interneurons, which regulate motor neuron activity [40, 41]. KCC2 is a key molecular determinant of GABAergic inhibition, which maintains a low intracellular Cl– concentration required for GABAA receptor-mediated hyperpolarization [9, 10]. Previous research has reported that KCC2 expression in the lumbar enlargement is markedly reduced after stroke, and acupuncture treatment can restore GABAergic inhibition and reduce neuronal excitability by upregulating KCC2 expression. This effect was abolished by the KCC2 antagonist D-Indo-oxy-5-sulfonic acid [18]. We further demonstrated that stroke upregulated NMDAR1 expression and induced a significant reduction in total KCC2 expression, membrane localization, and phosphorylation at Ser940 in the spinal ventral horn. Acupuncture treatment ameliorated these pathological changes. Importantly, the protective effect of acupuncture on KCC2 was impeded by NMDAR, but was restored by D-AP5, suggesting that this effect may involve suppression of NMDAR expression.
To explore how acupuncture might influence KCC2 regulation, we investigated two
key aspects: the activity and protein stability of KCC2. Phosphorylation at the
Ser940 site of KCC2 is essential for its membrane transport function and surface
stability. In rat hippocampal neurons, excessive glutamate-induced activation of
NMDAR has been shown to trigger Ca2+ influx, which subsequently activates
protein phosphatases to dephosphorylate Ser940 [11]. This process impairs KCC2
function and reduces its membrane localization, thereby reversing GABAA
receptor-mediated hyperpolarization and enhancing neuronal excitability. This
process can be blocked by okadaic acid, an inhibitor of PP1 and PP2A. We observed
an upregulation of PP1C
Building upon previous studies on acupuncture for PSSH, the present study systematically investigated how acupuncture treatment alleviates spasticity and promotes motor recovery through modulation of the NMDAR-PP1/Calpain1-KCC2 pathway. Our findings revealed a dual pathological role of NMDAR in promoting neuronal hyperexcitability: by facilitating Ca2+ influx that directly enhances neuronal excitation, and by impairing GABAergic inhibition through suppression of KCC2 function. Further analysis demonstrated that NMDAR downregulated KCC2 function through two pathways: upregulation of PP1 leading to Ser940 dephosphorylation, and activation of calpain1 promoting degradation of KCC2. Acupuncture treatment significantly reduced NMDAR1 expression, which was accompanied by a restoration of KCC2 activity and protein stability. Collectively, these effects converged to reduce the hyperexcitability of spinal ventral-horn neurons and contribute to the therapeutic efficacy of acupuncture in PSSH. Additionally, our findings also preliminarily offered a potential for future targeted interventions, particularly the development of pharmacological agents aimed at negatively modulating NMDAR-PP1/Calpain1 pathway to enhance KCC2 activity and stability for stroke-recovery management.
The intervention timing of acupuncture treatment for PSSH is also a critical issue. In this study, continuous acupuncture intervention was administered from day 4 to day 10 after MCAO, primarily based on observations from previous studies indicating that limb spasticity after MCAO begins to manifest on day 3 post-surgery, with a significant increase in MAS scores starting on day 3 and persisting until day 10 post-surgery [18, 36, 43]. Rats recover more rapidly than do clinical patients, and the period from day 3 to day 10 post-surgery may resemble the spasticity phase observed in clinical stroke patients. Furthermore, given that our primary focus was on observing gross motor function of the lower limbs, our observations were conducted up to day 10 post-surgery. Future studies involving fine motor skills may necessitate longer observation periods.
According to classical acupuncture theory, selecting acupoints and using needling techniques that are appropriate for the specific disease, are key factors in achieving therapeutic efficacy with acupuncture. Although this study was an animal experiment, it demonstrated that when treating lower-limb spastic hemiplegia after stroke, applying a motion-style needling technique (waggle needling) at GB34, which is located on the spastic antagonist muscle, is beneficial for alleviating post-stroke spasticity. Specifically, our findings indicated that future clinical strategies could validate approaches targeting the NMDAR-KCC2 pathway for PSSH.
Despite these insights, there were several limitations. This study primarily focused on the NMDAR-PP1/Calpain1-KCC2 pathway, however, the regulation of KCC2 may vary across different neuronal subtypes, so elucidation may necessitate the use of cell-type-specific markers and in situ detection techniques. Given that different NMDAR subunits can exert distinct effects on neuronal excitability, subunit-specific inhibitors or genetic approaches should be used to delineate the specific contributions. Ca2+ signaling is a key mediator of excitability regulation, but in the present work, we only observed the cumulative effect of Ca2+ influx via c-Fos and calpain1 activation. Future experiments combining Ca2+ imaging for real-time recording of neuronal Ca2+ dynamics will provide a more precise assessment of excitability changes. In addition, the inclusion of inhibitory synaptic markers would help to characterize how acupuncture modulates neural excitability. Acupuncture signals are primarily transmitted via the spinal dorsal horn; they may modulate ventral-horn motor neurons through local interneuronal networks, or ascend to brainstem or cortical centers and influence spinal excitability via descending pathways [44]. Further research combining viral tracing and multi-site electrophysiological recording would help clarify these cross-structural interactions. To avoid the confound of hormonal effects on neuronal activity, the present study used only male rats. Future work will include females in order to explore potential sex-related differences in the regulation of neuronal excitability by acupuncture. Moreover, to further validate acupoint specificity, more appropriate non-acupoint control sites will be considered. Our study used single-acupoint stimulation to provide mechanistic clarity, which differs from clinical acupuncture practice [45, 46]. In addition, we examined only consecutive acupuncture treatments from postoperative day 4 to day 10. Future research could investigate multi-acupoint protocols and varied treatment frequencies or durations to better inform clinical application.
This study systematically elucidated the potential mechanism underlying the therapeutic effects of acupuncture treatment in PSSH. Acupuncture effectively modulated the excitability of spinal ventral-horn neurons to ameliorate spasticity and improve motor function in MCAO rats with PSSH. These effects of acupuncture were associated with its modulation of the NMDAR-PP1/Calpain1-KCC2 pathway. Acupuncture treatment suppressed NMDAR expression and upregulated both the expression and function of KCC2, thereby alleviating spinal hyperexcitability after ischemic stroke.
PSSH, Post-stroke spastic hemiplegia; MCAO, Middle cerebral artery occlusion; KCC2, Potassium-chloride cotransporter 2; NMDAR, N-methyl-d-aspartate receptor; PP1, Protein phosphatase 1; TTC, 2,3,5-triphenyl tetrazolium chloride; RT-qPCR, Quantitative real-time pcr; c-Fos, Immediate early gene c-Fos; GABA,
The data that support the findings of this study are available in the Supplementary Material of this article.
Conceptualization, JLH and LXM; methodology, JLH, YXZ, and JSW; software, JYX, and MYC; validation, JYX; formal analysis, JLH; investigation, YXZ, JSW, and LHM; resources, LXM; data curation, YXZ and MYC; writing—original draft preparation, JLH; writing—review and editing, JLH and LXM; visualization, JLH and JYX; supervision, LHM; project administration, LXM; funding acquisition, LXM. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All animal procedures were performed in accordance with the NIH Guide for the Care and Use of Laboratory Animals and approved by the Ethics Committee for Animal Experimentation of Beijing University of Chinese Medicine (BUCM-2024101002-4017).
Not applicable.
This research was funded by the National Natural Science Foundation of China, grant number 82274655.
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/JIN46980.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.