




 , Hualin Yu 1,*
, Hualin Yu 1,*1 Second Department of Neurosurgery, First Affiliated Hospital of Kunming Medical University, 650032 Kunming, Yunnan, China
Abstract
Deep brain stimulation (DBS) is a common therapy for managing Parkinson’s disease (PD) in clinical practice. However, a complete understanding of its mode of action is still needed. DBS is believed to work primarily through electrical and neurochemical pathways. Furthermore, DBS has other mechanisms of action. This review explores the fundamental concepts and applications of DBS in treating PD, including its mechanisms, clinical implications, and recent research.
Keywords
- basal ganglia
- beta oscillations
- deep brain stimulation
- neuroprotection
- neurotransmitters
- Parkinson's disease
Parkinson’s disease (PD) is the second most prevalent neurodegenerative
disorder, and its occurrence rises in correlation with advancing age, with males
exhibiting a greater vulnerability to the condition [1]. At present, a conclusive
diagnostic approach for PD is lacking, hence clinical diagnosis remains the
fundamental basis for establishing a diagnosis [2, 3].
Healthcare professionals make a clinical diagnosis based on key symptoms and use
diagnostic criteria to rule out other possible causes [2, 3]. Typical motor
symptoms of PD include resting tremors, bradykinesia, and rigidity [4]. The
annual incidence of PD is estimated to range between 8.7 and 19 per 100,000
people worldwide, depending on research and statistical methods [5]. Currently,
the global population afflicted with PD exceeds 10 million [6]. It is
well-accepted among researchers that PD is a complex condition influenced by
multiple factors. These include genetic factors (autosomal dominant, autosomal
recessive, susceptibility genes), environmental factors (e.g., exposure to
hydrocarbons), constipation, physical activity, smoking (nicotine), and caffeine
intake [7, 8, 9]. The disease is assumed to be caused by initial damage to the
substantia nigra (SN), particularly in its compact area, leading to reduced SN
activity and concomitant loss of inhibition in other regions of the basal ganglia
[including the subthalamic nucleus (STN)], causing overactivity [10]. Typical
pathological changes in PD include the rapid loss of midbrain dopaminergic
(DAergic) neurons and the decades-long accumulation of Lewy bodies formed by
Located in the cerebrum, the basal ganglia (BG) are a group of nuclei that include the SN, the striatum (STR), the globus pallidus (GP), and the subthalamic nucleus (STN), as depicted in Fig. 1 [13]. They primarily integrate and process sensorimotor input from the cerebral cortex to select actions in motor and cognitive functions [14]. Different basal ganglia-thalamocortical loops (BG-TC loops) connect specific areas of the BG, the thalamus, and the cerebral cortex [14]. The classical model separates striatal medium spiny neurons (MSNs) into direct and indirect pathways by different projection targets [14, 15]. According to some researchers, sustaining a balance between direct and indirect paths is advantageous for movement coordination and execution [16]. In a healthy state (Fig. 1, and the left image in Fig. 2 (Ref. [14])), activation of the “direct pathway” (green in Fig. 1, and in the left image in Fig. 2) stimulates D1 [excitatory gamma-aminobutyric acidergic (GABAergic) and cholinergic] dopamine receptors (DRS), which project directly to BG output [15], thereby reducing the activity of the BG, relieving thalamic inhibition, and promoting movement. In contrast, the “indirect pathway” (red in Fig. 1 and in the left image in Fig. 2) activates D2 (inhibitory GABAergic) DRS that projects indirectly to the BG [15] via the external segment of the globus pallidus externus (GPe) and the STN [15]; this activation increases BG output, suppresses thalamic activity, and inhibits movement. The “hyper-direct pathway” (yellow in Fig. 1 and in the left image of Fig. 2) refers to the monosynaptic link between the motor cortex and the STN, which can also receive primary glutamaturgic synaptic inputs from the cortex [17, 18]. This pathway enables cortical inputs to reach the internal globus pallidus internus (GPi) and the substantia nigral pars reticulate (SNr) with relatively short latency, bypassing the STR [15]. The “hyper-direct pathway” has been hypothesized to be essential for inhibiting aberrant movements [18]. DAergic neurons serve a crucial role in the standard BG-TC circuit [15]. When DAergic neurons lose their normal regulatory function, motor and nonmotor symptoms may result. The appearance sequence of these symptoms is related to disease progression, and their combination can significantly impair the patient’s quality of life, as shown in Fig. 3 (Ref. [19, 20]) [9].
 Fig. 1.
Fig. 1.Basalganglionic and thalamocortical loop. The direct pathway (CTX-STR-GPi), the indirect pathway (CTX-STR-GPe-STN-GPi), and the hyper-direct pathway (CTX-STN) pathway. CTX, Cortex; STR, Striatum; GPe, Globus pallidus externus; GPi, Globus pallidus internus; STN, Subthalamic nucleus; SNc, Substantia nigra pars compacta. A simplified schematic diagram is illustrated in Fig. 2. Created with BioRender.com (https://www.biorender.com/).
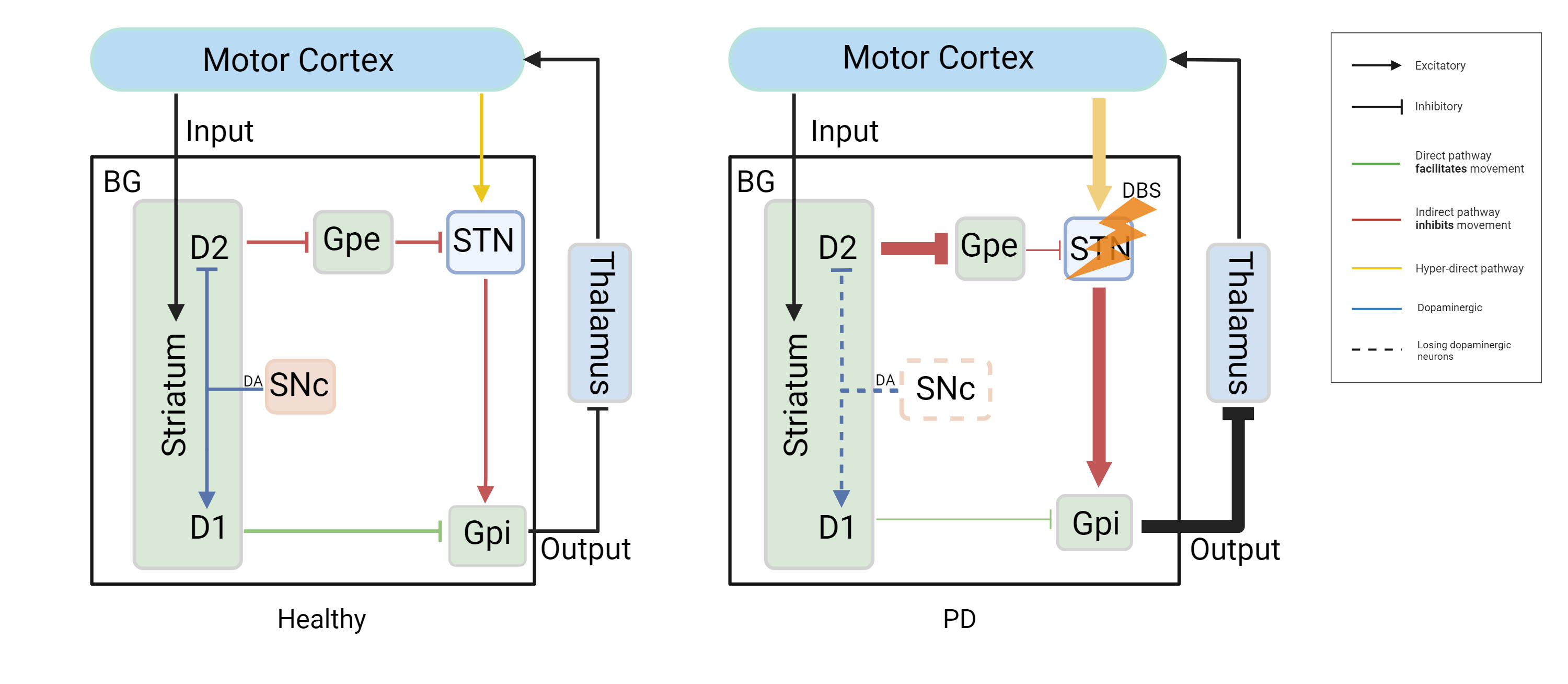 Fig. 2.
Fig. 2.The classical model of the BG. In a healthy state, D1-like DRS projects via the “direct pathway” to the BG output, reducing BG activity and promoting movement. In contrast, D2-like DRS projects via the “indirect pathway” from the Gpe to the STN and then to the BG output, increasing BG activity and suppressing movement. The “hyper-direct pathway” relates to the monosynaptic connection between the cortex and the STN. In PD, decreasing dopamine in the SNc results in decreased activity in the direct path and increased action in the indirect pathway, resulting in exaggerated output from the Gpi. Therefore, there is an excess inhibition of the thalamus and cortex, leading to movement suppression. In addition, the hyper-direct pathway shows excessive activity in PD [14]. DBS in the STN can alleviate motor symptoms. BG, Basal ganglia; PD, Parkinson’s disease; DRS, Dopamine receptors; DBS, Deep brain stimulation; DA, Dopamine. Created with BioRender.com (https://www.biorender.com/). Adapted from McGregor and Nelson [14].
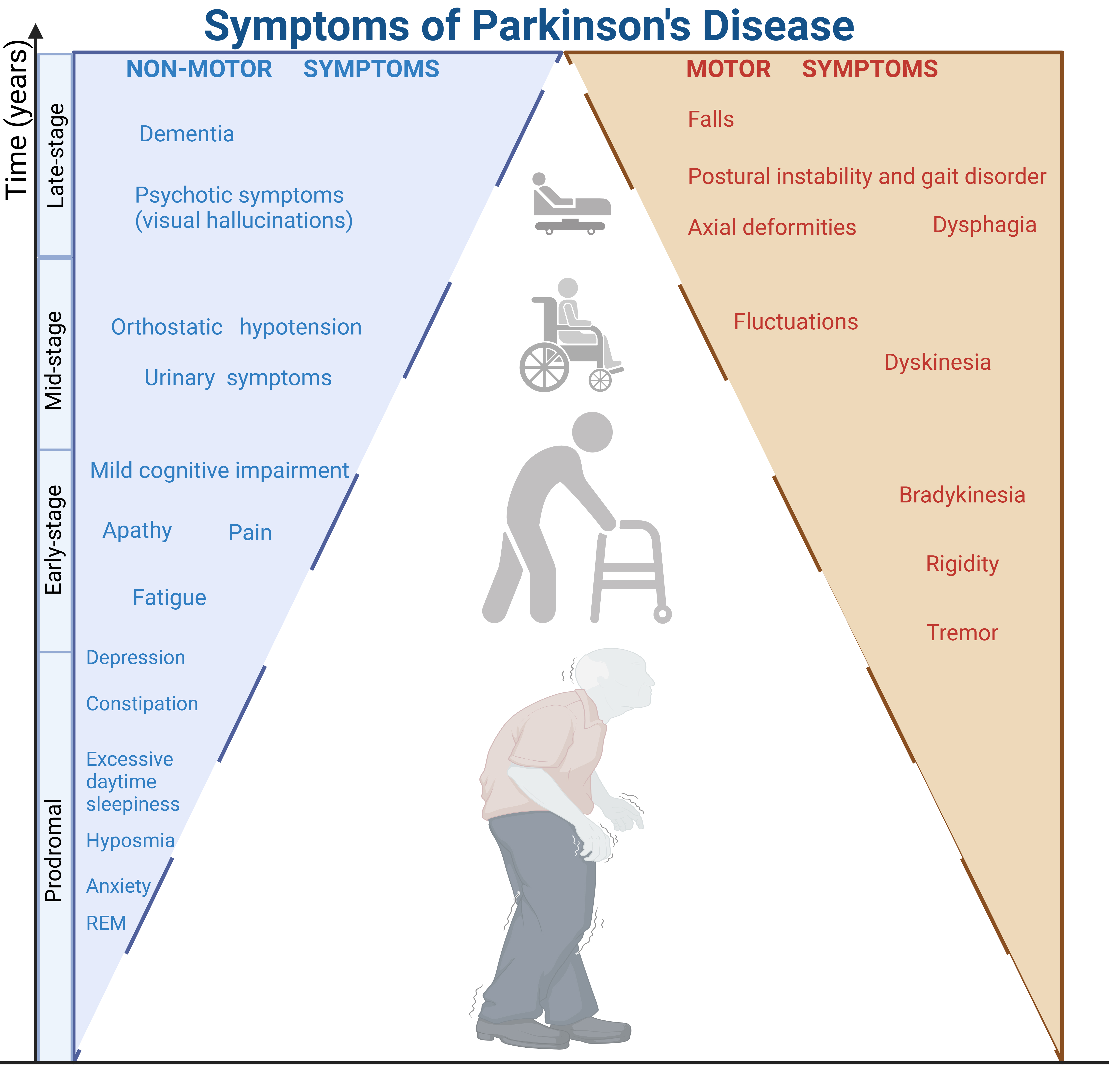 Fig. 3.
Fig. 3.Characterized by specific non-motor symptoms, the prodromal phase of PD can last for years, even decades. Typically, people over 50 develop motor symptoms and receive an early diagnosis of PD. In the mid-stage, patients experience worsening motor symptoms, increased non-motor problems, and motor complications induced by Levodopa (L-DOPA). Late-stage PD presents as a refractory motor impairment to L-DOPA [19, 20]. REM, Rapid eye movement; PD, Parkinson’s disease. Created with BioRender.com (https://www.biorender.com/).
Although PD is currently an incurable, progressive illness [9], active treatment can significantly improve the motor [21] and nonmotor [22] complaints of patients, ultimately, but temporarily, improving their quality of life [9, 23]. For early-stage PD patients, the main symptom treatments involve using levodopa and dopamine agonists to replace the lack of dopamine. This therapy aims to provide a sustained dopamine level. However, after the typical “honeymoon period” of dopamine treatment, patients inevitably experience motor and non-motor complications [7, 24, 25]. Therefore, surgical treatments including ablation techniques (thalamotomy and pallidotomy), and non-destructive deep brain stimulation (DBS) could be used in the later phases of PD; it is worth mentioning that restorative techniques (tissue transplantation and gene therapy) show unlimited opportunities and promise in preclinical studies. DBS is a powerful surgical treatment choice for late-stage PD patients who suffer from motor complications due to long-term drug treatment [13, 23, 25, 26].
For several neurological and psychiatric disorders, DBS is an effective treatment. Medical professionals can widely apply it to multiple conditions, including PD [23, 27], dystonia, severe obsessive-compulsive disorder (OCD), and resistant epilepsy [15, 16]. DBS has been used in over 160,000 patients worldwide to treat a variety of neurological and non-neurological conditions [28]. The number of patients receiving DBS has steadily increased each year. The DBS device comprises electrode(s) connected to an implantable pulse generator (IPG). The basic principle of treatment is to provide continuous or periodic stimulation to a single target (usually bilaterally) (Fig. 4) [13]. To treat PD, electrodes are implanted in the STN or GPi. Different electrical parameters for STN-DBS result in different effects on PD [29]. Standard clinical parameters include tonic continuous application of monophasic cathodic pulses at a frequency of 130 Hz and pulse-width of 60 ms, at an amplitude of 0.8–2.0 mA [30]. Electrode(s) provide continuous high-frequency stimulation for therapeutic effects [31, 32]. The specific procedures of the DBS surgery may vary slightly depending on the multi-disciplinary medical team using it and the type of equipment used. However, the main steps include preoperative evaluation, installation of a stereotactic head frame, imaging localization, skin incision and skull perforation, electrode implantation, intraoperative imaging or microelectrode recording, evaluation of stimulation effect, fixation, implantation of the pulse generator, and postoperative parameter adjustment [23, 33, 34].
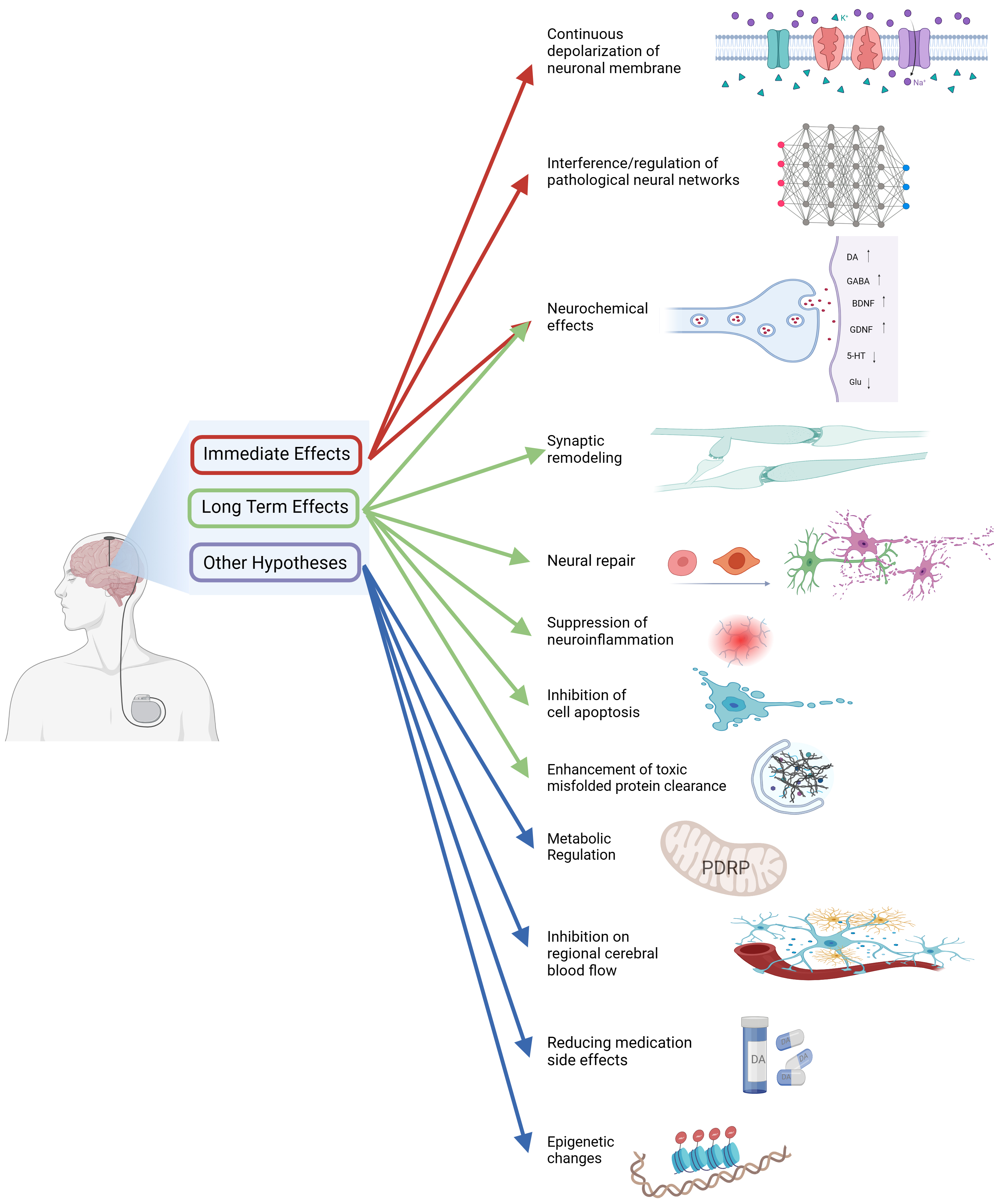 Fig. 4.
Fig. 4.DBS treats PD through multiple mechanisms. DBS, Deep brain stimulation; PDRP, PD-related metabolic pattern. Created with BioRender.com (https://www.biorender.com/).
DBS is a neurosurgical technique introduced in 1987 as a procedure for motor disorder, particularly tremor symptoms in PD patients [31]. In 1997, the U.S. Food and Drug Administration approved DBS to treat essential tremors, and in 2002, it also approved STN-DBS treatment for PD [16]. The three targets for PD-DBS include the STN, GPi, and ventral intermediate nucleus (VIM). However, STN and GPi are the two most popular clinical targets [13]; treatment selection should be guided by the patient’s signs to determine the most appropriate stimulation target [35].
Follow-up studies have shown that DBS can improve the motor performance of PD
patients in the long-term [36, 37], improve their lives [27, 38, 39, 40, 41], and is even
feasible and safe in elderly (
The adverse events most frequently linked to DBS include hemorrhage, infection, skin erosion, and issues associated with the hardware [23, 48, 49]. Furthermore, it is worth noting that there is a possibility of moderate side effects arising as a result of the dispersion of stimulation beyond the intended target region [23]. Nevertheless, in a broader sense, the advantages for individuals with PD after DBS surgery surpass the potential drawbacks [13, 23].
The current understanding of the mechanism of DBS therapy for PD is a subject of debate due to the intricate anatomical connections of the BG and the extensive impact of DBS on various areas within and beyond the motor circuit of the BG [50]. Although a considerable body of research has demonstrated that DBS may operate through the integration of several therapeutic processes, a comprehensive understanding of its precise workings needs to be elucidated. Hence, this article aims to critically examine the existing advancements in the field of PD DBS therapy and explore their potential to serve as a source of inspiration and guidance for the future development of next-generation DBS techniques.
The theoretical foundation of the inhibition-or-cessation theory is the BG-TC loop. Based on the classical model, dopamine signaling exerts a net inhibitory influence on the output of the BG via the GPi, thereby facilitating movement and mitigating symptoms associated with PD. This mechanism enables the execution of precise motor movements [14, 15]. The depletion of dopamine in the SN of individuals with PD leads to an activity imbalance between two pathways (Fig. 2, right image) [14]. In this imbalance, there is an elevation in D2 receptor activity within the indirect pathway, which subsequently suppresses the discharge of the GPe. As a result, there is an increase in the activity of the STN and an accompanying rise in the discharges of the GPi. Ultimately, this cascade of events contributes to the manifestation of motor dysfunction in PD patients [14]. Furthermore, the induction of tremor-like behavior is contingent upon two elements: the augmentation of signal transmission through the hyper-direct channel and the dopamine-dependent hyperpolarization of membrane potential. Both of these components are essential for the observed increase in burst firing inside the STN [17]. Therefore, the motor symptoms associated with PD can potentially be mitigated by the reduction of hyperactivity in the STN.
The inhibition or cessation of neuronal firing rates is substantiated by the use of pallidotomy as a technique for treating PD before the advent of levodopa [51, 52]. In 1994, Siegfried and Lippitz [53] published data supporting the efficacy of bilateral pallidal DBS for PD, showing that DBS may have fewer side effects than pallidotomy and is safe, specific, and reversible in elderly patients. In contrast to thalamotomy, DBS exhibited a reduced incidence of adverse responses (e.g., dysarthria, cognitive decline) and showed more pronounced therapeutic results [54], especially in the context of managing bilateral brain lesions [55]. There is a substantial body of experimental research that supports the notion that the inhibition or cessation of STN/GPi activity and the implementation of DBS effectively mitigate symptoms associated with PD [56, 57, 58, 59].
DBS operates by the inhibition or cessation of neuronal activity, a phenomenon that may be reasonably understood at the level of neuronal membrane potentials. Axons have a higher degree of sensitivity to stimulation than do to cell bodies. Furthermore, high-frequency DBS has been observed to have the effect of inhibiting cells near the implanted electrode while simultaneously stimulating the surrounding fibers [60]. In the GPe, there is a decrease in the firing rate, whereas in the GPi and STN, there is an increase in the firing rate. This leads to sustained depolarization of the neuronal membrane, inactivation of sodium channels [61], and heightened potassium currents. Consequently, there is functional inactivation and damage to STN/GPi neurons [52].
In a broad sense, the impact of DBS closely resembles that of cauterization, as it possesses the ability to inhibit endogenous synaptically mediated electrical activity. Hence, the aforementioned evidence indicates that high-frequency DBS has the capacity to inhibit or halt the firing of STN/GPi neurons. This leads to a complete or partial decoupling between electrical activity and synaptic function, thereby terminating or suppressing the transmission of pathological information among neurons. Consequently, the intended outcome of ameliorating motor symptoms is achieved (Fig. 2, right side).
The regular transmission of neuronal signals within the cerebral cortex is contingent upon the anatomical configuration and functional processes that facilitate the interconnection of individual neurons [62]. DBS can induce not only localized effects through the inhibition of neurons and activation of axons, but also elicit extensive effects on neural networks at a broader level [63]. Thus, the therapeutic mechanism of DBS for PD encompasses neural network dynamics at several levels, surpassing the exclusive influence of individual neurons. In order to comprehensively elucidate the mechanism of DBS, it is imperative to approach the analysis from the standpoint of neural network analysis.
Neural oscillation is a fundamental mode of neural communication [64] and frequently manifests itself as rhythmic fluctuations observed in electroencephalographs (EEGs) and local field potential (LFP) images. The phenomenon of oscillations among clusters of neurons can enhance the process of dynamic communication between neurons situated in distinct locations [65]. Neural oscillations can be categorized into different frequency ranges, including delta (1–4 Hz), theta (4–7 Hz), alpha (7–13 Hz), beta (14–30 Hz), and gamma (30–100 Hz) [66, 67]. Beta oscillations are commonly observed in several brain areas in both humans and other primate species. The areas that exhibit the greatest prominence include the somatosensory and motor cortices, thalamus, BG, and regions associated with the cerebellum [68]. Within the motor network, beta rhythms exhibit their highest intensity during periods of rest or tonic contractions, but decrease in magnitude during exercise and transition towards the occurrence of high-frequency oscillations within a higher frequency spectrum (such as gamma) [69, 70, 71]. Motor disorders can arise from the interference or disruption of sequential information that represents movement commands.
In typical situations, the BG-TC loop exhibits simultaneous processing of motor, cognitive, and affective input. Nevertheless, LFP measurements have shown that individuals with PD have notable alterations in functional connectivity within the BG as a result of a lack of dopamine. This inadequacy hinders the normal functioning of the BG circuit [15, 72]. Specifically, there is an observed elevation in the energy levels of beta rhythms. The decrease in dopamine level has been linked to a significant rise in tremor oscillatory power [69, 71, 73]. Additionally, decreased dopamine is associated with the synchronized firing of neuronal groups [72] and excessive synchronization with network oscillation, which is observed as beta-gamma phase-amplitude coupling (PAC) in surface recordings of the brain. These combined effects contribute to the manifestation of PD symptoms such as tremors, bradykinesia, rigidity, gait, and freezing [74, 75]. A resonance of roughly 20 Hz has been observed in the BG-TC loop of individuals diagnosed with PD [76]. The phenomenon of resonance facilitates the propagation and expansion of spontaneous pathogenic activity, resulting in the emergence of abnormal beta oscillations, such as high-frequency oscillations and gamma oscillations. This has been documented in previous studies [72, 76]. According to computational models, beta oscillations have been suggested to lead to a temporary decrease in the computational flexibility of neural networks. This decrease in flexibility may facilitate the preservation of existing states rather than the emergence of novel activity patterns [77]. Therefore, the foundation for DBS therapy in the management of motor problems in PD involves the disruption of beta oscillations and the mitigation of PAC [75].
According to Oswal et al. [78], DBS has been observed to interfere with and inhibit abnormal oscillations in the beta frequency range, while simultaneously increasing gamma power to alleviate symptoms associated with PD. The application of acute DBS elicits a specific neural-network response in individuals with PD, characterized primarily by a decrease in functional connectivity within the beta frequency band [79]. The hyper-direct channel serves as the primary conduit for delivering high-frequency beta oscillation signals from the cortex to the STN. Its excessive activity in PD patients may result in the production of low-frequency beta signals linked to movement disorders [17]. Thus it has been observed that DBS therapy has the potential to impede the hyper-direct pathway [78]. There is a growing body of research indicating that STN-DBS can be effective in ameliorating bradykinesia symptoms through the reduction of beta oscillations [80, 81, 82]. Additionally, DBS treatment can fine-tune the interaction between gamma oscillations and stimulation frequency [81], This, in turn, can lead to a decrease in cortex-amplitude coupling [83] and a reduction in the extent of phase-locking of beta activity in cortical motor neurons, ultimately enhancing cortical function [74, 84]. DBS has the potential to enhance overall connections, restore network architecture to a state of optimal health, enhance the connectivity between the thalamus and cortex, and diminish the influence of the STR over BG and cerebellar structures [83].
In brief, the application of high-frequency STN-DBS has the potential to modulate aberrant BG output, disrupt pathological synchronization, and inhibit the development and spread of beta oscillations [85]. Consequently, this intervention aims to restore the neural oscillation pattern of patients to a state of normalcy [83, 86, 87, 88]. It is important to acknowledge that DBS does not directly repair the diseased BG system. Rather, it modulates its function, regulates the dynamics of BG output, and modifies its capacity to restore information processing [50, 89, 90]. In autonomic nervous system studies, Herzog et al. [91, 92] demonstrated that STN-DBS effectively integrates the processing of sensory information in the basal ganglia circuitry related to the urinary bladder, leading to the normalization of perception of urinary bladder filling.
DBS can regulate the release of neurotransmitters in areas both near to, and distant from, the site of stimulation, including dopamine (DA), GABA, serotonin, Acetylcholine (Ach), and norepinephrine (NE). The modulation of these neurotransmitters eventually affects the functioning of individual neurons [93]. DBS has a wide range of effects on neurotransmitters, some of which are therapeutic and some of which could be harmful. At present, the field of neurotransmitter detection employs both noninvasive indirect measuring techniques, including positron emission tomography (PET) and magnetic resonance spectroscopy (MRS) scanning [94], and invasive direct measurements, including fast-scan cyclic voltammetry (FSCV) and microdialysis probes [95].
In individuals diagnosed with PD, there is a significant decrease of more than 50% in dopaminergic neurons located in the substantia nigra pars compacta (SNc) [96]. As a result, the administration of dopamine replacement therapy has emerged as the favored approach for treatment. The impact of continuous dopamine stimulation can be simulated by high-frequency stimulation (HFS), which has garnered significant attention due to its influence on dopamine function.
Empirical studies conducted on animals have demonstrated that DBS can enhance the release of dopamine in the SN and STR. According one study, STN-DBS has been observed to substantially elevate the concentration of extracellular dopamine and cause dopamine release in the STR of rats with PD that has been induced by 6-hydroxydopamine (6-OHDA). This conclusion was drawn by the utilization of analytical techniques such as microdialysis and voltammetry [97]. Furthermore, GPi-DBS has been observed to lead to an elevation in dopamine levels in rats [98]. One potential explanation, from rat studies, is that STN-DBS significantly enhances the SNc and facilitates the release of dopamine by directly activating the remaining nigrostriatal neurons [99]. Furthermore, DBS has been observed to elicit the release of dopamine in the STR of primates [100, 101, 102].
Nevertheless, the clinical outcomes of the effect of DBS on the release of dopamine in striatal dopaminergic neurons are inconsistent with the findings of animal studies. The available clinical evidence indicates that effective STN-DBS does not increase striatal dopamine levels [103, 104]. Martinez et al. [105] used microdialysis during the surgical procedure for GPi-DBS in order to assess and evaluate the levels of dopamine in the GPi both before and during the DBS intervention. The study demonstrated that a majority of patients, namely four of five, displayed a notable rise in pallidal dopamine levels after DBS of the globus pallidus internus. Nevertheless, it was shown that there was no significant association between enhanced rigidity and elevated levels of pallidal dopamine, suggesting that additional mechanisms might be implicated in the therapeutic outcomes of GPi-DBS.
Investigations have been conducted in rats with PD (produced by 6-OHDA) to investigate the impact of subthalamic nucleus high-frequency deep brain stimulation (STN-HFS) and Levodopa (L-DOPA). The results indicated that the administration of L-DOPA in an acute setting led to a considerable increase in the duration of increased striatal dopamine levels when combined with STN-HFS. Furthermore, this combination had a synergistic impact with L-DOPA, resulting in an increase and stabilization of extracellular dopamine levels in the STR. The restoration of the self-regulatory function of dopamine release at the presynaptic terminal of the STR may potentially lead to this result. Nevertheless, the efficacy of this feedback mechanism may diminish as the degradation of the nigrostriatal pathway advances [106]. Moreover, animal experimentation has demonstrated that STN-HFS leads to an elevation in dopaminergic receptors expression within the STR of rats [107]. Similarly, STN-DBS has been found to enhance dopamine metabolism in rats with 6-OHDA-induced PD [108].
The available pre-clinical research conducted on animals has indicated that STN-DBS has an impact on dopamine systems, both locally and distantly. However, findings from human clinical microdialysis studies mainly demonstrate local effects [93]. The available information indicates that the administration of levodopa has a comparable therapeutic effect on DBS in reducing beta-band power in the STN and GPi. The observed decrease in symptoms, including motor slowness, rigidity, and tremor suppression, is associated with the use of DBS therapy, as indicated by previous studies [76, 109, 110]. These findings suggest that there is potential efficacy of DBS in facilitating dopamine release.
The proper functioning of the central nervous system (CNS) relies on the equilibrium between excitatory and inhibitory (E/I) neurons at the neurotransmitter level. The disruption of the equilibrium between E/I is the primary factor contributing to the development of numerous CNS disorders. GABA serves as the primary inhibitory neurotransmitter in the CNS. The modulation of glutamatergic (glutergic) and DAergic neurons within the BG-TC loop is crucial in the regulation of motor function [111].
Research conducted on primates has demonstrated that HFS can fully suppress the autonomous electrical activity of neurons in the GPi by activating GABA(A) and GABA(B) receptors [112]. Using magnetic resonance spectroscopy, a clinical investigation identified substantially elevated levels of BG GABA in patients with PD-DBS relative to a control group [113]. Research conducted on patients who had GPi-DBS also validated findings of an augmentation in the release of GABA [114, 115].
The occurrence of degeneration of serotonin cells inside the dorsal raphe nucleus (DRN) is observed in conjunction with PD [116]. The serotonergic system is closely linked to the regulation of emotions and the development of depressive symptoms [117]. Evidence exists that suggests that bilateral STN-DBS has the potential to decrease the firing rate of DRN serotonergic neurons and elicit behaviors resembling depression in rodent models of PD, as well as in control subjects [118]. The application of STN-DBS over an extended time (10 weeks) has been observed to inhibit the activity of serotonergic neurons during calcium transient caliper measurements; the inhibition of the serotonergic cell phenotype may ensue, ultimately culminating in the display of depressive-like behaviors in mice with PD that has been induced by 1-methyl- 4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP) administration [119]. Patients who undergo STN-DBS display behaviors that are associated with the serotonergic and dopaminergic systems, such as depression, suicidal ideation, and impulsive behavior [120].
Although there is no direct connection between the STN and the DRN, research in rats suggests that there may be an indirect mechanism of serotonin regulation using relays in the lateral habenula (LHb) [121]. Further research is necessary to investigate the precise anatomical link between the STN and DRN to identify more accurate targets for DBS and mitigate the occurrence of adverse reactions [93].
ACh functions as a rapid-acting, localized neurotransmitter in the peripheral nervous system, specifically at the neuromuscular junctions and autonomic ganglia [122]. The primary role of central cholinergic neurotransmission is to modulate the excitability of neurons, regulate the release of neurotransmitters presynaptic before synapse, and coordinate neuronal firing, akin to the process of neuromodulation [123, 124]. This system is linked to a range of cognitive functions, such as memory, attention, executive function, and spatial visual function [125]. Cholinergic neurodegeneration is recognized as a pathological hallmark of Alzheimer’s disease [126] and is also associated with the progressive deterioration of cognitive function in PD [127], as well as the development of postural instability and difficulties with gait [128].
The pedunculopontine nucleus (PPN) is located in the upper pons in the dorsolateral part of the ponto-mesencephalic tegmentum and is a neural structure of remarkable diversity, consisting mainly of cholinergic, glutergic, and GABA components [129]. The PPN is linked to gait and balance control [129, 130], and it has been identified as a potential target for deep brain stimulation for treating movement and postural disorders [131]. The application of low-frequency stimulation to the PPN has shown promise in mitigating gait impairments among persons with PD [131]. The impact of STN-DBS on cholinergic neurons in the PPN remains uncertain [130]. However, according to clinical research, DBS targeting the STN has been observed to lead to enhancements in gait speed, distance, postural control, and balance among individuals with PD [132]. These improvements are not directly influenced by the activation of cholinergic neurons in the PPN or local alterations in cholinergic fibers within the STN [130]. Instead, experiments in mice have shown that these effects may be transmitted through other neural pathways [130]. Therefore, additional studies are necessary to clarify the impact of DBS on the cholinergic system in the pedunculopontine nucleus.
Noradrenergic (NE), arising in the neurons of the locus coeruleus (LC), is involved in various sensory, motor, behavioral, and cognitive processes [133]. Both PD and Alzheimer’s disease neuropathology have been found to exhibit a significant reduction in the NE pathway [134]. There is a possibility that the involvement is associated with motor and non-motor symptoms in PD [135, 136].
The interaction between STN-DBS and the monoaminergic (DA, NE) system has been observed, and previous animal investigations have demonstrated that a significant impairment in noradrenergic function in a PD rat model can diminish the therapeutic efficacy of STN-DBS [137]. The clinical data also has indicated that individuals with PD experienced weight gain after undergoing STN-DBS [138]. This weight gain may be linked to the effect of STN-DBS on the NE system in the locus coeruleus [139].
The present level of technology imposes limitations on the research of neurochemical-signal alterations induced by electrical stimulation. The accuracy and reliability of research results may be influenced by the precision, specificity, and validity of procedures used for the detection and analysis of neurotransmitters. Additionally, traditional DBS methods have the drawback of only delivering stimulation on a wide scale and are incapable of differentiating specific cell types or neurotransmitter systems within the area of interest. There are significant changes in the levels of neurotransmitters seen in individuals undergoing DBS for PD. The attainment of therapeutic changes in neurotransmitters is imperative for significantly improving clinical symptoms in patients with PD; for instance, STN-DBS alleviates motor symptoms of hemiparkinsonian monkeys by promoting activity of the dopaminergic system [102]. However, activation of non-target regions, could result in undesirable consequences; for instance, STN-DBS has been linked to mood-related side effects possibly caused by inhibition of DRN serotoninergic systems [118, 119, 120].
Therefore, future studies must take into account the distinctiveness of neurochemical cell types to enhance the precision of identifying the desired site for stimulation. A novel technique known as orientation selective deep brain stimulation (OS-DBS) was recently developed [140]. This technique offers opportunities for refining the ideal functional stimulation.
The key objective in the management of PD is to protect the dopaminergic neurons located in the SN. There is a proposition that suggests that DBS might demonstrate neuroprotective characteristics through the inhibition of DAergic neuron degeneration in the SN [10, 141].
Concerning DAergic neurons, STN-DBS has been proposed as a possible neuroprotective intervention based on rodent models of PD produced by toxin exposure [142, 143]. Also, it has been observed that the persistent application of STN-DBS in the A53T alpha-synuclein PD rat model leads to a decrease in the degeneration of tyrosine hydroxylase-positive (TH+) SN neurons [144]. This effect appears to be influenced by the precise placement of the electrode [142]. Furthermore, studies have shown evidence that STN-DBS has the potential to provide neuroprotection for dopaminergic cells in primates [145].
DBS may exert extensive neuroprotective effects. These consequences consist of the facilitation of neurotrophic-factor release, synaptic stimulation, inhibition of cellular apoptosis, attenuation of neuroinflammation, reduction of glutamate excitotoxicity, and augmentation of the clearance of hazardous misfolded proteins [146].
In PD rats, STN-DBS may have long-term therapeutic effects and neuroprotective effects by inducing increased release of neurotrophic factors [147]. A recent study from our group showed that brain-derived neurotrophic factor (BDNF) has neuroprotective effects on PD mice [148]. The administration of glial-cell line-derived neurotrophic factor (GDNF), by intracerebral infusion targeting the thalamus, has been seen to result in significant therapeutic advantages for patients diagnosed with PD [149].
The use of high-frequency DBS has been seen
to augment the synthesis and secretion of BDNF in glutamate synapses. This effect was observed in
cellular experiments [150]. Furthermore, DBS in rats has been shown to lead to an
increase in BDNF levels in both the nigrostriatal system and the primary motor
cortex [151]. The preformed fibril (PFF) model of rat
alpha-synuclein (
The preclinical research findings suggest that DBS may act as neurotrophic factors to exert neuroprotective effects. However, more clinical evidence is needed to validate this result.
Disturbances in the regulation of neural plasticity are prevalent among patients with PD, resulting in dysfunctions of synaptic transmission and neuronal activity [154]. Clinical research has established that DBS facilitates neuroplasticity in impaired motor circuits [155, 156].
Thirteen PD patients underwent a resting-state functional magnetic resonance imaging (fMRI) analysis to investigate connectivity changes resulting from DBS. The results showed that there was enhanced functional connectivity across premotor areas in response to STN-DBS [157]. Clinical research using fMRI has shown that stimulation reduces the strength of effective subthalamic nucleus afferents and efferents, while strengthening cortico-striatal, thalamo-cortical, and direct pathways [158]. The study recorded whole-cell patch-clamp data from STN neurons in rat-brain slices. The results showed that the application of HFS induced changes in synaptic plasticity in different subpopulations of neurons within the STN. The alterations included short-term potentiation (STP), long-term potentiation (LTP), and long-term depression (LTD) [159]. Also, researchers investigated gene-expression profiles in rat hippocampus using oligonucleotide microarrays and quantitative real-time polymerase chain reaction. They found that HFS can induce neuroplasticity effects at the molecular level by upregulating specific transcription factors [160].
DBS application for improving neuroplasticity will help in the development of new treatments for neurological and psychiatric disorders.
A recent article on stem cell activation suggested that DBS can enhance the neuronal repair process [161]. In 6-OHDA hemiparkinsonian rodents with stable dopaminergic dysfunction, bilateral long-term STN-DBS improved TH+ dopaminergic neurons and their axonal projections in the nigrostriatal and mesolimbic dopaminergic systems [162].
Neural precursor cells (NPCs) exhibit high responsiveness to electric fields, by undergoing proliferation, differentiation, and migration in response to application of electrical stimulation [163]. Iwasa et al. [161] suggested that DBS can stimulate the growth of NPCs [13]; Vedam-Mai et al. [164] found that DBS has the potential to improve symptoms in patients by activating NPCs, then, using rats, found that DBS led to local progenitor cell proliferation and microglial activation [165].
The role of DBS in facilitating NPCs will drive the development of neuro-regenerative medicine. However, due to experimental ethical requirements, there is no clinical evidence yet.
Neuroinflammation is now recognized as an important pathophysiological feature of PD, and microglia activation, astrocyte formation, and lymphocyte infiltration are involved in the inflammatory process and contribute to the neurodegeneration of SNc in PD [7, 166].
Animal studies showed that STN-DBS inhibited the neuroinflammatory process of PD by modulating glial cells (including astrocytes) in the GP, and this result was confirmed in an in vivo rat model and in a cell-culture study [167]. Another experimental study showed that STN-DBS inhibited microglia in the SN of PD rats, and normalized the levels of neuroinflammatory cytokines [168]. Anti-inflammatory cytokines such as Interleukin-13 and Interleukin-4 are upregulated after IN-DCN-DBS (Interposed nucleus-deep cerebellar nuclei-deep brain stimulation) in ataxic mice [169]. After the death of PD patients, postmortem STN samples showed that STN-DBS can lead to a decrease in the inflammatory process [170]. Hepcidin is a mediator of the chronic inflammatory process, a recent clinical study showed that in PD patients treated with DBS, the serum concentration of pro-hepcidin was significantly higher than in the control group and the best medical treatment group [171].
The review by Vedam-Mai et al. [172] suggested that DBS can modulate different subtypes of glial cells, including astrocytes, microglia, and macrophages, thereby altering their phenotype and function to promote therapeutic effects. Therefore, the modulating effect of DBS on neuroinflammation may contribute to delaying or blocking early disease progression in PD.
One hypothesis that has been suggested is that an overabundance of Glu activity in the BG leads to the generation of excitotoxins in SNc neurons, thereby facilitating the progression of PD, and also suggested that blocking this neurotoxicity could be a potential neuroprotective therapy for PD. The assertion is substantiated by electrophysiological and metabolic evidence that indicates that STN-DBS leads to a reduction in the excessive activity of glutamatergic neurons in non-human primates [173]. Furthermore, clinical investigations utilizing magnetic resonance spectroscopy have demonstrated that pontine glutamate levels are lower in a group of individuals with PD who have undergone DBS surgery than in a control group [113]. The effect of DBS on glutamatergic neurons is likely to be a result of stimulation.
In summary, excitatory inputs from the STN may lead to nigrostriatal dopaminergic neuron death, and DBS can block excitatory action of inputs from the STN [174]. Studying the precise mechanism by which DBS blocks the neurotoxic effects of Glu may aid in the development of drugs that target glutamate metabolism, both presynaptically and postsynaptically.
Apoptotic and autophagic degeneration was found in the substantia nigra of patients with PD [175]. STN-DBS induces autophagy and inhibits apoptosis, providing a molecular basis for its neuroprotective effects. This prevents phosphorylation of B-cell lymphoma (Bcl)-2, which increases its activity and dissociates it from Beclin1, a positive regulator of autophagy. This has been shown to occur when protein phosphatase 2A activation is inhibited in 6-OHDA-induced PD rats [176]. However, there are no clinical studies to show how DBS affects apoptosis in PD.
The formation of Lewy bodies in neurons is one of the characteristic
pathological manifestations of PD [177]. Lewy bodies are mainly composed of
In summary, the primary body of data substantiating the neuroprotective benefits of DBS is generated from research done in animal models. Postmortem studies [178] and clinical evidence [182] suggest that the use of DBS as a treatment intervention does not exhibit any neuroprotective effects in individuals with PD. This discovery highlights a potential constraint in evaluating the neuroprotective effects in clinical settings, since DBS is often used at the advanced stages of the illness, potentially missing the ideal therapy window. A recent clinical follow-up study from our group showed that the results of DBS in patients with slow and moderate progression were better than those with fast progression [183].
There may be two main reasons why neuroprotection has not yet been demonstrated in the preclinical and clinical stages: (1) animal models have inherent limitations that hinder their ability to accurately replicate the physiological and pathological processes of human diseases [184, 185]; (2) although the neuroprotective effect of DBS slows the rate of dopamine neuron loss after surgery, all nigrostriatal dopamine neurons were lost before the patient died, so there was no significant difference at the time of death [186].
If DBS is indeed neuroprotective, and whether the use of high-frequency stimulation (e.g., STN-HFS) in the early stages of PD can provide additional neuroprotection to delay or even arrest the progression of PD, needs to be further explored in future experimental studies.
Patients with PD exhibit a specific spatial covariance pattern of glucose metabolism in 18F-fluorodeoxyglucose and positron emission tomography imaging, termed PD-related metabolic pattern (PDRP). This is characterized by a relative increases in pallidum, thalamus, pons, and cerebellar metabolism and concurrent metabolic reductions in pre-motor and posterior parietal areas [187]. The quantification of PDRP expression has been shown to have a link to the severity of the clinical illness [187, 188]. STN and GPi-DBS have been shown to reduce abnormal brain network activity in PD by decreasing the expression of the PDRP [189, 190].
Astrocytes play a role in the modulation of neuronal firing and exert a direct influence on cerebral blood flow. This influence may result in either an augmentation or reduction of neuronal activity by mediating neurovascular coupling, as shown by PET studies [63]. A study combining high-resolution functional near-infrared spectroscopy and low-resolution EEG found that STN-DBS was associated with reduced blood volume in bilateral sensorimotor (SM), premotor (PM), and dorsolateral prefrontal (DLPF) cortex, whether the patient was performing a task or not [191]. Previous research using PET scans has established a correlation between decreased cerebellar blood flow and thalamic stimulation-induced suppression of unilateral symptoms [192]. DBS may be affecting metabolism by affecting (e.g., inhibiting) blood flow to brain regions, which in turn affects neuronal firing patterns and neural signaling, thereby alleviating the symptoms of PD.
The motor and neuropsychiatric challenges linked to the innervation of the striatal dopamine system are exacerbated and reinforced with prolonged dopamine agonist (DAergic) therapy in PD [25]. Furthermore, prolonged dopamine treatment may increase DRS tolerance, potentially reducing its therapeutic efficacy. DBS technology can regulate neuronal activity, decrease tolerance to dopamine replacement therapy, improve the effectiveness of dopamine agonist medications, and minimize adverse effects [25]. STN-DBS treatment for patients can reduce the levodopa-equivalent daily dose by approximately 39%, potentially decreasing gastrointestinal side effects [41].
By controlling gene expression in a reversible and heritable fashion, cellular epigenetic mechanisms operate independently of DNA coding sequences [193]. While modulating responses to a vast array of external stimuli, epigenetics regulates neuronal development and functions as an interface between the environment and the genome [194]. Among the regulatory mechanisms in epigenetics encompass DNA methylation patterns, modifications of histones, and expression of non-coding RNA [195]. Notably, the synuclein gene is especially susceptible to epigenetic modification [196]. DBS therapy has been shown to modulate neuronal activity by engaging certain epigenetic mechanisms, thereby leading to an amelioration of clinical symptoms [195]. The DNA methylation-states observed in the brains and blood samples of individuals with PD demonstrate a remarkable degree of consistency [195]. DBS-induced genes exhibit distinct DNA methylation patterns [197]. Furthermore, researchers have compared the expression of microRNA (miRNA) and splicing in the blood leukocytes of PD patients before and after DBS treatment [198]. They identified 11 miRs that were differentially expressed in DBS-treated PD patients and untreated PD patients, indicating a possible mechanism for the relevance of DBS in alleviating PD pathologic signs [198].
DBS has a multitude of effects on diverse neural elements in and around the anatomical nucleus, culminating in clinical improvement, according to the available evidence [199] (Fig. 4). STN-DBS for PD can relieve tremors within seconds, providing evidence to support the hypothesis that STN-DBS operates by directly interfering with neural transmission within the BG network involved in tremors [42, 200]. The manifestation of slower-responding symptoms is largely influenced by long-term mechanisms, such as synaptic plasticity and eventual anatomical remodeling [201].
Current research suggests that the mechanism of DBS treatment for PD is primarily influenced by its effect on brain neural-circuit structure and function. Given that “structure dictates function”, investigation into neuronal protection mechanisms holds great potential for integrating disease prevention, treatment, and recovery.
The investigation into the mechanism of DBS in the treatment of PD has been further advanced by technological advancements in computer science, medical imaging, research methods, and data collection that will aid in the precise localization of the lesion site and the identification of novel therapeutic targets. These developments will minimize the adverse effects of misplaced stimulation of non-target regions, and will maximize therapeutic stimulation. We foresee further expansion in the clinical implementation of DBS.
One promising field of investigation is the development of adaptive DBS algorithms. The algorithms can adjust stimulation settings in real-time by analyzing local field potentials, thereby enhancing the therapeutic effect while also minimizing potential negative reactions [202].
DBS is considered a less invasive alternative to ablative surgery. However, it still involves intrusion. Moving forward, the integration of emerging technologies, such as transcranial direct-current stimulation, repetitive transcranial magnetic stimulation, optogenetics stimulation [203], chemogenetic stimulation, and focused ultrasound technology [204, 205], may cause those practicing traditional DBS to reconsider their stance.
HP and RQ participated in design, and manuscript drafting. ZW, LM and JW participated in literature selection, JL and XG participated in mapping, YZ and MZ participated in figure design and literature review. HY participated in conception and design. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This research was funded by the Health Commission of Yunnan Province, Renowned Doctors Special Program, Yunnan High-level Talents Training Support Program, grant number KYRLMY20200026; Yunnan Applied Basic Research Project of Diagnosis and Treatment Center for Nervous System Diseases, grant number ZX2019-03-05; Health Commission of Yunnan Province, Science and Technology Talents and Platform Program, grant number 202105AC160029; Geng Xin, a Project for the Training of Technological Innovation Talents in Yunnan, grant number 202205AD160006; Reserve Program for Young and Middle-aged Academic and Technical Leaders (Project No. 202205AC160013).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.