



















1 Department of Neonatology, Lanzhou University Second Hospital, 730030 Lanzhou, Gansu, China
†These authors contributed equally.
Abstract
To address the unmet need for targeted therapeutic strategies for neonatal hypoxic–ischemic encephalopathy (HIE), we developed a brain-targeting lipid nanoparticle delivery system capable of silencing circular RNA homeodomain-interacting protein kinase 2 (circHIPK2) in astrocytes and investigated its ability to mediate neuroinflammation and improve neurological outcomes.
Dual-modified 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-poly (ethylene glycol)-neurotropic virus-derived peptide (DSPE-PEG2000-RVG29) peptide/mannose-functionalized lipid nanoparticles loaded with circHIPK2 small interfering RNA (M-R@siC-NPs) were constructed, and their physicochemical properties, stability, and biocompatibility were characterized. Using an in vitro oxygen-glucose deprivation (OGD) model and a neonatal murine hypoxic–ischemic brain damage (HIBD) model, we evaluated the effects of circHIPK2 silencing by the M-R@siC-NPs on the expression of two astrocyte activation markers, glial fibrillary acidic protein (GFAP) and interleukin-1β (IL-1β), via western blotting, quantitative reverse transcription-polymerase chain reaction (qRT-PCR), and immunofluorescence staining. Neurobehavioral recovery was assessed through righting reflex, negative geotaxis, and Morris water maze tests.
M-R@siC-NPs exhibited a uniform size distribution (134 nm), good blood–brain barrier penetrability, and astrocyte-targeting specificity. The nanoparticles effectively silenced circHIPK2 while demonstrating excellent colloidal stability and biosafety. In vitro, circHIPK2 knockdown by M-R@siC-NPs markedly suppressed OGD-induced astrocyte activation, reducing GFAP and IL-1β expression (p < 0.01). In HIBD mice, M-R@siC-NPs attenuated hippocampal astrocyte activation and improved motor coordination (shortened righting reflex latency, p < 0.0001) and spatial memory (increased platform crossings in Morris water maze, p < 0.0001).
The RVG29/mannose dual-modified M-R@siC-NPs precisely regulated astrocyte activation and attenuated neuroinflammation, effectively ameliorating brain injury in HIBD mice. This study establishes a novel RNA interference-based therapeutic strategy for targeted neuroinflammatory modulation, providing a promising translational platform for HIE treatment.
Keywords
- hypoxic–ischemic encephalopathy
- lipid nanoparticles
- astrocyte activation
- circHIPK2
- targeted gene therapy
Neonatal hypoxic–ischemic encephalopathy (HIE), triggered by perinatal asphyxia, poses a critical threat to neonatal health and represents a major cause of neonatal mortality and childhood disability [1, 2, 3, 4]. Its incidence ranges from 1–8% in developed countries and is reportedly as high as 26% in underdeveloped regions, with global mortality rates of 10–60%. Among survivors, at least 25% of affected infants suffer long-term neurodevelopmental sequelae, with 15–28% of pediatric cerebral palsy cases attributed to HIE [5, 6]. The pathological mechanisms involve energy metabolism dysfunction, excitotoxicity, oxidative stress, neuroinflammation, and apoptosis.
Current therapeutic options for HIE remain limited. While therapeutic hypothermia—the clinical gold standard—has demonstrated neuroprotective effects via the attenuation of astrocyte overactivation, 44–53% of treated infants still experience mortality or severe neurological deficits [7]. Stem cell transplantation, constrained by a narrow therapeutic window, dosage uncertainties, and individualized strategy challenges, faces limited clinical applicability [8]. These limitations underscore the urgent need for novel therapeutic approaches for HIE.
Astrocytes exhibit dual roles in central nervous system (CNS) inflammation
[9, 10, 11, 12, 13]. While they are initially protective during lesion isolation and
blood–brain barrier (BBB) restoration [14, 15], they become harmful under
ischemic stress in HIE. In the latter context, overactivated astrocytes release
pro-inflammatory cytokines such as tumor necrosis factor-
Circular RNAs (circRNAs), which are covalently closed structures lacking 5′
caps and 3′ poly(A) tails, have emerged as key regulators in brain disorders
due to their exceptional stability [21, 22, 23]. For example, circHIPK2 has
been identified as a key regulator of astrocyte activation through a molecular
mechanism involving miR-124-2HG sequestration and subsequent Sigma non-opioid
intracellular receptor 1 (SIGMAR1)-mediated endoplasmic reticulum
stress/autophagy pathway activation [24]. In lipopolysaccharide-induced
neuroinflammation models, activation of the cortical circHIPK2/SIGMAR1
axis correlates with upregulated TNF-
The BBB, although a critical protective mechanism, severely hinders drug delivery to the CNS [26, 27, 28, 29]. RNA interference (RNAi) technology offers therapeutic potential, yet the application of small interfering RNA (siRNA) as a drug faces clinical translation challenges due to rapid in vivo degradation and poor cellular uptake [30, 31]. Delivery of siRNA within nanocarriers can address these limitations through enhanced biocompatibility and modifiability, enabling efficient siRNA encapsulation and delivery [32, 33]. Mesoporous silica nanoparticles (MSNs) offer tunable pore structures and high drug-loading capacity, making them promising gene vectors [34, 35, 36, 37, 38]. Lipid nanoparticles (LNPs) have been validated for safety and efficacy and shown to improve targeted delivery while minimizing off-target effects [32, 39, 40, 41]. For example, glioma-targeting LNPs developed by Kuang et al. [42] demonstrated low systemic toxicity in clinical applications. However, delivery systems specifically for modulation of circHIPK2 expression remain unexplored, hindering the translational progress for circHIPK2-based therapies.
Recent advances have highlighted RVG29, a 29-amino-acid peptide derived from rabies virus glycoprotein, as a BBB-penetrating ligand. 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-poly (ethylene glycol)-neurotropic virus-derived peptide (DSPE-PEG2000-RVG29) binds to nicotinic acetylcholine receptors on endothelial cells of the BBB and facilitates receptor-mediated transcytosis [43, 44, 45, 46, 47]. Concurrently, overexpression of mannose receptor on astrocytes enables targeted delivery by mannose-decorated nanoparticles [48, 49, 50, 51].
In the present study, we engineered a dual-targeted siRNA nanoplatform consisting of RVG29 peptide/mannose-functionalized LNPs loaded with circHIPK2 siRNA (M-R@siC-NPs) as a potential therapy for HIE. This approach attempted to combine the advantages of LNPs and MSNs to achieve precise astrocyte targeting, suppress their pathological activation, and ameliorate neonatal hypoxic–ischemic brain injury. We tested the efficacy of M-R@siC-NPs both in vitro and in vivo, and the results indicate that this innovative strategy may provide novel therapeutic avenues for HIE and advance RNAi-based targeted gene therapy.
Aminated dendritic MSNs (NH2-MSNs, cat no. R-SG-89NH2), 1,2-dipalmitoyl-sn-glycero-3-phosphocholine
(DPPC, cat no. LP-R4-057), cholesterol,
1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[methoxy(polyethylene
glycol)-2000] (DSPE-PEG2000, cat no. R-H54510), DSPE-PEG2000-RVG29 (cat no. R9985), Cy5.5 (cat no. D10061), and M-PEG-lipid (cat. no. LP-R4-18) were
purchased from Ruixi Biotechnology Co., Ltd. (Xi’an, Shaanxi, China). Three siRNA (cat no. 15258)
sequences targeting circHIPK2 (circHIPK2 siRNA) and a non-homologous negative
control (siNC) without sequence homology to the target gene, along with
fluorescein amidite (FAM)-labeled siRNA, were obtained from Qingke Biotechnology
Co., Ltd. (Beijing, China). The siRNA-mate Plus transfection kit was acquired
from GenePharma Co., Ltd. (cat no. G04026, Shanghai, China). Chloroform (CHCl3
, cat no. CX1060)
and other analytical-grade reagents were sourced from Chongqing Chuandong
Chemical Group Co., Ltd. (Chongqing, China). Fetal bovine serum (FBS) was
acquired from Lanzhou Minhai Bio-Engineering Co.Ltd. (cat no. SA211.02, Lanzhou,
Gansu, China). Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12 (DMEM/F12,
cat no. 11320033), and 0.25% trypsin-ethylenediaminetetraacetic acid (EDTA, cat
no. 25200056) were purchased from Gibco BRL (Grand Island, NY, USA). The Cell
Counting Kit-8 (CCK-8) was obtained from Boster Biological Technology Co., Ltd.
(cat no. AR1199, Wuhan, Hubei, China). Trizol (cat no. RR036A), PrimeScript RT
reagent Kit (cat no. RR092S), and SYBR Green PCR Master Mix (cat no.RR820A) were
supplied by Takara Bio, Inc. (Shiga City, Shiga Prefecture, Japan). Quantitative
real-time polymerase chain reaction (qPCR) primers (cat no. TSE20240906-029) were synthesized by ZerKcorp
Biotech (Xi’an, Shaanxi, China). Agarose powder (cat no. GC205013), SerRed Nucleic
Acid Stain (10,000
Specific siRNAs were designed to silence the expression of circHIPK2. The efficacy of circHIPK2 silencing was evaluated using quantitative reverse transcription PCR (qRT-PCR) and western blotting. A total of four groups were established: three experimental groups transfected with each siRNA targeting circHIPK2 (circHIPK2-1, circHIPK2-2, and circHIPK2-3) and one control group without siRNA transfection. The siRNA-mate Plus transfection reagent was used to deliver the siRNAs into astrocytes. The siRNA demonstrating the most efficient silencing effect was selected for subsequent experiments (Table 1).
| Gene | Sense | Antisense |
| sicircHIPK2-1 | 5′-GGAAGAGUAAGCAGCACCA-3′ | 5′-UGGUGCUGCUUACUCUUCC-3′ |
| sicircHIPK2-2 | 5′-GAGACGGGAAUUAAGUCAA-3′ | 5′-UUGACUUAAUUCCCGUCUC-3′ |
| sicircHIPK2-3 | 5′-GCUUAGUCUUUGAGAUGUU-3′ | 5′-AACAUCUCAAAGACUAAGC-3′ |
siRNA, small interfering RNA; HIPK2, homeodomain-interacting protein kinase 2.
Agarose gel electrophoresis was performed to evaluate the siRNA loading capacity
of NH2-MSNs. At room temperature, siRNA (218 µg/mL) and NH2-MSNs
(5 mg/mL) were mixed at mass ratios (1:0, 1:10, 1:20, 1:25, 1:30, w/w), followed
by incubation for 30 min. After incubation, 6
Liposomes capable of crossing the BBB were prepared by mixing DPPC, cholesterol,
M-PEG-lipid, and DSPE-PEG2000-RVG29 at a mass ratio of 3:1:1:1, dissolving the
mixture in chloroform, and evaporating the solvent under vacuum to form a lipid
film. The film was hydrated with 4 mL PBS and sonicated for 20 min to obtain a
nanoemulsion. The emulsion was then emulsified under ice-cold conditions with 125
W ultrasonication for 8 min, followed by centrifugation at 6100
For nanoparticle assembly, circHIPK2 siRNA and NH2-MSNs were mixed
at a mass ratio of 1:30, incubated at room temperature for 10 min, and
electrostatically bound to form NH2-MSN-siRNA complexes. The liposomes were
then combined with NH2-MSN-siRNA at a 1:1 mass ratio, incubated at room
temperature for 10 min, and subjected to ultrasonication (3 min pulses with 5-s
intervals). After centrifugation at 6100
As controls, target-free LNPs (siC-NPs) were prepared using the same method but omitting DSPE-PEG2000-RVG29 and M-PEG-lipid. The full name of siC-NPs is non-targeted modified circHIPK2 siRNA-lipid nanoparticles, which serve as non-targeted control nanoparticles in this study. Their core carrier components are consistent with those of M-R@siC-NPs (mannose/RVG29 peptide dual-modified circHIPK2 siRNA-lipid nanoparticles), with the only difference being the lack of two targeted ligands, namely M-PEG-lipid and DSPE-PEG2000-RVG29. Both siC-NPs and M-R@siC-NPs carry circHIPK2 siRNA3. Fluorescently labeled Cy5.5-conjugated targeting (Cy5.5-M-R@siC-NPs) and non-targeting nanoparticles (Cy5.5-siC-NPs) were also synthesized for fluorescence imaging.
The Malvern Nano analyzer (cat no. ZEN-3600, Zeta-Sizer, Malvern Instruments, Malvern, UK) was used to measure particle size and surface zeta potential. For sample preparation, 10 µL of nanoparticle solution was spotted onto a 400-mesh carbon-coated copper grid, air-dried at room temperature, and imaged using transmission electron microscopy (TEM, cat no. HT7800, Hitachi, Tokyo Metropolis, Japan) to observe the morphology, structure, and size of NH2-MSNs and M-R@siC-NPs.
The colloidal stability of M-R@siC-NPs was evaluated by monitoring hydrodynamic
particle size changes via The Malvern Nano analyzer over 14 days (0, 2, 4, 6, 8, 10, 12, and 14 days).
Additionally, M-R@siC-NPs suspensions were separately incubated in DMEM/F12 cell
culture medium, PBS (pH 7.4), pure FBS, and artificial cerebrospinal fluid (aCSF)
at ambient temperature (25 °C). Morphological changes, including macroscopic
aggregation or precipitation, were observed and recorded daily for 14 days. This
temperature represents the typical ambient temperature used for simulating the
storage of nanoformulations and in vitro experiments, and it is also
consistent with the specifications on room temperature conditions for long-term
stability testing as outlined in the “Guidelines for the Stability Testing of
Pharmaceutical Preparations” of the Chinese Pharmacopoeia (25 °C
The siRNA concentration was quantified using a microvolume UV-Vis
spectrophotometer (cat no. DENOVIX DS-11, DENOVIX, Wilmington, DE, USA) at
260 nm. Encapsulation efficiency (EE, %) was calculated via centrifugation
(14,000
Drug-loading capacity (DLC, %) was calculated using the following formula:
C57BL/6 neonatal mice (specific pathogen-free grade) were purchased from Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences. All histopathological evaluations were guided by faculty members specializing in pathology from the School of Basic Medical Sciences, Lanzhou University. The animal experimental protocols were approved by the Animal Ethics Committee of the Second Hospital of Lanzhou University (approval number: D2024-439).
Primary astrocytes were seeded into 96-well plates at a density of 1
Nanoparticle solutions (20, 40, 60, 80, and 100 µg/mL), PBS (negative
control), or triple-distilled water (positive control) were added to 1.5-mL
eppendorf tubes (1 mL per group). Blood was collected from the orbital venous
plexus of healthy C57BL/6 mice using EDTA anticoagulant tubes. After mixing, the
blood was centrifuged at 3000
In strict accordance with the guidelines on non-clinical safety studies of pharmaceuticals issued by the International Council for Harmonisation of Technical Requirements for Pharmaceuticals for Human Use (ICH), these guidelines explicitly require that the potential effects of pharmaceuticals on major physiological functions (cardiovascular, respiratory, central nervous systems) be evaluated. Additionally, the study design and implementation refer to the Good Laboratory Practice for Non-Clinical Studies (GLP) promulgated by the National Medical Products Administration (NMPA) of China. Although this study does not belong to the anti-tumor field, the requirement in this guideline that nanopharmaceuticals should focus on carrier accumulation, organ-targeted distribution, and long-term toxicity is universally applicable. To evaluate the systemic toxicity of the nanoparticles, 10-day-old C57BL/6 neonatal mice were randomly divided into four groups (n = 3 per group, statistical analysis is conducted based on the complete set of sample data): a NC group, PBS group, siC-NPs group (80 µg/mL), and M-R@siC-NPs group (80 µg/mL). Nanoparticles were administered intravenously for 20 consecutive days. Blood samples were collected for complete blood count (CBC) analysis using a Mindray Veterinary Automated Hematology Analyzer (cat no. BC-2800vet, Mindray Biomedical Electronics Co., Ltd. Shenzhen, Guangdong, China). Serum was obtained via refrigerated centrifugation, and biochemical parameters were analyzed using an Automated Biochemical Analyzer (cat no. Chemray 240, Rayto Life Technology Co., Ltd. Shenzhen, Guangdong, China). Mice were anesthetized via intraperitoneal injection of 1% sodium pentobarbital at a dose of 50 mg/kg body weight. Anesthesia was confirmed when the mice exhibited no response to toe pinch and loss of corneal reflex. Subsequently, the mice were euthanized by cervical dislocation, in compliance with the Animal Ethics Guidelines of the Second Hospital of Lanzhou University. Death was verified by the absence of breathing and heartbeat for 5 consecutive minutes. After euthanasia, heart, liver, spleen, lung, kidney, and brain tissues were collected, fixed, and stained with H&E. Histopathological changes were quantitatively analyzed using the TissueFAXS Plus panoramic tissue cytometry system (cat no. TissueFAXS Plus, TissueGnostics GmbH, Vienna, Vienna State, Austria).
Primary astrocytes were seeded on confocal dishes at a density of 1
Ten-day-old C57BL/6 neonatal mice were randomly divided into three groups (n = 9 per group): a PBS group, Cy5.5-siC-NPs group, and Cy5.5-M-R@siC-NPs group. The corresponding agents were intraperitoneally injected into each group. At time points of 0, 1, 2, 4, 6, 8, 10, 12, 24, and 48 h post-injection, in vivo fluorescence imaging was performed using a small-animal imaging system (Vie-works Smart-LF, Eschborn, Hessen, Germany) to monitor nanoparticle accumulation and assess targeting specificity. Mice were euthanized when the peak intracranial fluorescence intensity was achieved, and brain tissues were harvested for ex vivo imaging. Heart, liver, spleen, kidney, and lung tissues were also collected and imaged using the Vie-works Smart-LF small-animal live imaging system to observe the Cy5 fluorescence distribution in major organs [52]. Brain tissue sections were further processed with 4’,6-Diamidino-2-Phenylindole (DAPI, cat no. D1306, Thermo Fisher Scientific Inc. Waltham, MA, USA) staining for pathological examination.
Primary astrocytes were isolated from postnatal day 2–3 C57BL/6 neonatal mice
anesthetized with isoflurane and disinfected with 75% ethanol. Whole brains were
dissected under a biosafety cabinet, minced in pre-cooled Hank’s Buffered Salt
Solution (HBSS) buffer, and digested with 0.25% trypsin-EDTA at 37 °C
for 5 min. The digestion was halted by adding DMEM/F12 medium containing 10%
FBS, followed by centrifugation at 1200
For the in vitro hypoxia–ischemia (HI) model, the cell culture medium was switched to serum-free medium 24 h before the cells were subjected to OGD. After two washes with PBS, the cells were incubated in serum-free and glucose-free medium and placed in a hypoxic chamber (1% O2, 5% CO2, 94% N2, cat no. Coy-HC-05, Coy Laboratory Products, Inc. Grass Lake, MI, USA) at 37 °C for 6 h. The medium was then exchanged with complete medium for 24–48 h of recovery culture before collection for further analysis.
For the in vivo HIBD model, 6–8-day-old C57BL/6 neonatal mice were anesthetized with 2% isoflurane. The left common carotid artery was permanently ligated with an 8-0 surgical suture. Post-surgery, mice were exposed to a hypoxic environment (8% O2, 92% N2, flow rate 1.5 L/min) in a chamber for 2 h. Successful establishment of the HIBD model was verified using laser speckle imaging. Mice were randomized to four groups: the sham group, HIBD group, siC-NPs group, and M-R@siC-NPs group. Brain morphology was observed at 3, 5, and 7 days post-surgery. Brain tissues were collected, sectioned, and subjected to H&E staining and immunofluorescence analysis for assessment of pathological changes.
After treatment, brain tissues and cells were washed 2–3 times with pre-cooled
PBS. Protein lysates were prepared using radioimmunoprecipitation assay (RIPA)
lysis buffer (Ailv Life, AIWB-012, Wuhan, Hubei, China) containing protease
inhibitors, incubated on ice for 30 min, and centrifuged at 12,000
Mice in the siC-NPs and M-R@siC-NPs groups (n = 12/group) received
intraperitoneal injections of LNPs (80 µg/mL) for 5 consecutive days,
followed by a 2-day pause, totaling four cycles over 28 days (100 µL per
dose). Post-treatment, three mice per group were euthanized for histopathological
analysis, including H&E staining and GFAP immunofluorescence staining to assess
glial activation. Samples from parallel groups (n = 3/group) were used for
western blot analysis to quantify GFAP and IL-1
This test was administered at the end of the first treatment cycle (day 5). Mice
were placed ventral-side down on a soft pad, and the time to resume prone
position was recorded (
This test was conducted post-modeling at the end of the first treatment cycle (day 5). Mice were positioned head-down at a 15° incline on a 30-cm slope. Latency to complete a 180° turn toward the top was measured (cutoff: 60 s). The results from three trials per mouse were averaged.
This test was performed on post-treatment day 28 using a 120-cm diameter pool
(water depth: 50 cm, temperature: 21
Statistical analysis was performed using GraphPad Prism 10.1.2 (GraphPad Software, Inc. La Jolla, CA,
USA). All data are presented as mean
In the development of functional nanoparticles for this study, the first
challenge was to identify highly efficient circHIPK2 silencing
sequences. For this purpose, primary astrocytes were transfected with three
distinct circHIPK2 siRNAs (circHIPK2-1, circHIPK2-2,
and circHIPK2-3). FAM-labeled siRNAs were utilized to track transfection
efficiency (Fig. 1A). qRT-PCR results showed that compared with the Control group
(without siRNA transfection), all three circHIPK2 siRNAs significantly
reduced the expression level of circHIPK2 mRNA in primary astrocytes.
Among them, circHIPK2 siRNA-3 exhibited the optimal silencing
efficiency, mediating an 80.13% decrease in circHIPK2 mRNA expression.
This efficiency was significantly higher than that of circHIPK2 siRNA-1
and circHIPK2 siRNA-2, with a statistically significant difference
(p
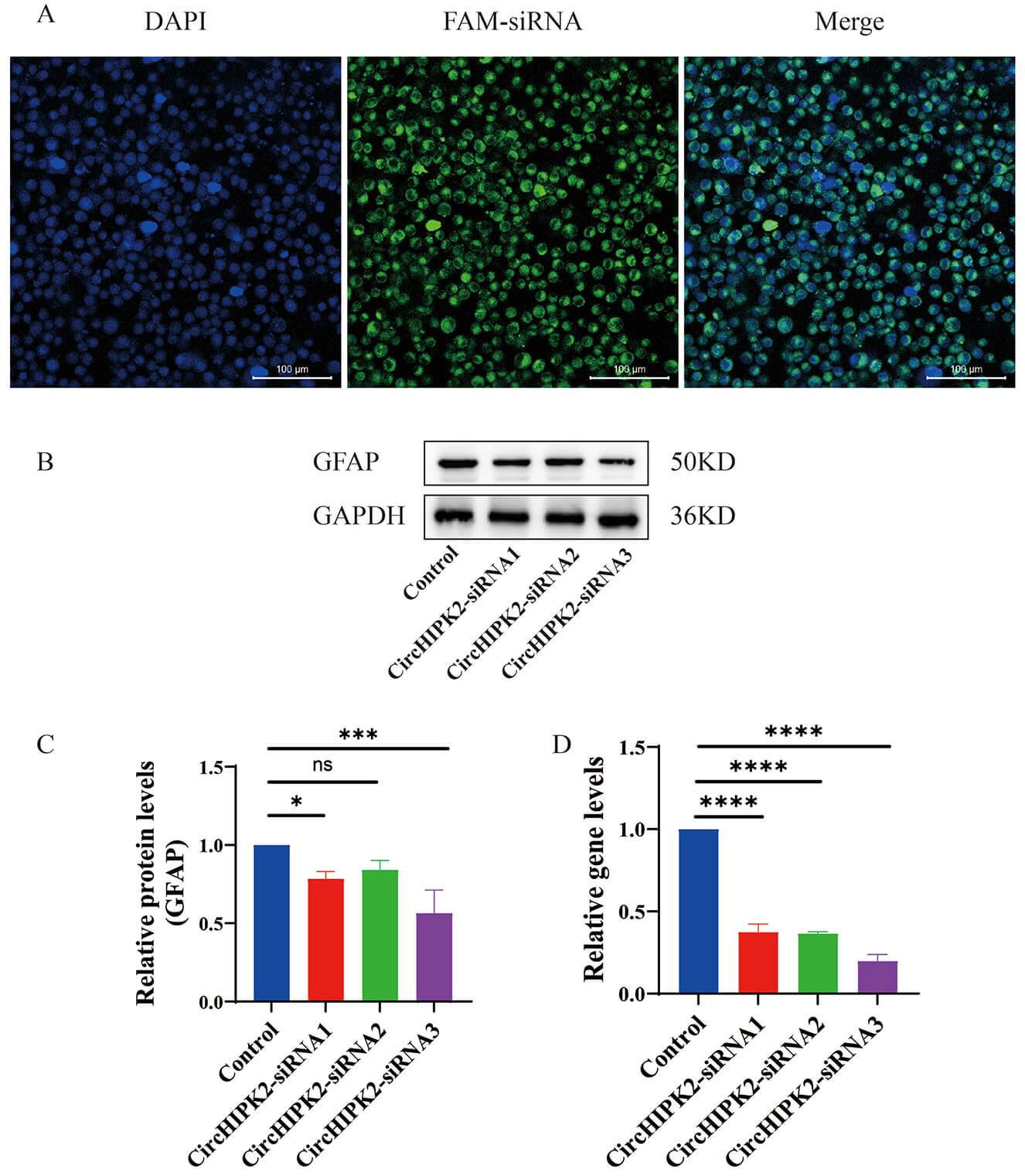 Fig. 1.
Fig. 1.
Silencing efficiency of three different circHIPK2 siRNA
sequences in astrocytes. (A) Cellular localization of FAM-labeled siRNAs (green
fluorescence) at 48 h post-transfection, with nuclei counterstained by DAPI
(blue). Scale bar = 100 µm. (B) Western blot analysis of glial fibrillary
acidic protein (GFAP) protein expression across groups. GAPDH served as a loading
control. (C) Quantitative analysis of GFAP protein levels normalized to GAPDH
expression (n = 3). (D) Quantitative reverse transcription PCR (qRT-PCR) analysis
of circHIPK2 mRNA expression (n = 3). Data are presented as mean
The physicochemical properties of the different synthesized nanoparticles are
summarized in Fig. 2. Dynamic light scattering (DLS) analysis revealed that the hydrodynamic diameters of
the NH2-MSNs, NH2-MSN-siRNA, and M-R@siC-NPs were 115.80
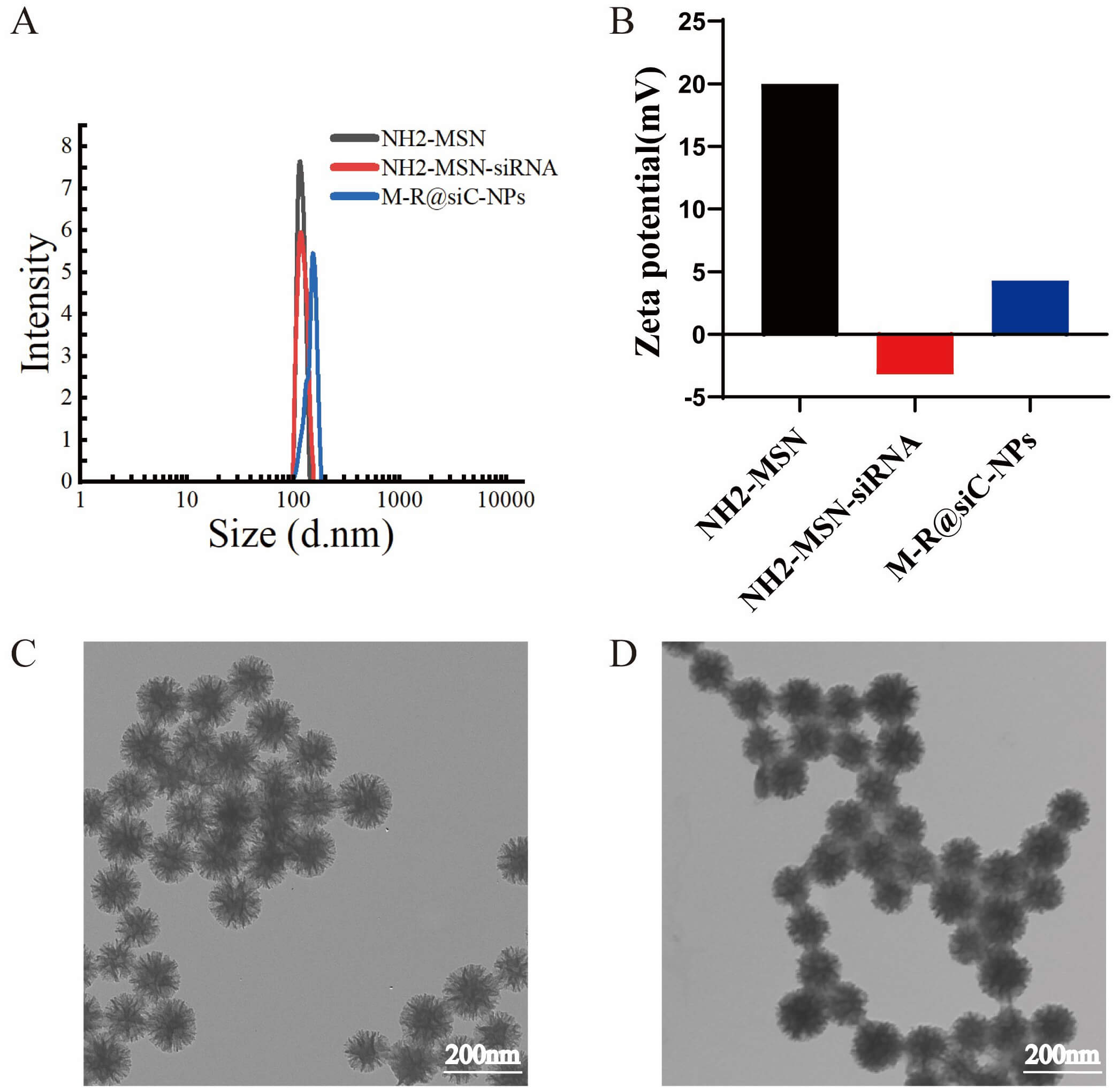 Fig. 2.
Fig. 2.
Physicochemical characterization of nanoparticles. (A)
Hydrodynamic size distribution of different nanoparticle formulations (n = 3;
data presented as mean
To determine the optimal mass ratio for binding circHIPK2 siRNA (218
µg/mL) with NH2-MSNs (5 mg/mL), agarose gel electrophoresis was
performed. The results demonstrated that when the mass ratio of siRNA to
NH2-MSNs reached 1:30 or higher, the fluorescence intensity of the siRNA
band decreased (Fig. 3A), indicating successful electrostatic binding between the
positively charged NH2-MSN and negatively charged siRNA. Thus, a 1:30 mass
ratio was employed in the preparation of nanoparticles for subsequent
experiments. To evaluate the encapsulation efficiency and drug loading capacity
of the nanoparticles, ultramicroscopic UV-Vis spectrophotometry was used to
quantify siRNA content (R2 = 0.9983). The calculated encapsulation
efficiency and drug loading capacity were 75.28%
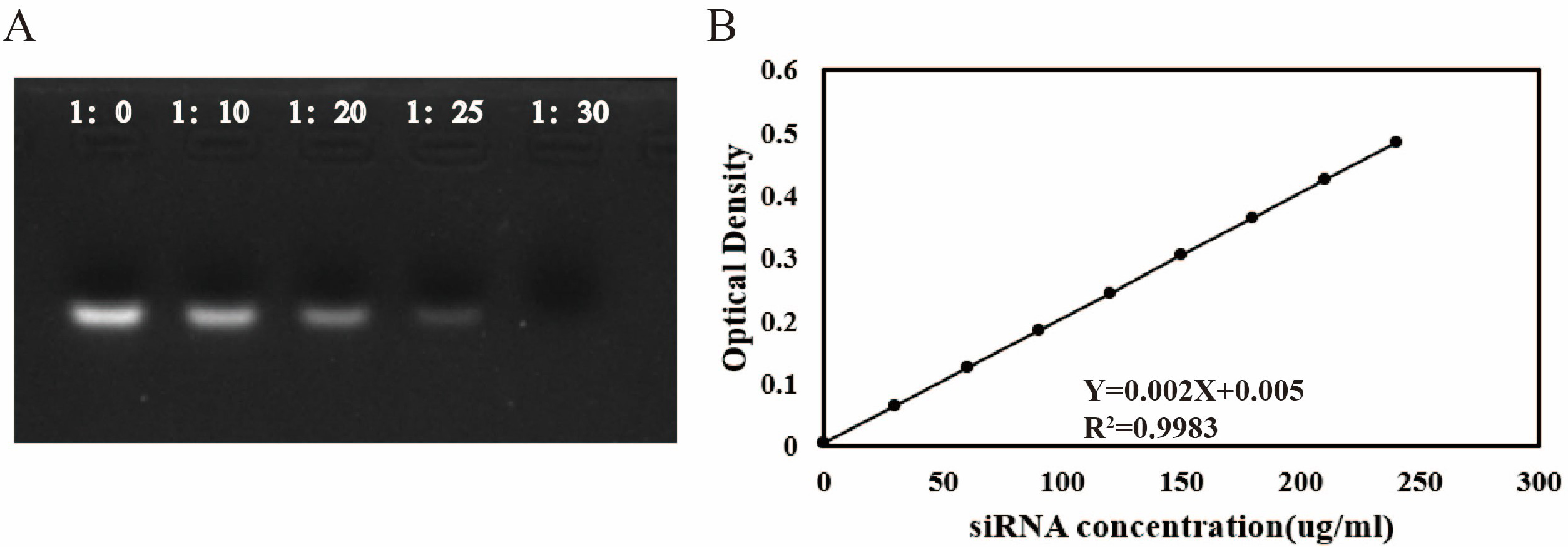 Fig. 3.
Fig. 3.
Characterization of siRNA encapsulation efficiency and drug
loading capacity of nanoparticles. (A) Agarose gel electrophoresis analysis of
circHIPK2 siRNA binding to amino-functionalized dendritic silica
nanoparticles (NH2-MSNs) at varying mass ratios (1:0, 1:10, 1:20, 1:25, and
1:30). Disappearance of the siRNA band intensity indicates successful loading.
(B) Standard curve for siRNA quantification: absorbance at 260 nm was measured
for gradient siRNA solutions, yielding a linear regression equation of Y = 0.002X
+ 0.005 (R2 = 0.9983, n = 3; data presented as mean
To evaluate the stability and biocompatibility of nanoparticles in physiological
environments, the morphology and dispersion of nanoparticle suspensions in
DMEM/F12, PBS, FBS, and aCSF were observed over 14 days. The results demonstrated
that M-R@siC-NPs maintained a uniform dispersion without significant aggregation
or precipitation in all tested media. Notably, no evident interaction between
nanoparticles and serum proteins or culture medium components was observed in FBS
and DMEM/F12, confirming effective surface modification (Fig. 4A–C). These
findings indicate that M-R@siC-NPs exhibit excellent long-term stability, making
them suitable for in vitro cell culture and in vivo CNS
delivery applications. Additionally, DLS analysis revealed no significant
fluctuations in the hydrodynamic diameter of M-R@siC-NPs throughout the 14-day
study period (p
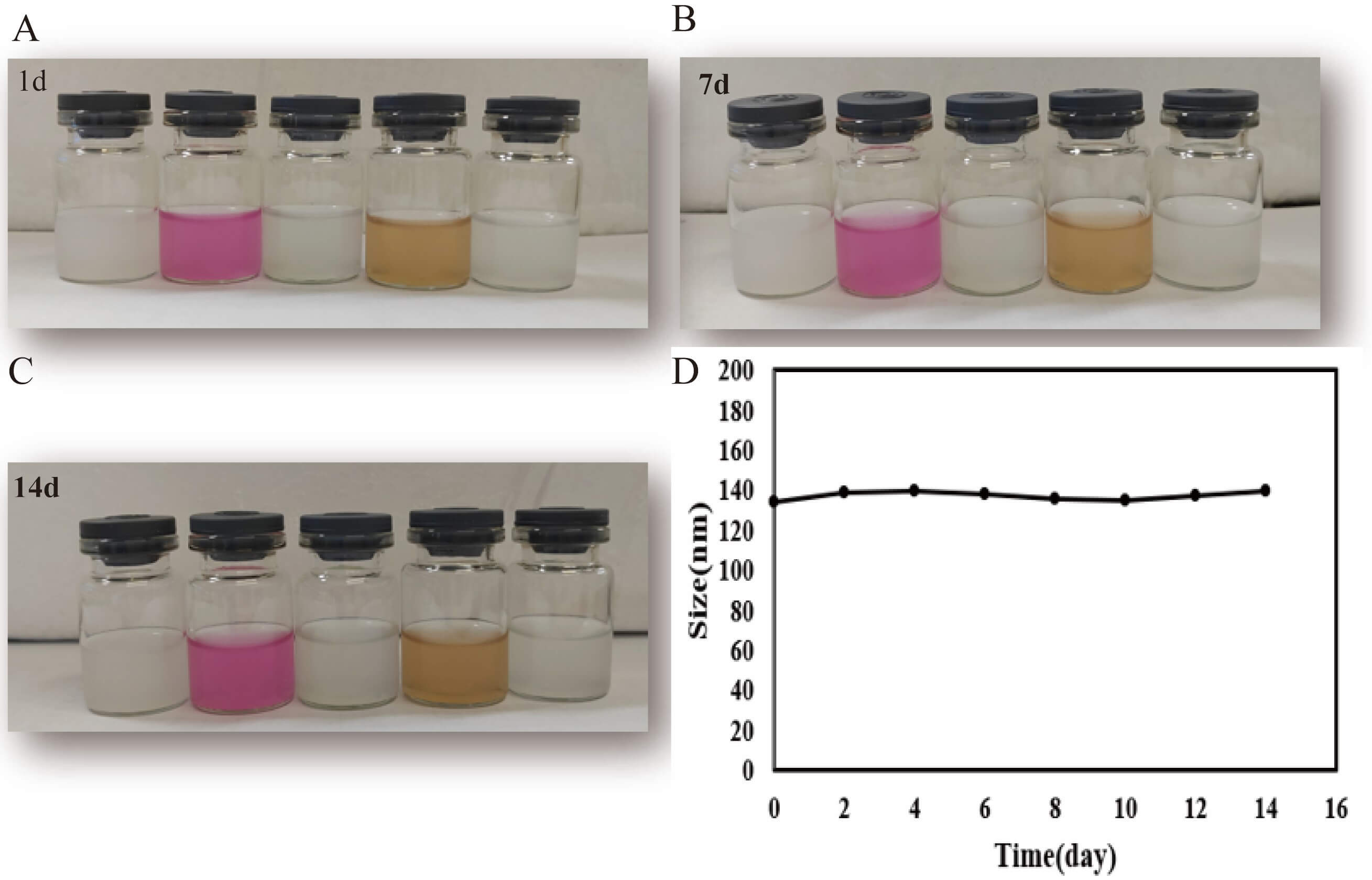 Fig. 4.
Fig. 4.
Physical stability of M-R@siC-NPs in various media. (A–C)
Macroscopic appearance of undiluted original dispersions of M-R@siC-NPs stored in
DMEM/F12 medium, PBS, FBS, and aCSF on days 1, 7, and 14 (stored at 25
°C in the dark). No visible aggregation or precipitation was observed in
any medium, confirming the excellent colloidal stability of the M-R@siC-NPs. (D)
Hydrodynamic size evolution of M-R@siC-NPs measured by DLS over 14 days (n = 3;
data presented as mean
The biocompatibility of LNPs was evaluated by adding them to cultures of primary astrocytes for 24 h, followed by cell viability analysis using the CCK-8 assay. The results demonstrated that astrocytes retained over 90% cell viability upon exposure to siC-NPs and M-R@siC-NPs at concentrations of 20–80 µg/mL. However, with nanoparticle concentrations of 100 µg/mL, astrocyte viability decreased to 89.01% (siC-NPs) and 87.59% (M-R@siC-NPs), and these reductions were significant compared with the viability of the control group (Fig. 5A). Based on these findings, 80 µg/mL was selected as the optimal concentration for subsequent experiments to ensure the biocompatibility of nanoparticle treatments (Fig. 5B).
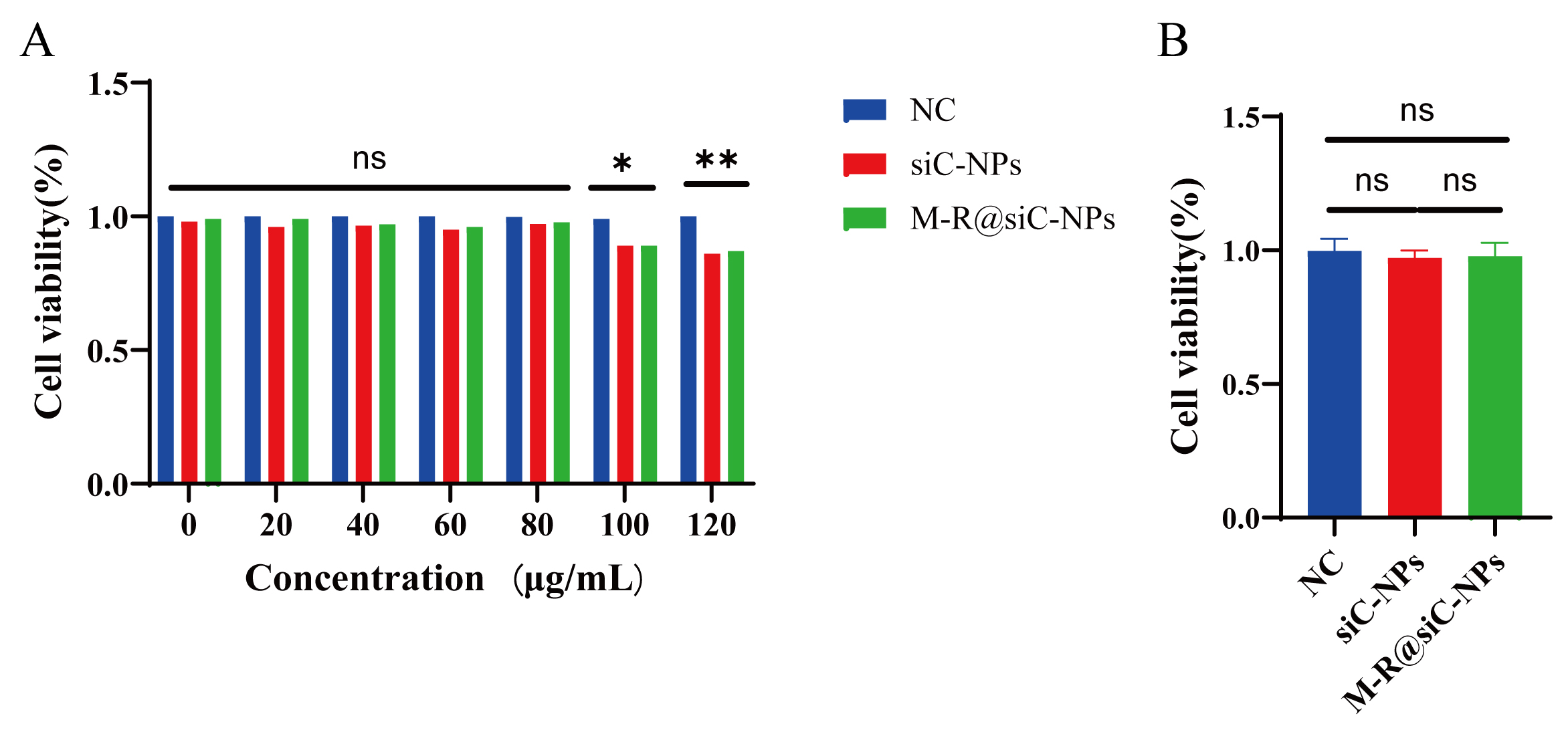 Fig. 5.
Fig. 5.
Nanoparticle cytotoxicity assessment for astrocytes. (A)
Effects of different concentrations (20, 40, 60, 80, 100, and 120 µg/mL) of
NC (normal control) group, siC-NPs, and M-R@siC-NPs on astrocyte viability,
assessed by CCK-8 assay (n = 3; data presented as mean
To evaluate the in vivo safety of the nanoparticles, a peripheral blood hemolysis assay was conducted to assess whether free nanoparticles in the bloodstream would induce acute erythrocyte destruction. At nanoparticle concentrations of 20, 40, 60, 80, and 100 µg/mL, the hemolysis rates remained less than 1.5% (Fig. 6A,B). From the appearance of centrifuged samples in Fig. 6A, the supernatants of all nanoparticle concentration groups remained colorless and transparent, which was completely consistent with the PBS negative control group. In contrast, the dark red supernatant observed in the positive control group (triple-distilled water) was absent in all nanoparticle groups. This macroscopic result directly confirms that even at the highest tested concentration (100 µg/mL), the nanoparticles did not induce visible red blood cell (RBC) lysis, indicating no significant hemolysis and initially verifying their hemocompatibility (Fig. 6A). The absorbance of the supernatants was measured at a wavelength of 540 nm using a microplate reader, and the hemolysis rate was calculated according to the formula. The results showed that the hemolysis rate of each concentration group was below 1.5%, which was far lower than the 5% safety threshold for hemolysis rate of biomaterials (Fig. 6B). The slight upward trend of hemolysis rate with increasing concentration was attributed to the slightly increased probability of non-specific contact between nanoparticles and RBC membranes as the nanoparticle concentration increased (from 20 to 100 µg/mL). This contact led to the release of a minimal amount of hemoglobin; however, the release volume was extremely low and did not reach the threshold for macroscopically visible color changes. These results confirmed that the nanoparticles did not induce erythrocyte destruction, indicating they offer an excellent in vivo safety profile.
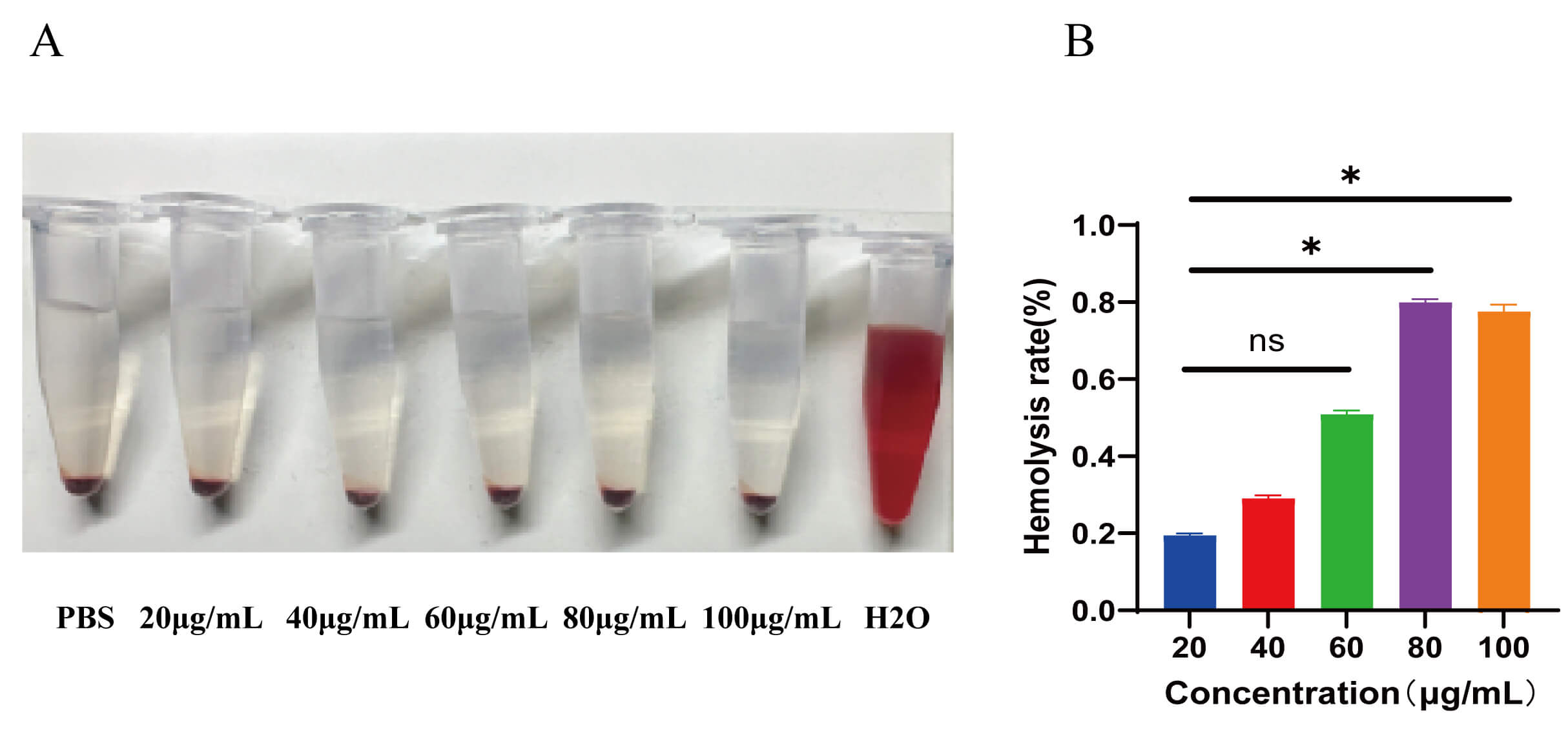 Fig. 6.
Fig. 6.
Hemolysis assessment upon nanoparticle exposure. (A) Hemolysis
assay setup: (left to right) negative control (1 mL PBS), nanoparticle-treated
groups (20, 40, 60, 80, or 100 µg/mL), and positive control (1 mL
ddH2O). After incubation at 37 °C for 3 h, samples were centrifuged
to observe hemolysis (red supernatant indicates erythrocyte lysis). (B)
Concentration-dependent hemolysis rates (n = 3; data presented as mean
For in vivo analysis of nanoparticle biocompatibility, this study
evaluated the functional status of major physiological organs in experimental
animals after daily intravenous injection of nanoparticles for 20 days. Blood
routine parameters (white blood cell [WBC] count, red blood cell [RBC] count,
hemoglobin [HGB], and platelet [PLT] count) and serum biochemical markers
(alanine aminotransferase [ALT], aspartate aminotransferase [AST], total
bilirubin [TBIL], albumin [ALB], alkaline phosphatase [ALP], creatinine [CREA],
blood urea nitrogen [BUN], and uric acid [UA]) were analyzed in both control and
LNP-treated groups. All values fell within the reference ranges for healthy mice,
with no adverse effects observed (Fig. 7). Statistical analysis revealed no
significant differences between groups (p
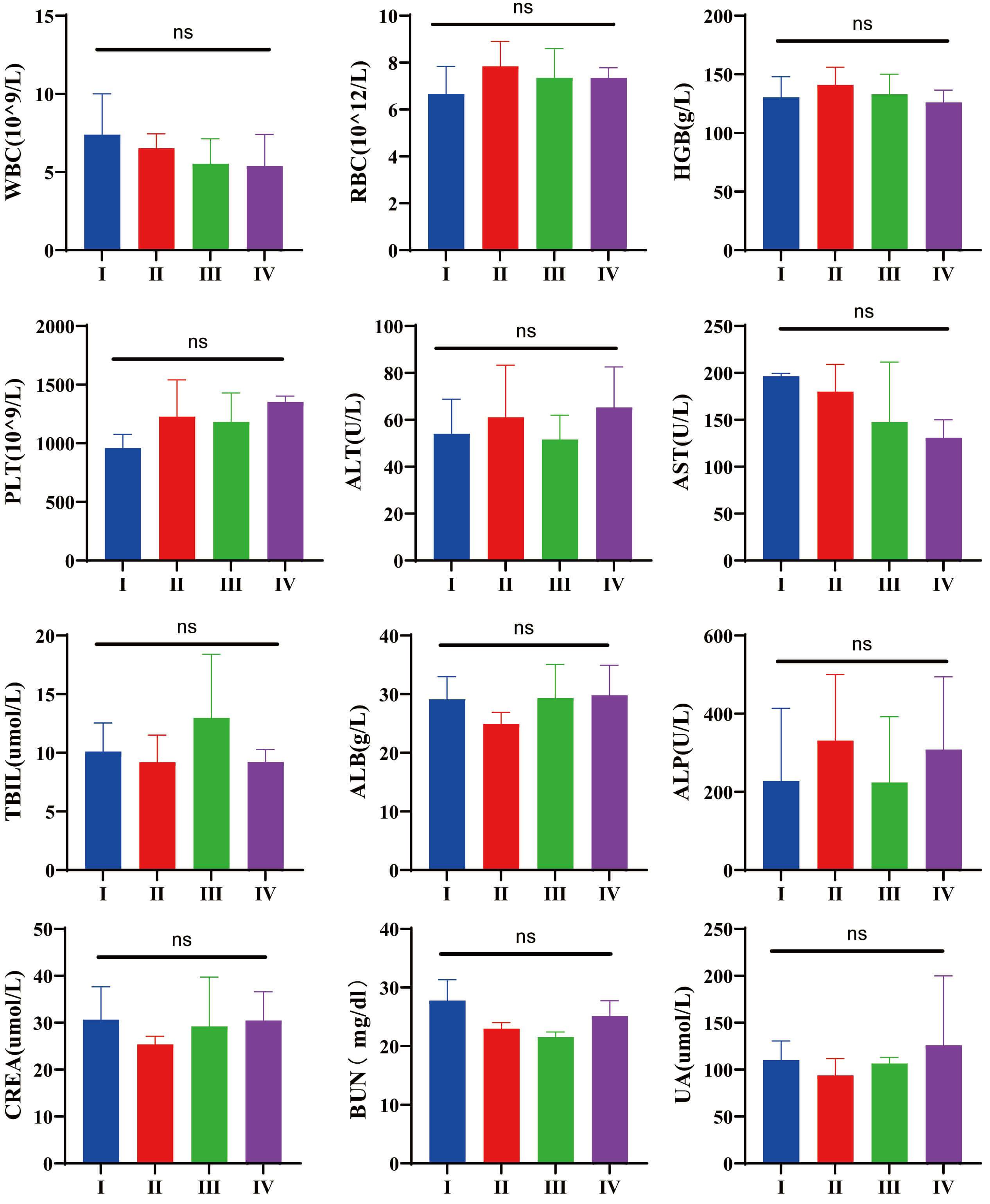 Fig. 7.
Fig. 7.
Toxicological evaluation of routine blood parameters and
hepatic/renal function indicators after exposure to different conditions. (I) NC
(normal control) group, (II) PBS-treated group, (III) siC-NPs-treated group, (IV)
and M-R@siC-NPs-treated group. No significant differences were observed in
routine blood parameters (WBC count, RBC count, HGB, and PLT count) or
hepatic/renal biochemical markers (ALT, AST, TBIL, ALB, ALP, CREA, BUN, and UA)
across the groups (ns: p
To assess the in vivo biosafety of the LNPs, histopathological analysis was performed on major organs (heart, liver, spleen, lung, kidney, and brain) harvested after 20 days of intravenous injection of the different particles. The H&E staining results demonstrated that, compared with the NC group, the nanoparticle-treated groups exhibited no significant histopathological abnormalities, showing no inflammatory cell infiltration, cellular edema, or necrosis in any organ (Fig. 8). These morphological observations confirmed that the LNPs caused no detectable cytotoxicity or structural damage to vital organs, underscoring their excellent in vivo biocompatibility and safety profile.
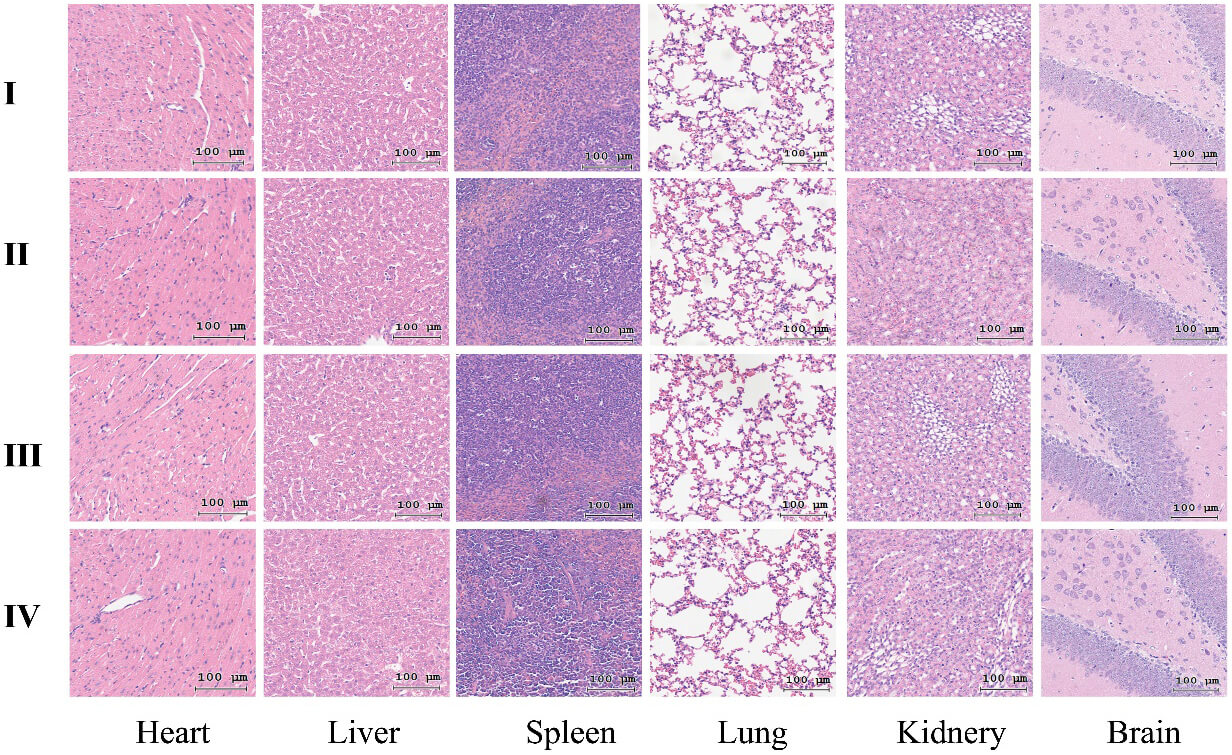 Fig. 8.
Fig. 8.
Histopathological analysis of major organs via H&E staining after exposure to nanoparticles in different experimental groups. (I) NC group, (II) PBS-treated group, (III) siC-NPs-treated group, and (IV) M-R@siC-NPs-treated group. Scale bars = 100 µm. H&E, hematoxylin and eosin.
Evaluations of cellular nanoparticle uptake efficiency and targeting revealed
distinct fluorescence patterns. No fluorescent signals were detected in the PBS
control group under laser scanning confocal microscopy (LSCM). In contrast,
significant intracellular fluorescence accumulation was observed in the groups
treated with Cy5.5-siC-NPs (80 µg/mL) and Cy5.5-M-R@siC-NPs (80
µg/mL). Quantitative analysis demonstrated that the mean fluorescence
intensity in these experimental groups was statistically higher than that in the
control group (Fig. 9A,B), with cellular uptake efficiency reaching 95.2%
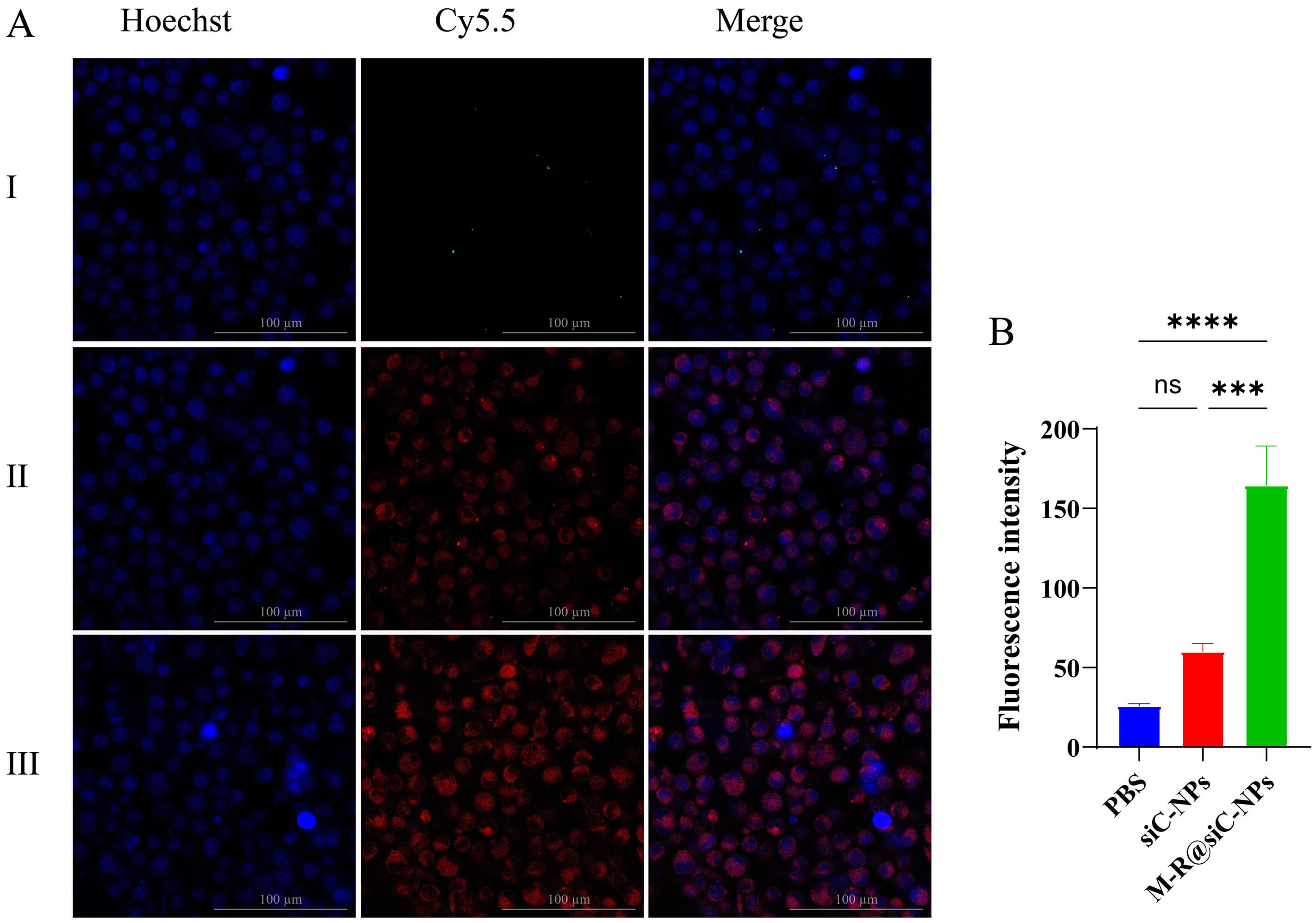 Fig. 9.
Fig. 9.
In vitro targeting evaluation. (A) LSCM images showing
the localization of Cy5.5-labeled siC-NPs (Group II) and Cy5.5-M-R@siC-NPs (Group
III) in astrocytes. Red fluorescence (Cy5.5) indicates nanoparticle distribution,
while blue fluorescence (DAPI) labels cell nuclei. Scale bar = 100 µm. (B)
Quantitative analysis of fluorescence intensity in Groups I (PBS control), II
(Cy5.5-siC-NPs), and III (Cy5.5-M-R@siC-NPs). Data are presented as mean
On the evaluation of the targeted delivery performance of the nanoparticles, the surface-modified Cy5.5-M-R@siC-NPs exhibited distinct advantages. LSCM imaging revealed significantly stronger fluorescence signals in the group treated with Cy5.5-M-R@siC-NPs compared with that treated with Cy5.5-siC-NPs. Fluorescence quantification confirmed that the intensity of Cy5.5-M-R@siC-NPs was three-fold higher than that of Cy5.5-siC-NPs (Fig. 9A,B), with a statistically significant difference observed. This outcome substantiates that the active targeting capability of M-R@siC-NPs can achieve drug delivery specificity.
In vivo live imaging revealed that intraperitoneally administered targeted nanoparticles (Cy5.5-M-R@siC-NPs) exhibited peak fluorescence intensity in the brain at 4 hours post-injection, with complete clearance observed by 48 hours (Fig. 10A-III). In contrast, non-targeted Cy5.5-siC-NPs displayed a delayed peak fluorescence at 6 hours, reduced intensity, and rapid clearance within 24 hours (Fig. 10A-II,C). Statistical analysis demonstrated that the brain accumulation of targeted nanoparticles was significantly higher than that of the non-targeted counterparts from 4–10 hours post-administration. These results confirm that the M-R@siC-NPs with brain-targeting modifications effectively penetrated the BBB, potentially offering enhanced cerebral bioavailability and an extended therapeutic window.
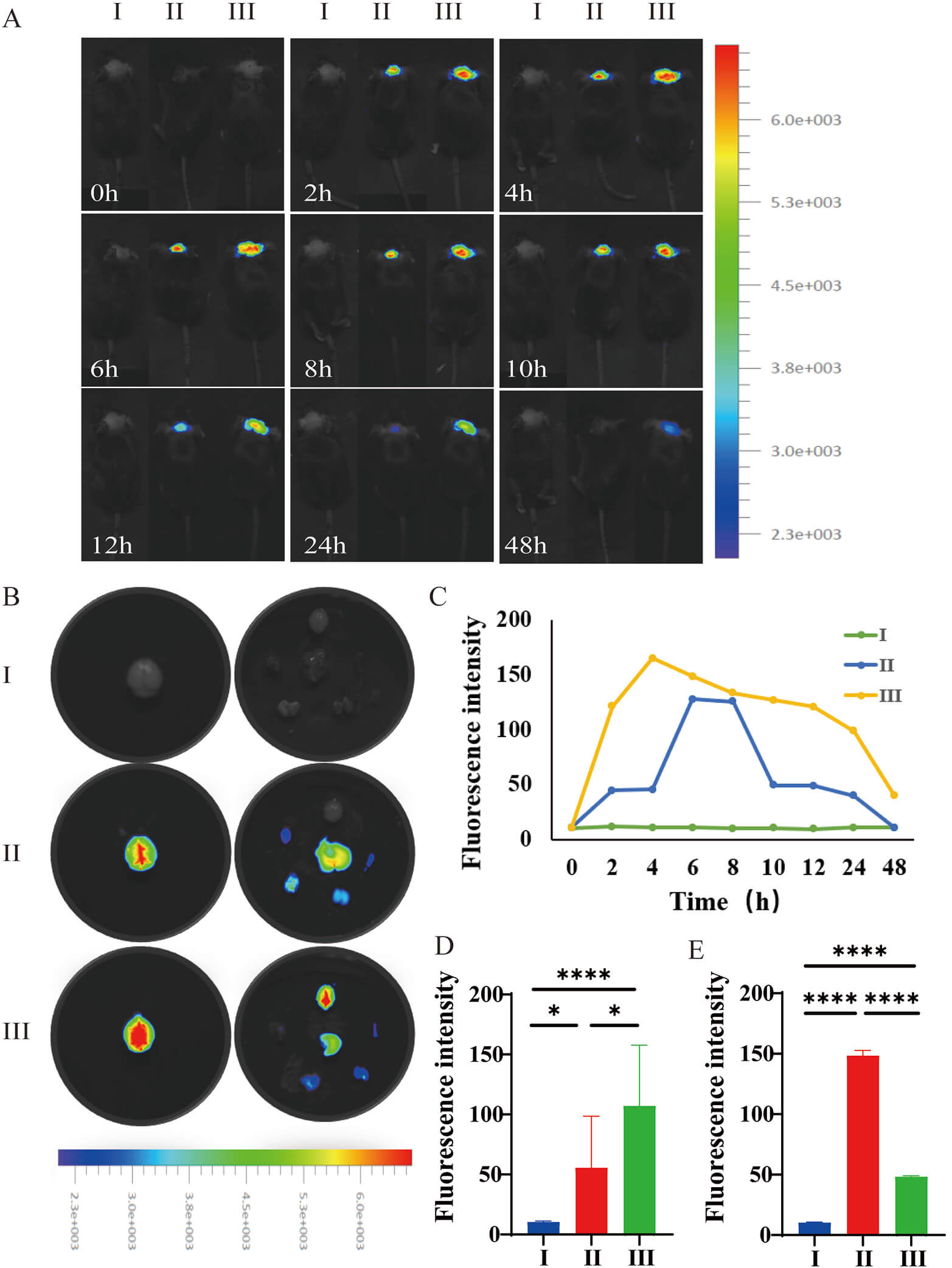 Fig. 10.
Fig. 10.
In vivo biodistribution and brain targeting efficiency
of nanoparticles. (A) Real-time fluorescence imaging of Cy5.5-labeled
nanoparticles in C57BL/6 neonatal mice after intraperitoneal injection (Groups I:
PBS; II: Cy5.5-siC-NPs; and III: Cy5.5-M-R@siC-NPs). Imaging was performed using
a chemiluminescence imaging system with excitation/emission wavelengths set to
675/720 nm (Cy5.5 channel). (B) Ex vivo fluorescence imaging of major
organs (heart, liver, spleen, lung, kidney, and brain) at the fluorescence
intensity peak (4–6 hours post-injection). (C) Dynamic changes in brain
fluorescence intensity (0–48 hours, region of interest [ROI] quantification, n =
9). (D) Quantitative analysis of brain fluorescence intensity across groups. (E)
Total fluorescence intensity comparison among major organs. Data are presented as
mean
Ex vivo fluorescence quantification further validated the brain-specific targeting efficacy of the modified nanoparticles (Fig. 10). Quantitative analysis of brain tissues revealed markedly stronger Cy5.5-M-R@siC-NPs fluorescence compared with Cy5.5-siC-NPs fluorescence (Fig. 10B,D), underscoring their superior BBB-penetrating capacity. Systemic biodistribution analysis (Fig. 10B,E) showed uniform distribution of the non-targeted nanoparticles across major organs, whereas Cy5.5-M-R@siC-NPs demonstrated selective enrichment in the brain, with statistically significant differences in fluorescence intensity observed. The results of this spatial distribution analysis support the notion that the brain-targeting ligands on M-R@siC-NPs minimize peripheral organ exposure while enhancing brain-specific delivery.
Microscopic localization in brain tissues provided insight into the BBB-penetration mechanism of M-R@siC-NPs (Fig. 11). Frozen sections stained with DAPI (blue fluorescence, nuclear counterstain) revealed distinct red fluorescence signals from Cy5.5-M-R@siC-NPs, with pronounced accumulation in the brain parenchyma and perivascular regions. Compared with controls and non-targeted nanoparticles, targeted nanoparticles exhibited concentrated fluorescence in the brain parenchyma, particularly near the BBB. The BBB restricts passive diffusion of most molecules; however, the enhanced retention of Cy5.5-M-R@siC-NPs in brain tissues suggests active transport mechanisms. The unique lipid composition and surface modifications of these nanoparticles likely enable receptor-mediated interactions with endothelial cells lining the BBB, facilitating transcytosis and sustained cerebral retention (Fig. 11).
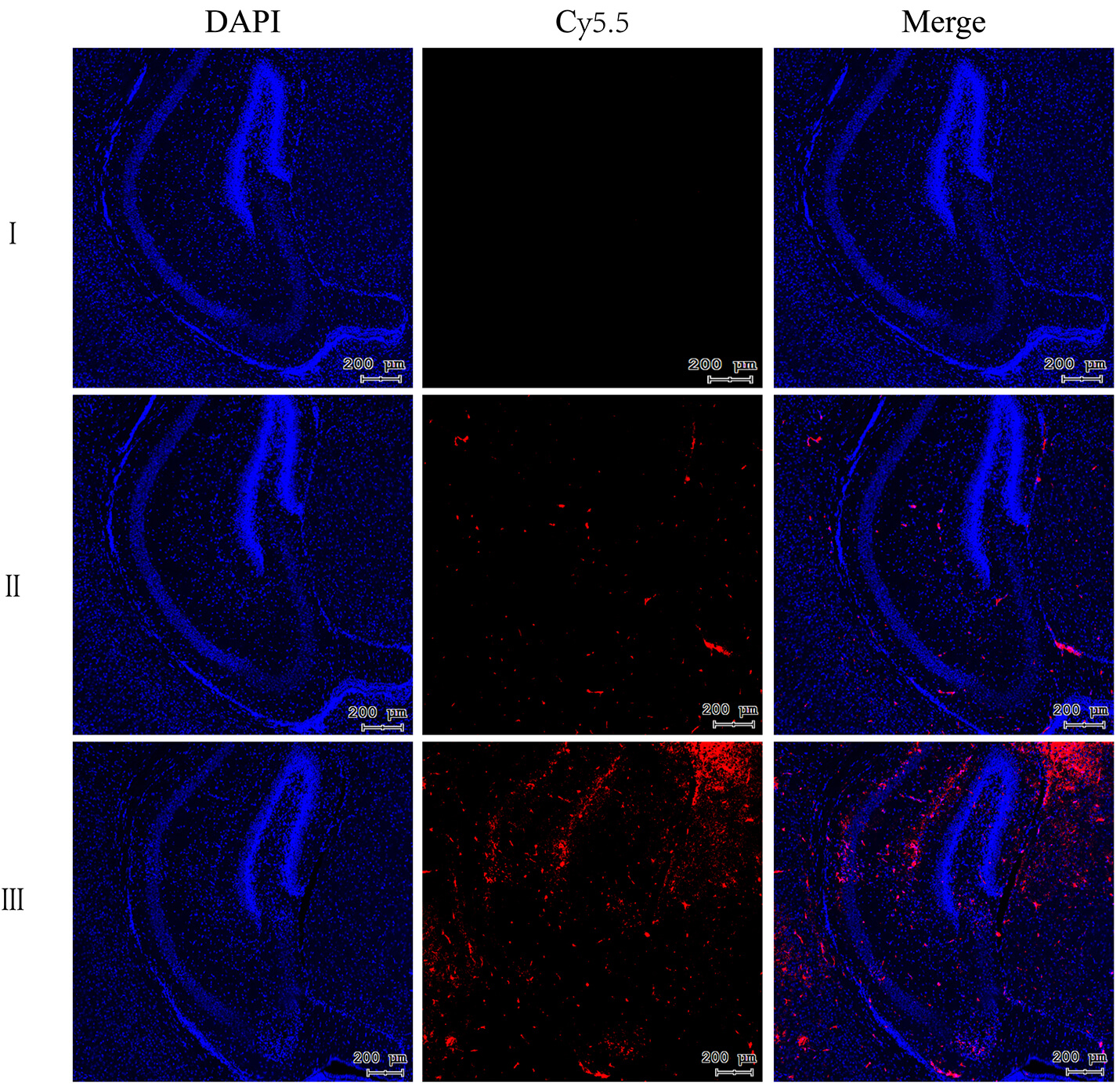 Fig. 11.
Fig. 11.
Fluorescence intensity of Cy5 (red) in the brains of mice in different treatment groups. Scale bar = 200 µm. DAPI (blue) served as nuclear counterstaining. I: PBS-treated group; II: Cy5-siC-NPs-treated group; III: Cy5-M-R@siC-NPs-treated group.
Primary astrocytes were successfully isolated and purified from the cerebral
tissues of C57BL/6 neonatal mice. Immunofluorescence staining confirmed astrocyte
identity: GFAP-labeled astrocytic processes (green, Fig. 12A-I),
phalloidin-stained F-actin cytoskeleton (red, Fig. 12A-II), and DAPI-stained
nuclei (blue, Fig. 12A-III). Merged images (Fig. 12A-IV) revealed that primary
astrocytes exhibited characteristic star-shaped or polygonal morphology with
large cell bodies, prominent nuclei, and multiple filopodia-like processes in
culture. Quantitative analysis demonstrated that
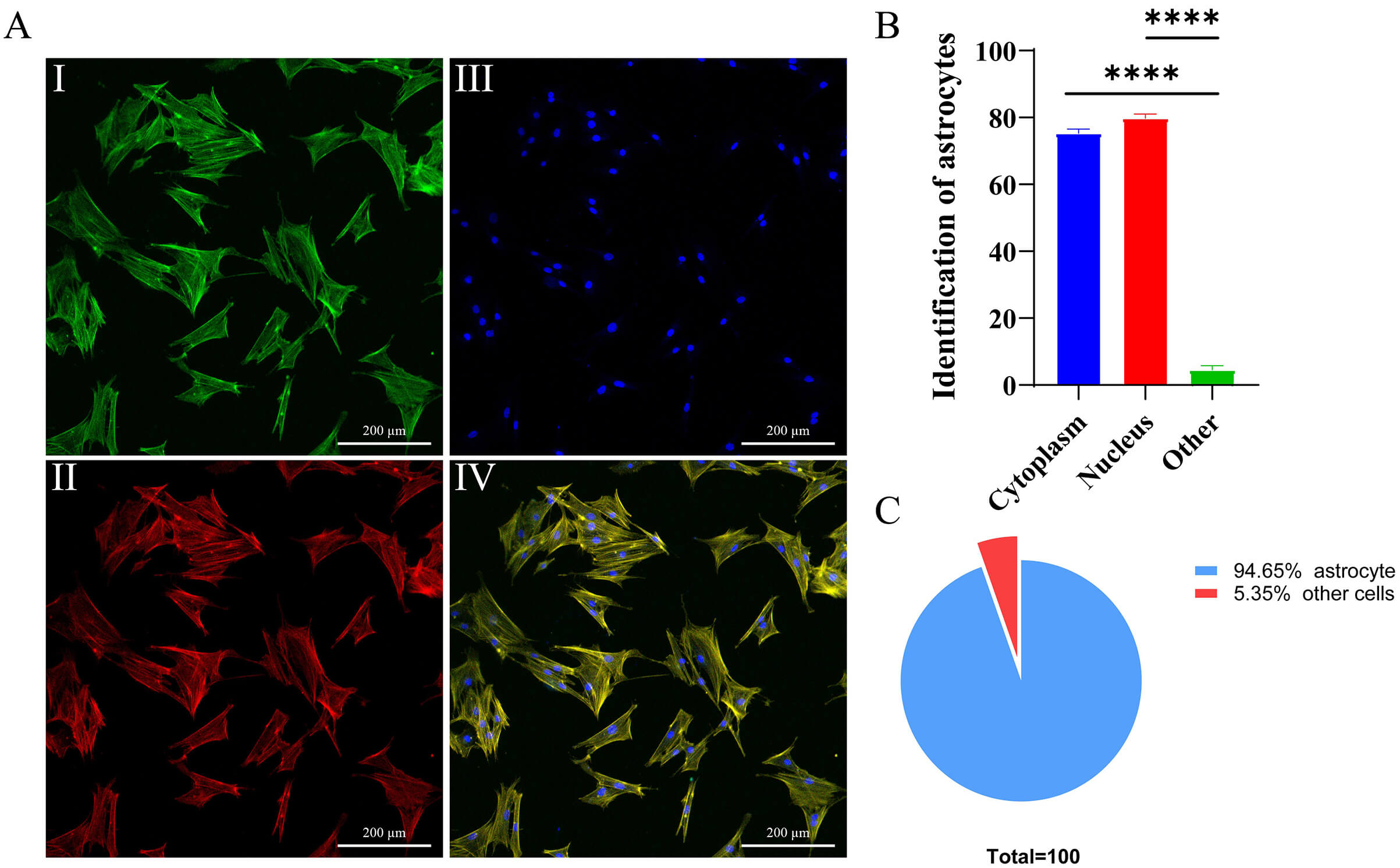 Fig. 12.
Fig. 12.
Morphological characterization and purity analysis of primary
astrocytes. (A) Immunofluorescence analysis: A-I: GFAP (astrocyte marker,
green); A-II: phalloidin-labeled cytoskeleton (red); A-III: DAPI-stained nuclei
(blue); A-IV: merged tri-channel image. (B) Antibodies labeled with GFAP bind to
specific proteins in the cytoplasm (not the nucleus) of primary astrocytes, and
the proportion of positive cells is observed under a fluorescence microscope, ****p
To assess the therapeutic potential of M-R@siC-NPs, primary astrocytes were
subjected to OGD treatment. The experimental groups included NC, OGD (model),
siC-NPs-treated, and M-R@siC-NPs-treated groups. The outcomes were evaluated
based on GFAP expression, IL-1
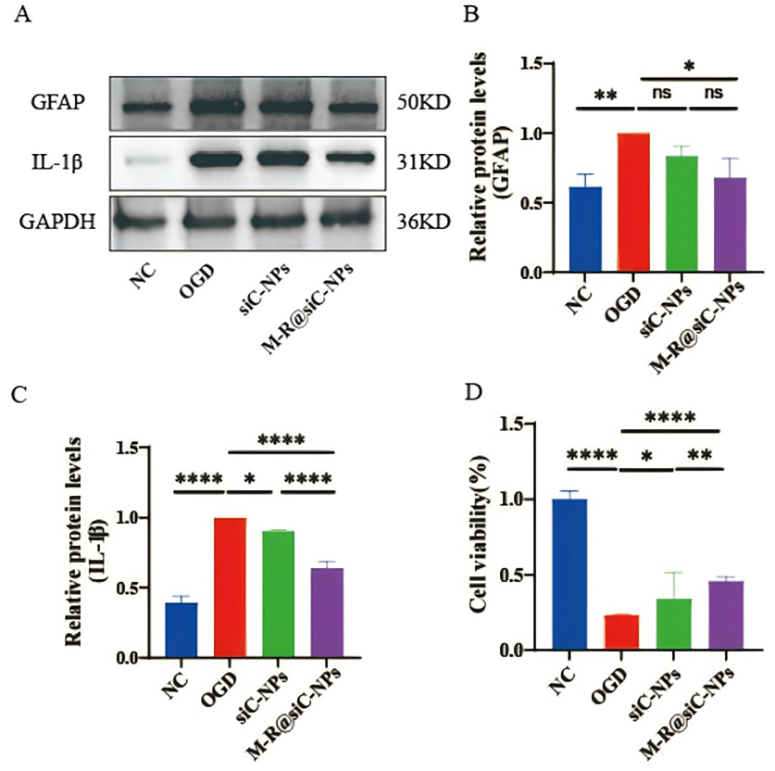 Fig. 13.
Fig. 13.
In vitro expression of GFAP and interleukin-1
In the OGD model of primary astrocytes, Western blot results demonstrated that
the OGD model group exhibited significantly increased protein levels of GFAP and
IL-1
These findings demonstrate that M-R@siC-NPs can alleviate astrocyte activation and neuroinflammation via targeted gene silencing, while promoting the functional recovery of damaged cells.
The murine HIBD model was established using the Rice-Vannucci method [53]. Cortical blood flow was quantified using laser speckle flowmetry at 24 hours post-surgery. Compared with those in the sham-operated group, HIBD mice exhibited a statistically significant reduction in left cortical blood flow (Fig. 14A,B), confirming successful model induction.
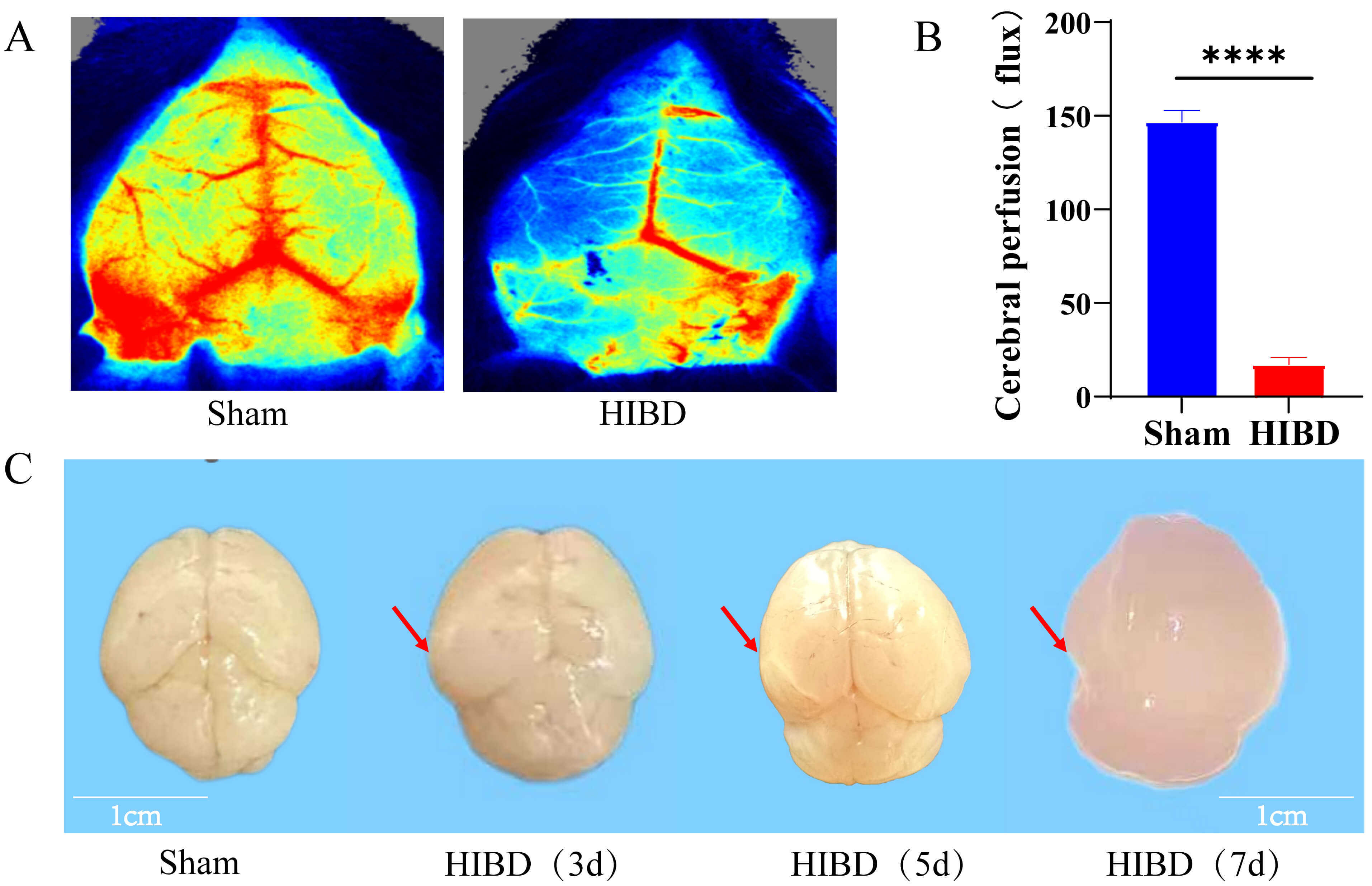 Fig. 14.
Fig. 14.
Cerebral blood flow perfusion and pathological morphological
changes in HIBD model mice. (A) Comparative laser speckle flowmetric images of
cerebral blood flow in sham-operated and HIBD groups. (B) Quantitative analysis
of left cerebral blood flow perfusion. (C) Macroscopic observations of HIBD brain
tissues at 3, 5, and 7 days post-surgery (arrows indicate: 3 d edema, 5 d
infarction, 7 d tissue softening/loss). Scale bar = 1 cm. Data are presented as mean
Post-mortem examination of brain tissues revealed progressive pathological changes. Sham-operated mice displayed normal brain morphology, characterized by a uniform texture, smooth surfaces, and an intact structure. In contrast, the brains of HIBD mice exhibited time-dependent deterioration, including mild cerebral edema at 3 days post-HIBD (indicative of ischemia–hypoxia injury), ipsilateral infarction foci at 5 days (reflecting vascular occlusion and neuronal necrosis), and exacerbated tissue damage at 7 days, including softening of the brain parenchyma, regional necrosis, and structural disintegration (Fig. 14C). These findings collectively validate the reliability of the HIBD model for studying hypoxic–ischemic brain injury progression.
To build upon our in vitro findings, the therapeutic efficacy of M-R@siC-NPs was evaluated in a C57BL/6 HIBD model. The mice were divided into four groups: sham-operated, HIBD, siC-NPs-treated, and M-R@siC-NPs-treated. At 5 days post-treatment, brain tissues were subjected to H&E staining. The brain tissues of sham-operated mice exhibited normal cellular morphology with tightly arranged neurons and no injury. The brain tissues of HIBD mice showed pyknotic nuclei, deep staining, and inflammatory cell infiltration. While the brain tissues of siC-NPs-treated mice displayed reduced injury severity, including decreased inflammatory infiltration and improved edema, the brain tissues of M-R@siC-NPs-treated mice demonstrated significant improvement, including an intact brain architecture, minimal neuronal damage, orderly cellular arrangement, and markedly reduced inflammatory cell infiltration (Fig. 15A).
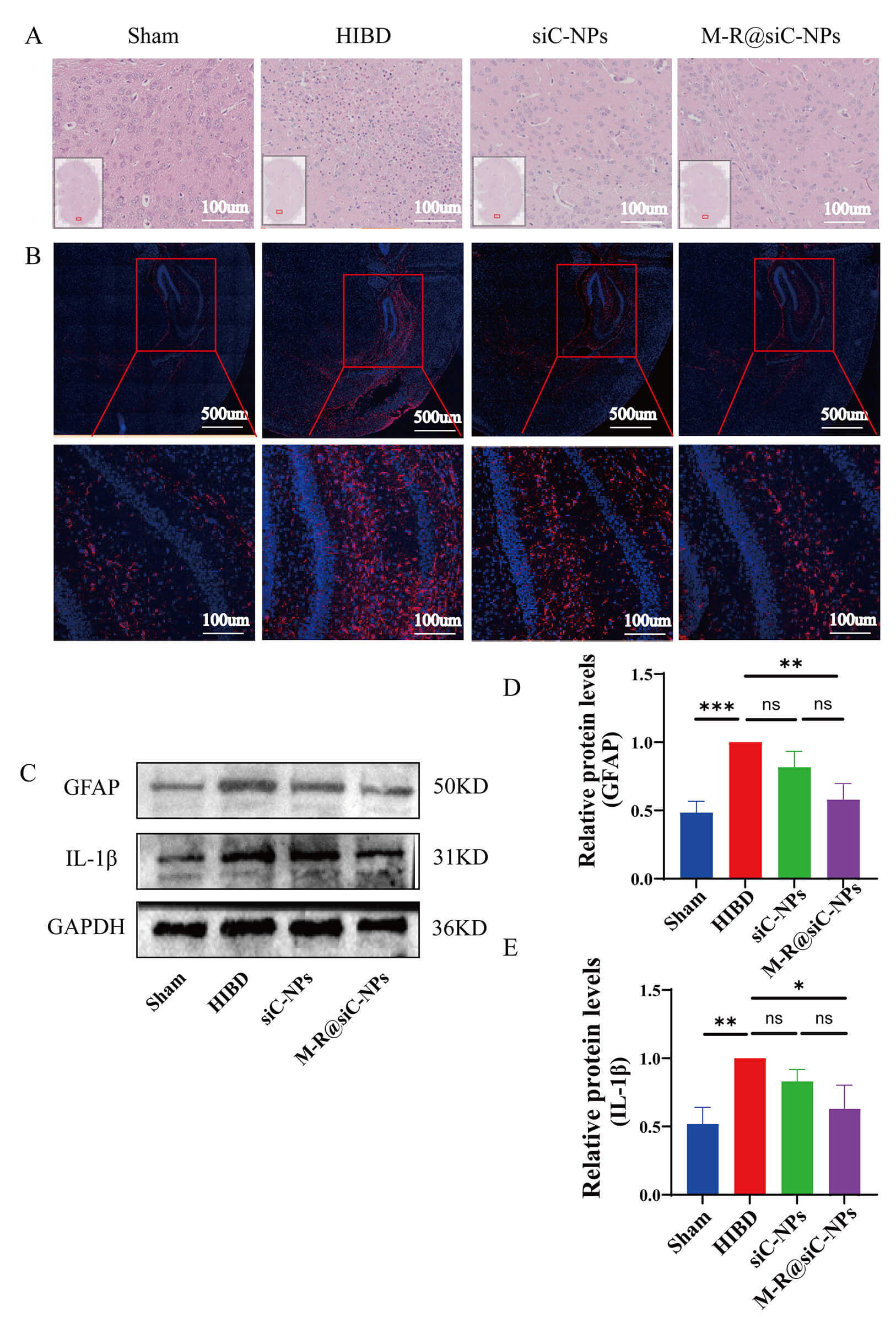 Fig. 15.
Fig. 15.
Histopathological changes and GFAP/IL-1
Immunofluorescence analysis revealed distinct astrocyte activation states across groups (Fig. 15B). Weak GFAP fluorescence in brain tissues from sham-operated mice indicated quiescent astrocytes. In contrast, intense GFAP signals in brain tissues from HIBD mice reflected hypoxia–ischemia-induced astrocyte activation and secondary neuroinflammation. While treatment with siC-NPs reduced the GFAP intensity compared with that in HIBD mice, treatment with M-R@siC-NPs achieved the most pronounced suppression, with significantly weaker GFAP fluorescence than in both the HIBD and siC-NPs-treated groups, underscoring the superior efficacy of targeted nanoparticles for inhibiting astrocyte activation.
Western blot analysis confirmed these trends in an astrocyte activation marker
(GFAP) and an inflammatory cytokine (IL-1
The righting reflex test evaluates sensorimotor development by measuring the latency to rightward rotation after placement in a supine position. Sham-operated mice exhibited rapid righting reflexes with significantly shorter latency times compared with HIBD mice (Fig. 16A), which showed prolonged latencies, indicating impaired neuromuscular control and balance due to neurological injury. Treatment with siC-NPs and M-R@siC-NPs reduced latency times, with the M-R@siC-NPs-treated group demonstrating the most marked improvement.
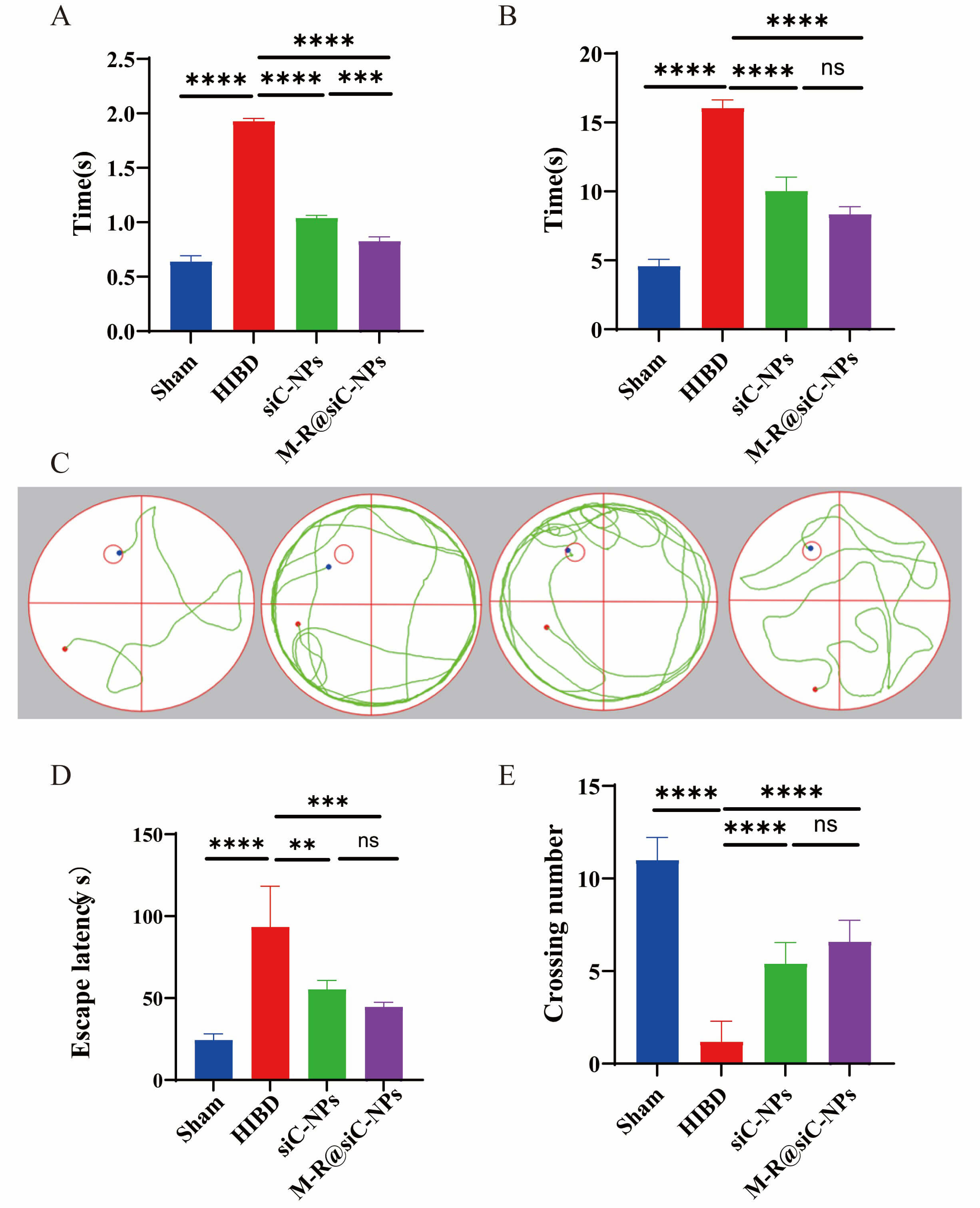 Fig. 16.
Fig. 16.
Therapeutic effects of M-R@siC-NPs on behavioral and cognitive
functions in HIBD model mice. (A) Righting reflex test: M-R@siC-NPs treatment
significantly shortened latency compared with that in the HIBD model group
(p
Using the negative geotaxis test, the mice in each group were assessed for their ability to climb upward on an inclined plane. The HIBD mice displayed significantly longer escape latencies than the sham-operated control mice (Fig. 16B), reflecting compromised neuromuscular strength and coordination. While both treatment groups showed improved latencies compared with the HIBD mice, treatment with M-R@siC-NPs led to statistically significant recovery, suggesting enhanced motor function restoration.
To evaluate spatial learning and memory recovery, the Morris water maze test was performed over 5 days (Fig. 16C). During training (days 1–4), sham-operated mice exhibited progressively shorter escape latencies, indicating efficient spatial memory formation. In contrast, the HIBD mice displayed prolonged latencies (Fig. 16D) and disorganized swimming paths, reflecting hippocampal dysfunction. Treatment with siC-NPs or M-R@siC-NPs reduced latency times and increased platform-crossing frequency. Notably, M-R@siC-NPs-treated mice showed the greatest improvements in learning and memory retention during the probe trial (day 5, Fig. 16E).
In the mouse HIBD model, the siC-NPs group exhibited significantly less improvement in brain pathological lesions and neurological function recovery compared to the M-R@siC-NPs group. Combined with in vivo fluorescence imaging results—where the cerebral accumulation of M-R@siC-NPs was 3 times that of siC-NPs—it was further confirmed that the BBB penetration capability and astrocyte-targeting ability, mediated by M-PEG-lipid and DSPE-PEG2000-RVG29, are the key factors underlying the superior in vivo therapeutic efficacy of the targeted nanoparticles. Moreover, the presence of siC-NPs clearly verifies that this efficacy advantage originates from the targeted modification, rather than the carrier or siRNA itself.
Current HIE therapies are associated with multiple limitations. For example, the gold standard approach, hypothermia, can only be employed in neonates within 6 h post-birth, and its use is still associated with 44–53% mortality/severe disability rates and infection risks [7, 54, 55, 56, 57, 58]. While experimental drugs (allopurinol, melatonin) have shown promise, long-term safety data for these therapeutics are lacking [59, 60, 61]. Stem cell therapies encounter ethical/immunological barriers [62, 63]. This study successfully constructed brain-targeting LNPs (M-R@siC-NPs) for astrocyte-specific delivery of circHIPK2 siRNA, which exhibited significant therapeutic efficacy in a murine HIBD model. The dual-targeting nanoplatform efficiently traversed the BBB, silenced circHIPK2 expression, inhibited astrocyte activation, and attenuated neuroinflammation, providing a novel strategy for neonatal HIE therapy (Fig. 17). In contrast with existing therapies, M-R@siC-NPs offer superior targeting, efficacy, and safety. Compared with glioma-targeted liposomes specifically, M-R@siC-NPs exhibit greater siRNA encapsulation and brain accumulation (RVG29-mediated hippocampal targeting), minimizing systemic toxicity.
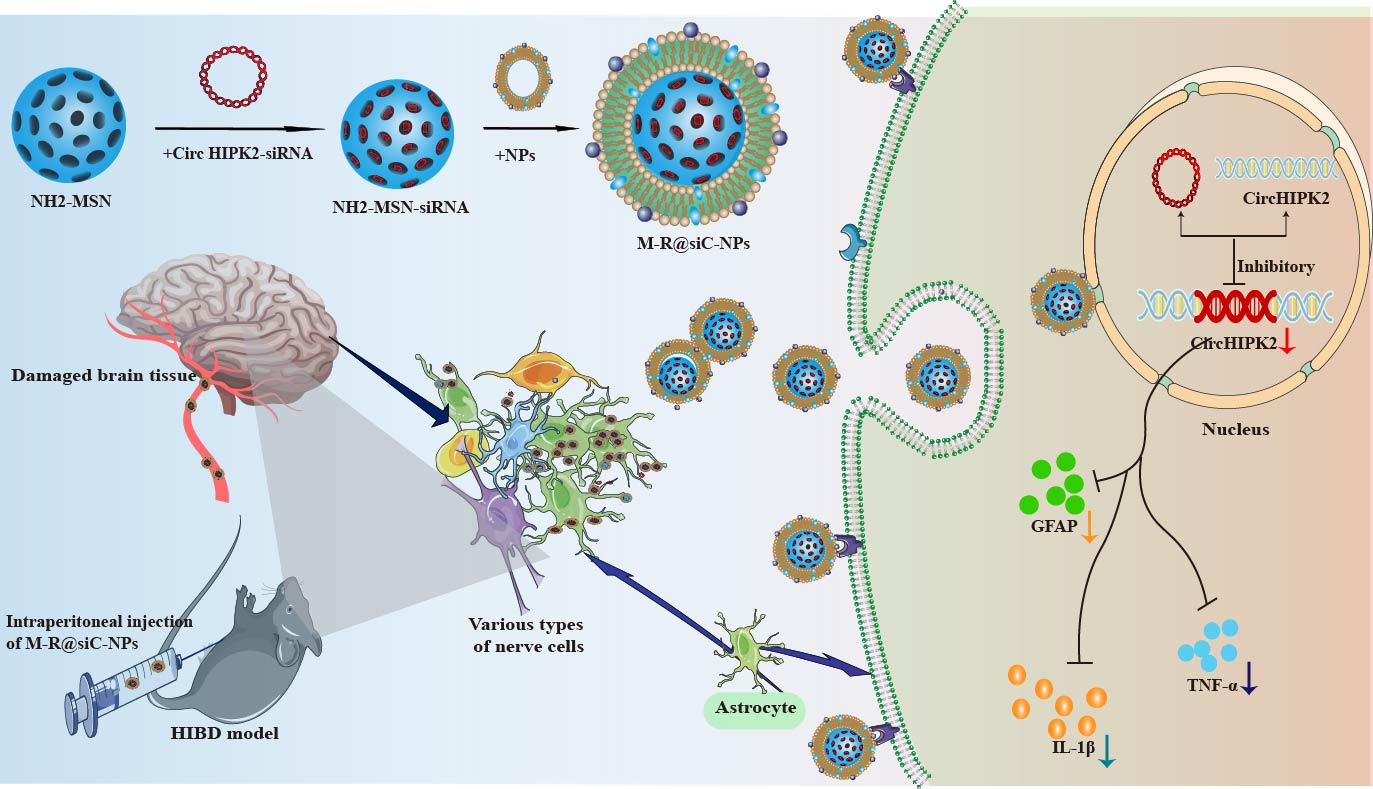 Fig. 17.
Fig. 17.
Schematic illustration of the molecular mechanism by which
M-R@siC-NPs alleviate HIBD through a circHIPK2 silencing-mediated
reduction of astrocyte activation. TNF-
Neuroinflammation is central to ischemia–reperfusion injury in HIE. During the
pathogenesis of HIE, astrocytes undergo aberrant activation, characterized by
upregulated GFAP expression and the release of pro-inflammatory mediators (e.g.,
TNF-
The BBB poses a critical barrier to CNS drug delivery, limiting cerebral bioavailability while increasing systemic toxicity [65, 66]. Although RNA interference holds promise, the application of siRNA as therapeutic agents has faced limitations including poor membrane permeability, rapid degradation, mononuclear phagocyte system clearance, renal excretion, and off-target effects [67, 68]. To address these limitations, we developed M-R@siC-NPs incorporating MSNs. MSNs exhibit exceptional drug-loading capacity due to their ordered mesopores, high surface area, and modifiable silanol groups [34, 69, 70, 71, 72, 73, 74]. In this study, positively charged NH2-MSNs achieved high siRNA loading via electrostatic interactions with negatively charged siRNA.
Liposomes, with their amphiphilic lipid bilayer structure, enable co-delivery of hydrophilic/hydrophobic agents while enhancing BBB penetration [75, 76]. Targeted modifications using ligands (e.g., polymers, peptides) further improve brain delivery [38, 77]. M-R@siC-NPs were functionalized with a Glucose Transporter 1 (GLUT1)-targeting M-PEG-lipid and the BBB-penetrating DSPE-PEG2000-RVG29. RVG29 mediates BBB transcytosis via receptor recognition, while M-PEG-lipid directs astrocyte targeting, conferring superior specificity compared with non-targeted siC-NPs.
Cellular uptake of nanoparticles is influenced by the physicochemical properties
of the nanoparticles, such as surface charge and size [78, 79, 80, 81]. While sub-100 nm
particles are better able to penetrate the BBB, 100–150-nm particles exhibit
prolonged circulation. However, even larger particles (
The assessment of “nanorisk” requires a comprehensive evaluation of
nanoparticle trafficking dynamics, organ-specific accumulation patterns, and
toxicity mechanisms in biological systems [90]. Safety assessments in the present
study revealed that after treatment with M-R@siC-NPs at concentrations of 20–80
µg/mL,
Activated astrocytes critically regulate sterile inflammation and IL-1
Currently, the treatment of HIBD primarily relies on two major strategies:
neuronal protection and broad-spectrum anti-inflammation, both of which have
significant limitations. On one hand, neurons exhibit extremely poor tolerance to
hypoxia-ischemia—intracellular ATP depletion exceeds 80% within 2 hours of
ischemia. Traditional neurotrophic factors such as brain-derived neurotrophic
factor (BDNF) can only delay neuronal apoptosis by approximately 15%–20% due
to restricted BBB permeability and downregulation of target receptors. Although
antioxidants like N-acetylcysteine can temporarily reduce oxidative stress, they
fail to block cascading necrosis via the mitochondrial pathway; clinical
translation trials have shown that their 90-day neurological function improvement
rate is only 5.7% [97, 98, 99]. On the other hand, while broad-spectrum
anti-inflammatory strategies can inhibit early pro-inflammatory factors
IL-1
As core regulators of CNS homeostasis, astrocytes play a crucial role in HIBD.
They clear over 80% of glutamate in the synaptic cleft by highly expressing
glutamine synthetase (GS), thereby inhibiting excitotoxicity; secrete IL-6 and
C-X-C motif chemokine ligand 12 (CXCL12) to regulate neuroinflammation; and
maintain BBB integrity through the modulation of vascular endothelial growth
factor (VEGF) and TGF-
In this study, a novel mannose/RVG29 peptide dual-modified lipid nanoparticle M-R@siC-NPs was innovatively constructed to overcome the bottlenecks of traditional interventions. Leveraging the high expression of mannose receptors (MR) on the surface of astrocytes, the mannose modification enables specific endocytosis of the nanoparticles; combined with the RVG29 peptide, which enhances brain penetration capability, this system can efficiently deliver circHIPK2 siRNA into astrocytes. M-R@siC-NPs demonstrate great potential in inhibiting astrocyte activation, exerting anti-inflammatory effects, and alleviating HIBD-induced brain damage, holding promise as an innovative strategy for the treatment of HIE.
The present study has some limitations that must be considered and addressed in future research. The sample size in this study was small, and the long-term biodistribution and toxicity of M-R@siC-NPs were not evaluated. Moreover, the mechanisms by which M-R@siC-NPs interact with the BBB remain unclear. Future work should include expanded cohorts, assessment of scale-up feasibility, proteomics analyses for pathway elucidation, and in vivo imaging to visualize BBB traversal.
In conclusion, this study successfully developed M-R@siC-NPs with dual functions based on the inclusion of RVG29 peptide and mannose-PEG-lipid, offering good BBB penetration and astrocyte-targeting efficiency. These nanoparticles achieved efficient circHIPK2 silencing while exhibiting excellent stability and biosafety. Mechanistically, the results of this study indicate that M-R@siC-NPs reduce astrocyte activation and mitigate neuroinflammation, effectively ameliorating brain injury in HIBD model mice. This work not only establishes a novel RNA interference-based targeted therapy for neuroinflammation but also provides a foundation for the clinical translation of circHIPK2 as a therapeutic target in HIE. Overall, M-R@siC-NPs represent a breakthrough in HIE therapy, combining BBB penetration, astrocyte specificity, and robust siRNA delivery. This dual-targeted system overcomes multiple limitations of conventional treatments, demonstrating transformative potential for precision gene therapy in neonatal brain injury.
WB, Western blot; qRT-PCR, Quantitative reverse transcription polymerase chain reaction; CCK-8, Cell counting kit-8; CREA, Creatinine; CNS, Central nervous system; PEG, Polyethylene glycol; BBB, Blood–brain barrier; GLUT1, Glucose transporter 1; HIBD, Hypoxic–ischemic brain damage; HIE, Hypoxic–ischemic encephalopathy; GFAP, Glial fibrillary acidic protein; NP, Nanoparticle; OGD, Oxygen-glucose deprivation; PBS, Phosphate-buffered saline; PLT, Platelet; ALB, Albumin; ALT, Alanine transaminase; AST, Aspartate transaminase; RVG, Rabies virus glycoprotein; siRNA, Small interfering RNA; IL-1
The data that support the findings of this study are available from the corresponding author upon reasonable request.
YXD, FHS, SXW, YTZ and XL designed the research study. YXD, DYQ, DF and YJS performed the research. ZHC and RCM provided help and advice on the WB experiments. DYQ, DF and YJS provided help and advice on the H&E experiments. YXD and FHS analyzed the data. YTZ and FW demonstrated outstanding efforts in critical revision of the article and grant preparation. ZHC, RCM, DYQ, DF, YJS, and FW have actively participated in the conception or design of the research, as well as the acquisition, analysis, and interpretation of the study data. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work. All authors have read and approved the final manuscript for publication.
All animal experiments were approved by the Ethics Committee of the Second Hospital of Lanzhou University (Approval No. D2024-439) and were performed in strict accordance with the Regulations on the Administration of Laboratory Animals and the international standards for the care and use of laboratory animals established by AAALAC.
We would like to thank the Second Hospital of Lanzhou University for providing the experimental site for this study. Additionally, we appreciate Medjaden, Inc. for offering scientific editing services for this manuscript.
This research received no external funding.
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/JIN45212.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.