1. Introduction
Among age-related neurodegenerative disorders, Parkinson’s disease (PD) stands
out as the second-highest occurring disorder. The distinctive clinicopathological
features of PD involve the gradual and specific loss of dopaminergic (DA-ergic)
neurons in the midbrain and the substantia nigra pars compacta (SNpc) regions of
the brain. Additionally, a notable characteristic is the pathological buildup of
-synuclein (-syn) aggregates [1]. Consequently, the dorsal
striatum and other target areas have a dopamine deficiency, which causes chief
motor dysregulations such as bradykinesia, stiffness, and tremors. Furthermore,
non-motor manifestations, like depression and sleep disturbances in PD patients,
are also evident [1].
Numerous studies have revealed that the brains of PD patients and animal models
with synucleinopathies contain an abnormal accumulation of
P-Ser129--syn, a pathologically active phosphorylated form of
-syn [2, 3, 4, 5]. In healthy brains, -syn undergoes minimal
phosphorylation, while there are dramatically elevated
P-Ser129--syn levels during PD pathology in brains afflicted with
Lewy pathology [2, 6]. This suggests a potential correlation between this
post-translational modification and heightened deposition of -syn,
coinciding with Lewy body formation and the beginning of the neurodegenerative
process. Current strategies, including treatments inhibiting -syn’s
aberrant aggregation, have generated significant interest as potential
interventions that could potentially slow or arrest the advancement of the
pathological condition.
The 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced mouse model is
widely used due to its ability to replicate several key features of PD, including
DA-ergic neuron degeneration and motor deficits. This model is well-characterized
and allows for the study of neurodegenerative processes and potential therapeutic
interventions. While specific -synucleinopathy models, such as
transgenic mice expressing human -syn, provide insights into the role
of -syn aggregation in PD, the MPTP model remains valuable for its
simplicity, reproducibility, and relevance to DA-ergic system degeneration [7, 8]. We
selected the MPTP model for its established utility in investigating
neurodegeneration and evaluating neuroprotective strategies, which are critical
aspects of PD research.
Oxidative stress mediates vital contributions in PD pathogenesis while
activating glycogen synthase kinase-3 beta (GSK-3). Increased
GSK-3 leads to -syn toxicity and the dysregulation of the
Wnt/-catenin signaling mechanism. The canonical Wnt/-catenin
signaling process, essential for embryonic developmental and tissue homeostasis
processes, also controls neuronal function in the central nervous system (CNS)
[9]. This pathway is an intriguing candidate that appears to be dysregulated in
PD and has been linked to several neuropathologies [10]. Research suggests
activating this pathway could restore neurogenesis and enhance brain self-repair
capacity [11], making it a promising therapeutic candidate for countering
neurodegenerative conditions. Furthermore, the pathogenesis of PD is associated
with the activity of GSK-3, a key regulator of the Wnt/-catenin
signaling pathway. GSK-3 restricts -catenin’s cytoplasmic
stabilization and, hence, nuclear migration, effectively preventing the
conventional Wnt pathway activation [12]. GSK-3 inhibition has been shown
to promote -catenin’s stabilization, thus promoting Wnt
signaling-mediated neuroprotective benefits in PD models [13]. Elevated expression
of GSK-3 in the nigral neurons of post-mortem PD brains and its aberrant
phosphorylation at the Tyrosine 216 (Tyr216) is also evident in the striatum of individuals with
PD [14, 15]. GSK-3 activity is strictly controlled through the negative
regulation occurring via GSK-3 Serine-9 phosphorylation at the
N-terminal region [16]. Pharmacological agents inhibiting GSK-3activation may mitigate neurodegeneration in PD [17].
While neuronal autophagy is widely recognized as a defensive and beneficial
mechanism for the physiological functioning of the nervous system, its
paradoxical involvement in neuronal cell death has become increasingly evident.
Numerous studies employing environmental toxins, hereditary PD-risk genes, and
postmortem PD brain samples have illuminated the contributions of autophagy in
instigating PD-associated DA-ergic neuronal loss [18, 19]. Particularly noteworthy is the
identification of autophagic-regulated loss of nigral neurons in PD patients
[20]. Oxidative stress induced by MPTP or dopamine toxicity has been demonstrated
to amplify the formation of autophagic vacuoles, intensifying autophagic activity
and subsequently leading to neuronal cell death [21, 22]. This dual role of
autophagy, serving both neuroprotection and contributing to neurodegeneration,
underscores its multifaceted involvement within the intricate framework of
neuronal processes.
The drug targeting strategies in PD treatment have limited efficacies and are
often associated with side effects. Phytochemicals present alternative, more
efficacious, and safer treatment strategies against PD pathology. Owing to its
diverse neuropharmacological characteristics, ranging from anti-oxidative to
anti-inflammatory, the phenolic aldehyde vanillin exhibits promise against the
pathophysiology of PD. The aim of this study was to determine if vanillin
inactivates GSK-3, thus potentiating the Wnt/-catenin
signaling in MPTP-induced nigrostriatal degeneration while protecting the
DA-ergic system. By examining critical pathways implicated in PD pathogenesis,
such as increased -syn expression, autophagic-neuronal loss, and
Wnt/-catenin/GSK-3 activity, we seek to understand the
underlying mechanism of therapeutic potential of Vanillin in PD.
2. Materials and Methods
2.1 Ethical Statement Regarding Animal Usage
All animal handling and experimental procedures in this study followed standard
techniques and protocols. The experiments involving C57BL/6 mice were conducted
in accordance with the protocols approved by the Institutional Animal Ethics
Committee (IAEC) of Jawaharlal Nehru University (JNU), New Delhi (approval code:
10/GO/ReBi/99/CPCSEA/March 10, 1999). Every effort was made to minimize animal
suffering and ensure their well-being throughout the study. We have followed
ARRIVE guidelines 2.0.
2.2 Experimental Animals and Dietary Regimen
65 animals Male Mice (C57BL/6) were used in the present study. Male C57BL/6 mice, weighing 25–30 grams and aged 2.5–3 months were raised and
housed under controlled laboratory conditions. The mice were provided free access
to a standard mouse pellet diet and filtered water ad libitum throughout the
study. The housing environment maintained a temperature of 21 2
°C, a humidity level of 55%, and followed a continuous 12-hour
light/dark cycle. All animal-related procedures were conducted in compliance with
the regulations of Central Laboratory Animal Resource (CLAR) at Jawaharlal Nehru
University (JNU), New Delhi, India. To ensure proper adaptation, the mice
underwent a one-week acclimatization period prior to the commencement of the
experimental phase. Furthermore, before the start of the experiment, the mice
were appropriately trained on all behavioural equipment to ensure optimal
performance and accurate results.
2.3 Experimental Design and Treatment Groups
Adult male C57BL/6 mice were randomly assigned to five groups:
Control Group: Received normal saline (i.p.) as a vehicle.
MPTP Group: Received intraperitoneal (i.p.) injections of MPTP (79F3538,
Sigma-Aldrich, St. Louis, MO, USA) (20 mg/kg) once a day for 5 consecutive days
[23].
Vanillin Group: Vanillin (BCBV5242, Sigma-Aldrich, St. Louis, MO, USA) (60 mg/kg
body weight, p.o.) was administered daily for 21 days, dissolved in normal
saline.
MPTP+Vanillin Group: Mice were subjected to MPTP once every 24 hours for five
consecutive days via intraperitoneal administration, followed by daily oral
administration of Vanillin for 21 days at a dose of 60 mg/kg body weight.
MPTP+L-DOPA (Levodopa/3,4-Dihydroxy-L-phenylalanine) Group: Mice were subjected
to MPTP once every 24 hours for five consecutive days via intraperitoneal
administration, followed by daily oral administration of L-DOPA (PHR1217,
Sigma-Aldrich, St. Louis, MO, USA) for 21 days at a dose of 5 mg/kg body weight.
The assignment of mice to the respective groups was randomized to minimize bias
and ensure a balanced distribution of characteristics among the experimental
cohorts.
2.4 Evaluation of Motor Behaviour
2.4.1 Forced Swim Test (FST)
The Forced Swim Test (FST) was conducted using a transparent glass cylinder with
dimensions of 25 cm in height and 13 cm in diameter [24]. The cylinder was filled
with water to a depth of approximately 18 cm, maintained at a temperature of 22
°C. Each mouse was individually placed in the cylinder, and the swimming
behaviour was systematically video recorded and analyzed by an unbiased observer
unaware of the experimental groups. The behaviour was recorded for a duration of
6 minutes. The immobility time (defined as the duration during which an animal
spends floating with minimal movements to keep its head above the surface of the
water) of the mouse during the final 4 minutes of the test was manually recorded.
2.4.2 Cylindrical Test
Animals were placed in a glass cylinder that was 30 cm high and 20 cm inside
[25]. The mice were left undisturbed in the cylinder for a period of 5 minutes,
during which their behaviour was recorded. An observer who was unaware of the
experimental group counted the instances in which each forepaw was utilized to
start weight-shifting motions, such as landing and wall contact.
2.4.3 Wire Hanging Test
The test is based on the concept that neuronal degeneration in the basal ganglia
circuits leads to motor impairments [26]. The hanging wire test is used to assess
muscle strength and prehensile reflex, evaluating the animal’s ability to grasp a
stretched horizontal wire with its forepaws and sustain suspension. Mice were put
on a horizontal rod measuring 5 5 mm area, 35 cm long, and positioned
between two poles of 50 cm height. The suspension time, i.e., duration until
mouse drops, was recorded during the test. This parameter is a reliable indicator
for detecting neuromuscular abnormalities associated with motor strength.
2.5 Western Blot
Protein expression was determined as per our previous procedure [27].
50 µg of proteins were fractionated by Sodium Dodecyl
Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) and transferred onto a
Polyvinylidene fluoride (PVDF) membrane (18289A10, Bio-Rad, Boston, MA, USA). The blots were blocked and incubated
with the primary antibodies at 4 °C overnight. The next
day, the blots were incubated with Horseradish peroxidase (HRP)-tagged
anti-rabbit secondary antibodies (1:2000; catalogue number 7074S, CST, Danvers, MA, USA) for 2
hours at room temperature. The blots were then developed on an X-ray film in the
darkroom using a luminol-based enhanced chemiluminescence (ECL) substrate. The
blots were then scanned, and density was measured using ImageJ 1.8.0 software
(LOCI, University of Wisconsin, Madison, WI, USA). The expression of the proteins
was normalized against the expression of housekeeping protein -actin.
The list of primary antibodies used for immunoblotting is given in Table 1. The original figures of Western blot can be found in the
Supplementary Materials.
Table 1.
List of antibodies used in Western blotting.
| Antibody |
Host |
Product details |
Dilution |
| -catenin |
Rabbit |
CST, Danvers, MA, USA, Cat # 6B3 |
1:1000 |
| Glycogen synthase kinase-3 beta (GSK-3) |
Rabbit |
CST, Danvers, MA, USA, Cat # 27C10 |
1:1000 |
| P-Ser9-GSK-3 |
Rabbit |
Affinity Biosciences, Cincinnati, OH, USA, Cat # AF2016 |
1:250 |
| Wnt-3a |
Rabbit |
GeneTex, Irvine, CA, USA, Cat #GTX64367 |
1:500 |
| P-Ser129--synuclein (-syn) |
Rabbit |
Affinity Biosciences, Cincinnati, OH, USA, Cat # AF3285 |
1:1000 |
| Microtubule-associated protein 1A/1B-light chain 3 (LC3) |
Rabbit |
Invitrogen, Rockford, IL, USA, Cat # PA1-16931 |
1:1000 |
| p62/Sequestosome (SQSTM1) |
Rabbit |
Invitrogen, Rockford, IL, USA, Cat # PA5-20839 |
1:1000 |
| -actin |
Rabbit |
Abcam, Waltham, MA, USA, Cat # ab8227 |
1:10,000 |
2.6 Immunostaining
Mice were anaesthetized with thiopentone sodium (173277, Neon Laboratories Ltd.,
Mumbai, Maharashtra, India) (150 mg/kg body weight, i.p.). After that, the mice
were transcardially perfused with normal saline, followed by a 4%
paraformaldehyde (PFA) (00547007-1, Thermo Fisher Scientific, Mumbai,
Maharashtra, India) dissolved in phosphate-buffered saline (PBS). Tissues were
cut into 20 µm thick slices with a cryotome, mounted on 1% gelatin-coated
glass slides, and immunostained. Sections were blocked in a humid chamber for 1 h
with 5% bovine serum albumin (BSA) (3534981, SRL Pvt. Ltd., Taloja, Maharashtra,
India) and then incubated with primary antibodies overnight at 4 ℃. The next day,
tissue sections were rinsed with PBS and incubated with a secondary antibody at
room temperature for 2 h. After that, they were counterstained with
4′,6-diamidino-2-phenylindole (DAPI) (Batch No 034M4031V, Sigma-Aldrich, Burlington, MA, USA) for 30 min. For immunostaining, tissue
sections were mounted with fluorescent mounting media and visualised under a
confocal microscope (Eclipse Ti-2, Nikon, Melville, NY, USA). The list of antibodies used
for immunostaining is given in Table 2.
Table 2.
List of antibodies used in Immunostaining.
| Antibody |
Host |
Product details |
Dilution |
| P-Ser9-GSK-3 |
Rabbit |
Affinity Biosciences, Cincinnati, OH, USA, Cat # AF2016 |
1:500 |
| P-Ser129--syn |
Rabbit |
Affinity Biosciences, Cincinnati, OH, USA, Cat # AF3285 |
1:500 |
| LC3 |
Rabbit |
Invitrogen, Rockford, IL, USA, Cat # PA1-16931 |
1:1000 |
2.7 Statistical Analysis
Throughout the experiment, an observer blinded to the study design and treatment
condition performed all behavioural scoring, examination of the histopathological
characteristics, and cell counting. GraphPad Prism software version 7 was used to
conduct the statistical analysis (GraphPad Software, Inc., San Diego, CA, USA).
All the data is presented as a mean standard error of the mean (SEM). To
assess the significant differences between the control, MPTP, Vanillin, MPTP +
Vanillin and MPTP+L-DOPA groups, a one-way analysis of variance (ANOVA) was used,
followed by a Tukey test that assumes the data has a normal distribution.
*p 0.05, **p 0.01, and ***p 0.001 were
used to signify significant differences when compared to the control, and
#p 0.05, ##p 0.01, and
###p 0.001 when compared to the MPTP groups.
3. Results
3.1 Vanillin Ameliorates MPTP-Induced Motor Impairments
To determine the potential neuroprotective effects of Vanillin on motor balance
and coordination in the MPTP-induced PD model, we performed the FST, wire
hanging, and cylindrical test at the end of the dosing (Fig. 1). Our FST results
(Fig. 1B,C; F = 213.3) show that the MPTP-intoxicated mice showed more
immobility time (p 0.001) compared to the control mice. On the other
hand, Vanillin (p 0.001), MPTP+Vanillin (p 0.05), and
MPTP+L-DOPA (p 0.01) treated mice showed less immobility compared to
the MPTP-intoxicated mice. Wire hanging test results (Fig. 1D,E; F = 44.29)
reflected a similar trend. MPTP-intoxication resulted in a considerably decreased
latency to fall (p 0.001) compared to the control mice, whereas
Vanillin (p 0.001), MPTP+Vanillin (p 0.01), and
MPTP+L-DOPA (p 0.05) groups showed an increased latency to fall
compared to the MPTP-treated mice. Similarly, cylinder test results (Fig. 1F,G;
F = 48.05) showed that mice treated with MPTP were more static and
exhibited lesser (p 0.01) rearing compared to the control mice. In
contrast, Vanillin (p 0.05), MPTP+Vanillin (p 0.05),
and MPTP+L-DOPA (p 0.05) treated mice were less static and showed
more rearing compared to the mice subjected to MPTP injections. These results
indicate Vanillin attenuated MPTP-induced motor dysfunction in MPTP-lesioned
mice.
 Fig. 1.
Fig. 1.
Effect of Vanillin on behavioral characteristics in
mice with MPTP-induced PD. (A) Schematic representation of MPTP injection,
Vanillin treatment, and timeline of the study. (B) Image representation for FST.
(C) Graphical representation of immobility time (sec.) in mice of all the groups.
(D) Image representation for wire hanging test. (E) Graphical representation of
the latency to fall in sec. in mice of all the groups. (F) Image representation
for cylinder test. (G) Graphical representation of the numbers of rearing between
different groups. Results are presented as mean SEM. Statistical
difference is shown by **p 0.01 and
***p 0.001 when compared to the
control mice and #p 0.05;
##p 0.01 and
###p 0.001 when compared to
the MPTP treated mice. MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; FST,
Forced Swim Test; SEM, standard error of the mean; L-DOPA,
Levodopa/3,4-Dihydroxy-L-phenylalanine; PD, Parkinson’s disease.
3.2 Vanillin Decreases the Pathological P-Ser129--Syn
Protein Expression
The dysregulations in -syn are critical in PD pathogenesis. More
specifically, -syn aggregation is linked to neuronal dysfunction and
degeneration in PD. -syn undergoes several post-translational
modifications. Phosphorylation of the serine 129 residue has been recognized as a
crucial pathological marker for PD. The expression of the pathological form of
-syn, which is phosphorylated, leading to its aggregation in DA-ergic
neurons, was assessed using western blotting and immunohistochemical staining in
the present study (Fig. 2). The western blot results revealed the higher
expression of the P-Ser129--syn in the
MPTP-intoxicated PD mice group in contrast to the vehicle control group (Fig. 2A,B; F = 7.919, p 0.01) whereas Vanillin (p 0.05),
MPTP+Vanillin (p 0.05) and MPTP+L-DOPA (p 0.05) treated
mice showed lower expression of the P-Ser129--syn compared to
MPTP-treated mice group. Furthermore, immunohistochemical staining of
P-Ser129--syn also showed increased immunoreactivity in
MPTP-induced PD mice compared to the vehicle control group (F = 8.634; Fig. 2C,D, p 0.01). Whereas in the Vanillin (p 0.01),
MPTP+Vanillin (p 0.05) and MPTP+L-DOPA (p
0.01) treated group, the immunoreactivity of P-Ser129--syn was
found to be significantly reduced in comparison with the MPTP-treated
group. These findings demonstrate that Vanillin treatment significantly
alleviated the expression of the P-Ser129--syn, indicating that
Vanillin mitigated the aberrant P-Ser129--syn protein expressions
in MPTP-treated mice.
 Fig. 2.
Fig. 2.
Effect of MPTP administration and Vanillin treatment on the
P-Ser129--syn expression in the SNpc of mice brain. (A) The role
of MPTP and Vanillin administration on the P-Ser129--syn levels in
the substantia nigra pars compacta (SNpc) of the mouse brain (B) Bar graphs
depict the comparative expression of P-Ser129--syn (Fold change)
in SNpc of the mice of all the groups. (C) Representative images showing
P-Ser129--syn immunoreactivity in the SNpc of the mice of all the
groups (Scale bar: 20 µm). For visualization, the secondary antibody Goat
anti-rabbit immunoglobulin G (IgG) (Alexa fluor® 488) was used,
followed by counterstaining with 4′,6-diamidino-2-phenylindole (DAPI). (D)
Bar graphs depict the comparative expression of P-Ser129--syn
immunoreactivity between the groups. Values are given in the form of mean
SEM (n = 3), **p 0.01 vs. controls and #p 0.05,
##p 0.01 vs. MPTP group. -syn,
-synuclein; SNpc, substantia nigra pars compacta.
3.3 Vanillin Hinders GSK-3 Activity and Facilitates
P-Ser9-GSK-3 Phosphorylation
We investigated the expression of GSK-3 and P-Ser9-GSK-3
in the SNpc region of mice. Our western blot findings indicated that
MPTP-administered mice had significantly elevated GSK-3 protein
expression compared with the control group (Fig. 3A,B; F = 9.37, p
0.05). Vanillin (p 0.01), MPTP+Vanillin (p 0.05) and
MPTP+L-DOPA (p 0.01) treated mice showed significantly decreased
GSK-3 protein expression compared to the MPTP-treated group. The
enzymatic activity of GSK-3 is modulated by phosphorylation.
Phosphorylation at serine-9 significantly reduces the enzymatic activity of
GSK-3. The results from the western blot also showed a significant
decrease in the protein expression of P-Ser9-GSK-3 (Fig. 3A,C,
p 0.001) in MPTP-exposed mice in contrast with the control group.
The protein expression level of P-Ser9-GSK-3 was substantially
increased in Vanillin (p 0.05), MPTP+Vanillin (p 0.05)
and MPTP+L-DOPA groups compared to the MPTP-intoxicated mice. Similarly, our
immunofluorescence results also showed a significant decrease in the expression
of P-Ser9-GSK-3 (Fig. 3D,E, p 0.001) in
MPTP-intoxicated mice in contrast to the control group. The immunoreactivity of
P-Ser9-GSK-3 was substantially increased in Vanillin (p 0.001), MPTP+Vanillin (p 0.05) and MPTP+L-DOPA (p
0.01) mice in comparison to the mice treated with MPTP. These results indicate
that Vanillin treatment negated the GSK-3 activity and mediated
P-Ser9-GSK-3 phosphorylation in MPTP-treated mice. Fig. 3D
represents representative immunofluorescence images of
P-Ser9-GSK-3.
 Fig. 3.
Fig. 3.
Effect of MPTP administration and Vanillin treatment on the
GSK-3 and P-Ser9-GSK-3 expression in the mice SNpc. (A)
The representative western blot images of GSK-3 and
P-Ser9-GSK-3 protein expression in the mice of all the groups.
(B,C) Bar graphs depict the relative GSK-3 and
P-Ser9-GSK-3 protein expression (Fold change) in the SNpc region of
mice of all the groups. (D) Representative confocal image of
P-Ser9-GSK-3 expression between the mice of all the groups (Scale
bar: 20 µm). For visualization, the secondary antibody Goat anti-rabbit IgG
(Alexa fluor® 488) was used followed by counterstaining with DAPI. (E) Bar graphs depict the relative
fluorescence intensity of P-Ser9-GSK-3 expression in SNpc of mice
of all groups. Values are given as mean SEM (n = 3), ns, non-significant,
*p 0.05, ***p 0.001 vs. control and #p 0.05, ##p 0.01 and ###p 0.001 vs.
MPTP group.
3.4 Vanillin Potentiates the Wnt/-Catenin Pathway in
MPTP-Intoxicated Mice
Aberrations in the Wnt/-catenin signaling pathway have been
demonstrated to have a vital role in the pathophysiology of PD. In this study, we
assessed the expression of Wnt-3a and -catenin in the SNpc area of mice.
Our western blot results revealed that MPTP-lesioned mice showed significantly
decreased protein expression of Wnt-3a (Fig. 4A,B; F = 29.96, p
0.001) compared to the control group. Inversely, Vanillin (p 0.001),
MPTP+Vanillin (p 0.01) and MPTP+L-DOPA (p 0.05) treated
mice showed increased protein expression of Wnt-3a compared to the MPTP-lesioned
mice. -catenin plays a vital role in the regulation of
Wnt/-catenin signaling. The expression of -catenin was also
significantly downregulated in MPTP-intoxicated mice compared to the control
group (Fig. 4A,C; F = 10.98, p 0.001). The protein
expression of -catenin was upregulated in Vanillin
(p 0.001), MPTP+Vanillin (p 0.01) and MPTP+L-DOPA
groups compared to the MPTP-intoxicated mice. These observations
indicate that Vanillin treatment in MPTP-induced mice significantly
upregulated the protein expression of Wnt-3a and -catenin. These data
indicate that the neuroprotective effects of Vanillin in vivo appear to
be mediated through the activation of the Wnt/-catenin signaling
pathway.
 Fig. 4.
Fig. 4.
Effect of MPTP intoxication and treatment with Vanillin on
-Catenin & Wnt-3a protein expression levels. (A) Representative
western blot images of -Catenin & Wnt-3a protein expression in the SNpc
region of mice brain. (B,C) Bar graphs depict the comparative expression of
-Catenin & Wnt-3a protein (Fold change) in SNpc of mice of all the
groups. Data is expressed as mean SEM (n = 3), ns, non-significant,
***p 0.001 vs. controls and #p 0.05,
##p 0.01 and ###p 0.001 vs. MPTP groups.
3.5 Vanillin Decreases Autophagic-Cell Death by Modulating the
LC3Π/Ι and p62 Protein Expression
The only known autophagy protein marker that specifically interacts with
autophagosomes is light chain 3 (LC3-II). Upon exposure to reactive oxygen species (ROS),
microtubule-associated protein 1A-LC3-I is converted to
microtubule-associated protein 1B-LC3-II, which triggers
autophagy. LC3-II indicates the presence of autophagosomes, which help clear the
cell of accumulated proteins and defective organelles. We determined the effect
of MPTP exposure and Vanillin treatment LC3-II expressions by performing western
blot analysis and immunohistochemistry. Our immunofluorescence results show that
MPTP treatment significantly increased LC3 expression and punctate distribution
in the cytoplasm of DA-ergic neurons (Fig. 5A,B; F = 15.87, p
0.001). The number of LC3 puncta was significantly reduced in Vanillin
(p 0.001), MPTP+Vanillin (p 0.05), and MPTP+L-DOPA (p 0.01) groups in contrast to the MPTP
group. Similarly, our western blot result also shows that LC3-II/I ratio
was significantly higher (Fig. 5C,D; F = 11.67, p 0.05) in the
MPTP-treated group in contrast to the control group, suggesting increased
autophagosome formation and thus increased autophagy in DA-ergic neurons. The
LC3-II/I ratio was significantly reduced in Vanillin (p 0.01),
MPTP+Vanillin (p 0.05), and MPTP+L-DOPA (p 0.05) groups
compared to the mice treated with MPTP, indicating reduced autophagic-cell death
in DA-ergic neurons. To validate our observation, we also monitored the
expression of the p62 protein, which acts as a cargo protein autophagy substrate,
so decreased p62 expression may be associated with increased autophagy. Our
western blot findings indicate that the protein expression of p62 was markedly
reduced in the MPTP group (Fig. 5E; F = 27.82, p 0.001) compared
with the control. However, the expression of p62 was higher in Vanillin
(p 0.01), MPTP+Vanillin (p 0.05), and MPTP+L-DOPA
(p 0.001) treated mice than the MPTP exposed mice, indicating
Vanillin treatment modulated the MPTP-induced aberrant p62 proteins expressions.
These data indicate that LC3-II binds to p62 and selectively mediates the
autophagy-dependent degeneration of DA-ergic neurons in the MPTP group, which was
reversed when these mice were treated with Vanillin.
 Fig. 5.
Fig. 5.
Effect of MPTP and Vanillin on the LC3 and p62 expression in the
SNpc region of the mice brains. (A) The representative confocal images of LC3
expression in the mice of all the groups (Scale bar: 20 µm). (B) Bar graph shows the comparative
expression of LC3 immunoreactivity in SNpc region of the mice of all the groups.
(C) Representative western blot images of LC3 Π/Ι protein (Fold
change) and p62 protein in the Control, MPTP, Vanillin, MPTP+Vanillin and
MPTP+L-DOPA treated groups. (D,E) Bar graphs depict the comparative protein
expression of LC3 Π/Ι and p62 protein (Fold change) in SNpc region
of the mice in all the groups. Results are expressed as mean SEM (n = 3),
*p 0.05, **p 0.01, ***p 0.001 and vs.
controls and #p 0.05, ##p 0.01 and
###p 0.001 vs. MPTP treated mice.
4. Discussion
In the present study, we have determined the potential neuroprotective effects
of Vanillin in PD by assessing its effect on motor balance and coordination in
the MPTP-induced PD mice model. Combining results from all three behavioral
tests—the FST, wire hanging, and cylindrical tests allows for a more
comprehensive assessment of the animal’s motor function. In coherence with
previous studies, we found that MPTP administration induces depressive behavior
and impaired motor activity [28]. However, vanillin effectively reversed these
behavioral and motor dysfunctions. A similar motor deficit alleviating effect of
vanillin has been previously reported in a rotenone-induced rat model of PD [29].
PD, which involves the aberrant accumulation of -syn inside the SNpc,
which causes the selective and gradual death of neurons [30]. The
post-translational modification of -syn is a crucial factor in its
aggregation [31]. Phosphorylated -syn, particularly
P-Ser129--syn, is a key hallmark of PD and related
synucleinopathies [31, 32]. Research has demonstrated that the administration of
MPTP leads to elevated levels of P-Ser129--syn in mice SNpc [33].
Consistent with these, our immunofluorescence and western blot results also
showed a significant increase in P-Ser129--syn expression in
MPTP-intoxicated mice. In contrast, Vanillin treatment significantly ameliorated
the P-Ser129--syn expression in MPTP-induced mice.
The Wnt/-catenin signaling is implicated in the pathogenesis of PD,
regulating ventral midbrain precursor development, proliferation, and
differentiation [34, 35, 36]. Dysregulated Wnt/-catenin signaling is reported
to be involved in the development of PD [37, 38, 39]. This signaling is altered by
GSK-3, an inhibitor of this crucial pathway, which phosphorylates and
ubiquitinates -catenin. The inactivation of GSK-3 increases
-catenin levels, potentially offering neuroprotective effects [40].
Reports suggest that activating the Wnt/-catenin may facilitate the
differentiation and regeneration of DA-ergic neurons [41, 42]. This highlights
the potential of targeting Wnt/-catenin signaling components, as well as
GSK-3, which may be an effective therapeutic intervention for PD [34].
Blocking GSK-3 reduced -syn protein expression and,
consequently, prevented cell death in PD disease models [14]. Neuroinflammation,
involving activated astrocytes and microglia, contributes to -syn
fibril formation [43]. GSK-3, an enzyme implicated in this process,
exacerbates neuroinflammation when overactive. A recent study shows that
inhibiting GSK-3 via Ser9 phosphorylation reduces astroglial and
microglial reactivity, lowering pro-inflammatory cytokine levels and ROS, thereby
offering neuroprotection [44]. These findings suggest that GSK-3
inhibitors could be a promising therapeutic approach to mitigate
neuroinflammation and slow the progression of neurodegenerative diseases.
Notably, in vivo, studies have also revealed that human
GSK-3(S9A) mutated form exhibit elevated
P-Ser129--syn and p-Tau in TH-positive DA-ergic neurons in mice as
they age [17, 45]. These findings underscore the intricate interplay between
GSK-3, -syn phosphorylation, and the subsequent
neurodegenerative processes, shedding light on potential targets for therapeutic
intervention. Phosphorylation regulates the activity GSK-3 activity. The enzymatic
GSK-3 activity gets reduced following the phosphorylation at serine-9
[46]. The results from both our immunofluorescence and western blot analysis
showed a decrease in the protein levels of P-Ser9-GSK-3 in
MPTP-intoxicated mice. The MPTP-induced aberrant protein levels of
P-Ser9-GSK-3 was markedly alleviated following the Vanillin
treatment. Next, we assessed the protein levels of Wnt-3a, -catenin in
the SNpc region of mice. Our results revealed that MPTP-lesioned mice had
significantly reduced protein expression of Wnt-3a. Vanillin was significantly
able to rescue this decreased Wnt-3a levels. -catenin protein levels
were also considerably downregulated in MPTP-intoxicated mice. These reduced
levels of -catenin upregulated following Vanillin treatment in
MPTP-exposed mice. These findings imply that the neuroprotective properties
imparted by Vanillin in vivo may be largely mediated by the activation
of the Wnt/-catenin signaling, mainly by suppressing the GSK-3
activity.
Although neuronal autophagy is primarily recognized as a defensive mechanism, it
can paradoxically contribute to neuronal cell death. Numerous studies have
revealed the involvement of dysregulated autophagy in DA-ergic neuronal
degeneration [47, 48]. Notably, PD patients showed autophagy in nigral neurons [49].
Studies have observed that MPTP administration elevates LC3-II expression and
decreases p62 expression, indicating the activation of the autophagy-lysosomal
pathway [50, 51]. Furthermore, a recent study indicates that induced autophagy
can cause cell death in MPP+-exposed human neuroblastoma cells [52]. Our
immunofluorescence findings demonstrate that increased LC3 immunoreactivity in
the MPTP-induced mouse model is consistent with these observations. Vanillin
treatment significantly lowered the LC3 immunoreactivity in the
MPTP-administrated group. Reduced p62 expression and an elevated LC3-II/LC3-I
ratio indicate increased autophagic activity. Our results also show that the
expression of LC3/I protein was elevated and Vanillin treatment
significantly reduced the ratio of LC3/I protein expression in
MPTP-intoxicated mice. Additionally, p62 levels were decreased in the
MPTP-administrated rats, which further shows the increased autophagic-cell death
in MPTP-induced mice, and Vanillin administration markedly increased the p62
expressions. The effect of autophagy and autophagic cell death in dopaminergic
neurodegeneration appears to be contingent upon the particular cellular setting
and the initial causal cause. Improving the comprehension of autophagy stress
[53] and the mechanisms governing autophagic-cell death holds promise for
developing therapeutics to restore DA-ergic neuronal homeostasis in PD.
Furthermore, additional physiologically relevant knowledge on the autophagic
pathway’s activity status in PD patients is required to assess whether promoting
or suppressing autophagy would be more beneficial in reducing the symptoms of the
disease and delaying its progression. Future studies aimed to addressing the
present dispute will be critical in translating these discoveries into viable
therapies for PD.
In this study, we primarily focused on assessing the activity of GSK-3
and the activation of the Wnt/-catenin pathway in the context of
MPTP-induced nigrostriatal degeneration, as these pathways are known to be
involved in neuroprotection. While our study aimed to investigate the protective
effect of Vanillin on the dopaminergic system by targeting GSK-3
activity and the Wnt/-catenin pathway, demonstrating the co-expression
of related signaling molecules with dopaminergic neuron-specific markers would
have provided stronger evidence. Thus, future studies should incorporate
techniques such as immunofluorescence co-staining with specific markers for
dopaminergic neurons to further validate our findings and enhance the
understanding of Vanillin’s potential therapeutic effects on PD. Additionally,
our study focused specifically on the SNpc region, and the mechanisms directly
related to the Wnt/-catenin signaling pathway. However, we did not
investigate this pathway in the striatum region, which indeed plays a crucial
role in the dopaminergic system and the pathophysiology of PD in the striatum.
Future studies should consider examining this pathway in the striatum to provide
a more comprehensive understanding of PD and to identify additional therapeutic
targets against PD.
5. Conclusions
In conclusion, our study highlights the intricate mechanisms involved in PD
etiology and the potential therapeutic role of Vanillin. The aberrant
accumulation of -syn in the SNpc contributes to neurodegeneration in
PD. Vanillin treatment effectively mitigates the elevated levels of
phosphorylated P-Ser129--syn. Moreover, our study of
Wnt/-catenin signaling highlights the critical role it plays in PD
pathogenesis and highlights the possibility of using GSK-3 as a target
for neuroprotection and neurorestoration. GSK-3, implicated in both
-syn phosphorylation and -catenin degradation, emerges as a
critical player in PD pathophysiology. Vanillin treatment not only mitigates
GSK-3 activity but also positively modulates Wnt/-catenin
signaling, suggesting its neuroprotective effects. The dual role of autophagy in
neuroprotection and neurodegeneration, revealing its involvement in DA-ergic
neuronal degeneration in PD. Treatment with vanillin significantly reduces
abnormal autophagic-cell death, suggesting a possible function for vanillin in
reestablishing DA-ergic neuronal homeostasis. Thus, by inhibiting GSK-3
activity and reducing autophagic cell death in DA-ergic neurons, vanillin may
stimulate the Wnt/-catenin signaling pathway, which in turn may explain
its neuroprotective effect in our study Fig. 6. This study enhances our
understanding of the complex interplay of molecular pathways in PD and
underscores Vanillin’s promising therapeutic implications for the disease.
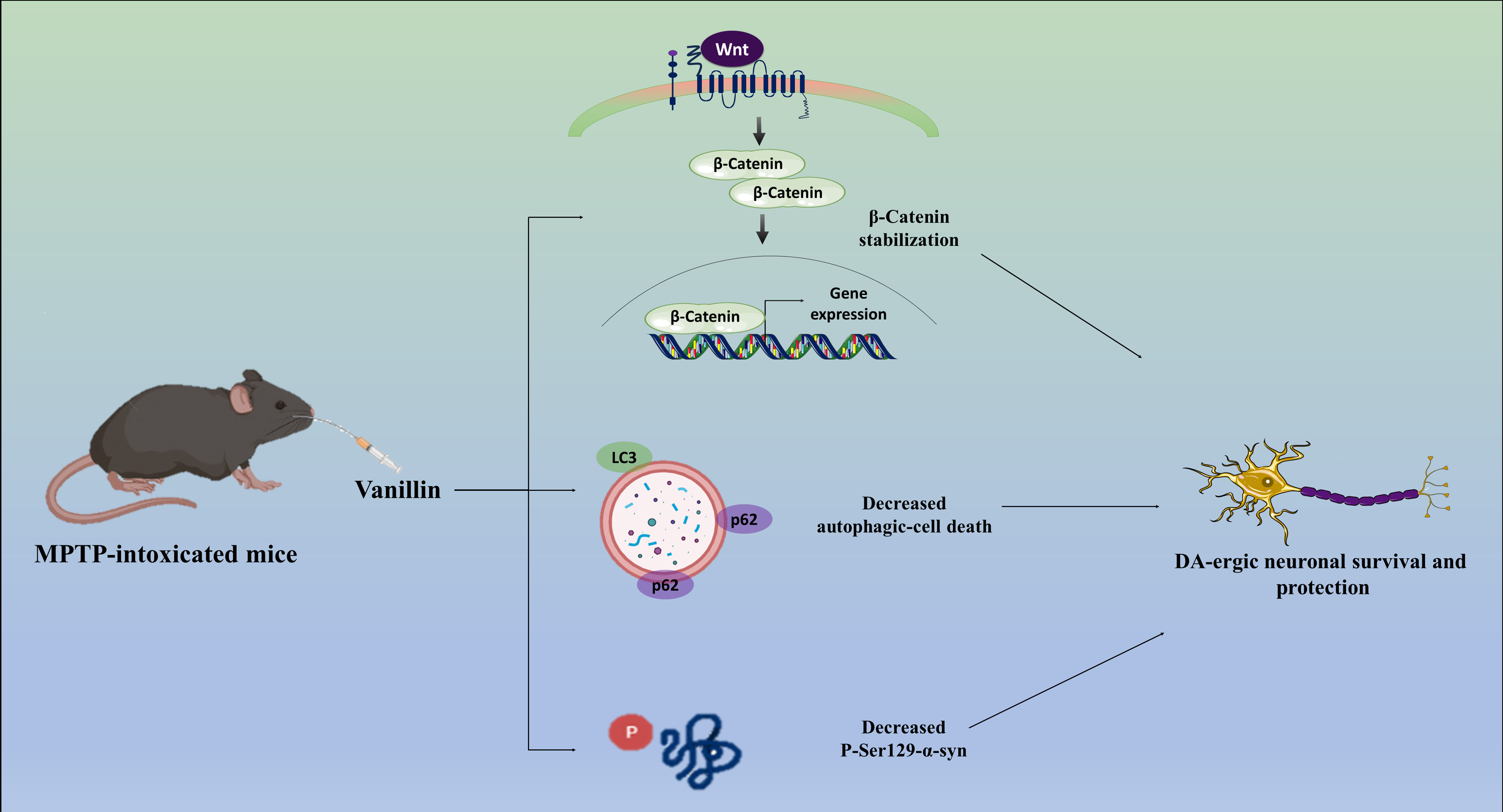
 Fig. 6.
Fig. 6.
Possible mechanisms that mediate the neuroprotective action of
Vanillin on MPTP-intoxicated PD mice model. DVL, Dishevelled; CK-1, casein kinase
1; APC, adenomatosis polyposis coli; Lrp, lipoprotein receptor-related protein;
TrcP, transducin repeats-containing protein; Ub, ubiquitin; Axin, axis inhibition protein; DA-ergic, dopaminergic.
Availability of Data and Materials
All the data produced during the study are already included in the manuscript,
and no further data is required to reproduce the results.
Author Contributions
ACM conceptualized, supervised the study and edited the final manuscript. LR performed the experiments and wrote the manuscript. Both authors read and approved the final manuscript. Both authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Ethics Approval and Consent to Participate
All the experiments were carried out on C57BL/6 mice following standard guidelines and regulations of the Institutional Animal Ethics Committee (IAEC), Jawaharlal Nehru University (JNU), New Delhi (10/GO/ReBi/99/CPCSEA/March 10, 1999). All the Standard methods and protocols were followed for animal handling and experiments. All efforts were made to reduce animal suffering.
Acknowledgment
The authors would like to acknowledge the Central Instrumentation Facility (CIF), School of Life Sciences, Jawaharlal Nehru University, New Delhi, India.
Funding
This study was supported by the Department of Biotechnology (DBT), Govt. of India (BT/PR38493/TRM/120/465/2020), and (BT/PR47726/CMD/150/26/2023), Ministry of Science and Technology (Govt. of India).
Conflict of Interest
The authors declare no conflict of interest.