





1 Department of Child Health Care, Children’s Hospital of Chongqing Medical University; National Clinical Research Center for Child Health and Disorders, Ministry of Education Key Laboratory of Child Development and Disorders; Chongqing Key Laboratory of Child Neurodevelopment and Cognitive Disorders, 400010 Chongqing, China
Abstract
Autism spectrum disorder (ASD) is a neurodevelopmental condition characterized by deficits in social interaction, communication, repetitive behaviors, and narrow interests. This study aimed to investigate the impact of the Hypoxia-inducible factor-1 alpha (HIF-1α) inhibitor (PX-478) on ASD-like behaviors in rat offspring exposed to prenatal hypoxia (PH).
Pregnant rats were randomly assigned to control or PH groups, with the latter experiencing six hours of hypoxia on the 17th day of gestation. Offspring were further treated with PX-478 treatment initiated at one week (+1 w) or three weeks (+3 w) after birth. Hippocampal histology was assessed using hematoxylin and eosin (HE) staining, while protein levels of HIF-1α and phosphatase and tensin homolog (PTEN) were analyzed via western blotting. The concentration of vascular endothelial growth factor (VEGF) was measured using an Enzyme-Linked Immunosorbent Assay (ELISA) kit.
PX-478 treatment significantly improved spatial memory, learning, and social ability, while reducing anxiety-like behavior in PH-exposed offspring rats. HE staining revealed that PX-478 treatment decreased the number of hippocampal neurons necrosis in offspring. However, PX-478 treatment at one week post-birth led to decreased body weight and elevated levels of alkaline phosphatase (ALP) and Alanine aminotransferase (ALT) in offspring rats, whereas no significant effect was observed after three weeks of treatment. Additionally, PX-478 treatment resulted in reduced HIF-1α protein levels in the hippocampus and VEGF concentration in the serum of PH-exposed offspring rats, along with elevated PTEN protein levels.
The findings suggest that PX-478 treatment attenuated autism-like behavior in offspring. HIF-1α might play an important role in autism-like behavior induced by prenatal hypoxia, which may be realized by inhibiting PTEN activity.
Keywords
- HIF-1α
- PX-478
- prenatal hypoxia
- autism spectrum disorder
Autism spectrum disorder (ASD) is a neurodevelopmental condition characterized by deficits in social interaction, communication impairments, repetitive behaviors, and restricted interests [1]. The global prevalence of ASD has been steadily increasing annually [2], with reported rates of 2.7% among children in the United States [3] and approximately 0.7% in China [4]. Despite the rising incidence, the etiology of ASD remains elusive, and no specific pharmacological treatments are available. Additionally, the high cost of specialized care facilities places significant psychological and financial burdens on families and society [5]. Therefore, understanding the underlying mechanisms of ASD is essential for its prevention and management.
Hypoxia-inducible factor-1 alpha (HIF-1
PX-478, an inhibitor of HIF-1
Pregnant Sprague Dawley (SD) rats, weighing 220
The female and male rats were paired overnight in a 2:1 ratio (1 male and 2 females), and the females with vaginal plugs observed the following morning were designated as being at day 0.5 of pregnancy. Subsequently, pregnant rats were randomly assigned to one of four groups (n = 5 each): the control group (pregnant rats received no specific intervention, and offspring were administered the same volume of physiological saline on the 1st and 3rd weeks post-birth); the PH group (pregnant rats were exposed to hypoxia for 6 h on the 17th day of pregnancy, and offspring received the same volume of physiological saline on the 1st and 3rd weeks post-birth); the PH and PX-478 group (PH+1 w inhibitor; pregnant rats exposed to hypoxia for 6 h on the 17th day of pregnancy, and offspring were orally administered PX-478 (B6004, APExBIO, Houston, TX, USA; 30 mg/kg, continuously for one week) starting at one week post-birth); and the PH and PX-478 administration at 3rd-week groups (PH+3 w; pregnant rats were exposed to hypoxia for 6 h on the 17th day of pregnancy, and offspring were orally administered PX-478 (30 mg/kg, continuously for one week) starting at the 3rd week post-birth).
According to a previous study [11], a hypoxic box containing padding, food, and water was prepared in advance for PH treatment. Then, the 17-day pregnant rats were placed in the hypoxic box, and the nitrogen and oxygen mixture gas (10% O2 and 90% N2) was pumped into the box. Following air removal, an oxygen concentration detector (CY-12C, Jinan Jincheng Security Equipment Co., Ltd, Jinan, Shandong, China) was used to monitor the oxygen concentration in the hypoxic box, which was maintained at 10%. The pregnant rats stayed in a hypoxia box for 6 h.
At the 6th week post-birth, behavioral tests were conducted on offspring from each group. Five offspring were randomly chosen from each group, comprising 2 males and 3 females. Behavioral tests were conducted between 9:00 and 10:00 in the morning. With the exception of the open field test, all other experiments required a one-day adaptation period before commencement. Following the completion of each behavioral experiment, rats were given a 24-hour rest period.
According to previous study [15], a spherical toy was positioned inside the toy box, while untreated offspring of matching age and sex were placed within an unfamiliar mouse box. Subsequently, the offspring were positioned in the middlebox. The durations of playtime for the offspring in both the unfamiliar mouse box and toy box were recorded.
According to previous study [16], a cubic open field box constructed of gray
wood (100 cm in length, 100 cm in width, and 45 cm in height) was utilized for
the test. The bottom of the open field box was partitioned into 25 square grids
delineated by black lines, each measuring 20
According to previous study [17], the target box was positioned at the entrance of the experimental platform and was appropriately marked for subsequent examination. Offspring were gently situated at the center of the platform. If an offspring failed to locate the target box within 1 minute, they were gently guided to the target box and allowed to remain there for 15 seconds. Conversely, if an offspring successfully located the target box within the specified time, they were permitted to remain inside for 15 seconds. Offspring underwent training twice daily, and after four days of training, they were subjected to the assessment. The activity of the offspring within a 4-minute period was recorded without any guidance. Finally, the number of trial and error attempts and the latency to escape of the offspring were tallied.
After eight weeks, euthanasia of the offspring was performed using an overdose of pentobarbital sodium injections (30–50 mg/kg; 230301, New Asia Pharma, Shanghai, China), as described in a prior study [18]. Subsequently, hippocampus and blood samples were collected from all animals. For serum separation, blood samples underwent centrifugation at 2500 rpm for 10 minutes, and the resulting supernatant was collected [19]. The VEGF, alkaline phosphatase (ALP), and aspartate transaminase (AST) levels in the serum were determined using a commercial kit (Nanjing Jiangcheng Bioengineering Institute, Nanjing, China) following the manufacturer’s instructions.
The hippocampi were fixed in 4% paraformaldehyde (Sigma-Aldrich, St. Louis, MO, USA) at room temperature for 48 hours, followed by conventional paraffin embedding and sectioning. Subsequently, the paraffin sections were dewaxed and rehydrated using xylene (Sigma-Aldrich, St. Louis, MO, USA) and ethanol (Sigma-Aldrich, St. Louis, MO, USA). Hematoxylin and eosin (HE; 023J5431, Sigma-Aldrich, St. Louis, MO, USA) staining was conducted, and the sections were sealed with neutral gum. The structure of the hippocampus was observed under an optical microscope (KB0801003, Olympus, Tokyo, Japan) in each group.
The proteins from the hippocampus were extracted using radioimmunoprecipitation
lysis buffer (RIPA; P0013B, Beyotime, Shanghai, China). In brief, hippocampus were
transferred to pre-cooled microtubes containing an appropriate volume of chilled
RIPA buffer. Tissue disruption was achieved using a gentle tissue homogenizer or
motorized pestle under continuous cooling. Then, the homogenate was incubated on
ice for 30 minutes. Gentle agitation during this period facilitated the release
of proteins from subcellular compartments. Post-lysis, samples were centrifuged
at 14,000 rpm for 15 minutes at 4 °C to pellet insoluble debris and
intact organelles. The resultant supernatant, containing the soluble protein
fraction, was carefully aspirated. Following protein concentration determination
with the BCA protein colorimetric assay kit (P0009, Beyotime, Shanghai, China),
the proteins were separated on Sodium dodecyl sulfate (SDS)-polyacrylamide gels
(MP10W12, Sigma-Aldrich, USA) and transferred to polyvinylidene fluoride (PVDF)
membranes (03010040001, Sigma-Aldrich, USA). Subsequently, the membranes were
blocked with 5% bovine serum albumin (BSA) blocking buffer (V900933,
Sigma-Aldrich, USA) and then incubated with primary antibodies (anti-PTEN and
HIF-1
SPSS 25.0 (IBM Corp., Chicago, IL, USA) and GraphPad Prism 8.0 software
(GraphPad Software, Inc., San Diego, CA, USA) were used to analyze and plot the
experimental data. The obtained data were described by mean
The body weights of offspring rats in each group were analyzed initially, and
the results are shown in Fig. 1A,B. The results reveal that offspring in the PH,
PH+1 w, and PH+3 w inhibitor groups exhibited significantly lower body weights
compared to the control group. Furthermore, offspring in the PH+1 w and PH+3 w
inhibitor groups showed a significant decrease in body weight compared to the PH
group. These findings suggest that both PH and the
HIF-1
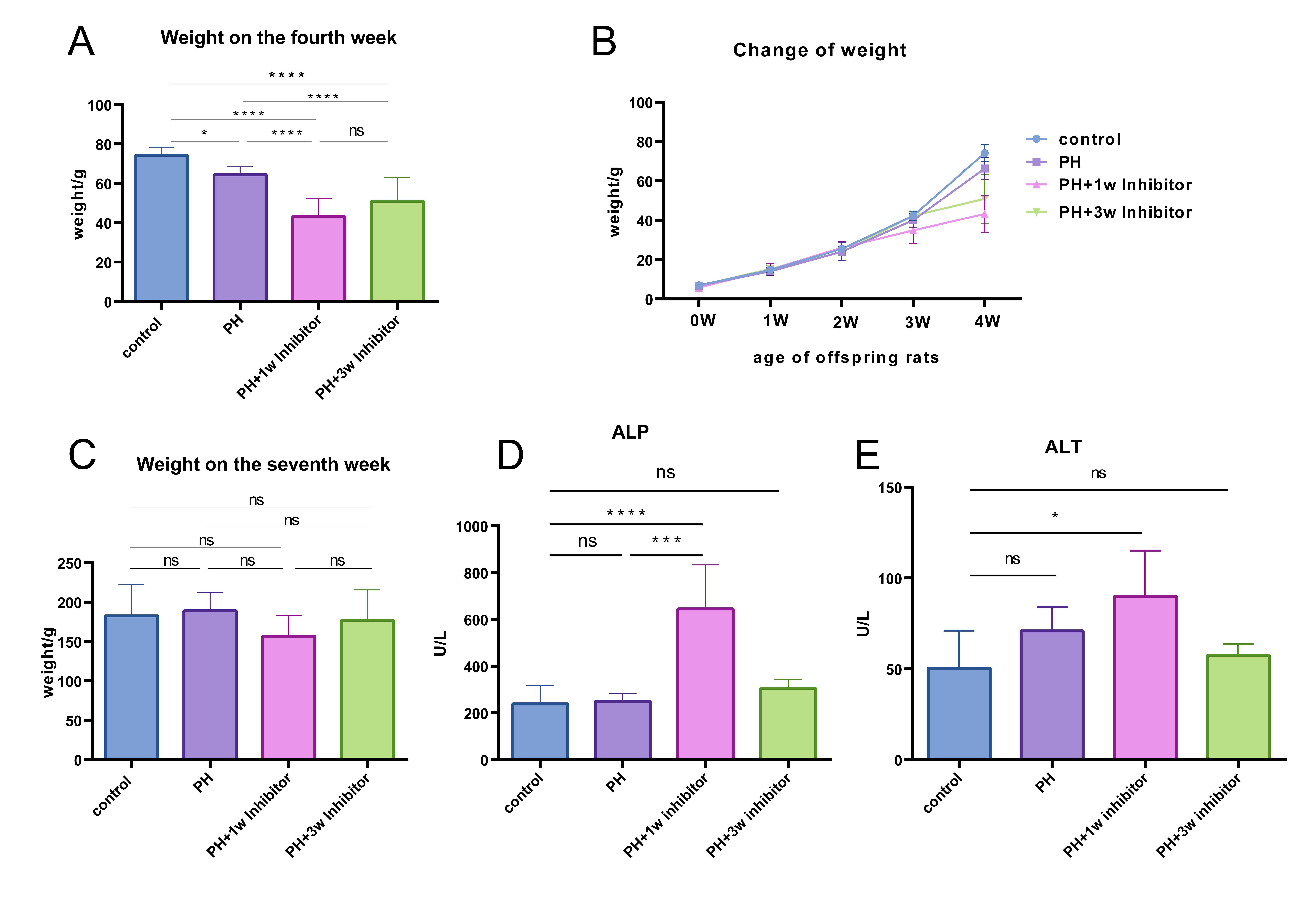 Fig. 1.
Fig. 1.
Effects of PX-478 on the growth of offspring rats. (A) Weight
of offspring rats at four weeks. (B) Trend of changes in body weight of rats over
four weeks. (C) Weight of offspring rats at seven weeks. (D) ALP levels in the
serum of offspring rats. (E) Alanine aminotransferase (ALT) levels in the serum of offspring rats. The
significance of differences between groups was analyzed using the one-way ANOVA
test. ns, p
The HE staining results (Fig. 2) revealed that in the control group, brain tissue cells exhibited orderly and continuous arrangement with intact cell morphology. In contrast, brain tissue cells of mice in the PH group displayed disordered arrangement, nuclear pyknosis and disappearance, blurred nuclear structure, vacuoles, and necrotic cells in the cytoplasm. However, these pathological changes were alleviated after treatment with PX-478, suggesting that PH could impair neural cell development in offspring brain, and early inhibitor administration post-birth may mitigate PH-induced damage to hippocampal neurons in offspring.
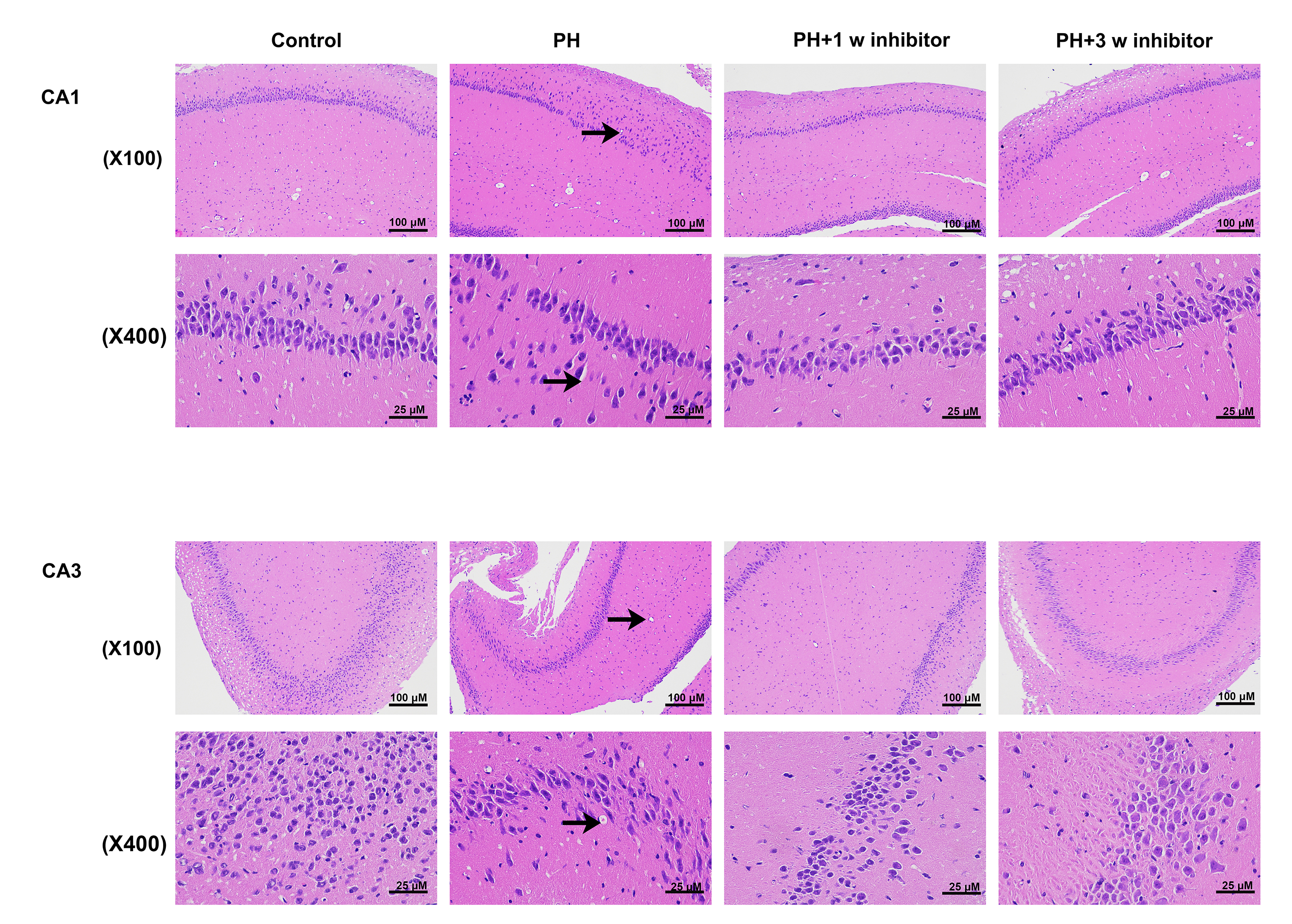 Fig. 2.
Fig. 2.
Effects of PX-478 on brain development of PH-treated offspring
rats. HE staining of the hippocampal CA1 and CA3 neurons of the offspring rats.
The black arrow represented the vacuoles. Magnification 100
Through behavioral tests on offspring, it was observed that PH treatment notably increased self-grooming time (Fig. 3A), toy area stay time (Fig. 3B), and familiar-area stay time (Fig. 3D), while decreasing central area stay time (Fig. 3C). However, in the PX-478 treated group, self-grooming time (Fig. 3A), toy area stay time (Fig. 3B), and familiar area stay time (Fig. 3D) were significantly reduced, while central area stay time (Fig. 3C) was increased in PH-treated offspring. Similarly, results from the Barnes maze test revealed that trial and error times (Fig. 3E,H) and escape latency (Fig. 3F,G) of offspring in the PH group were markedly elevated, whereas PX-478 1 w or 3 w treatment significantly reduced trial and error times (Fig. 3E,H) and escape latency (Fig. 3F,G) of PH-treated offspring. These findings indicate that PH treatment compromises offspring’s spatial memory ability, which can be mitigated by postnatal administration of PX-478 along with improving learning ability. No significant difference was observed between PX-478 1 w and 3 w treatments.
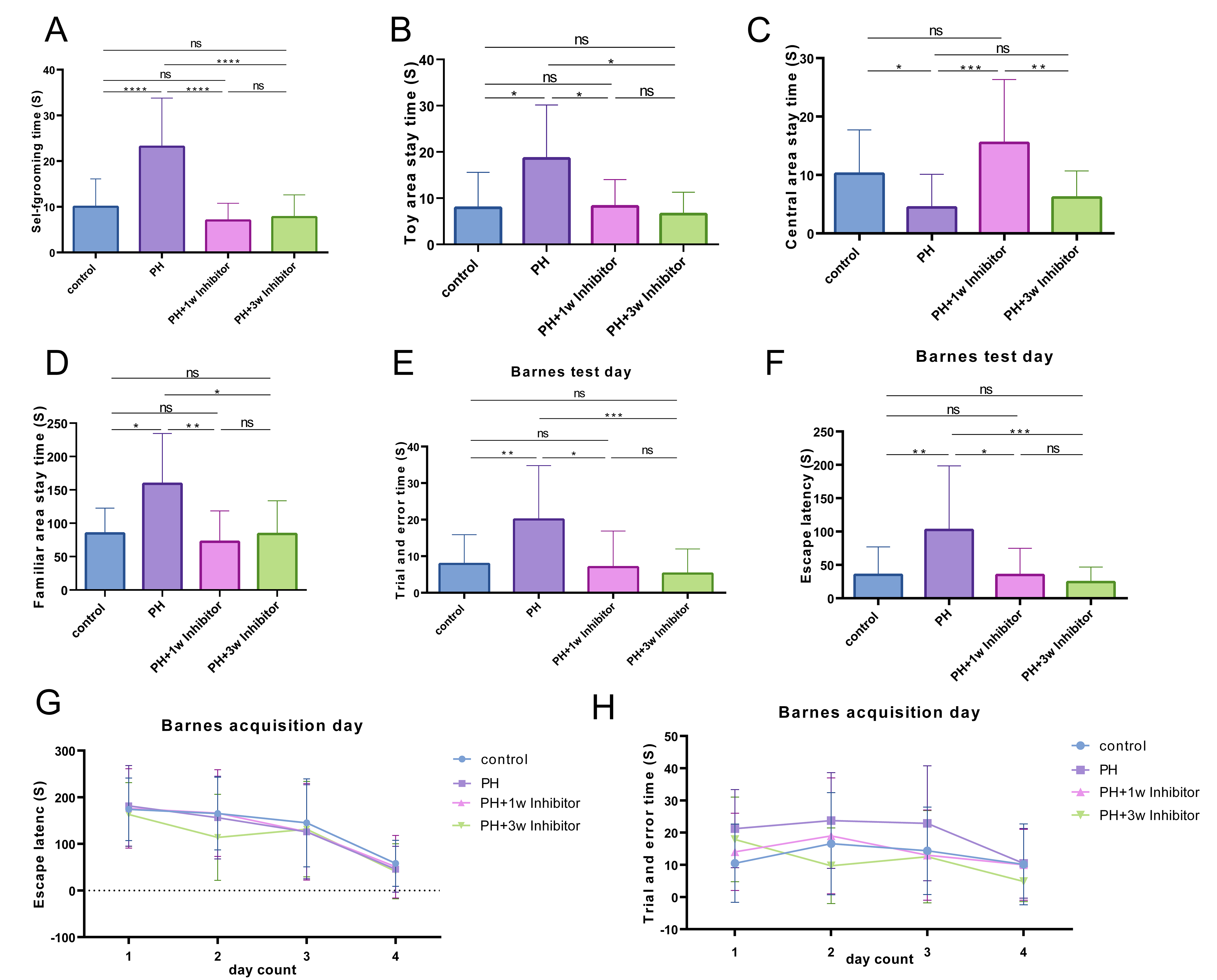 Fig. 3.
Fig. 3.
Effects of PX-478 on the behaviors of PH-treated offspring rats.
The self-grooming time (A) and central area stay time (C) of offspring rats were
measured using the open field test. The toy area stay time (B) and the familiar
area stay time (D) of offspring rats were assessed using the three-chamber social
interaction test. The trial and error times (E,H) and escape latency (F,G) of
offspring rats were evaluated using the Barnes maze test. The one-way ANOVA test
was used to analyze the significance of differences between groups. ns,
p
It was observed that PH treatment markedly elevated HIF-1
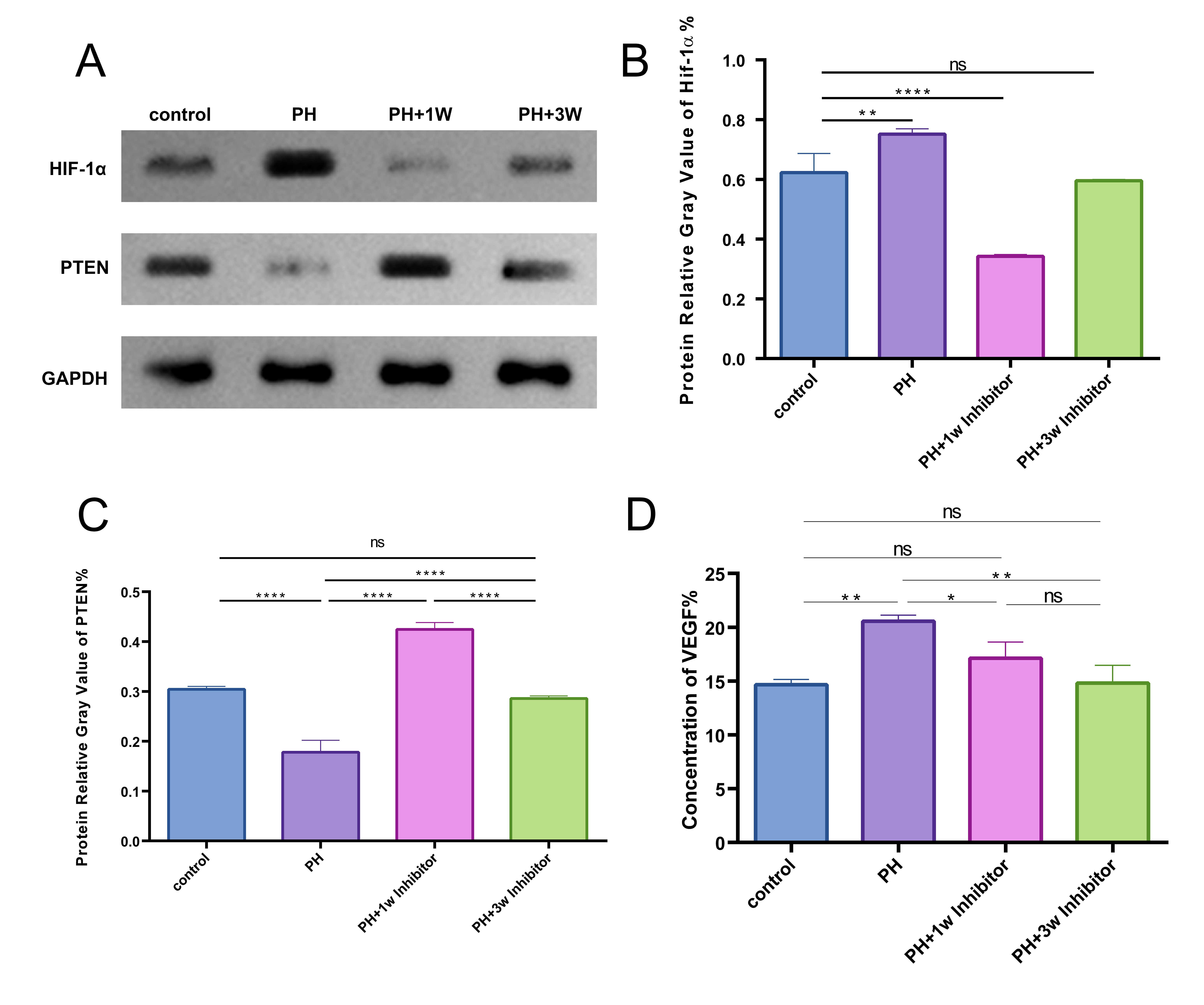 Fig. 4.
Fig. 4.
Effects of PX-478 on the HIF-1
The findings of this study underscore the potential of PX-478 in ameliorating
PH-induced autistic behavior in offspring, notably enhancing their spatial memory
and learning capabilities. However, it’s noteworthy that PX-478 treatment induced
weight loss and impaired liver function in offspring during the early stages,
although these effects attenuated over time. Mechanistically, early post-birth
administration of PX-478 mitigated hippocampal neuron damage induced by PH by
modulating HIF-1
The occurrence of ASD may stem from various factors, among which hypoxia emerges
as a notable environmental contributor [20]. HIF-1
HIF-1
Furthermore, ASD occurrence has been linked to inflammatory factors [34]. A
clinical meta-analysis revealed significantly elevated VEGF concentrations in the
peripheral blood of children with ASD compared to healthy controls. PX-478,
through HIF-1
However, there were still some limitations in this study. PX-478 has certain
side effects that can affect mouse weight and liver function. Although these side
effects may disappear as mice grow, they need to be closely monitored in future
clinical applications. In addition, our study only focused on verifying the
trends of changes between major molecules, the deep molecular mechanisms were not
yet precise. Further analysis is needed to determine the expression of
HIF-1
In conclusion, this study confirmed that the use of PX-478 can reverse the
autism-like behavior of offspring after experience of prenatal hypoxia by
regulating HIF-1
All data in this study were provided in the manuscript.
All authors participated in the design, interpretation of the studies and analysis of the data and review of the manuscript. YD made substantial contributions to the conception and design of the work and revised the article. YY performed the research, built animal models, prepared figures, analyzed the data and wrote the manuscript. JC and TYL supervised the study and were responsible for the interpretation of data for the work. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
This study protocol has been reviewed and approved by the Ethics Committee of Children’s Hospital of Chongqing Medical University (IACUC Issue No: CHCMU-IACUC20221122004).
Not applicable.
This work was supported by the National Natural Science Foundation of China (82272590) and the Natural Science Foundation of Chongqing Science and Technology Bureau (CSTB2022NSCQ-MSX0976).
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/j.jin2309165.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.