


 , Wouter Huiting 1, Jan N. Keijser 1, Csaba Nyakas 2,3, Marieke J. G. van Heuvelen 4, Eddy A. van der Zee 1
, Wouter Huiting 1, Jan N. Keijser 1, Csaba Nyakas 2,3, Marieke J. G. van Heuvelen 4, Eddy A. van der Zee 11 Department of Neurobiology, Groningen Institute for Evolutionary Life Sciences (GELIFES), University of Groningen, 9747 AG Groningen, The Netherlands
2 Research Center for Molecular Exercise Science, Hungarian University of Sports Science, 1123 Budapest, Hungary
3 Department of Morphology and Physiology, Health Science Faculty, Semmelweis University, 1085 Budapest, Hungary
4 Department of Human Movement Sciences, University of Groningen, University Medical Center Groningen, 9713 AV Groningen, The Netherlands
Abstract
Beneficial effects of whole-body vibration (WBV) on brain and musculoskeletal health in mice have been demonstrated, but underlying mechanisms remain relatively unrevealed. WBV improves attention and memory performance in mice, putatively through stimulation of the cholinergic system. Here, we investigated the effects of WBV on the septo-hippocampal cholinergic system.
Young C57BL/6 mice (8 weeks old) were subjected to 10 min WBV/day (mechanical vibration: 30 Hz; ~0.1-μm peak-to-peak displacement), 5X/week for 5 weeks. In Experiment 1, choline acetyltransferase (ChAT)-immunoreactivity in the septum and hippocampus was analyzed either 2 or 24 h after the last WBV session. Pseudo-WBV-treated mice (same handling procedure as WBV, but no vibrations) served as controls. In Experiment 2, the longitudinal profile of ChAT-immunoreactivity was analyzed in the hippocampus after 1, 2, 3, 4, or 5 weeks of WBV. In addition, synaptophysin immunostaining was performed at either 2 and 5 weeks of WBV. Mice housed 1/cage during the entire experiment served as controls. The balance-beam test was used to monitor the functional impact of WBV. In Experiment 3, a Y-maze reference-memory test was performed after 5 weeks of WBV to obtain a functional cognitive outcome measure of WBV. Pseudo-WBV treated mice served as controls.
In Experiment 1, ChAT-immunoreactivity was significantly enhanced after the last WBV session of the 5-week period. This was found in the septum, Cornu Ammonis 1 (CA1), CA3, and dentate gyrus, and was dependent on layer and time-point (2 or 24 h). Experiment 2 revealed that, ChAT-immunoreactivity was lower after 2 weeks of WBV, whereas it was significantly higher after 5 weeks (similar to in Experiment 1). Immunostaining for synaptophysin, a marker for synaptic density, was also significantly higher after 5 weeks of WBV, but not significantly lower after 2 weeks, as was ChAT. WBV-treated groups performed significantly better than did controls on the balance beam from week 3 onwards. Experiment 3 showed that WBV-treated mice had better spatial-reference memory performance in the Y-maze test than did pseudo-WBV controls.
Our results indicate that WBV stimulates the septo-hippocampal cholinergic system in a gradual and dynamic way that may contribute to improved spatial-memory performance. This finding suggests that WBV, by upregulation of the septo-hippocampal cholinergic system, may be considered a valuable therapeutic strategy to enhance brain functions in aging, neurodegenerative, and other brain diseases.
Keywords
- acetylcholine
- cholinergic system
- motor coordination
- passive exercise
- synaptophysin
Long-term physical activity has been widely acknowledged to be a low-cost and easily affordable strategy to improve cognitive functions [1]. The benefits of physical activity in elderly populations have been examined in relation to general health, brain functions, and risk of neurogenerative diseases [2]. However, despite the rapid development of the fitness and sport industry to emphasise the availability and affordability of new exercise strategies, in practice it is still very hard to implement exercise interventions for elderly populations. This is a multifactor problem for several reasons. First, subjects with limited physical or cognitive capabilities often refrain from performing physical activity. In addition, a lack of motivation or willingness to engage in physical activity, or the combination of both factors, often occurs. A generally less demanding alternative may be whole-body vibration (WBV). WBV is considered to be a form of passive exercise in which a well-controlled mechanical stimulus is provided by special vibrating platforms [3, 4]. In clinical experiments, WBV was found to stimulate the musculoskeletal system, resulting in increased muscle strength [5], motor coordination [6], hormonal responses [7], and bone mineral density [8]. Translational WBV research has provided additional insights into the underlying (neuro) biological mechanisms; similar effects of WBV on osseous and muscle parameters have been observed in rodent models [9]. WBV may also affect the central nervous system, including various domains of cognitive functions and behavior, in a manner that is similar to that of physical activity. Recently, the relevance of translational WBV, in relation to brain functioning and brain disorders, has been highlighted and reviewed [4, 10, 11, 12]. Long-term (5–12 weeks) WBV in rodents alleviates degeneration of neurons and glial cells and the accumulation of proinflammatory cytokines [13, 14, 15, 16, 17]. In addition, increased levels of neurotrophic factors and neurogenesis markers were observed after WBV [13, 18, 19]. A range of improved memory functions and reduced anxiety-like behavior has also been observed in rodents as a beneficial effect of WBV [13, 15, 16, 20, 21]. Although it is not entirely clear what mechanism leads to these cognitive benefits, WBV seems to exert prominent effects on the hippocampus, a pivotal region for memory and learning functions [13, 14, 15, 22, 23, 24].
From earlier studies, it is known that WBV may lead to an acute release of various neurotransmitters [18, 25, 26, 27, 28]. Furthermore, our previous exploratory study indicated that cholinergic activity, based on the expression level of the rate-limiting acetylcholine-producing enzyme choline acetyltransferase (ChAT), was elevated in the cortex and amygdala after 5 weeks of WBV [29]. The cholinergic system enhances attention and plays a crucial role in learning and memory processes [30]. It can be activated in response to external environmental stimulation, including exercise [31, 32]. Therefore, the positive impact of WBV on learning and memory processes, shown in both rodents [13, 15, 16, 20] and humans [33, 34], may, in part, be due to the enhanced activity of the cholinergic system.
In order to extend the knowledge of the effect of WBV on the cholinergic system, and hence cognition and other domains of behavior regulated by the cholinergic system, we performed a study consisting of three exploratory experiments using ChAT expression in the hippocampus as an indirect measure of cholinergic activity of septal cholinergic-projecting cells. The primary aim of these experiments was to elucidate the effects of WBV on hippocampal cholinergic neurotransmission and spatial-reference memory functions. In addition, we evaluated synaptic density using synaptophysin as a marker. Both ChAT and synaptophysin levels were determined by way of immunohistochemistry. Quantification of synaptophysin immunoreactivity has been shown to be a reliable measure of synaptic density [35, 36]. Moreover, hippocampal synaptophysin immunoreactivity correlates positively with hippocampal ChAT expression [37, 38].
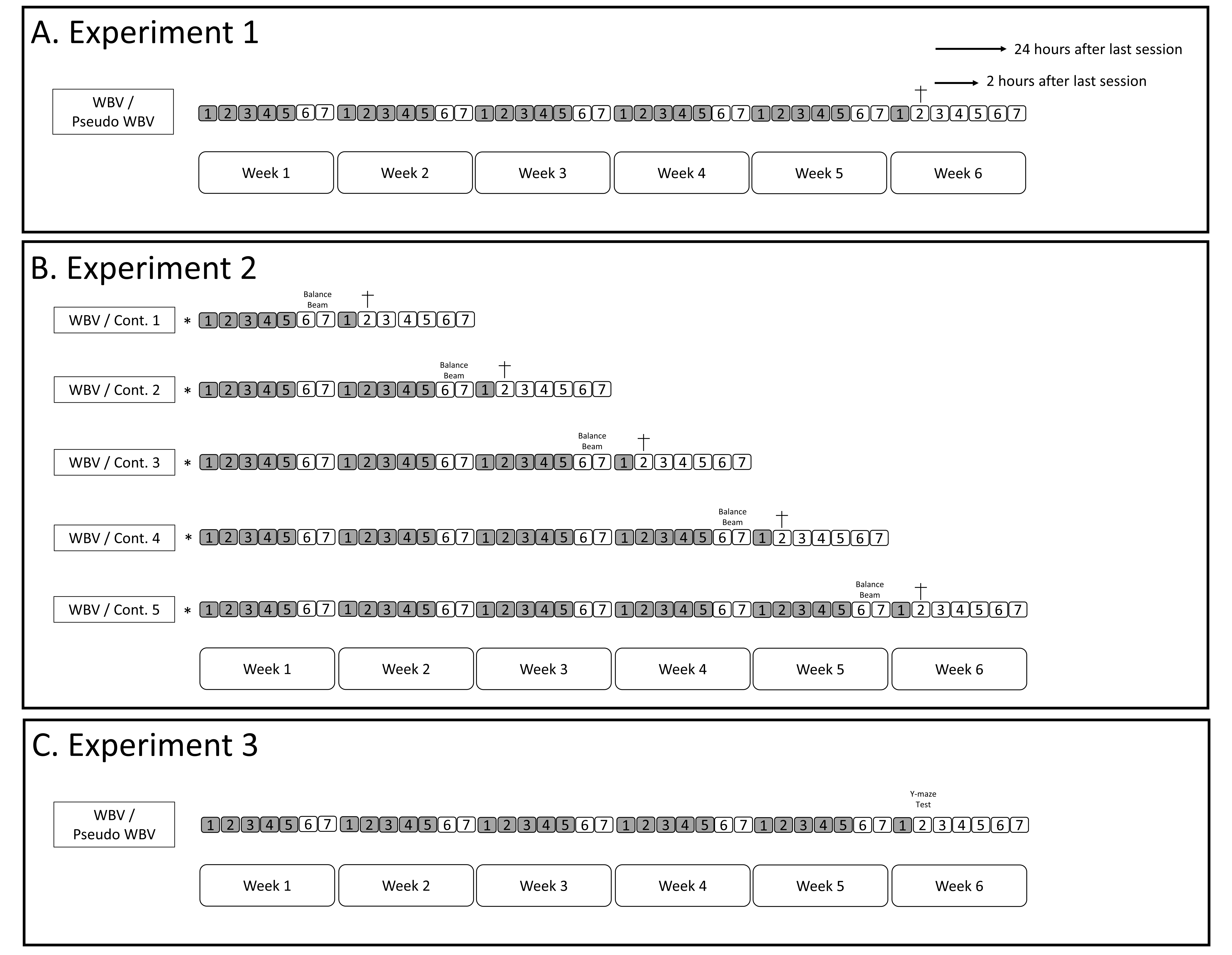
The present study was separated into three experiments. First (Experiment 1), in order to validate the time point of termination after the last WBV session, the long-term effects (5 weeks) of WBV on cholinergic activity in the medial septum and hippocampus were studied in young mice. For this, mice were randomly assigned by a randomizer application to four experimental groups: two WBV-treated groups (n = 7 and 8) and two pseudo-WBV-treated groups (n = 7 and 8). After 5 weeks of WBV, a WBV and a pseudo-WBV control group were terminated 2 h after the last WBV session, and the other two groups (WBV and pseudo-WBV) were terminated 24 h after the last WBV session. This exploratory study was aimed at determining the temporal dynamics of WBV on cholinergic activity by investigating its effects at either 2 or 24 h after the last WBV session. This experimental design is shown in Fig. 1A.
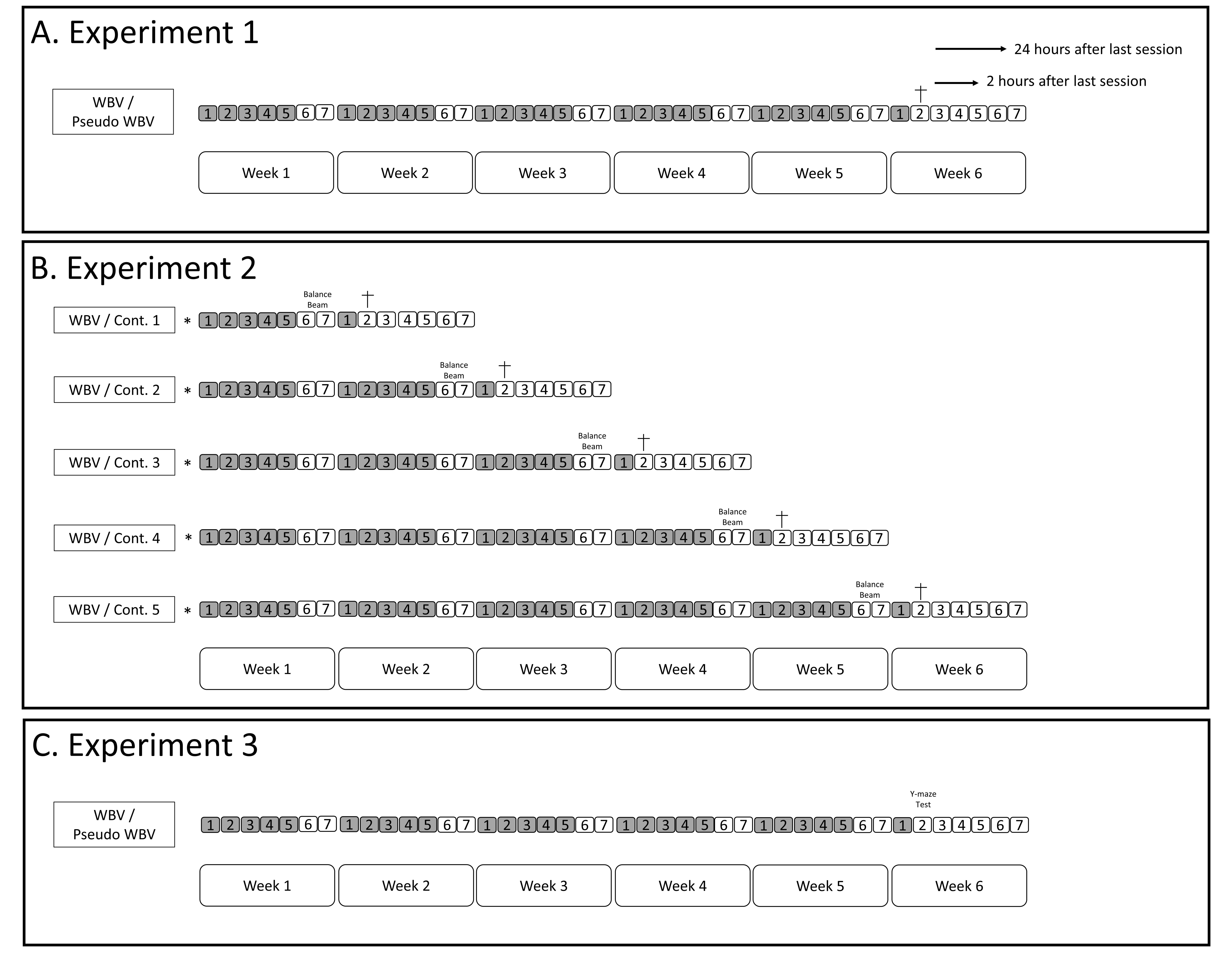 Fig. 1.
Fig. 1.
Experimental design. This study consisted of three separated experiments. Gray blocks indicate days with a whole-body vibration (WBV) session of 10 min, and open blocks indicate days without WBV sessions. In Experiment 1 (A), 8-week-old male C57BL/6 mice were assigned to four experimental groups: two WBV-treated groups (n = 8/gp) and two pseudo-WBV-treated groups (n = 7/gp). Mice were terminated after 5 weeks of WBV. A WBV group + its corresponding pseudo-WBV group was terminated either 2 h or 24 h after the last WBV session (†). In Experiment 2 (B), 8-week-old male C57BL/6 mice were randomly distributed among 10 experimental groups: 5 groups underwent WBV and 5 corresponding groups served as home-cage controls (n = 8/gp). Mice were euthanised after 1, 2, 3, 4 or 5 weeks of WBV (24 h after the last WBV session). A baseline measure of motor coordination was obtained for mice in all groups before the start of intervention (indicated by *). Motor coordination of mice in all groups was reassessed at the end of the intervention, one or two days before termination (†). Brain tissue was collected for immunohistological purposes in the first and second experiments (†). In Experiment 3 (C), 8-week-old male C57BL/6 mice were randomly assigned to two experimental groups: a WBV treated group and a pseudo-WBV treated group (n = 13/gp). Spatial-reference memory was analyzed by a Y-maze reference-memory test at the end of 5 weeks of WBV intervention. The WBV intervention continued during the Y-maze test period, but was always done after the test session in order to avoid direct interference of the intervention with the test performance.
In Experiment 2, the long-term effects of WBV on cholinergic activity (and motor coordination) were studied with a longitudinal design. Mice were randomly assigned to 10 experimental groups (n = 8 each): 5 WBV-treated groups and 5 home-cage control groups. A balance-beam test was performed to assess motor coordination. A baseline measurement was obtained for all experimental groups at the start of the intervention, and a correspondent home-cage control group and a WBV group were tested every week before euthanasia. From the second week onwards, after each week, a WBV and a home cage control group were terminated 24 h after the last WBV session (Fig. 1B). This exploratory study was designed to investigate the longitudinal, weekly development (from week 1 to week 5) of ChAT-immunoreactivity in the hippocampus (dentate gyrus, CA3, and CA1). Next, based on the obtained ChAT results (see below in Section 3.2 Experiment 2), we analyzed synaptic density in either week 2 or 5 using synaptophysin as a marker. In addition, we knew from our previous study [20], that the body weight of mice was not affected by 5 weeks of WBV intervention with daily 10-min sessions. However, we did not examine this previously by the comparison of WBV mice and home-cage controls on a weekly basis. We weighed the mice every week in Experiment 2.
The long-term effects of WBV on spatial reference memory were investigated in Experiment 3. Mice were randomly divided into two experimental groups (n = 13/gp): a WBV-treated group and a pseudo-WBV group. The mice were tested in a Y-maze after 5 weeks of WBV to assess reference-memory function. The experimental design is shown in Fig. 1C.
One hundred and thirty-six C57BL/6 male mice were used in these experiments
(Experiment 1, n = 30; Experiment 2, n = 80; Experiment 3,
n = 26). The mice were 8 weeks old at the start of the WBV
interventions. All mice were maintained in the same housing conditions:
individually housed in cages (30
We adhered to the reporting guidelines for WBV studies in animals [39]. Mice
underwent the same WBV procedure in all three experiments, including a single,
daily WBV session of 10 min, 5X/week. No habituation to the device before the
start of the WBV protocol took place. The WBV device had been used by our
research group earlier [20, 29, 40] and it consisted of a power amplifier (V406
Shaker Power Amplifier, Ling Dynamic Systems Ltd,
Hertfordshire, UK) and an oscillator (LEVELL R.C. Oscillator Type TG200DMP, Levell electronics Ltd, Hertfordshire, UK) attached to a cage with 12
removable compartments (6.5
 Fig. 2.
Fig. 2.
WBV device. The whole-body vibration device used in this study consists of 3 main compartments (A): (1) the oscillator (panel B – upper device); (2) the power amplifier (B – lower device), and (3) the cage with 12 removable compartments (C). This system provides a constant frequency of 30 Hz and amplitude of 54 µm, and allows for the treatment of 12 mice simultaneously without physical interactions.
The balance-beam test was used to measure motor coordination in Experiment 2. It served as an outcome measure of muscle performance as well as an indicator of the functional impact of the WBV treatment (based on previous experiments [20, 40]). A 1-m long, 4-mm wide, wooden beam was placed horizontally 80 cm above a container filled with paper used for protecting the mice if they fall. A safe cage, which served as motivation for the mouse to cross the beam, was placed at the far end of the beam. Firstly, mice were allowed to familiarise themselves to the safe cage for 1 min. Secondly, the mice were placed in the middle of the beam to allow them to become familiar with the test environment. Finally, mice were placed at the near end of the beam facing the safe cage. The time to cross to the safe cage was recorded. Balance-beam sessions were carried out at the beginning of light phase.
A Y-maze task was used to assess spatial-reference memory functions, as described previously [41]. Briefly, the Y-maze task was carried out in a tubular, transparent, Plexiglas area consisting of a start arm and two test arms that formed the Y. The arms were positioned at 120° angles from each other and were 5 cm in diameter and 27.5 cm long. A start box was attached to the start arm. A food reward consisting of a small crumb of regular food (0.05–0.1 g) was placed in one of the test arms as bait. Food crumbs were also placed below the perforations at the end of the other test arm. This ensured that both arms would smell the same and thereby helps to avoid discrimination between the baited and non-baited arms based on smell. A small rim (1 cm high) was placed at the end of the arms to prevent visual inspection of the bait. A guillotine door was installed halfway down each arm. When the mouse chose an arm, the other arm was closed by the guillotine door, which was operated by the researcher.
Each mouse underwent two habituation trials on the first day to familiarise them with the experimental situation. In the first habituation trial, the mouse was placed in the start box, from which it was allowed to explore one of the two test arms, which was baited with a small crumb of food. The other arm was closed. After the mouse consumed the bait, it was allowed to roam back to the start box. The second habituation trial was immediately after the first, and the other test arm was opened and baited. After the habituation, mice performed a training protocol for seven days consisting of one training session/day. Each training session consisted of 6 trials. During the entire training phase, the role of the left–right test arms (baited or not baited) was randomly switched. A valid visit or choice was considered to have occurred when the mouse placed all of its paws in a test arm (and the experimenter closed the other arm). After the mouse returned to the start box, the start arm attached to the start box was closed, preventing the mouse from reentering the maze. After cleaning both the start and test arms and re-baiting the same arm, the test procedure was repeated again, allowing the mouse to re-explore the maze. A correct choice occurred when the mouse visited the baited arm. Correct choices for each training session were used as final outcome measures and determined the longitudinal learning curve of the mouse. In addition, the area under the curve (AUC) was calculated to describe the overall performance of the mouse in the Y-maze task. All Y-maze procedures were carried out at the beginning of the light phase.
Mice were terminated with CO2 [42], directly followed by transcardial perfusion with saline (0.9% NaCl solution and 1 unit/mL heparin) (02405822, Apotheek Albert Schweitzer Ziekenhuis, Dordrecht, The Nederlands) and then with a 4% para-formaldehyde solution (158127, Sigma-Aldrich, St. Louis, MO, USA). Brains were collected and fixed in 4% para-formaldehyde solution for 24 h. After fixation, the brains were washed in phosphate buffer solution for 24 h, then were dehydrated in 30% sucrose solution. Samples were frozen with liquid nitrogen and stored at –80 °C until sectioning. Coronal sections of 20 µm were cut by cryostat, and free-floating sections were stored in phosphate-buffered solution containing 0.2% sodium azide at 4 °C until the immunohistochemical staining procedure.
The cholinergic marker ChAT (the rate-limiting enzyme for the production of acetylcholine) was visualised in both experiments with immunohistochemistry. Before the incubation of the primary antibody, sections were preincubated in 0.01 m Tris-buffered saline (TBS) containing 5% normal rabbit serum (NS01L, Sigma-Aldrich, St. Louis, MO, USA) and 0.2% Triton X-100 (TX) (X-100, Sigma-Aldrich, St. Louis, MO, USA). Primary antibody (Goat antibody ChAT, 1:400; Millipore Sigma, Burlington, NJ, USA) was incubated in 0.01 M TBS, 5% normal rabbit serum, and in 0.2% Triton X for 3 h in a 37 °C water bath, followed by 1 day at room temperature and 2 days in a cold room at 4 °C. Secondary antibody (Rabbit anti goat, 1:400; Jackson ImmunoResearch, Baltimore Pike, West Grove, PA, USA) was incubated in 0.01 M TBS containing 1% normal rabbit serum and 0.2% Triton X for 4 h at room temperature, then allowed to stand overnight in a cool room.
Synaptophysin, an integral membrane protein in the presynaptic vesicles of neurons, was stained to visualise presynaptic boutons and served as an indicator of synaptic density in Experiment 2. Sections were preincubated in 0.01 M TBS, 3% bovine serum albumin (A4503, Sigma-Aldrich, St. Louis, MO, USA), and 0.5% TX for 1 h. Primary antibody (mouse antibody, 1:1000; Millipore Sigma, Burlington, NJ, USA) was incubated in 3% bovine serum albumin and 0.5% TX in 0.01 TBS for 3 h at room temperature, followed by 3 days incubation at 4 °C. Secondary antibody (Goat anti mouse, 1:400; Jackson ImmunoResearch, Baltimore Pike, West Grove, PA, USA) was incubated in 0.01 TBS for 2 h at room temperature.
Before incubation of the primary antibodies, brain sections were prewashed in 0.01 M TBS followed by incubation of 0.3% H2O2 for 30 min. Finally, all sections were incubated with avidin-biotin peroxidase complex for 2 h (Vestastain ABC kit, Vector, Burlingame, CA, USA) and visualised with 0.075 mg/mL 3,3′-diaminobenzidine (Sigma-Aldrich, St. Louis, MO, USA) solution. During the procedures, sections were extensively rinsed in 0.01 M TBS or in TBS containing 0.2% TX. Sections were mounted on microscopy slides, dehydrated in gradients of ethanol-xylol solutions and cover slipped for microscopic analyses.
Quantification of the immunohistochemical data was performed with a Quantimet system (Leica DFC 365FC camera, Leica Biosystems, Nussloch, Germany) integrated into a microscope system (Olympus BH2 microscope, Olympus-Life science, Shinjuku-ku, Tokyo, Japan) run by Quantimet software (Quantimet 570, Leica Biosystems, Nussloch, Germany). Field-size analysis was performed to ensure that Quantimet data collection was performed objectively. No significant difference in field sizes were found between experimental- and home-cage or pseudo-WBV control groups.
Optical density for ChAT was analyzed in the granular and molecular layers of dentate gyrus inner blade (DGI) for Experiment 1; and in both the DGI and outer blades (DGO) for Experiment 2. In addition, ChAT optical density was measured in the oriens and pyramidal layers of CA1 and CA3 subregions for both experiments, as well as in the medial septum (cell body and dendrites were separated) for Experiment 1.
The optical density for synaptophysin was obtained in the molecular layers of
DGI and DGO blades for Experiment 2. These regions were selected because they
were representative for the ChAT results and the high quality of the
synaptophysin immunoreactivity. The obtained data were corrected for a specific
background staining in the corpus callosum. Analysis was performed on pictures
taken at 200
All statistical analyses were done using Microsoft Excel (Microsoft Office Professional Plus 2019, Microsoft Cooperation, Redmond, WA, USA). Student’s independent
t-tests were performed to evaluate the differences between the WBV and pseudo-WBV
or home-cage control groups. Outliers (more than 2 Standard Deviations (SD) from
the mean) were removed. Significance was defined as p
In Experiment 1, we determined whether septal and hippocampal ChAT immunoreactivity was altered after 5 weeks of WBV. In addition, we determined whether the time of termination (either 2 or 24 h after the last WBV session) influenced the outcome. Hippocampal ChAT expression was measured in subregions of the dorsal hippocampus (DGI, CA3, and CA1) and in the medial septum. ChAT-immunoreactivity visualised the cholinergic fibers arising from the cholinergic cells of the medial septum that innervated the different hippocampal subregions.
ChAT-positive immunostaining was not changed in the cholinergic cell bodies
after WBV, either at the 2-h hour or the 24-h time point. In contrast, a
significant increase in the axons and dendrites of the medial septum was found in
the mice terminated 2 h after the last WBV session (p
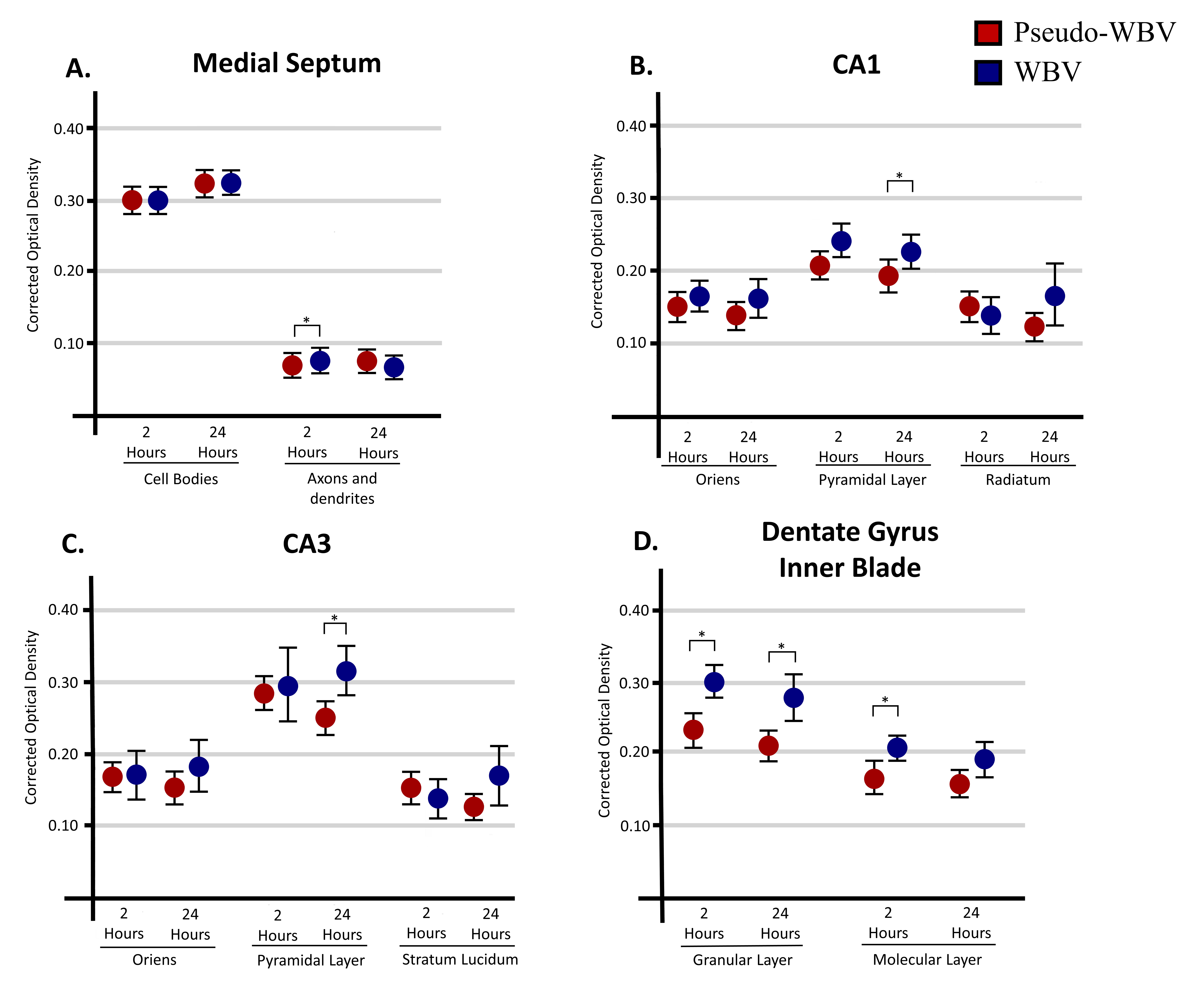 Fig. 3.
Fig. 3.
Experiment 1—ChAT immunostaining in the medial septum and
parts of the hippocampus. Effects of 5 weeks of whole-body vibrations (WBV)
intervention on the progression of choline acetyltransferase (ChAT)
immunostaining in mice terminated after either 2 or 24 h after the last WBV
session in the cell bodies and dendrites of medial septum (A), three layers
of Cornu Ammonis 1 (CA1) (B), and CA3 (C), as well as in the granular and molecular
layers of the dentate gyrus inner blade (DGI) (D). Cholinergic activity was
significantly higher in the dendrites and axons of the medial septum in the 2-h
group (A). WBV significantly increased ChAT levels in the pyramidal layer
of CA1 (B) and CA3 (C) subregions in the 24-h group. ChAT level was
significantly higher in the granular layer of DGI both in the 2- and 24-h groups
(D). The molecular layer of DGI was only significantly affected by WBV in
the 2-h group. Data are presented as mean
In Experiment 2, we investigated the longitudinal profile of ChAT immunoreactivity and motor coordination after 1, 2, 3, 4, or 5 weeks of WBV. In order to obtain comparable data of a possible longitudinal ChAT fluctuation induced by WBV over 5 weeks, as well as exclude the potential effects of other factors on ChAT level such as daily handling, stress, or novel environment, undisturbed mice served as home-cage controls.
Mice were weighted every week to assess possible effects of WBV on body weight.
A daily 10-min session of WBV, 5 days/week, for 1–5 weeks of treatment had no
significant effect on body weight. A balance-beam test was performed for
evaluation of motor coordination in this second experiment. Balance-beam
performance was not significantly different between the WBV and home-cage
controls before treatment or after the first or second week of WBV. A
significantly shorter walking time (indicating better motor coordination and
performance) was found in the WBV-treated group than in the home-cage group from
the third week onward (weeks 3 and 4: p
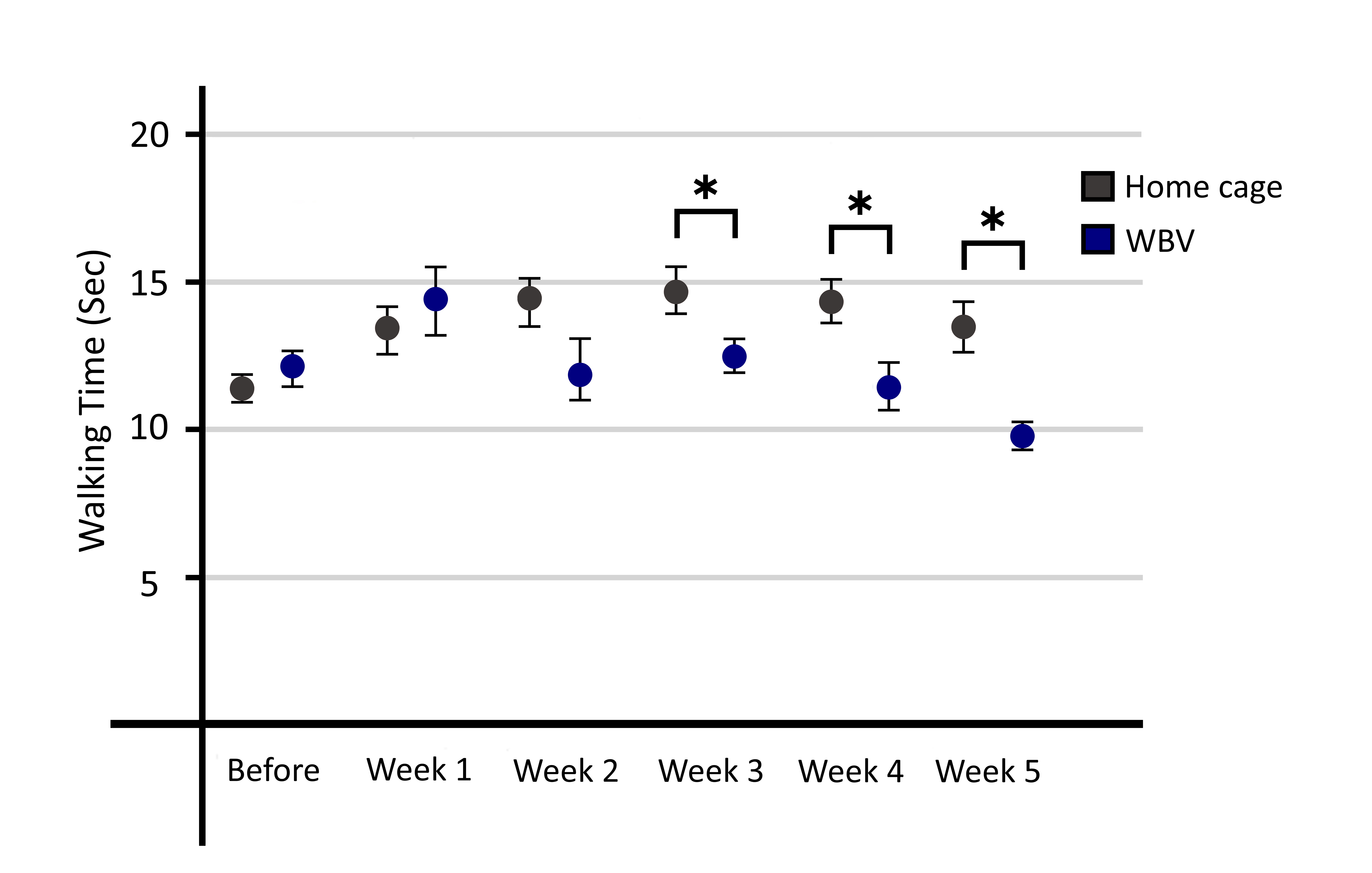
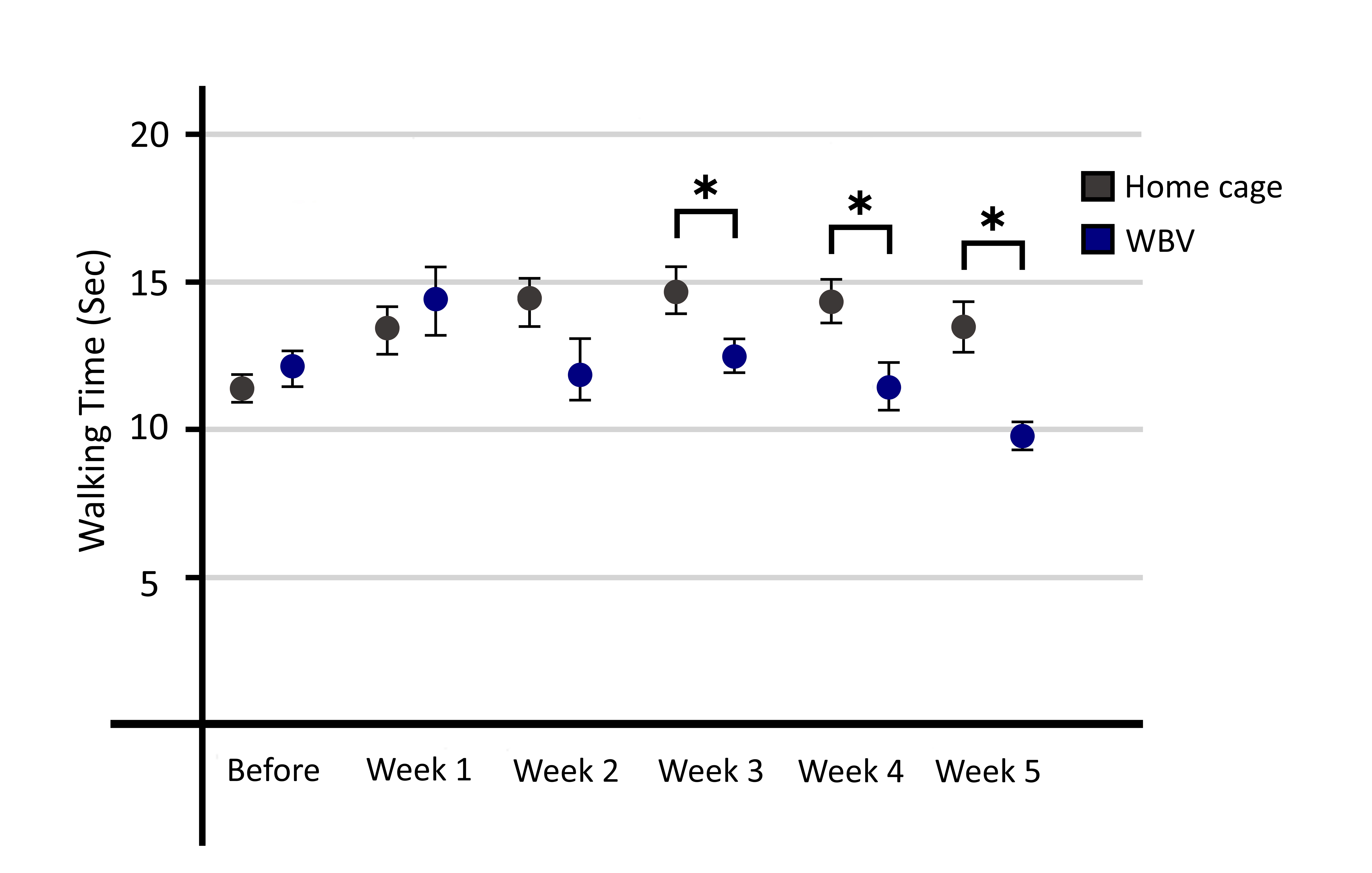 Fig. 4.
Fig. 4.
Experiment 2—Walking time on the balance beam as measure of
motor coordination. Effects of 1–5 weeks of whole-body vibrations (WBV)
intervention on progression of motor coordination in the balance-beam test,
showing the functional impact of WBV. Balance-beam performance was significantly
improved from week 3 onward by the WBV intervention. Data are presented as mean
We further scrutinised the impact of WBV on ChAT immunoreactivity in Experiment
2. To determine the dynamics of ChAT levels over the 5-week intervention period,
we measured the optical density of ChAT staining in subregions of the dorsal
hippocampus (DGI, DGO, CA3, and CA1) at either week 1, 2, 3, 4 or 5. Home-cage
mice served as controls to provide the most stable ChAT level for comparison. The
home-cage controls showed a significantly higher level of ChAT-positive staining
after 2 weeks of the experiment than did the WBV-treated group. This significant
difference was detected in the stratum oriens of CA1 and CA3 (CA1: p
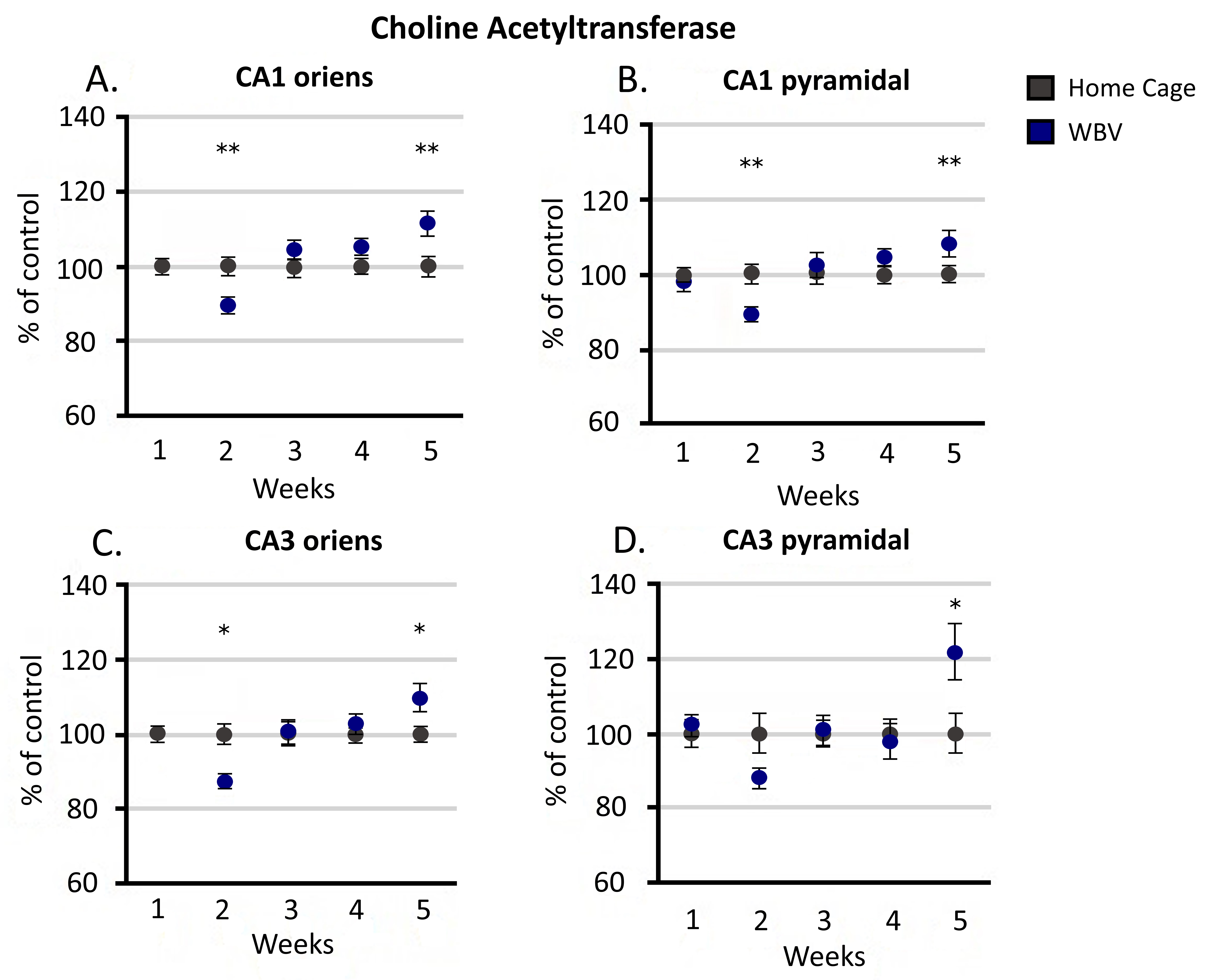
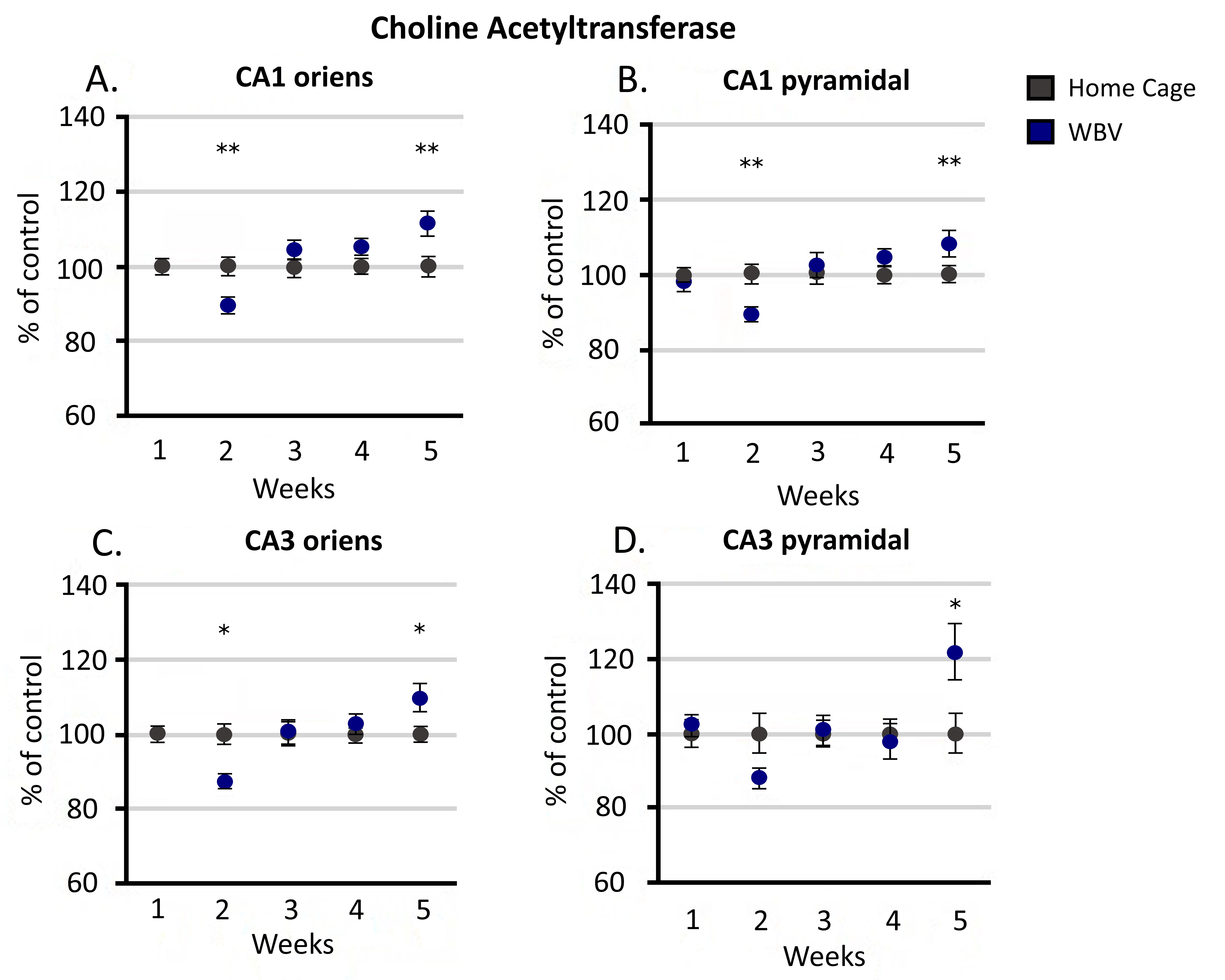 Fig. 5.
Fig. 5.
Experiment 2—ChAT immunostaining in the hippocampal CA1 and
CA3 subregions. Effects of 1–5 weeks of whole-body vibration (WBV) intervention
on the progression of choline acetyltransferase (ChAT) immunostaining in the CA1,
stratum Oriens (A), and pyramidal layer (B); and in the CA3, stratum
oriens (C), and pyramidal layer (D). ChAT levels were significantly
lower in the WBV group after 2 weeks and higher in the WBV group after 5
weeks. All data are presented relative to the baseline level (100%) of
the home-cage control groups. Data are presented as mean
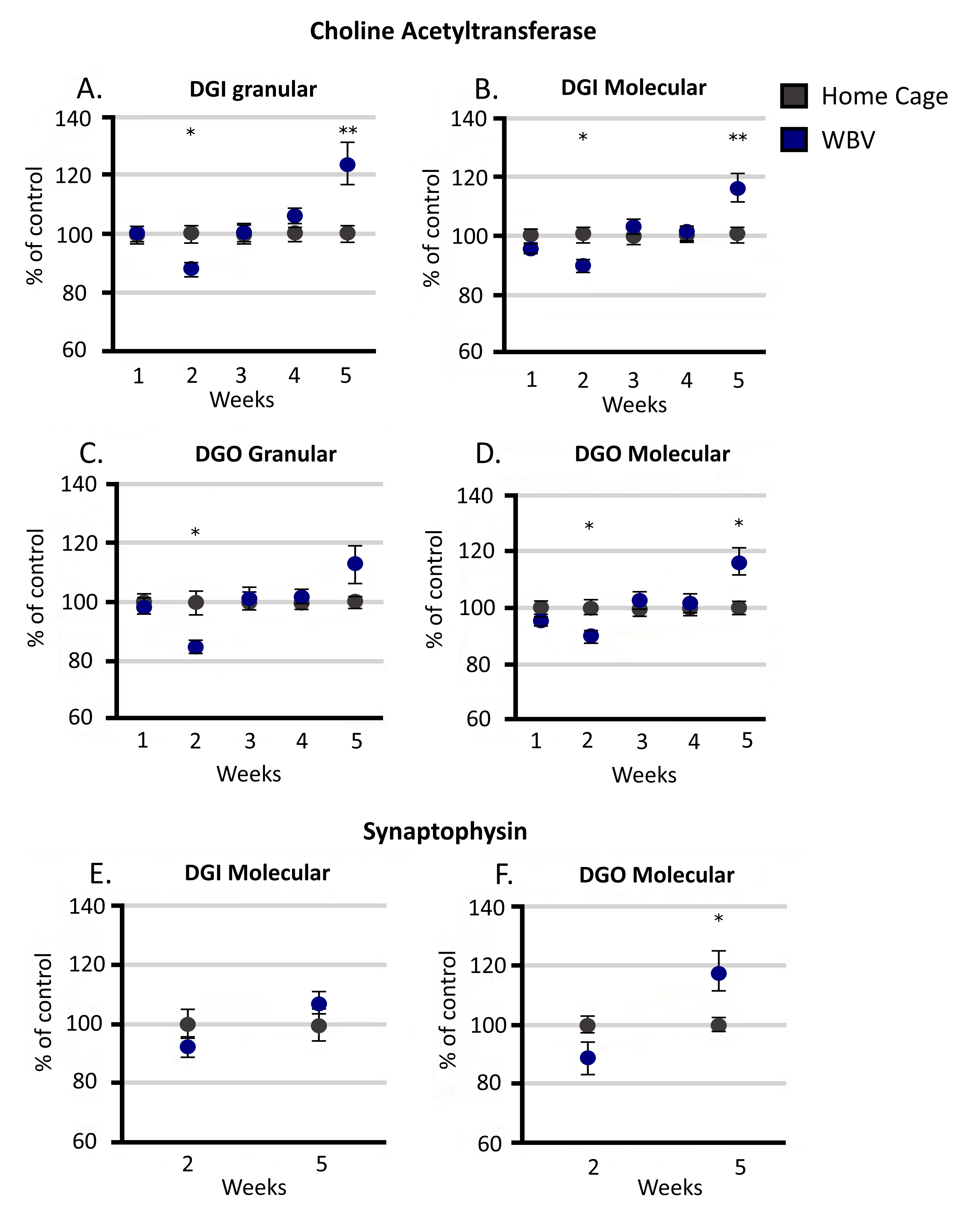
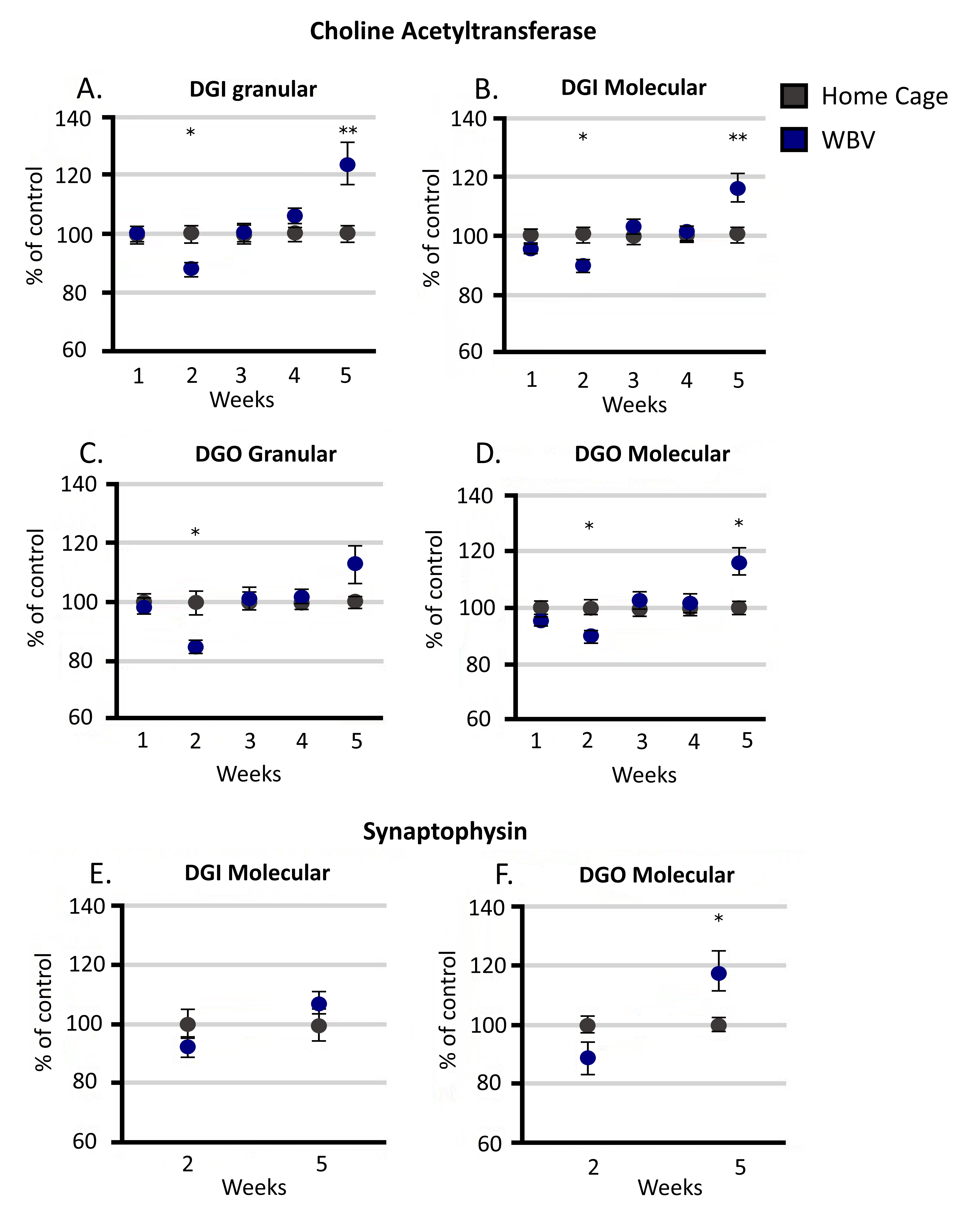 Fig. 6.
Fig. 6.
Experiment 2—ChAT and synaptophysin immunostining in the
hippocampal dentate gyrus subregions. Effects of 2 or 5 weeks of whole-body
vibration (WBV) intervention on the progression of choline acetyltransferase
(ChAT) immunostaining in the granular (A) and molecular layers (B) of
the dentate gyrus inner (DGI) blade, as well as in the granular (C) and
molecular layers (D) of dentate gyrus outer (DGO) blade. Bottom panels: the
effects on synaptophysin immunostaining in the molecular layer of DGI (E)
and DGO blades (F). ChAT levels were significantly lower in the
WBV group after 2 weeks; ChAT levels were significantly higher in the WBV group
after 5 weeks. Level of synaptophysin was only higher in the DGO blade after 5
weeks of WBV. All data are presented relative to the baseline level
(100%) of home-cage control groups. Data are presented as mean
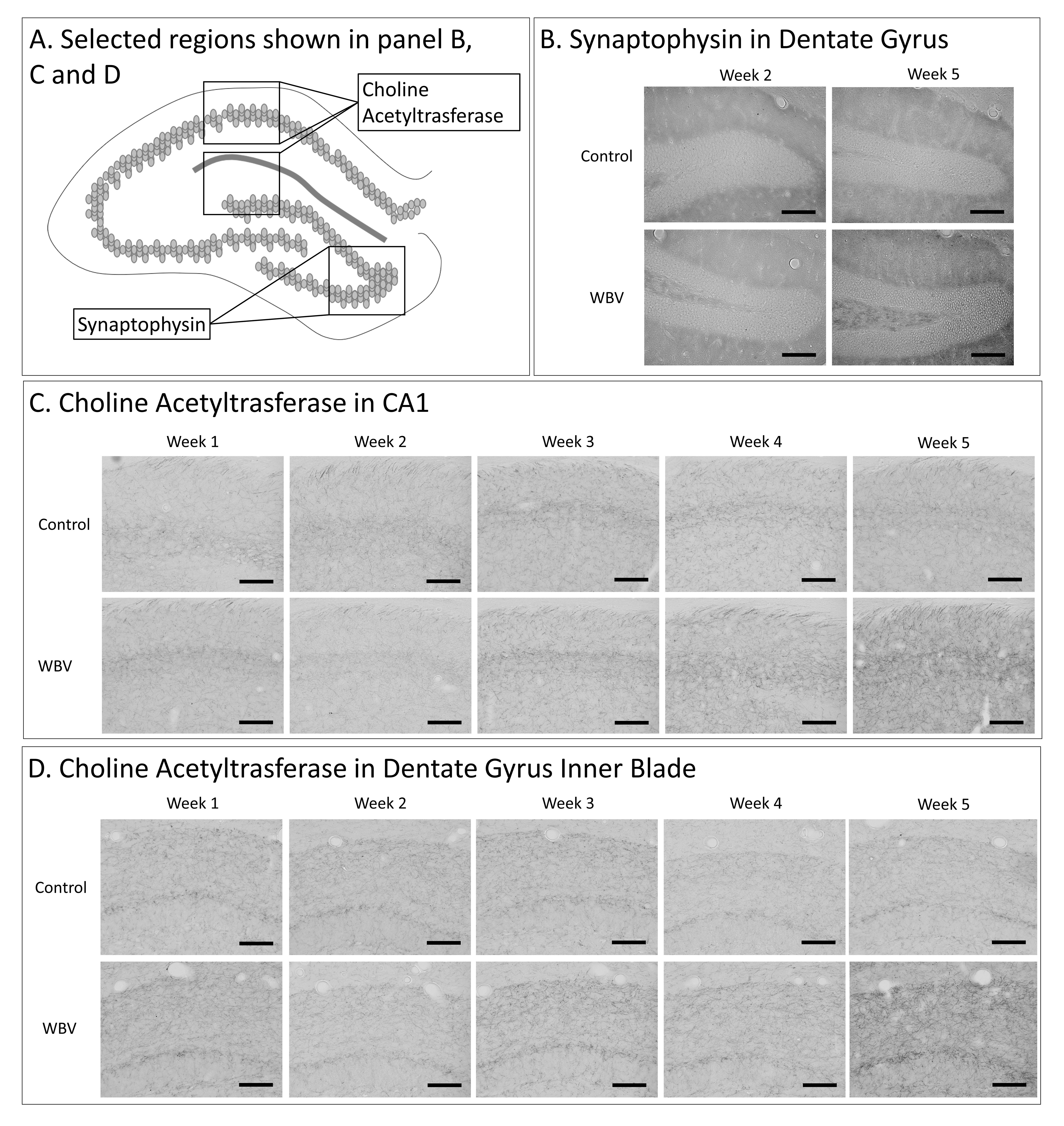 Fig. 7.
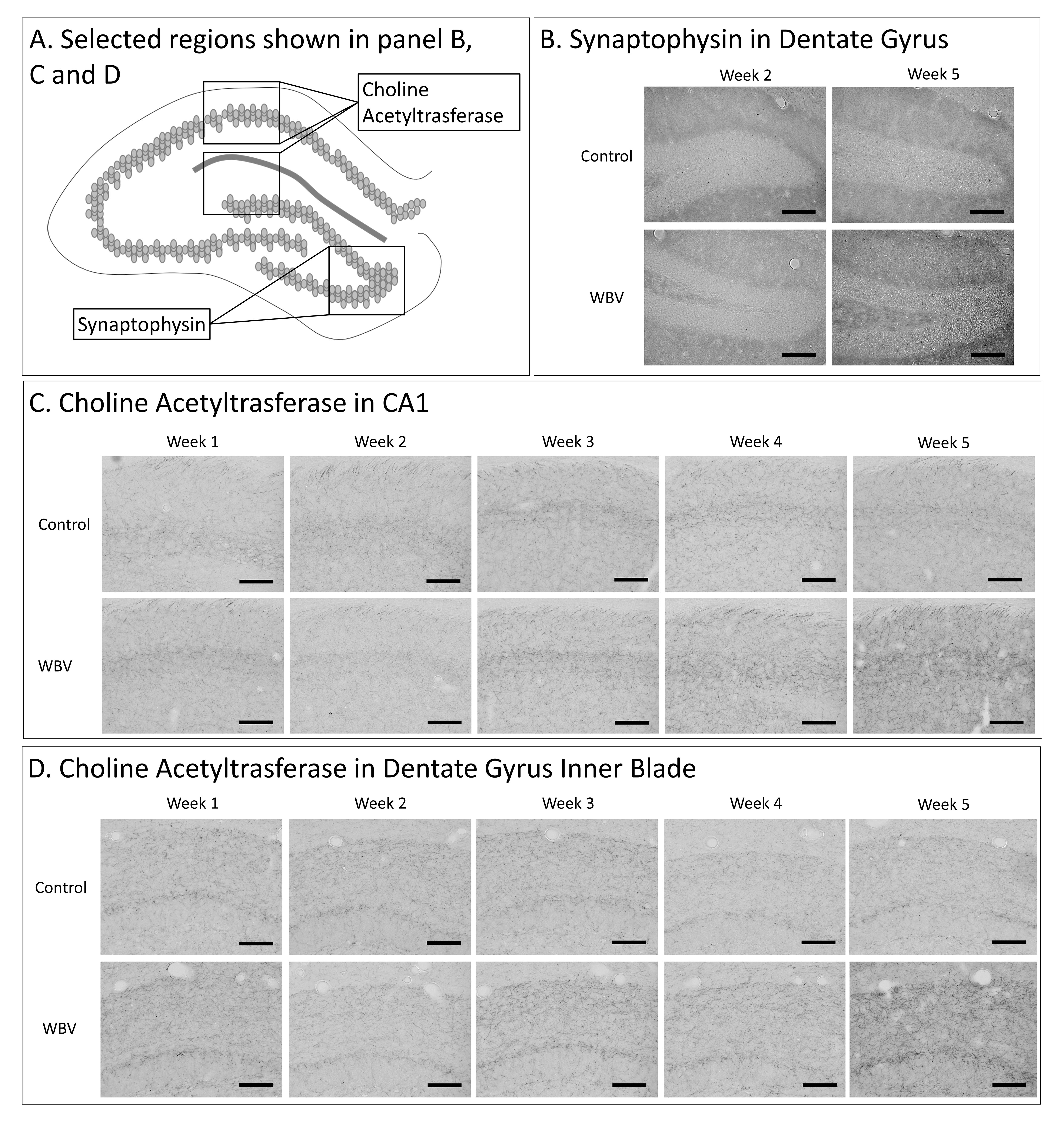
Fig. 7.
Experiment 2—Representative pictures of ChAT and synaptophysin
immunostaining in selected hippocampal regions. Schematic overview of
hippocampal regions of interests for choline acetyltransferase (ChAT) and
synaptophysin (A). Representative images of the synaptophysin
immunostaining in the dentate gyrus in weeks 2 and 5 are shown in (B).
Representative images of ChAT immunostaining from week 1 to week 5 are visualised
in the CA1 (C) and dentate gyrus inner (DGI) blade (D) hippocampal
subregions. Images were taken at 200
The aim of Experiment 2 was to analyze the longitudinal progression of ChAT
immunostaining (as an indicator of cholinergic activity) induced by WBV.
Quantification of the ChAT results showed significant differences in weeks 2 and
5. To examine whether a different marker would show a similar pattern, we
performed an additional exploratory staining to assess synaptic density of two
representative subregions: the molecular layers of DGI and DGO blades.
Synaptophysin was stained and analyzed on either week 2 or 5 in the DGI and DGO
subregions. Synaptophysin did not show significant alteration after 2 weeks of
WBV despite a reduction in expression (Fig. 6E,F). In contrast, similarly to ChAT
immunostaining, synaptophysin was significantly higher in the WBV-treated group
after 5 weeks than in the home cage controls (DGO: p
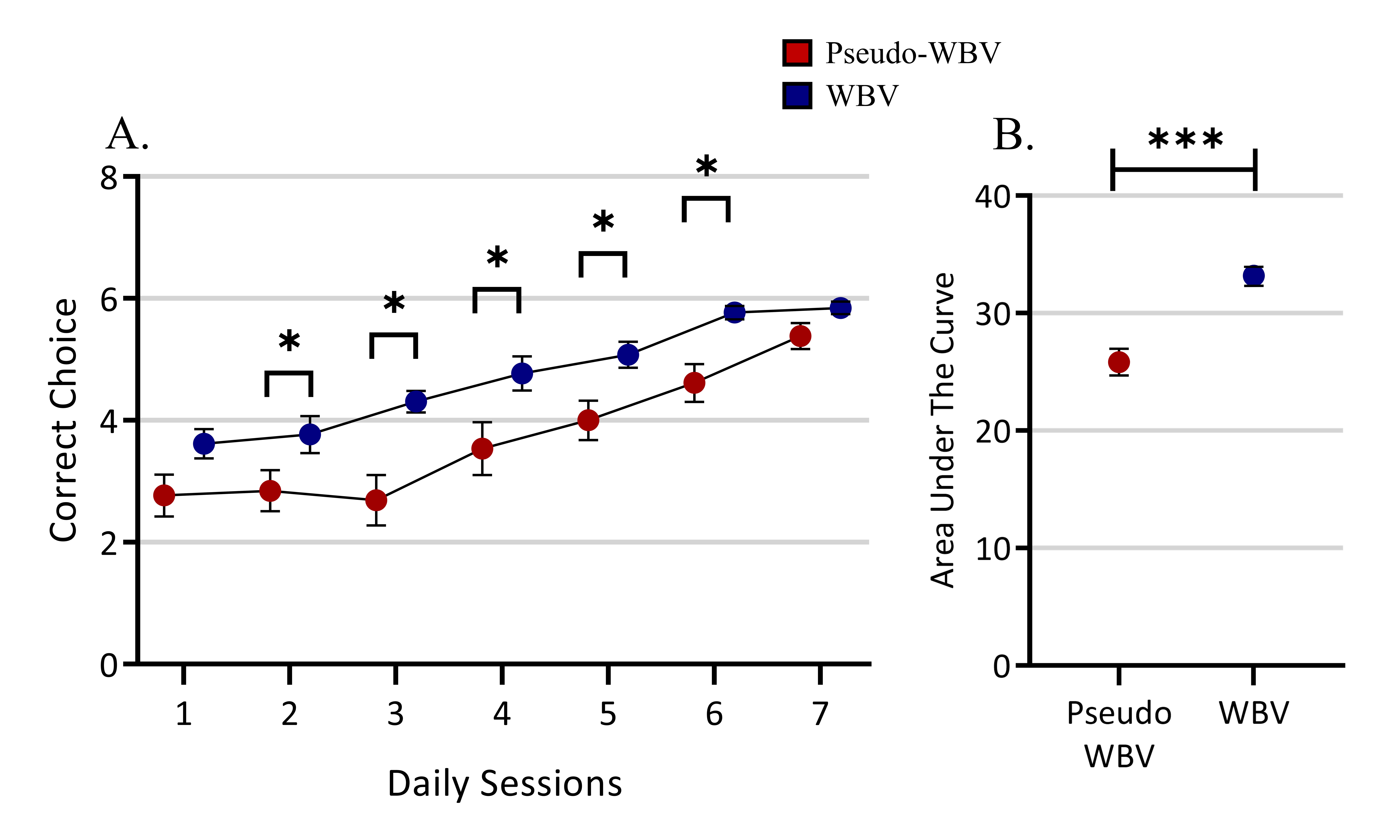
In Experiment 3, we investigated the effects of a 4-week-long WBV intervention
on spatial reference memory functions in a Y-maze task. Spatial memory
performance was not significantly different in the WBV and pseudo-WBV control
groups in the first and last sessions of the Y-maze task. However, significantly
more correct choices were made by the WBV-treated group than by the pseudo
WBV-treated group from the second session to the sixth session (session 2–6
p
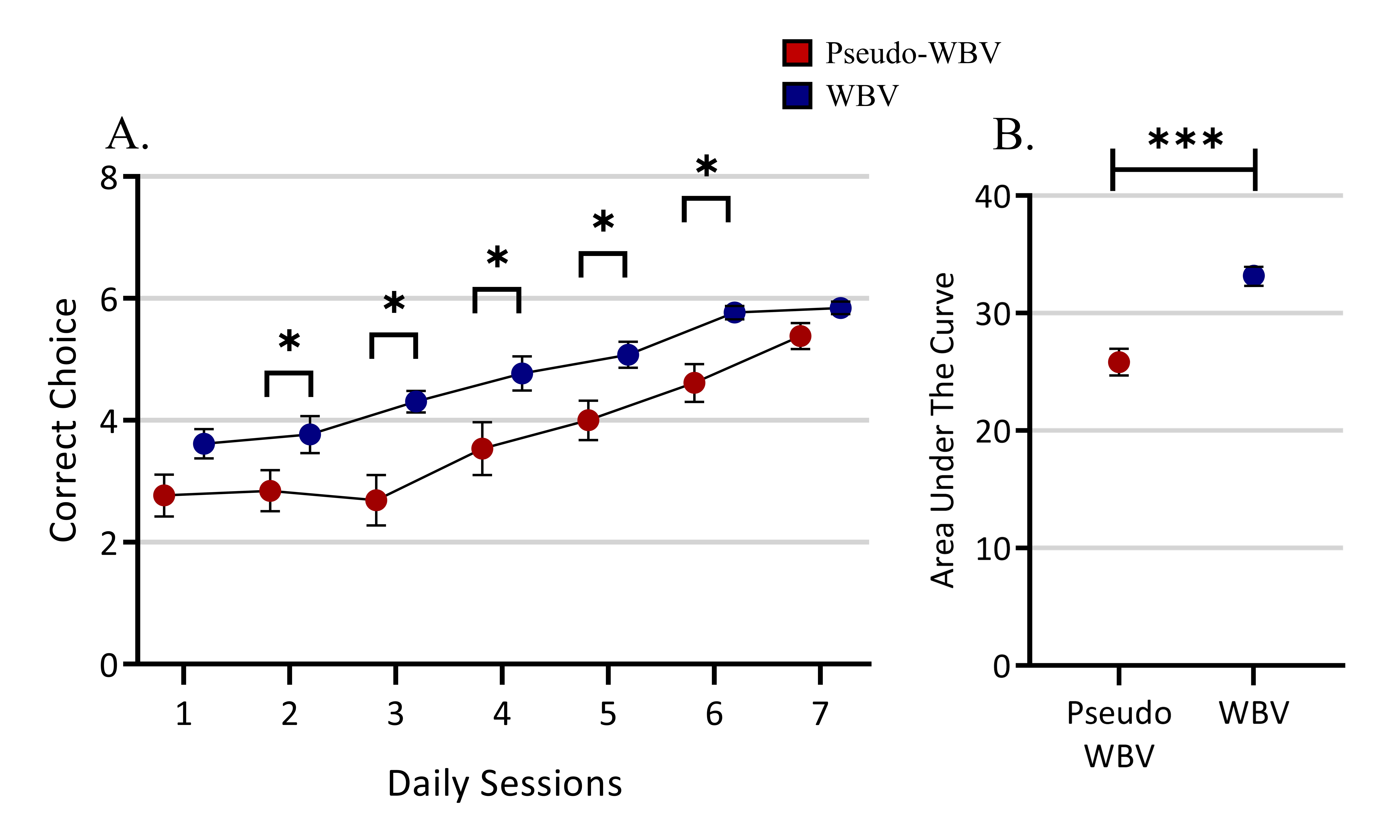 Fig. 8.
Fig. 8.
Experiment 3—Spatial reference memory performance in the
Y-maze. Effects of 5 weeks of whole-body vibration (WBV) intervention on
spatial reference memory functions in the Y-maze task.
WBV-treated mice made significantly more correct choices from session 2 to
session 6 than did the pseudo WBV-treated controls (session 2–6: p
The purpose of this study on the hippocampal cholinergic system, using ChAT as a marker, was to continue and to extend the findings of our previous study of ChAT in the cortex and amygdala (cholinergic fibers arising from the nucleus basalis innervating the cortex and amygdala) [29]. The current study was designed to address three research issues: (1) the dynamics of WBV on ChAT levels at either 2 or 24 h after the last WBV session of a 5-week-long intervention (Experiment 1); (2) the longitudinal impact of 1 to 5 weeks of WBV intervention on ChAT levels (Experiment 2); and (3) the effects of WBV (5 weeks) on spatial reference memory functions in the Y-maze task (Experiment 3). In addition, we examined hippocampal synaptophysin expression, body-weight development, and motor coordination (Experiment 2). The results showed that the septum and the DG responded with an increase in ChAT-immunostaining in the group terminated at 2 h after WBV. Notably the CA1 and CA3 pyramidal layers responded 24 h after WBV, as did the DGI in the granular layer. Furthermore, our longitudinal experiment showed that cholinergic activity had a dynamic fluctuation during the 5-week period of WBV intervention. WBV increased ChAT and synaptophysin immunoreactivity after 5 weeks. In contrast, WBV seemed to attenuate ChAT levels after 2 weeks of WBV. The WBV intervention did not affect body weight, but improved motor coordination from the third week on. In Experiment 3, we found that WBV improved spatial reference memory functions in the Y-maze task. Taken together, our data demonstrated that in young mice, 10 min WBV, daily, with low intensity and of a sinusoidal nature for 5 consecutive weeks, was able to stimulate the cholinergic system and synaptic density in the hippocampus, as well as memory functions and motor coordination.
Our findings of improved motor coordination are in line with our previous studies using devices that generate mechanical vibration with a frequency of 30 Hz and an amplitude of 50–200 µm. We should note that the present findings primarily show the dynamics of balance-beam improvement, rather than a direct WBV-specific improvement. Several studies focusing on the direct effects of WBV interventions on motor coordination in rodents have been reported; using the same vibration platform and similar protocols, we enhanced motor coordination after 5 weeks of WBV, as was found in young C57BL/6 mice [20, 29] and CD1 mice [40]. We previously found similar improvements in the motor coordination of aged rats after 5 weeks of WBV [15]. Furthermore, increased muscle strength and healing had been reported after short- and long-term (from 2 to 4 weeks) WBV interventions by others [43, 44]. In the present study, we found increased motor coordination after 3 weeks of WBV, which seems to be consistent with the existing literature. That finding indicates that the total duration of the WBV intervention is an important factor. We conclude that rodents need to undergo at least 3 weeks of WBV to obtain long-term effects on neuromuscular performance, at least when using the current WBV protocol. Hence, when using a duration shorter than 3 weeks, it might be that the impact of WBV is suboptimal or even slightly adverse if 10-min daily sessions are used.
ChAT immunohistochemistry was used to indirectly assess the level (or potential) of cholinergic neurotransmission in cholinergic fibers/terminals in the hippocampus and in the medial septum. Five weeks of WBV intervention induced an increase of ChAT optical density in the Dentate Gyrus (DG), CA3, and CA1 hippocampal subregions, as well as in the medial septum. The medial septum projects through the fornix to the hippocampus, forming the septo-hippocampal pathway. Since the level of ChAT immunoreactivity correlates positively with cholinergic activity [45], our finding indicates a higher level of activity of these cholinergic cells, and hence increased acetylcholine release upon demand. It was found that ChAT expression was significantly increased in the axons and dendrites of the medial septum in the group that was terminated 2 h after the last WBV session, but it was not significantly altered in the cell bodies. This higher level of ChAT in the dendrites and can be explained by ChAT being produced within the soma and transported to the nerve terminals to synthesise acetylcholine. The observation that ChAT in the medial septum was significantly higher in the 2-h-terminated group suggests that ChAT synthesis is a rapid response to a WBV session. Since ChAT levels were significantly higher in the hippocampus at both in both the 2 h and 24 h groups, a relatively rapid but also longer-lasting transport to the terminals is suggested. It is currently unclear what the impact of a longer duration of our WBV protocol would do to ChAT levels, although the results do not suggest that a ceiling effect in ChAT levels had been reached. The current ChAT observations are in agreement with our previous finding of increased ChAT levels (and hence most likely cholinergic activity) in the cortex and amygdala after 5 weeks of WBV [29]. Apparently different cholinergic subsystems (i.e., the nucleus basalis and medial septum) respond similarly to WBV. The entry point by which WBV activates the cholinergic cells is currently unclear, besides, the vibratory stimuli from the periphery reaches the thalamus and subsequently the motor and somatosensory cortical areas that send out projections to other cortical and subcortical regions including the hippocampus [46, 47]. Most likely, the thalamus also connects to the ascending cholinergic system, stimulating hippocampal functioning and contributing to attention, an integral aspect of learning and memory. Improved cognitive functioning by WBV can be (in part) explained by this projection. Moreover, the DG is functionally one of the most prominent hippocampal regions, facilitating neurogenesis throughout adult life [48]. Cholinergic stimulation is known to have a positive effect on both young and adult hippocampal neurogenesis and functioning [49]. Also, long-term active exercise has been demonstrated to be a significant strategy for inducing neurogenesis [50], and for attenuating the loss of cholinergic fibers innervating the hippocampus [31, 51]. In contrast, ChAT optical density was significantly reduced after 2 weeks of WBV. This paradoxical result suggests that the increase in ChAT expression over time is not a linear process but depends on some form of depletion followed by a compensatory increase. If each WBV session induces a boost in acetylcholine release requiring subsequent ChAT activity to bring the acetylcholine levels back to baseline, it could come with a temporal lack of sufficient amounts of ChAT followed by de novo synthesis of ChAT, which increases with the number of WBV sessions. This dip in ChAT may relate to exercise-induced fatigue and to its recovery; and might be an important aspect to take into consideration for planning short-term WBV protocols in future studies. Whether ChAT expression could be even further enhanced by additional WBV sessions after week 5 remains currently unknown but seems possible, providing room for optimizing this WBV protocol.
Significantly higher levels of synaptophysin were detected after 5 weeks of WBV, but not after 2 weeks. Lower levels of synaptophysin were visible after 2 weeks, similar to results detected in the ChAT activity after 2 weeks. This finding indicates an improved degree of synaptic density and most likely remodeling after 5 weeks of WBV [52]. A number of studies have shown upregulated hippocampal synaptophysin after aerobic and resistance exercise [53], as well as after short-term environmental enrichment [54]. Furthermore, active exercise counteracts decreased expression of synaptophysin in stress [55], stroke [56], and cortical infarcts [57]. Our data further support the effectiveness of WBV as a therapeutic intervention for the brain in the context of enhanced synaptic density and, hence, synaptic plasticity. This increase in level of synaptophysin could contribute to the reported WBV-induced plasticity in hippocampal slices [22, 23].
The Y-maze test was used here to evaluate spatial reference memory functions in response to WBV intervention. The number of correct choices was significantly increased from the second session to the fifth session in the WBV-treated group. This difference was most pronounced in the second and third sessions. In contrast, there was no difference from controls in the first and last training sessions. This result suggests that the WBV-treated mice have a significantly improved learning curve and hence better spatial reference memory. Several studies have demonstrated the beneficial effects of WBV on spatial memory and hippocampal functioning in aging [15], in postoperative cognitive decline, and in the models of stroke [14, 16] and restraint stress [13]. The modulation of the cholinergic system plays an essential role in hippocampal memory functions including memory consolidation and retrieval [58, 59]. This improvement in spatial memory functions, at least in part, could be explained by the observed increase in hippocampal cholinergic activity. Taken together, the data suggest that WBV, over time, strengthens the cholinergic system by enhancing the presence of the rate-limiting enzyme ChAT. Upon demand, more acetylcholine can be released, improving the rate of acquisition as well as boosting memory processes. The hypothesis of Y-maze-induced acetylcholine release could be tested by way of microdialysis, which could be a next step in this line of research.
The current results corroborate the view that WBV can be seen as a form of passive exercise. Ample data have demonstrated various effects of active exercise on hippocampal functioning in rodents. It is in line with the finding that traditional active-exercise interventions (i.e., treadmill running, wheel running, rotarod running, or strength training performed on a vertical ladder) positively impact the hippocampus [60, 61, 62, 63, 64, 65]. A recurrent finding is that active-exercise interventions promote cell proliferation and neurogenesis in the dentate gyrus [62, 66, 67]. These exercise-induced hippocampal changes seem to be associated with enhanced synaptic plasticity and improved spatial learning [64, 67, 68, 69]. In the case of WBV, no additional physical activity is required, which makes it an interesting and valuable intervention for those not able or willing to perform active exercise.
Some limitations of the present study need to be addressed. The control mice in Experiment 2 did not receive pseudo-WBV treatment, which most likely also affects ChAT activity to some extent due to handling, the experience of novelty, stress, and increased physical activity. Hence, the difference between the WBV and the control group cannot be considered vibration-specific. However, we chose to avoid these pseudo-WBV-related factors as they do not contribute to the primary aim of the second experiment (providing a longitudinal profile, which requires the most stable baseline control). Of note: a pseudo-WBV group was used as a control in the first and third experiments, revealing the vibration-specific effects on ChAT and on spatial memory. Nevertheless, this discrepancy might contribute to a more complicated comparison of the actual change in ChAT immunostaining in Experiments 1 and 3 vs Experiment 2. Furthermore, we note that we only used immunohistochemistry as a technique to visualize proteins, which allows for a semi-quantitative approach only to determine protein concentration. Perhaps future experiments could use other methods such as microdialysis, activity assays, western blotting or electrophysiological measurements to further quantify the level of cholinergic activity and degree of synaptic plasticity.
Finally, the strength of our current study is its integrative experimental design that results in consistent findings based on three separate exploratory experiments with emphasis on: (1) the validation of the time points of termination; (2) the weekly changes in ChAT expression and motor coordination during the 5 weeks of WBV; and (3) the beneficial consequences of this WBV intervention for spatial reference memory performance, to which the changes in expression of ChAT and synaptophysin most likely contribute.
Our data demonstrated that daily 10-min WBV sessions for 5 consecutive weeks, using mechanical vibration with a frequency of 30 Hz and (vertical) amplitude of 14–54 µm, is an effective strategy, at least in young mice, to improve cholinergic activity, synaptic density, and spatial memory functions. Our data shed new light on the longitudinal development of this improvement, with a temporal decline followed by recovery and subsequent enhancement of ChAT expression. Furthermore, our findings indicate that the cholinergic system can respond rapidly to WBV stimuli that seem to last, at least, for 24 h. The data are consistent with those in the literature that show that WBV can be an effective treatment strategy to stimulate the brain, including the hippocampus, and to improve cognition and motor coordination.
Taken together, these exploratory experiments provide support for the relevance of passive WBV exercise as a non-pharmacological strategy to stimulate the cholinergic system and synaptic density. This suggest that WBV may be considered a therapeutic strategy in neurodegenerative disorders such as Parkinson’s, Alzheimer’s, or other, brain diseases. The improvements in brain functioning reported after different WBV interventions may be explained, at least in part, by the improved septo-hippocampal cholinergic system function and hippocampal plasticity. Future studies, using integrative experimental designs, need to focus on how the cholinergic system could be upregulated by WBV in animal models with pathological brain conditions.
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
ChAT, Choline acetyltransferase; WBV, Whole-body vibration; AUC, Area Under the Curve; DG, Dentate Gyrus; TBS, Tris-Buffered Saline; TX, Triton X-100; DAB, diaminobenzidine; DGI, Dentate Gyrus Inner-blade; DGO, Dentate Gyrus outer blade; CA1, Cornu Ammonis 1; CA3, Cornu Ammonis 3.
The study conception and design were contributed by EAvdZ, MJGvH, CN. Material preparation, data collection and part of the analysis were performed by WH, JNK. The first draft of the manuscript, literatures search, interpretation of the data, design of the figures and part of the data analysis were performed by TO. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All experimental procedures were evaluated and approved by the national Central Authority for Scientific Procedures on Animals (CCD) and by the local Institutional Animal Welfare Body of University of Groningen (approval number: DEC5685).
We thank Folkert Postema for his valuable contribution to this study and Dr. Gernot Riedel for providing the WBV equipment.
This research received no external funding.
The authors declare no conflict of interest. Eddy A. van der Zee is serving as one of the Editorial Board members. We declare that Eddy A. van der Zee had no involvement in the peer review of this article and has no access to information regarding its peer review. Full responsibility for the editorial process for this article was delegated to Gernot Riedel.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.