




1 Department of Infectious Diseases and Microbiology, School of Public Health, University of Pittsburgh, Pittsburgh, PA 15261, USA
†These authors contributed equally.
Abstract
Infection of astrocytes by Human Immunodeficiency Virus (HIV-1) remains a topic of debate, with conflicting data, yet instances of astrocytes containing viral DNA have been observed in vivo. In this study, we aimed to elucidate potential routes through which astrocytes could be infected and their ability to produce infectious particles using primary human astrocytes.
We infected primary astrocytes derived from either neuroprogenitor cells (NPCs) or induced pluripotent stem cells (iPSCs) that express both C-X-C chemokine receptor type 4 (CXCR4) and the C-C chemokine receptor type 5 (CCR5) coreceptors, using either cell-free HIV-1 virus directly or cell-associated virus indirectly through infected macrophages and microglia.
Low-level infectivity by cell-free viruses was primarily attributed to a defect in the entry process. Bypassing HIV-specific receptor-mediated entry using pseudotyped viruses resulted in productive infection and the release of infectious particles.
These findings suggest that astrocytes may be one of the potential sources of neurotoxicity in HIV-associated neurocognitive disorders (HAND) and could possibly act as reservoirs for HIV in the central nervous system (CNS).
Keywords
- HIV-1 neuropathogenesis
- astrocytes
- virus replication
- iPSCs-derived microglia
- macrophages
Human Immunodeficiency Virus (HIV-1) enters the central nervous system (CNS) early in infection through infected circulating monocytes, a process described by the ‘Trojan Horse’ model [1, 2]. Once these monocytes cross the blood-brain barrier, they differentiate into infected brain macrophages [3]. Macrophages can then sustain productive HIV-1 infection potentially infecting and/or exposing other glial cells and triggering neuroinflammatory responses [4, 5]. HIV-induced neuroinflammation has been identified as one of the primary causes of neurodegeneration in HIV patients [6]. Astrocytes, the most abundant type of glial cells in the brain, fulfill numerous supportive functions in the CNS and play a critical role in maintaining the blood-brain barrier [7, 8]. Astrocytes, along with microglia, have been implicated as potential targets or reservoirs of HIV in the CNS, although this remains a topic of controversy [9, 10, 11, 12].
Microglia and macrophages have been identified as the primary target cells for HIV-1 in the CNS in numerous studies [9, 13, 14]. However, the potential for astrocytes to be directly infected with HIV and whether they can produce replication-competent virus remains unclear. Several studies have indicated that astrocytes likely lack the CD4 receptor required for infection [4, 15, 16]. Nevertheless, in vivo detection of viral genetic material in astrocytes has been reported [10, 17], suggesting that astrocytes may contribute to the viral reservoir in the CNS.
One hypothesis proposes that astrocytes may ingest material from infected macrophages or microglia and slowly release viral components over time, acting as a continuous source of viral material that does not rely on reverse transcription [18]. Another study suggested the possibility of HIV infecting astrocytes via a CD4-independent pathway [16], while a third study described HIV transmission from lymphocytes to astrocytes through a C-X-C chemokine receptor type 4 (CXCR4)-dependent/CD4-independent mechanism [19]. However, the question of whether astrocytes can be infected with HIV has been contested. Early findings suggested that astrocytes were impermeable to cell-free HIV and could not be infected with either M- or T-tropic strains [15]. Other studies have failed to detect viral genetic material in ex vivo analyses of astrocytes isolated from infected brain tissue [9] and have not observed any membrane fusion interactions between astrocytes and virions [18].
In this study, we investigated the susceptibility of primary astrocytes derived
from either neural progenitor cells (NPCs) or induced pluripotent stem cells
(iPSCs) to HIV-1 infection and their ability to produce infectious virus
particles. Our findings demonstrate that both models of human astrocytes are
capable of supporting productive HIV-1 infection, albeit at a very low level
(
Primary human astrocytes were differentiated from human neuroprogenitor cells (NPCs, ABC-TC372-1), purchased from Accegen Biotech (Fairfield, NJ, USA) using the protocol developed by Hammond et al., 2002 [20]. The NPCs were confirmed to express the neural stem-cell marker Nestin and passed routine mycoplasma contamination tests and the test results were negative. Briefly, NPCs were plated on poly-D-lysine (PDL)-treated 6-well plates with glass coverslips and cultured in 2 mL of 90% KnockOut DMEM, 10% fetal calf serum (FCS), 0.1% L-Glutamine, and 0.1% penicillin/streptomycin (Gibco, Grand Island, NY, USA). Half of the media was replaced every 48 hours. NPCs were allowed to differentiate for four weeks until they exhibited a full astrocyte phenotype. Aliquots of differentiating NPCs were characterized every 7 days. Confirmation of the astrocyte phenotype was performed based on morphology and quantitative real time-polymerase chain reaction (qRT-PCR) analysis of Glial Fibrillary Acidic Protein (GFAP) transcripts four weeks post-differentiation and results are presented in Fig. 1A,B.
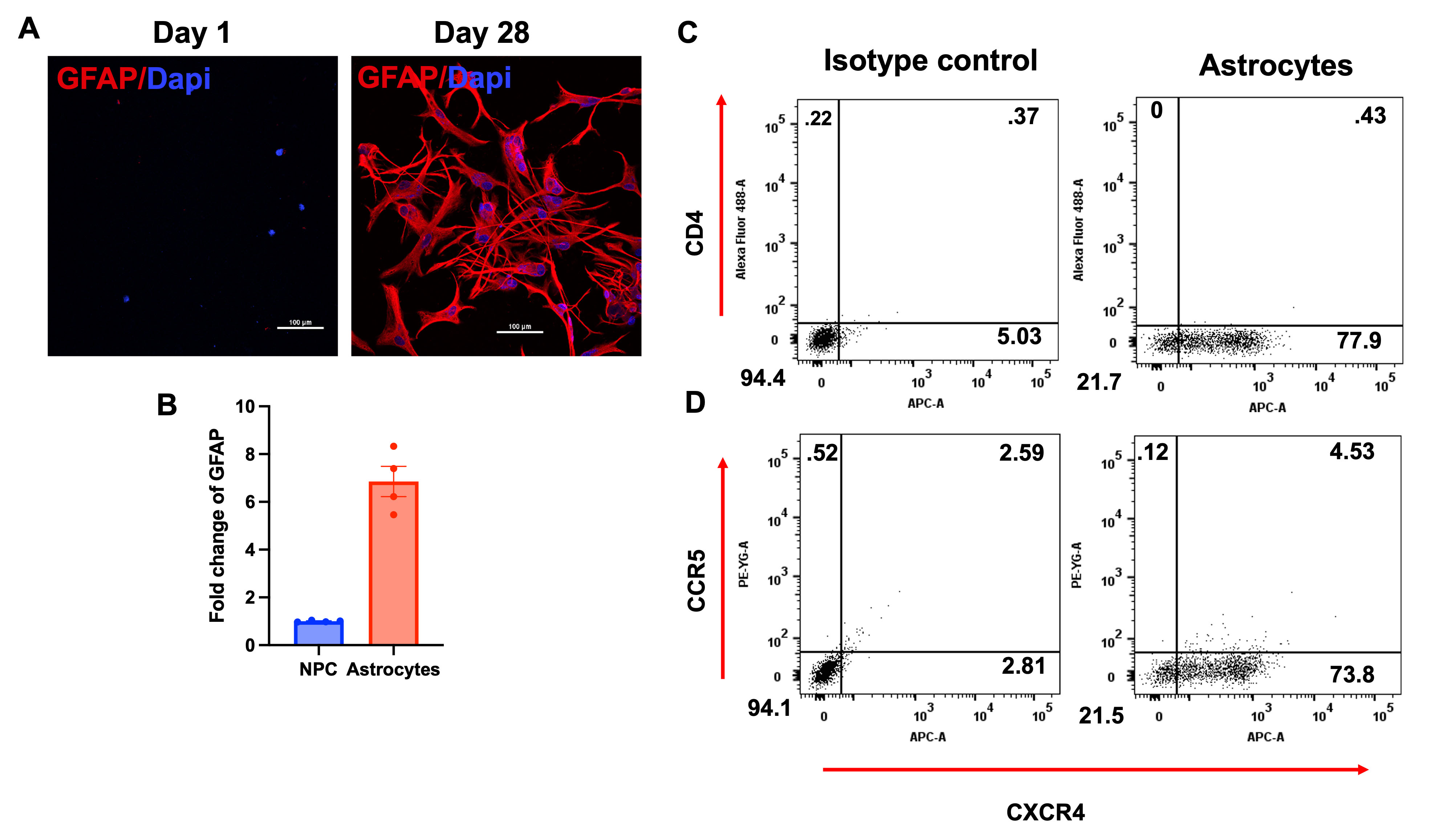 Fig. 1.
Fig. 1.
Generation and characterization of primary astrocytes. (A) Confocal images of NPCs and differentiated astrocytes expressing the astrocytic marker GFAP (red) at day 1 (left) and day 28 (right). (B) Quantification of GFAP expression by NPCs and Astrocytes through qRT-PCR. (C) Flow cytometry of Astrocytes stained for CD4-FITC and CXCR4-APC (right), versus isotype control (left). (D) Flow cytometry of primary astrocytes stained for CCR5 and CXCR4 coreceptor (right), versus isotype control (left). Numbers reflect percentage of cells in that quartile. N = 4, scale bar = 100 µm. Abbreviations: DAPI, 4′,6-diamidino-2-phenylindole; GFAP, Glial Fibrillary Acidic Protein; NPCs, neuroprogenitor cells; FITC, Fluorescein isothiocyanate; CCR5, C-C chemokine receptor type 5; CXCR4, C-X-C chemokine receptor type 4; APC, allophycocyanin; qRT-PCR, quantitative real-time polymerase chain reaction.
Healthy donor-derived human iPSC line (BYS0113, ATCC, Manassas, VA, USA) was cultured on dishes coated with hESC-qualified Matrigel in mTeSR medium (STEMCELL Technologies, Vancouver, Canada). Neural progenitors were generated from iPSC cultures following the protocol described by Jin et al. [21]. These neuroprogenitors were then differentiated into astrocytes over three weeks using the STEMDiff Astrocytes differentiation kit (STEMCELL Technologies, Vancouver, Canada), as per the manufacturer’s instructions. The differentiated astrocytes were subsequently matured for an additional three weeks using the STEMDiff Astrocytes maturation kit (STEMCELL Technologies, Vancouver, Canada), also following the manufacturer’s instructions. The matured iPSC-derived astrocytes (iAstrocytes) were used for experiments within 2–3 weeks post-maturation. For the generation of macrophages and microglia (myeloid) precursors from the human iPSC line (ATCC, BYS0113), we utilized the STEMDiff Hematopoietic differentiation kit (STEMCELL Technologies, Vancouver, Canada), following the manufacturer’s instructions. Myeloid precursors were differentiated into macrophages over a period of two weeks in a medium composed of DMEM/F12 (Gibco, Grand Island, NY, USA) supplemented with 2 mM GlutaMAX (Gibco, Grand Island, NY, USA) and 100 ng/mL macrophage colony-stimulating factor (M-CSF) (Peprotech, Cranbury, NJ, USA), according to Haenseler et al. [22]. Similarly, myeloid precursors were differentiated into microglia over three weeks in a medium composed of DMEM/F12 supplemented with 2 mM GlutaMAX (Gibco), 100 ng/mL Interleukin-34 (IL-34) (Peprotech, Cranbury, NJ, USA), and 10 ng/mL granulocyte-macrophage colony-stimulating factor (GM-CSF) (Millipore, Burlington, MA, USA), as previously established [22]. Once in maturation medium, iPSC-derived cells lose their ability to proliferate and become terminally differentiated. STR authentication and karyotyping of the iPSC line were performed by ATCC (Manassas, VA, USA). Cells were regularly tested for mycoplasma contamination the test results were negative, indicating that the cells were free of contamination. Cells were used within 30 passages for this study.
Flow cytometry was employed to characterize the expression of surface markers on human primary astrocytes crucial for binding and fusion, including CD4, C-C chemokine receptor type 5 (CCR5), and CXCR4. Astrocytes were dissociated using TripLE (Gibco, Grand Island, NY, USA) non-enzymatic dissociation reagent, transferred to clear flow tubes, and centrifuged at 1000 RPM for 5 minutes. After aspirating TripLE, cells were incubated with monoclonal wash solution (90% PBS, 10% FCS, 0.1% sodium azide) containing CD4-FITC (Fluorescein isothiocyanate), CCR5-phycoerythrin (PE), and C-X-C chemokine receptor type 4-allophycocyanin (CXCR4-APC) antibodies. Following a 45-minute incubation at 4 °C, cells were washed and fixed with 4% paraformaldehyde (PFA) (ThermoFisher Scientific, Waltham, MA, USA). Stained cells were analyzed using a FACSymphony A5 SE (BD Biosciences, Franklin Lakes, NJ, USA) with isotype-stained cells serving as controls for analysis.
To confirm the differentiation of primary astrocytes from NPCs, cells were stained for GFAP, an astrocyte-specific marker. Astrocytes plated on coverslips were fixed in 4% PFA for at least 15 minutes and permeabilized in a buffer containing 90% PBS, 10% FCS, and 0.1% Triton X-100. Next, cells were incubated with 1:1000 dilution of chicken anti-GFAP primary antibody (Abcam, Cambridge, UK) at 4 °C for 24 hours, followed by washing and staining with 1:500 dilution of the secondary antibody, Goat anti-Chicken Cy5 (Jackson ImmunoResearch Labs, West Grove, PA, USA). Slides were washed twice in PBS, stained with 4′,6-diamidino-2-phenylindole (DAPI), and mounted on coverslips using glycerol-based mounting media. Images were acquired using a Nikon A1-R confocal microscope (Nikon, Tokyo, Japan), and maximum intensity Z-projections were generated using ImageJ-FIJI software (version 2.14.0, National Institutes of Health, Bethesda, MD, USA). The images shown are representative of cultures generated from three independent experiments.
HEK-293T cells (CRL3216, ATCC, Manassas, VA, USA) were transfected with 5 µg of HIV-1 viral plasmids (pNL43 for X4-tropic, pNL43-YU2 Env or pNL43-BaL Env for R5-tropic) using PolyJet transfection reagent (SignaGen, Frederick, MD, USA) as described [23]. To identify productively infected cells, HIV-1 proviral plasmids were constructed to express fluorescent reporters: enhanced green fluorescent protein (EGFP) in plasmids pNL43 and pNL43-YU2 Env-EGFP, or near-infrared fluorescent protein (iRFP) in plasmid pNL43-BaL Env-iRFP. Forty-eight hours post-transfection, the supernatant was harvested, centrifuged to remove cellular debris, filtered through a 0.2-micron syringe filter (ThermoFisher Scientific, Waltham, MA, USA), and stored at –80 °C. To enhance astrocyte tropism, pseudotyped viruses were generated by cotransfecting 1.50 µg of Vesicular Stomatitis Virus Glycoprotein (pVSV-G-Env) expression plasmid with the above-mentioned proviral DNA [24]. Pseudotyped HIV particles interact with ubiquitous phosphatidylserine and low-density lipoprotein receptors (LDLRs) on the target cell surface, inducing endocytosis of the virion into the astrocyte [25]. All viruses were titrated in U87MG CD4+ CCR5+ cells (ARP-4035, NIH-AIDS Reagents Program, Germantown, MD, USA) to determine the multiplicity of infection (MOI). The cell species identification was carried out by ATCC (Manassas, VA, USA) through Cytochrome oxidase 1 (CO1) gene barcoding. All cell lineages utilized in this study were routinely tested for mycoplasma contamination with all outcomes being negative.
Four weeks post-differentiation, astrocytes were infected with 0.1 or 1 MOI of virus. Eighteen hours post-infection, cells were washed with PBS to remove unbound virus and maintained in astrocyte medium. Supernatant was collected every 48 hours until 10–15 days post-infection, and coverslips were collected for imaging. The remaining cells were harvested for RNA extraction using the MirVana kit (ThermoFisher Scientific, Waltham, MA, USA) following the manufacturer’s recommendations. RNA concentration and purity were assessed using a NanoDrop 2000 spectrophotometer (ThermoFisher Scientific, Waltham, MA, USA). cDNA was synthesized from 300 ng of total RNA using a high-capacity cDNA reverse transcription kit (ThermoFisher Scientific, Waltham, MA, USA) in a total reaction volume of 20 µL. qRT-PCR was performed using TaqMan Universal PCR master mix (ThermoFisher Scientific, Waltham, MA, USA) and the appropriate TaqMan assays or primers (refer to Table 1) with 2 µL of the cDNA reaction mixture. PCR assays were conducted on an ABI ViiA 7 real-time PCR system (Applied Biosystems, Waltham, MA, USA) with the following cycling conditions: initial activation of Taq DNA polymerase at 95 °C for 10 min, followed by 40 cycles of denaturation at 95 °C for 15 s and annealing/extension at 60 °C for 1 min. Results were normalized to the expression of the endogenous control Ribosomal Protein Lateral Stalk Subunit P0 (RPLP0).
| Target gene | Description | 5′-3′ Sequence |
| GFAP | Assay ID | Applied Biosystems TaqMan® Gene Expression Assays ID: Hs00909233_m1 |
| RPLP0 | Assay ID | Applied Biosystems catalog number 4310879E |
| SS-Gag | Primer F | TCTCTAGCAGTGGCGCCCGAACA |
| Primer R | TCTCCTTCTAGCCTCCGCTAGTC | |
| Probe | CGGGAG TACTCACCAGTCGCCGCCCCTCGCC CTCCCG | |
| MS-Gag | Primer F | CTTAGGCATCTCCTATGGCAGGAA |
| Primer R | TTCCTTCGGGCCTGTCGGGTCCC | |
| Probe | GGGCCTTCTCTATCAAAGCAACCCACCTCCAGGCCC |
Abbreviations: GFAP, Glial Fibrillary Acidic Protein; RPLP0, ribosomal protein lateral stalk subunit P0; SS-Gag, single-spliced Gag; MS-Gag, multiply spliced Gag.
To assess the role of cell-associated infection of human astrocytes, we co-cultured mature iAstro either with HIV- infected iMac or iMG in a 1:1 ratio. Briefly, iMac and iMG cultured in 6 well-plates were infected with HIV-BaL-Env-iRFP MOI 1 for 18 hrs, washed and maintained in medium for 10 days. Cells were then harvested, counted and added to mature iAstro previously labeled with Calcein and Hoechst (Thermo-Fisher Scientific, Waltham, MA, USA) per manufacturer’s recommendation. Co-cultures (1:1 ratio) were maintained for an additional 7 days and coverslips were harvested and fixed for at least 15 min in 4% PFA, and permeabilized. Cells were incubated with 1: 1000 dilution of rabbit Iba-1 primary antibody (Fujifilm Wako Chemicals, Richmond, VA, USA) at 4 °C for 24 hrs, followed by washing and staining with secondary antibody, Goat Ab to rabbit-Cy3 (Jackson ImmunoResearch Labs, West Grove, PA, USA), in a 1:500 dilution of buffer. The slides were washed twice in PBS and mounted on coverslips using glycerol-based mounting media. The remaining cells in the wells were harvested with TripLE, washed, fixed with 4% PFA and subjected to flow cytometry to quantitate infectivity.
Culturing and differentiation of NPCs into astrocytes were characterized by morphological changes and the expression of GFAP, an astrocyte-specific marker, over a period of four weeks (Fig. 1A,B). Subsequently, we assessed the expression of the HIV receptor (CD4) and coreceptors (CCR5 and CXCR4) using flow cytometry and specific markers. The results indicate a lack of CD4 and CCR5 expression, while more than 75% of cells express CXCR4, compared to the isotype control (Fig. 1C,D). These findings suggest that fully differentiated primary astrocytes do not express the canonical HIV receptor CD4 or the CCR5 co-receptor on their cell surface. Instead, over 75% of the cells express CXCR4, indicating that neurotropic viruses (R5-tropic) likely enter these cells through alternative pathways.
To evaluate the susceptibility of primary astrocytes to HIV infection and their
ability to produce infectious virions, we exposed these cells to both R5- and
X4-tropic HIV strains. Astrocytes were infected with HIV-1 YU2 EGFP
(CCR5-utilizing strain) and HIV-1 NL43 EGFP (CXCR4-utilizing strain) reporter
viruses with and without VSV-G-Env complementation, as described in the methods.
Infectivity was monitored by assessing EGFP expression in cells using microscopy
in conjunction with GFAP staining (Fig. 2A,B). Results indicate very low levels
of infection (
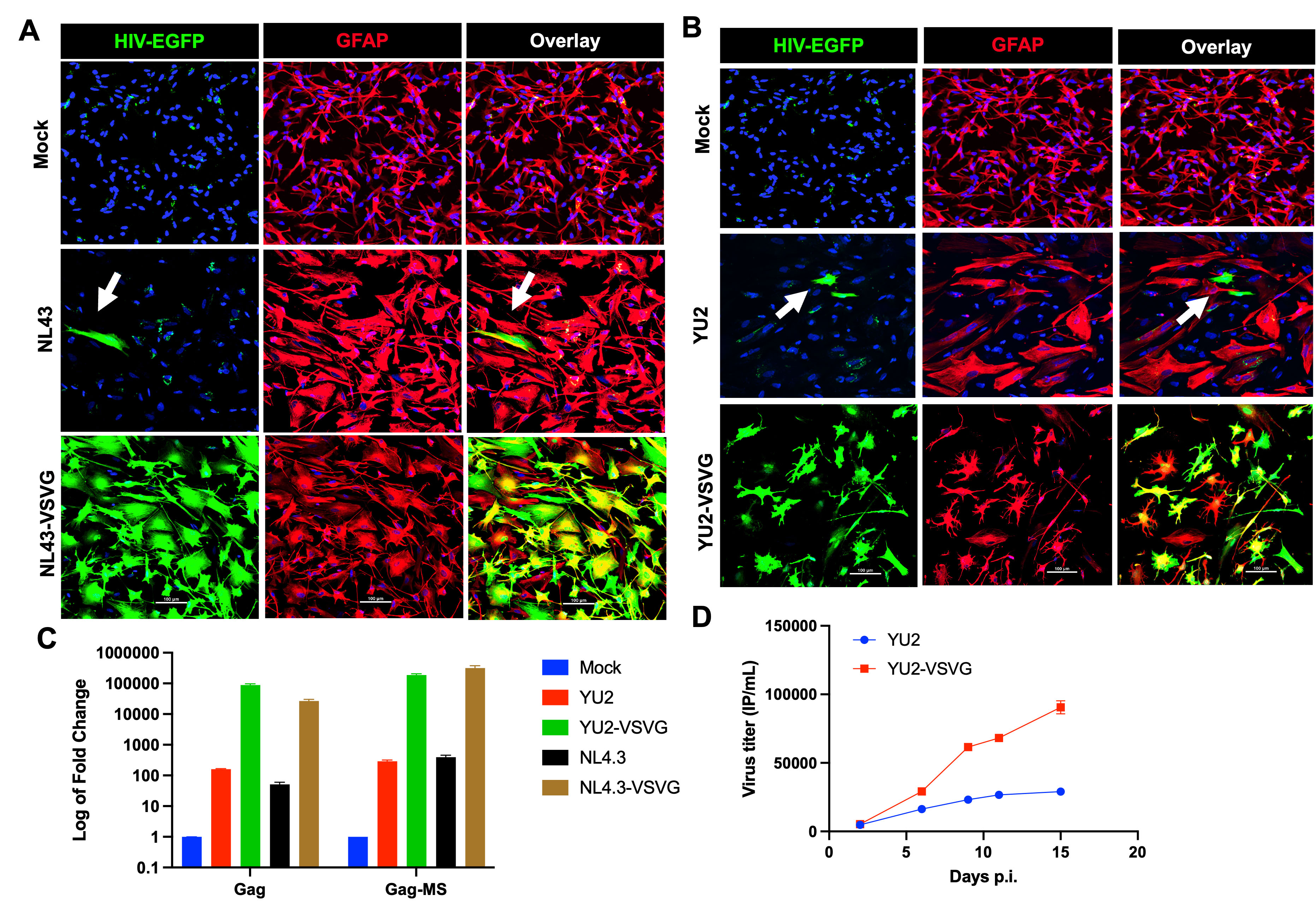 Fig. 2.
Fig. 2.
Cell-free HIV-1 infection of primary astrocytes and viral replication. (A) Confocal images of primary astrocytes stained with GFAP (red, Cy5) and Nucleus (DAPI, blue) upon exposure to HIV-1 NL43-EGFP, HIV-1 NL43-EGFP complemented with VSV-G-Env, or no virus (mock). (B) Confocal images of primary astrocytes stained with GFAP (red, Cy5) and DAPI (blue) upon exposure to HIV-1 NL43(YU2)-EGFP, HIV-1 NL43(YU2)-EGFP (green) complemented with VSV-G-Env, or no virus (mock). (C) Quantification of Gag transcript and multiply spliced transcripts in astrocytes exposed to HIV-1 at day 15, post-infection by qRT-PCR. (D) Assessment of cumulative infectious particles released in the supernatant over the period of 15 days post-infection. N = 4, scale bar = 100 µm. White arrows show HIV-infected astrocytes. Abbreviations: EGFP, enhanced green fluorescent protein; HIV-1, Human Immunodeficiency Virus; YU2-VSV-G, YU2 strain pseutotyped with the envelope glycoprotein of vesicular stomatitis virus; MS, multiply spliced; VSV-G-Env, envelope glycoprotein of vesicular stomatitis virus.
Next, we investigated whether the virus particles produced by infected astrocytes were infectious. Supernatants collected at various time points post-infection were used to infect HIV-permissive U87MG cells (Fig. 2D). U87MG cells infected with different dilutions of virus generated by astrocytes showed EGFP expression, indicating that astrocytes are capable of producing infectious virus particles up to 15 days post-infection. These results collectively suggest that once infected, astrocytes may serve as long-lived viral reservoirs.
iPSCs are emerging as promising tools for
developing more reliable in vitro models to study neuropathogenesis [24, 25]. These cells can be cultured in chemically defined media to generate various
cell lineages resembling those found in human brain tissue, including astrocytes
and microglia [26]. In this study, iPSC-derived astrocytes, referred to as
iAstro, were infected with R5-tropic HIV virus with and without the VSV-G-Env complementation.
Specifically, iAstro were exposed to HIV-1 NL43 BaL-near-iRFP reporter virus with and without VSV-G-Env complementation.
Infectivity was assessed by monitoring iRFP expression in cells using microscopy
in conjunction with GFAP staining (Fig. 3A). These results confirm very low
levels of infection (
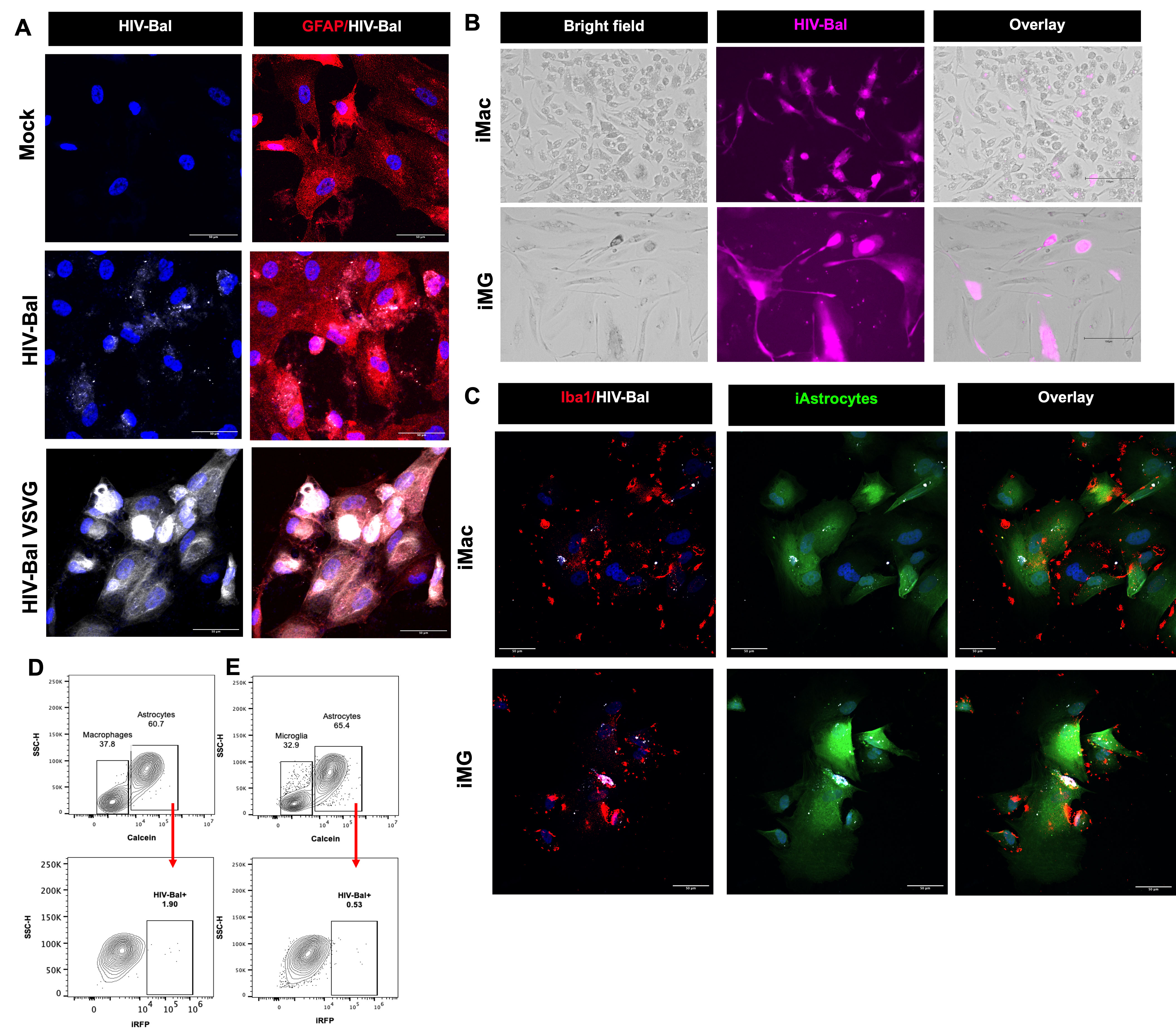 Fig. 3.
Fig. 3.
Cell-associated infection of astrocytes. (A) Confocal images of iPSC-derived astrocytes (iAstro) stained with GFAP (red, Cy3) and DAPI (blue) upon exposure to cell-free HIV-1 NL43(BaL)-iRFP, HIV-1 NL43(BaL)-iRFP (white) complemented with VSV-G-Env at 1 MOI, or no virus (mock). (B) Representative images of iPSC-derived macrophages (iMac) and microglia (iMG) upon infection with HIV-1 NL43(BaL)-iRFP (pink) at 1 MOI. (C) HIV-infected iMac and iMG were harvested 7 days post infection and co-cultured with calcein-labeled iAstro (green) for additional 10 days. Representative confocal images depict HIV-infected (white) iMac and iMG (Iba-1+, red) in close contact with iAstro (calcein+, green). Representative flow cytometry plot to identify calcein-positive iAstrocytes in cell suspension from co-culture with (D) macrophages or (E) microglia. iAstrocytes population was gated (upper panel) and further separated by iRFP expression to assess the percentage of iAstrocytes that were positive for HIV-infection (lower panel). Scale bar = 50 µm in (A,C), and scale bar = 150 µm in (B). Abbreviations: iPSC, induced pluripotent stem cells; MOI, multiplicity of infection; iRFP, near-infrared fluorescent protein; SSC-H, side scatter pulse-height.
The prevailing consensus is that astrocytes are generally resistant to HIV infection because they lack the requisite surface receptors necessary for virion fusion [18, 19]. Our flow cytometry analyses confirmed that primary astrocytes exhibit negligible levels of CD4 or CCR5, with significant expression of CXCR4. This supports previous evidence indicating that wild-type HIV cannot effectively fuse with astrocytic membranes through conventional receptor-mediated pathways [16, 18]. However, the observation of strong fluorescence in primary astrocytes and induced astrocytes (iAstro) treated with VSV-G-Env virus provides compelling evidence that the virus utilizes alternative membrane-mediated entry mechanisms to integrate its genetic material into the host astrocytes. The observation of strong fluorescence in primary astrocytes and induced astrocytes (iAstro) treated with VSV-G-Env virus provides compelling evidence that the virus employs alternative membrane-mediated entry mechanisms to integrate its genetic material into host astrocytes. Furthermore, since the fluorescent reporter gene is expressed only upon viral DNA integration, the robust colocalization of enhanced green fluorescent protein (eGFP) or near-iRFP with GFAP decisively demonstrates that, once internalized, the virus can utilize astrocytes to support viral transcription, leading to the production of viral proteins and infectious particles.
Interestingly, we observed a low level of cell-free infection of astrocytes with
wild-type NL43, BaL-Env, and YU2-Env viruses (
There has been speculation that HIV could potentially infect astrocytes through a CD4-independent pathway that is not well-defined. Schweighardt and Atwood [27] demonstrated that immortalized astrocytes (SVG-A) could be infected when exposed to wild-type X4 virus, albeit through a significantly less efficient process compared to traditional receptor-mediated fusion. They further showed that infection rates were markedly enhanced by artificially expressing CD4 and CXCR4 receptors on the membrane of SVG-A cells, indicating that the presence of the traditional receptor complex greatly facilitated infection [27]. Although cell-free HIV-1 viruses lacking VSV-G Env infected less than 1% of astrocytes, as evidenced by fluorescent signals detected via confocal microscopy and Fluorescence-activated cell sorting (FACS), we cannot rule out the possibility that additional viral particles were initially captured, internalized, and later released, as observed in dendritic cells (DCs) [28], This possibility warrants further investigation.
Furthermore, significant cytotoxicity was observed in all virus-treated astrocytes. It has been suggested that in HIV-associated neurocognitive disorders (HAND) pathology, the primary role of astrocytes may not be their potential to become infected, but rather their robust and prolonged inflammatory response when exposed to HIV [4]. This study demonstrates that regardless of their susceptibility to infection, astrocytes have the capacity to induce neuropathology through their reaction to the virus. Our findings indicate that virus-treated astrocytes exhibit a phenotype consistent with reactive astrogliosis, a typical astrocytic response to CNS insults. Reactive astrogliosis is generally aimed at limiting tissue damage; however, if this response persists excessively, it can become maladaptive, promoting neuroinflammation and apoptosis [29]. The evidence suggesting reactive astrogliosis in our study supports the hypothesis that astrocytes can detect and respond to HIV in a manner that potentially exacerbates cellular damage.
Moreover, there has been limited research into the co-culture of neurons, astrocytes, and microglia, and the potential implications of their interactions in neuro-HIV. Liddelow et al. [30] demonstrated that activated microglia could prompt astrocytes to adopt a neurotoxic phenotype similar to the proinflammatory phenotype observed in activated macrophages. Through co-culturing infected astrocytes with neurons and microglia, we aim to investigate whether infected astrocytes might induce or contribute to neuronal cytotoxicity, as suggested by previous studies [31, 32, 33]. Astrocytes play a multifaceted role in the CNS, and their involvement in HAND and dementia remains largely uncharted territory. Our forthcoming experiments seek to shed light on the role of these enigmatic cells.
Collectively, our study findings support the hypothesis that various astrocyte models can be infected with cell-free HIV, albeit at significantly low levels, despite their lacking appropriate surface receptors. Our experiments using VSV-G-Env-pseudotyped HIV unequivocally demonstrate that astrocytes can be infected when provided with a mechanism for the virus to penetrate the cell membrane. Furthermore, our results provide evidence that astrocytes may act as a reservoir for HIV, either by retaining viral material for subsequent release or potentially by becoming infected and producing new virions themselves. Additionally, our imaging data indicate that even at very low levels of infection, HIV can induce astrogliosis.
The datasets used and analyzed in the present study are available upon request from the corresponding author.
RSR and VA conceptualized and designed the experiments. RSR, SS, and MCEW conducted the research, analyzed and interpreted the data. RSR, SS, and VA drafted the manuscript. All authors contributed to the editorial revisions of the manuscript. All authors have read and approved the final version of the manuscript. All author has sufficiently contributed to the work and agrees to be accountable for all aspects of the research.
Not applicable.
We thank the NIH HIV Reagent Program, Division of AIDS, NIAID, NIH, Drs. HongKui Deng and Dan Littman for providing experimental reagents. We acknowledge the Center for Biological Imaging of the University of Pittsburgh for providing the instrumentation necessary for imaging acquisition. During the revision of this work, the authors used Grammarly AI tool in order to check spell and grammar of all the text. After using this tool, the authors reviewed and edited the Introduction and Discussion sections as needed and took full responsibility for the content of the publication.
This research received no external funding.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.