

1 Department of Rehabilitation Medicine, The Second Affiliated Hospital of Nanchang University, 330006 Nanchang, Jiangxi, China
2 School of Clinical Medicine, Nanchang Medical College, 330004 Nanchang, Jiangxi, China
Abstract
Evidence suggests that aging contributes to decreased cerebral blood flow and brain oxyhemoglobin (HbO2) in the association cortices during rest. However, the influence of aging on functional brain activation is still controversial. The objective of this study was to investigate the age-related dependence of HbO2 across distinct motor control conditions in both primary and association cortices.
Using functional near-infrared spectroscopy (fNIRS), this study assessed HbO2 level changes within the primary somatosensory cortex (PSC), primary motor cortex (PMC), supplementary motor cortex (SMC), prefrontal cortex (PFC) and dorsolateral prefrontal cortex (DLPFC) under various motor control conditions. Analysis examined changes in the concentration of HbO2 measured by fNIRS during rest, motor execution (ME), motor passivity (MP) and motor imagery (MI) with elbow flexion in 30 younger (21.5 ± 1.17 years old) and 30 older (60.9 ± 0.79 years old) adults.
During motor execution HbO2 was higher in younger adults than older adults in bilateral PMC, bilateral PFC, left PSC, left SMC and left DLPFC (p < 0.05). During motor passivity, HbO2 was higher in younger adults than older adults in bilateral PMC, left PSC and left SMC (p < 0.05). During motor imagery, HbO2 was higher in younger adults than older adults in bilateral PFC and bilateral DLPFC (p < 0.05).
This study provided evidence that HbO2 levels are different in the primary and association cortices during different motor control conditions in young and old adults and that HbO2 levels in different brain regions under different motor control conditions can be influenced by age.
Keywords
- functional near-infrared spectroscopy
- motor execution
- motor passivity
- motor imagery
The human brain comprises approximately 2% of the total body weight, yet it consumes 20% of the body’s oxygen [1, 2]. The basis of life phenomenon is the energy material ATP generated in a cell, while oxygen (O2) is essential for the continued production of ATP. A few minutes of ischemia or hypoxia can lead to irreversible damage or even death of brain tissue [3]. The supply of O2 to cells is through breathing, which transports inhaled air containing O2 into the alveoli within the lungs, where O2 enters the blood through gas exchange and is transported throughout the body by the arterial blood. O2 in the capillaries is exchanged with the tissues again, to then be taken up by the cells. The majority of O2 molecules in the blood are reversibly bound to oxyhemoglobin (HbO2) in red blood cells, with a fraction dissolved in the plasma. When the brain is thinking, a certain area of the cerebral cortex is activated to complete the corresponding thoughts. At this time, the HbO2 in the cortex of this area will change, with the degree of change reflecting the intensity of regional activation [4, 5]. Therefore, HbO2 is a critical indicator of brain tissue activity and function and its regulation is essential for maintenance of normal wakefulness and cognitive function [6, 7]. Investigating changes in HbO2 metabolism in healthy individuals can contribute to the understanding of spontaneous alterations in neural activity and responses to stimuli during brain development, maturation and aging processes [8, 9]. Moreover, HbO2 holds significant implications for the mechanisms underlying neurological disorders such as cerebrovascular disease, mental illness and neurodegenerative disease.
Different studies have presented varying perspectives on HbO2 metabolism during the aging process. Some investigators argue that HbO2 metabolism decreases with age, possibly due to continuous low perfusion and oxygen supply resulting from atherosclerosis or age-related cell loss (manifested as reduced gray matter volume) or changes in cellular function (manifested as decreased oxygen utilization) [10, 11]. Alternatively, it has been suggested that despite progressive atrophy of brain structure with age, brain function remains relatively stable for an extended period [12, 13]. This implies a complex relationship between aging and HbO2 metabolism [14]. Compared to younger adults, brain activation levels in certain regions of older adult brain tend to decrease, often accompanied by increased activation in other areas. Here, the hypothesis is that older individuals compensate for age-related neural changes by recruiting additional neural circuits or utilizing alternative pathways [15]. Since motor and cognitive tasks require the delivery of adequate oxygen and glucose to specific areas of the brain, it is thought that blood flow to specific areas should be related to enhanced metabolic activity in those areas performing a task [16]. However, this process depends on complex and poorly understood brain vascular regulation mechanisms which may change with aging and disease. A common explanation for the mismatch between cerebral blood flow and oxygen consumption during neuronal excitation is that blood flow increases beyond oxygen consumption to compensate for the depletion of oxygen reserves in brain mitochondria [17].
Aging is associated with structural and functional alterations in the brain. Consequently, it is important to explore strategies that harness the brain’s inherent plasticity and adaptability and employ continuous training methods to reshape brain function [18, 19]. Thus, individuals with impaired brain function can improve and reshape that function through continuous motor training. Currently, commonly used clinical motor controls include active, passive and imagined movements [20]. Active movements require individuals to have some degree of voluntary motor ability and are suited to late-stage recovery. In passive movement, individuals do not need to actively participate, as the entire process is carried out by therapists or therapeutic equipment, making it ideal for early-stage brain injury patients with limited motor function. Imagined movement involves patients mentally simulating actions without producing actual limb movements, making it suitable for brain injury patients with impaired motor function [21, 22]. Combined with the previous research results on the brain activation characteristics of these three motor tasks, the regions of interest (ROIs) in this study are primary somatosensory cortex (PSC), primary motor cortex (PMC), supplementary motor cortex (SMC), prefrontal cortex (PFC) and dorsolateral prefrontal cortex (DLPFC) [23, 24].
Functional near-infrared spectroscopy (fNIRS) is based on a neurovascular coupling mechanism, where local changes in brain functional activity lead to corresponding alterations in local cerebral blood flow [25, 26]. Oxygenated and deoxygenated hemoglobin in the cerebral cortex blood vessels absorb near-infrared light to varying degrees, causing near-infrared light attenuation. By analyzing the correlation between light attenuation and changes in chromophore concentration in the tissue, fNIRS can quantitatively measure changes in HbO2 concentration in brain tissue, thus reflecting the level of brain activity [27, 28]. fNIRS provides real-time monitoring of HbO2 metabolic levels. It is portable, easy to operate and meets the requirements of spatiotemporal resolution of brain function research, making it highly valuable and widely applicable in this field of research [29].
Batula et al. [30] used fNIRS to compare the brain activation levels of motor execution and motor imagery in the upper and lower limbs and found that motor execution activated motor areas to a greater extent than motor imagery and that the differences in the upper limbs were more obvious than those of the lower limbs. Holtzer et al. [31] used fNIRS to compare changes in cerebral oxygenation of the PFC during walking and walking while talking (dual-task) in older and younger adults and found that oxygenation levels were increased in the PFC during walking while talking when compared to only walking. Additionally, the authors found that this increase was greater in younger compared to older adults [31]. Zhang et al. [32] used fNIRS to explore age-related changes in cerebral cortex activation during finger-to-thumb opposition movement and found that the average beta values of the left and right PFC in an older group were significantly higher than for those in a younger group. Similarly to Holtzer et al. [31], some have found that older individuals demonstrate smaller changes in cerebral oxygenation during finger tapping when compared to younger individuals, which is consistent with the lower voxel activation observed in functional magnetic resonance imaging (fMRI) [33, 34]. Taken together, existing evidence on age-related differences in cerebral oxygenation suggest that younger adults demonstrate greater cerebral oxygenation during motor tasks and findings seem to vary based on the type of motor condition studied.
Here, the aim was to evaluate HbO2 changes in five areas of the cerebral cortex (PSC, PMC, SMC, PFC and DLPFC) in younger and older adults that completed three different motor conditions (motor imagery, motor execution and motor passivity).
As mentioned above, the use of fNIRS to examine age-related differences in cerebral oxygenation in different motor control conditions is not new. A unique aspect of this study was the evaluation of multiple motor control conditions (motor execution, motor passivity and motor imagery) and targeting the following ROIs: PSC, PMC, SMC, PFC and DLPFC. Also, while there is a large literature on walking and age-differences in fNIRS [35, 36], less is known concerning elbow flexion and multiple motor conditions. However, upper limbs account for 60% of the body’s total body functions and have a direct impact on the quality of life. The elbow flexion–extension movement is intrinsic to many daily activities such as tooth brushing, face washing and eating. No previous study has reported age-related change in elbow flexion and multiple motor modes, although this activity is common in daily life. Therefore, this study used an elbow flexion task to gain a deeper understanding of the role and significance of HbO2 metabolism and its association with the aging process. Specifically, the following hypothesis was evaluated: HbO2 levels in different brain regions under different motor control conditions is influenced by age.
Based on Causse et al.’s [37] description of age in fNIRS research, 30
younger (mean age 21.5
Prior to the study, investigators provided each subject with a detailed explanation of the specific requirements and instructions for the motor tasks. Subjects underwent repeated motor control training to ensure that they could each accurately perform the three different motor control tasks. During the study, investigators were responsible for each subject completing the motor control task at the same speed, frequency and intensity to ensure data reliability. Additionally, subjects were required to rest quietly for at least five minutes prior to performing any motor task. During the execution of the active, passive and imagined elbow flexion tasks and rest period, changes in brain HbO2 were monitored.
In the active elbow flexion task, subjects performed repetitive elbow flexion-extension movements as prompted by computerized voice instructions. Specifically, when the computer voice instructed “flex” participants repeated quick and vigorous elbow joint movements from the starting position of 0° extension to 90° flexion and back for 20 s. When the computer voice instructed “rest” participants kept their elbow joint in a fully extended position and relaxed for 20 s. The passive elbow flexion task involved an examiner who assisted subjects in moving their elbow joint to achieve the same angles and speed as in the active elbow flexion task based on computerized voice prompts. In the imagined elbow flexion task, subjects were instructed to imagine flexing their right elbow for 20 s without activating the elbow flexor muscles. Each participant repeated the “flex-rest” sequence three times, with a 20 s rest period between any two motor tasks to allow HbO2 to return to a baseline level. The resting task involved no execution of any physical or mental tasks, essentially representing a state of rest. For the sake of comparison, the procedures for the three tasks were identical, the only difference being whether they involved mental or physical activity related to elbow flexion.
A NirSmart-6000A device (Danyang Huichuang Medical Equipment Co., Ltd., Danyang, Jiangsu, China) was used to continuously measure and record the HbO2 level in subjects while they performed the various tasks [39]. The system comprises near-infrared light sources and avalanche photodiodes as detectors. The light source probes emitting light at wavelengths of 730 nm, 805 nm and 850 nm, with a sampling rate of 11 Hz. The experiment utilized 18 light source probes and 12 detection probes, forming 31 effective channels, with an average distance of three centimeters between the emitters and detectors (ranging from 2.7 to 3.3 cm). The placement of the optodes on the head followed the international 10–20 system for electrode placement [40].
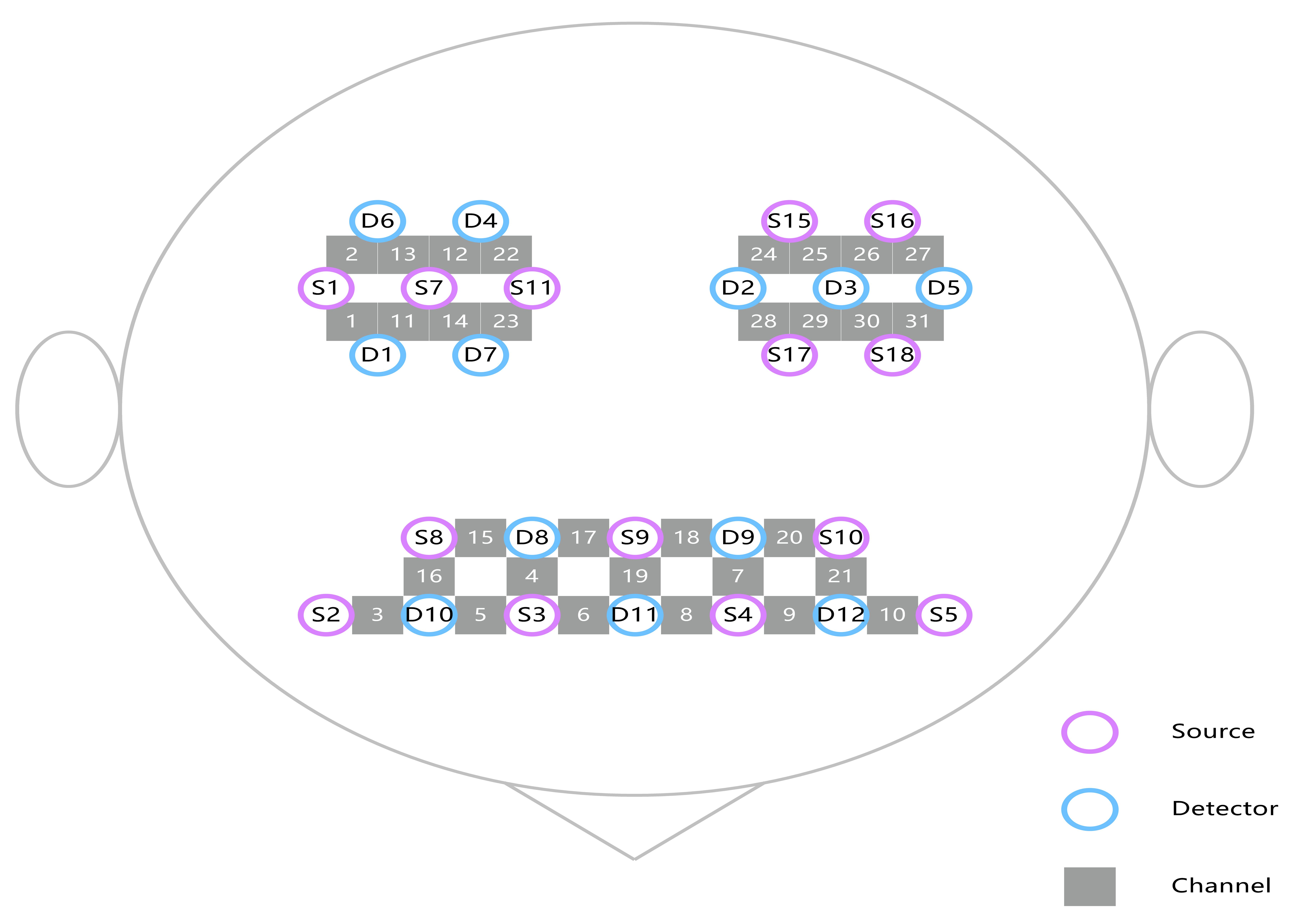
Based on previous researches [23, 24], the study primarily focused on specific ROIs within the brain. These ROIs included the PSC (Brodmann area: 1.2.3), PMC (Brodmann area: 4), SMC (Brodmann area: 6 area), PFC (Brodmann area: 10) and DLPFC (Brodmann area: 46). Each subject had fifteen channels in the frontal lobe and eight channels each in the left and right temporoparietal lobes. The arrangement of the channels is given in Fig. 1 and the channels corresponding to the ROIs are listed in Table 1.
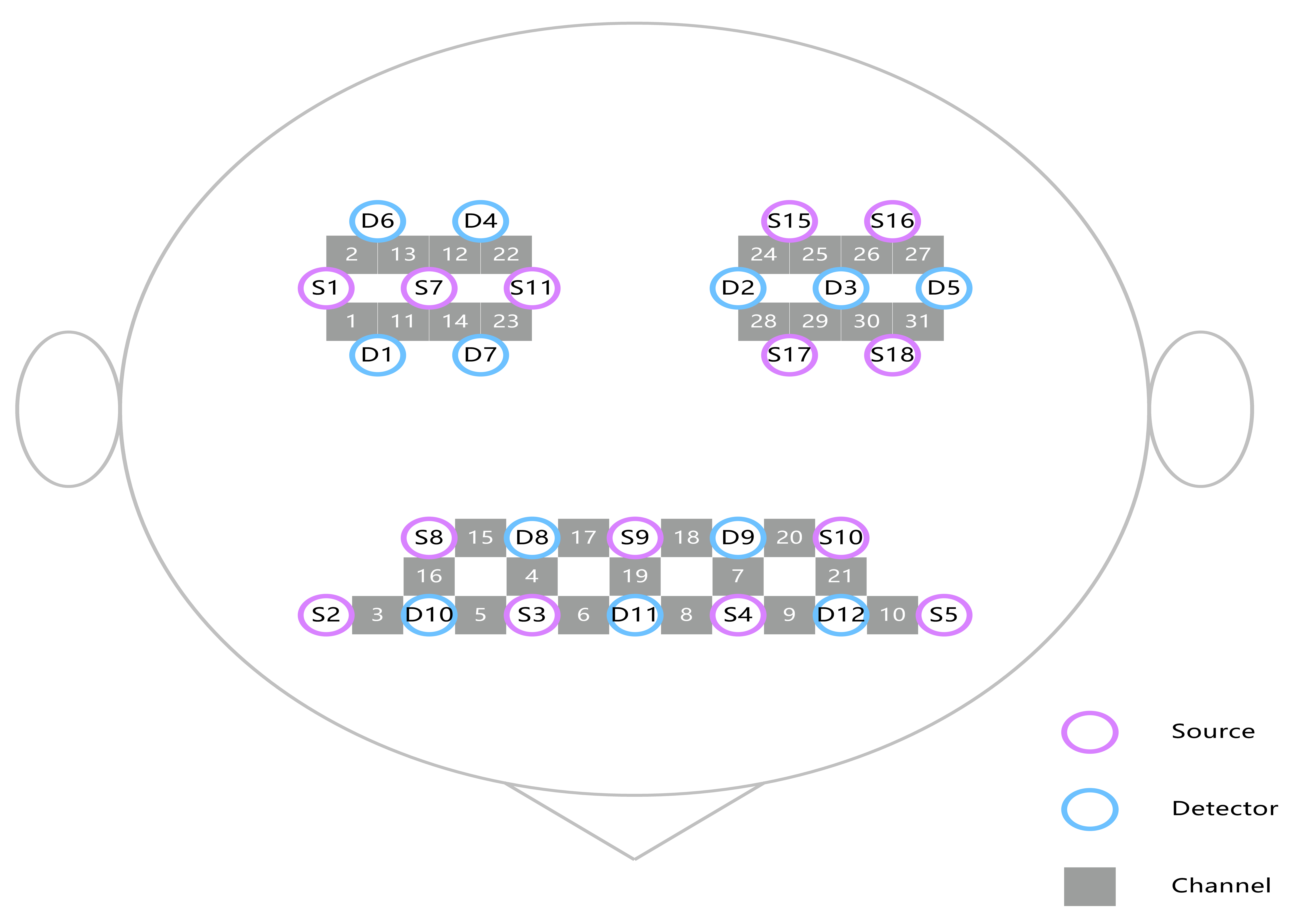 Fig. 1.
Fig. 1.
Location of the 31 channels according to the international 10–20 standard system.
| Brodmann area | Cortex name | Right hemisphere channels | Left hemisphere channels |
| 1, 2, 3 | Primary somatosensory cortex | 21, 2, 13 | 25, 26, 27 |
| 4 | Primary motor cortex | 22 | 24 |
| 6 | Supplementary motor cortex | 1, 11, 14, 23 | 28, 29, 30 |
| 10 | Prefrontal cortex | 4, 5, 6, 17 | 7, 8, 9, 18 |
| 46 | Dorsolateral prefrontal cortex | 3, 15, 16 | 10, 20, 21 |
Raw intensity measurements at 730, 805 and 850 nm were first low-pass filtered with a finite impulse response filter. A cutoff frequency at 0.14 Hz eliminated possible respiration and heart rate signals and unwanted high-frequency noise [41]. Using a combined independent component analysis/principal component analysis, environmental and equipment noise and signal drifts were then removed from the raw intensity measurements.
The modified Beer-Lambert law was then used to calculate the relative hemoglobin concentration changes in HbO2 [42]. The hemodynamic response function initial time was set to zero seconds and the end time was set at 20 s (with “–2–0 s” reserved as the baseline state and 0–20 s as the time for a single block paradigm). HbO2 concentrations for each block paradigm were superimposed and averaged to generate a block average result.
Statistical analysis proceeded by calculating the block average of HbO2
during the active, passive, imagined elbow flexion tasks and rest period. The
block average of the rest period gave the baseline value and the three task block
averages were then each used as the activation value. The HbO2 concentration
for each block paradigm was superimposed and averaged to generate a block average
result. The subtraction of the mean baseline value for HbO2 from the mean
activation value produced the change in HbO2 during the activation period.
These data were compared between younger and older groups by independent
t-test. All data are expressed as mean
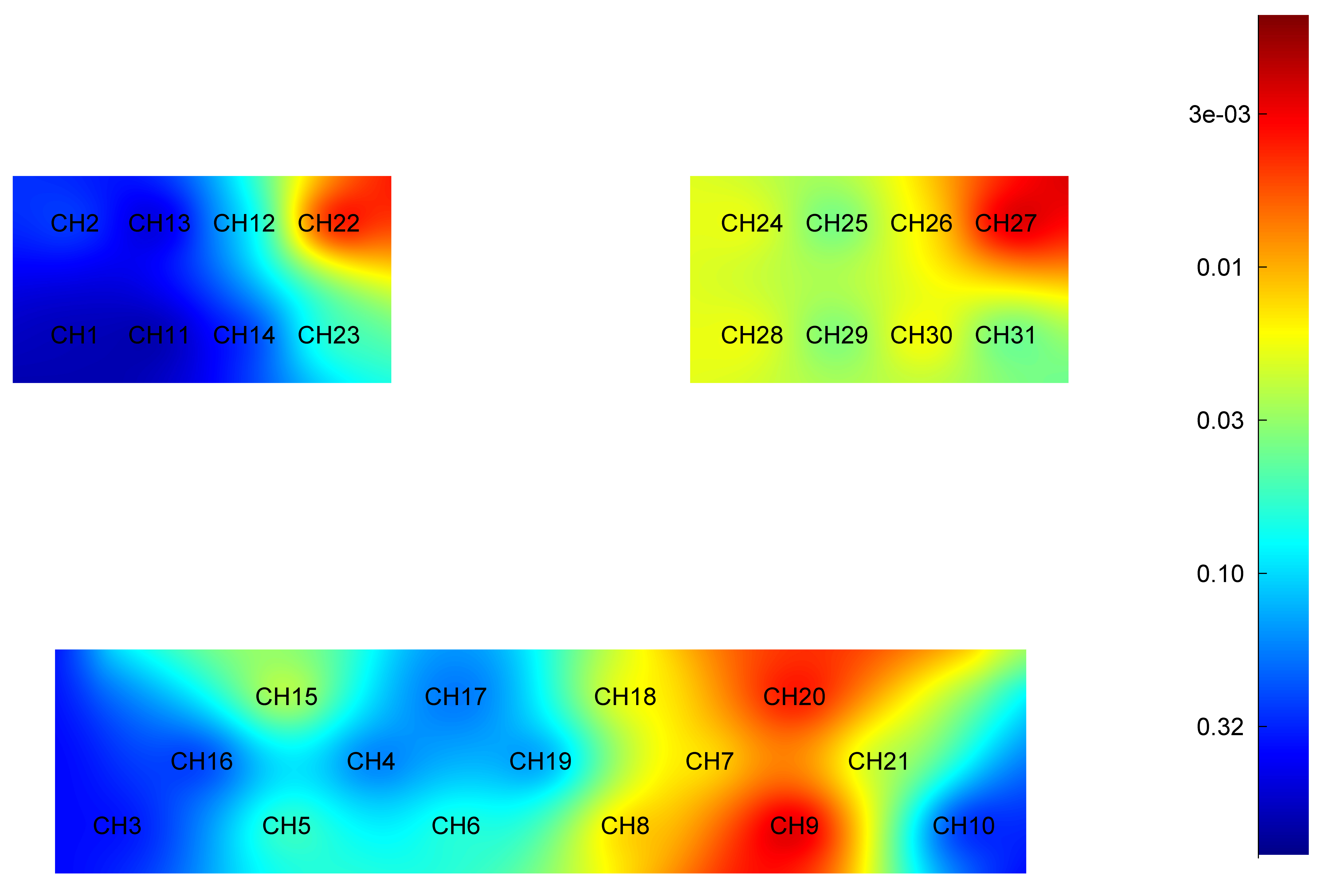
The independent sample t-test results for the HbO2 level during active elbow flexion in the younger and older groups are given in Table 2. Differences in the HbO2 level during active elbow flexion between the younger and older groups are illustrated in Fig. 2.
| Cortex | Younger | Older | t | p |
| Right PSC | 0.014 |
–0.001 |
1.407 | 0.165 |
| Right PMC | 0.056 |
–0.010 |
4.067 | 0.001 |
| Right SMC | 0.015 |
0.001 |
1.426 | 0.159 |
| Right PFC | 0.044 |
0.018 |
3.010 | 0.004 |
| Right DLPFC | 0.026 |
0.006 |
1.942 | 0.057 |
| Left PSC | 0.048 |
0.003 |
3.432 | 0.001 |
| Left PMC | 0.072 |
0.006 |
2.945 | 0.005 |
| Left SMC | 0.032 |
–0.006 |
3.569 | 0.001 |
| Left PFC | 0.048 |
0.001 |
4.626 | 0.001 |
| Left DLPFC | 0.048 |
0.020 |
2.736 | 0.008 |
PSC, primary somatosensory cortex; PMC, primary motor cortex; SMC, supplementary motor area; PFC, anterior prefrontal cortex; DLPFC, dorsolateral prefrontal cortex.
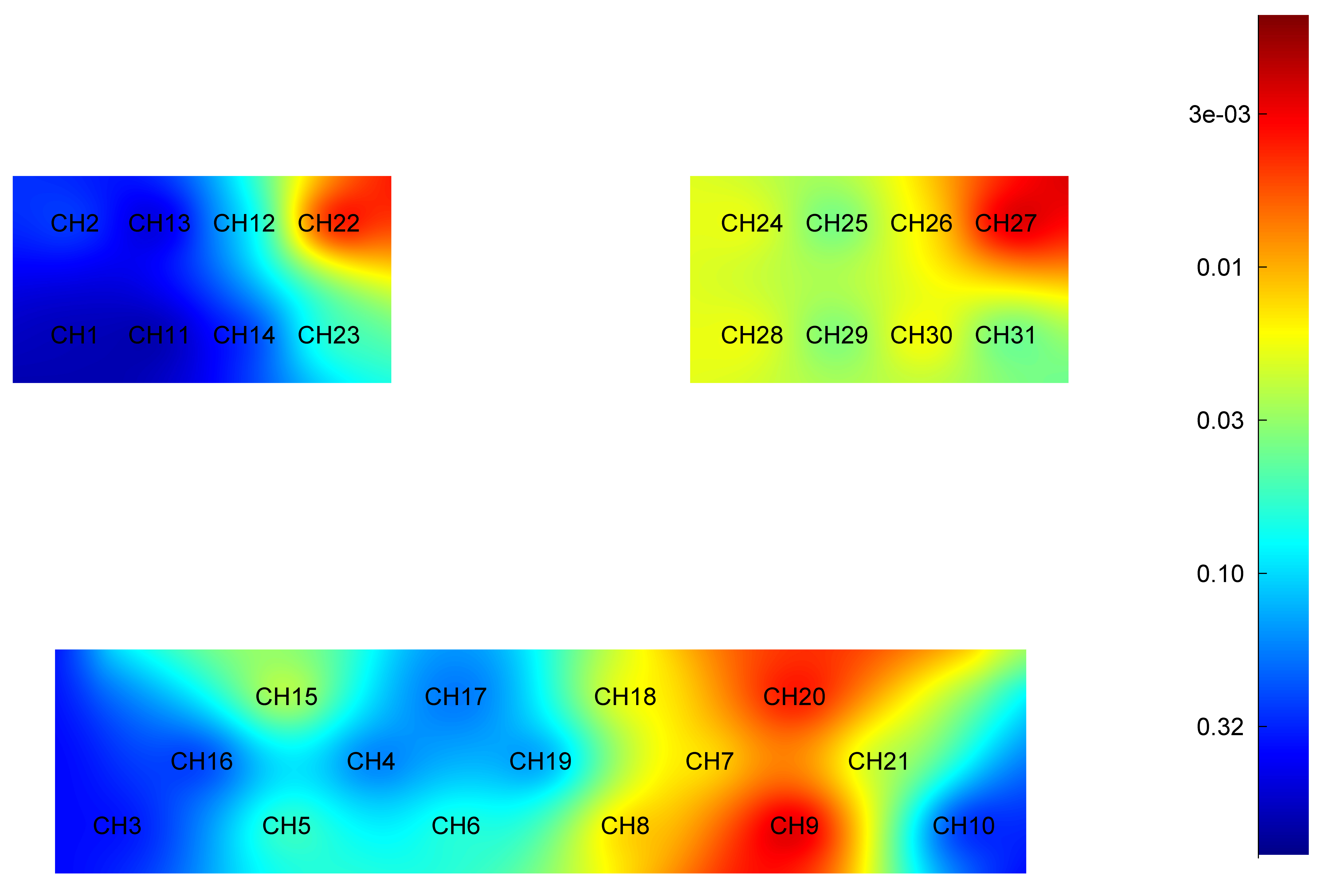 Fig. 2.
Fig. 2.
Difference in HbO2 level of each brain region during active elbow flexion between younger and older groups. Red: Greater significant difference. HbO2, oxyhemoglobin; CH, Channel.
Younger group: The mean and standard deviation of HbO2 in the cortex areas
(right PSC, right PMC, right SMC, right PFC, right DLPFC, left PSC, left PMC,
left SMC, left PFC, left DLPFC) are as follows: 0.014
Older group: The mean and standard deviation of HbO2 in the cortex areas (right
PSC, right PMC, right SMC, right PFC, right DLPFC, left PSC, left PMC, left SMC,
left PFC, left DLPFC) are as follows: –0.001
Specifically, the HbO2 level was significantly higher in younger adults when compared to older adults in cortex area: right PMC (t = 4.067, p = 0.001), left PMC (t = 2.945, p = 0.005), right PFC (t = 3.010, p = 0.004), left PFC (t = 4.626, p = 0.001), left PSC (t = 3.432, p = 0.001), left SMC (t = 3.569, p = 0.001) and left DLPFC (t = 2.736, p = 0.008).
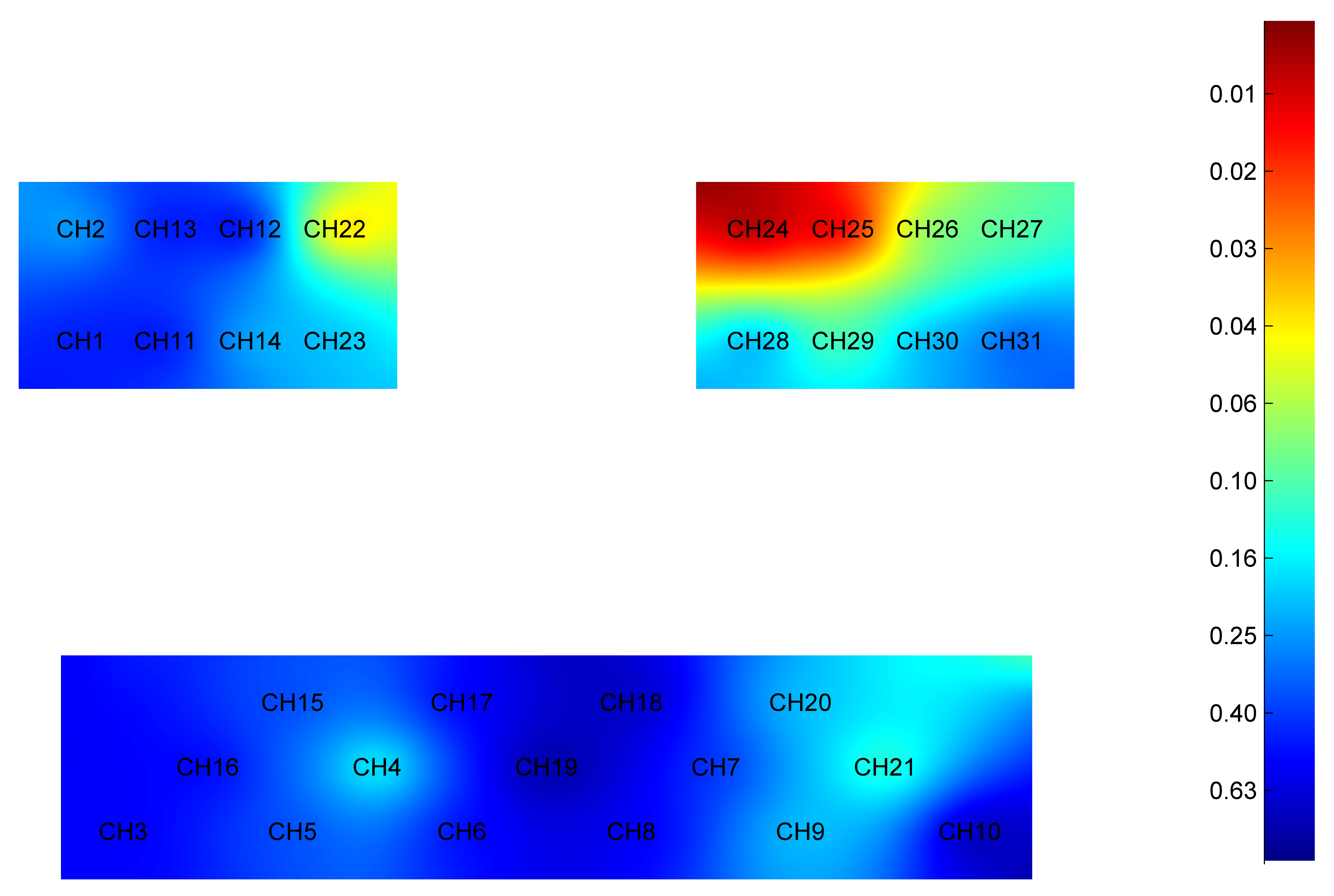
The independent sample t-test results for the HbO2 level during passive elbow flexion in the younger and older groups are given in Table 3. The differences in the HbO2 level during passive elbow flexion between the younger and older adults are illustrated in Fig. 3.
| Cortex | Younger | Older | t | p |
| Right PSC | –0.013 |
–0.016 |
0.299 | 0.766 |
| Right PMC | 0.010 |
–0.026 |
3.252 | 0.002 |
| Right SMC | –0.004 |
–0.016 |
1.345 | 0.184 |
| Right PFC | 0.007 |
–0.005 |
1.063 | 0.292 |
| Right DLPFC | 0.003 |
–0.002 |
0.387 | 0.700 |
| Left PSC | 0.040 |
–0.003 |
3.202 | 0.002 |
| Left PMC | 0.057 |
–0.009 |
3.951 | 0.001 |
| Left SMC | 0.023 |
–0.001 |
2.063 | 0.044 |
| Left PFC | 0.004 |
–0.005 |
0.741 | 0.462 |
| Left DLPFC | 0.031 |
0.013 |
1.666 | 0.101 |
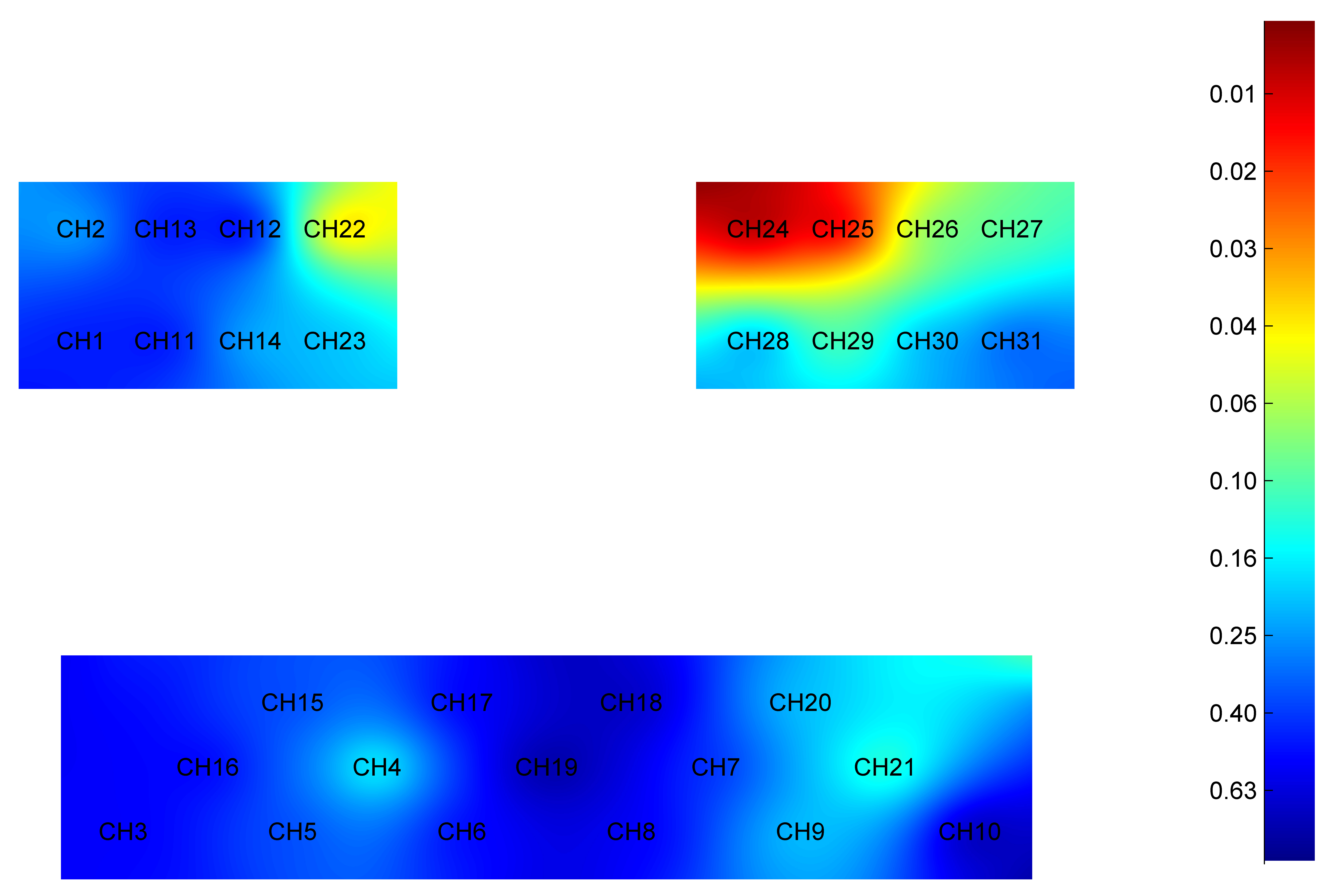 Fig. 3.
Fig. 3.
Difference in HbO2 level of each brain region during passive elbow flexion between younger and older groups. Red: Greater significant difference. HbO2, oxyhemoglobin; CH, Channel.
Younger group: The mean and standard deviation of HbO2 in the cortex areas
(right PSC, right PMC, right SMC, right PFC, right DLPFC, left PSC, left PMC,
left SMC, left PFC, left DLPFC) are as follows: –0.013
Older group: The mean and standard deviation of HbO2 in the cortex areas (right
PSC, right PMC, right SMC, right PFC, right DLPFC, left PSC, left PMC, left SMC,
left PFC, left DLPFC) are as follows: –0.016
Specifically, the HbO2 level was significantly higher in younger adults when compared to older adults in cortex areas: right PMC (t = 3.252, p = 0.002), left PMC (t = 3.951, p = 0.001), left PSC (t = 3.202, p = 0.002), and left SMC (t = 2.063, p = 0.044).
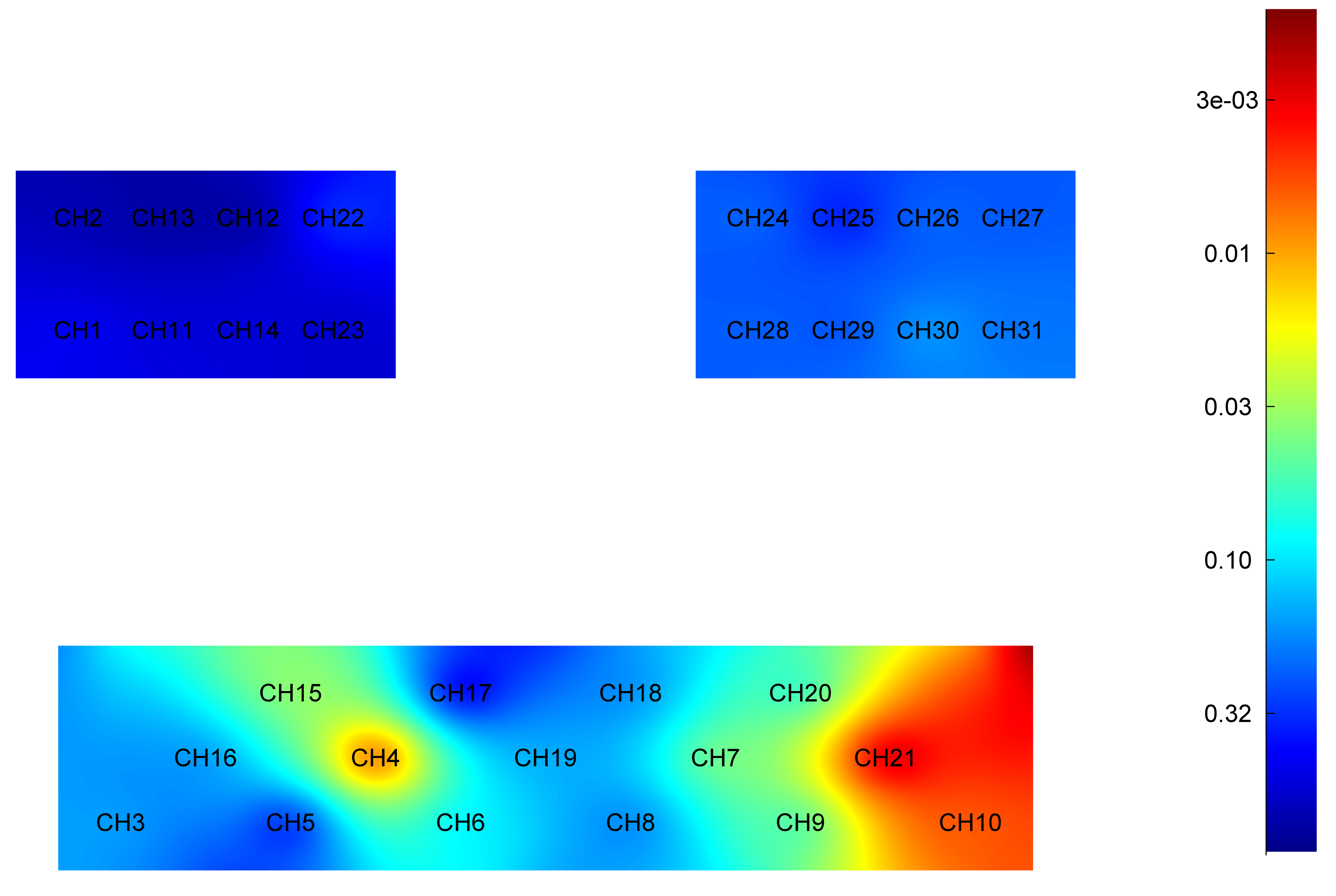
The independent sample t-test results for the HbO2 level during imagined elbow flexion in the younger and older groups are given in Table 4. The differences in the HbO2 level during imagining elbow flexion between the younger and older adults are illustrated in Fig. 4.
| Cortex | Younger | Older | t | p |
| Right PSC | 0.001 |
–0.001 |
0.139 | 0.890 |
| Right PMC | 0.017 |
–0.001 |
1.397 | 0.168 |
| Right SMC | 0.011 |
0.003 |
0.940 | 0.351 |
| Right PFC | 0.027 |
0.005 |
2.180 | 0.033 |
| Right DLPFC | 0.030 |
0.005 |
2.911 | 0.005 |
| Left PSC | 0.011 |
–0.004 |
1.586 | 0.118 |
| Left PMC | 0.020 |
–0.001 |
1.630 | 0.108 |
| Left SMC | 0.018 |
0.003 |
1.954 | 0.056 |
| Left PFC | 0.026 |
0.000 |
2.878 | 0.006 |
| Left DLPFC | 0.046 |
0.007 |
4.382 | 0.001 |
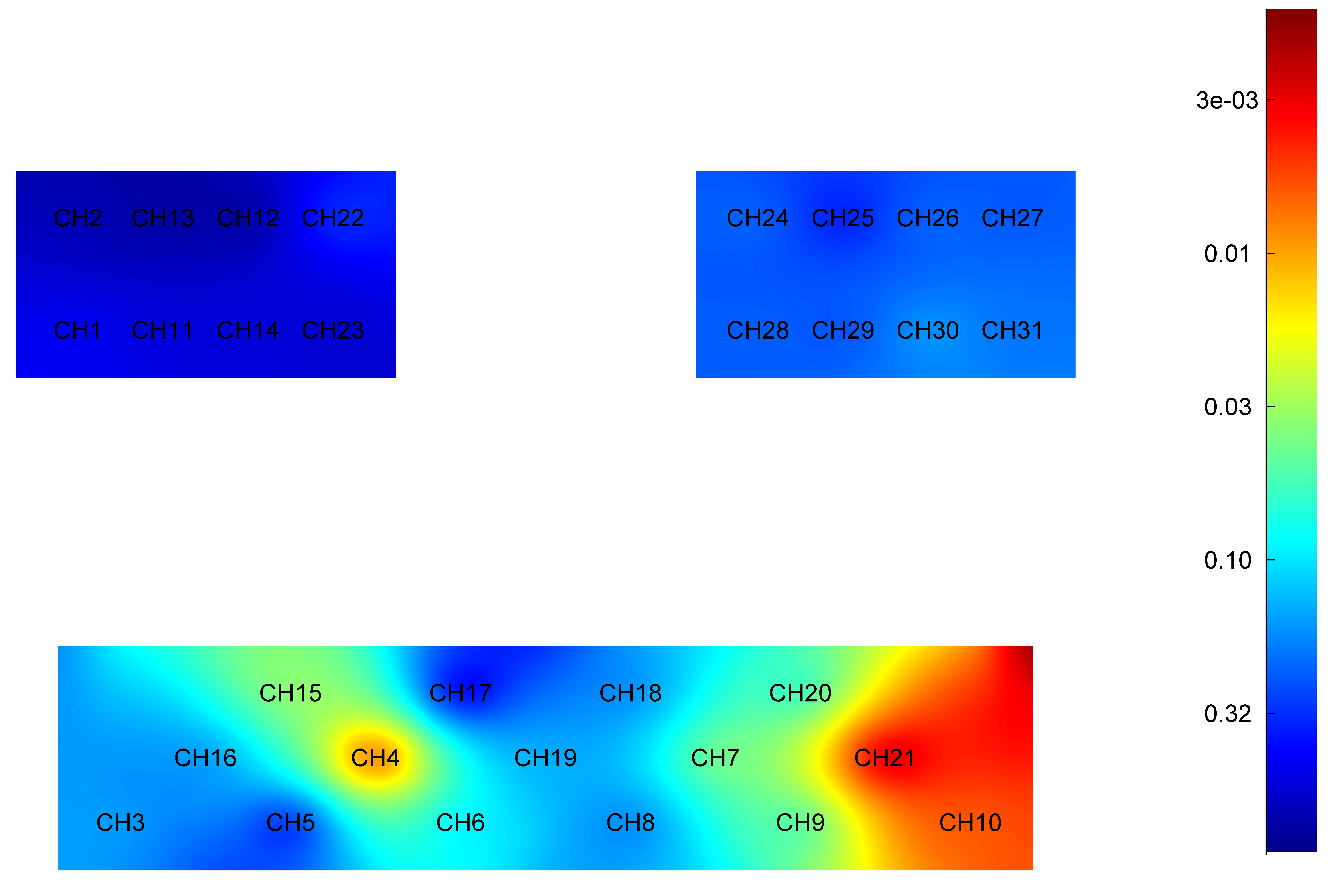 Fig. 4.
Fig. 4.
Difference in HbO2 level of each brain region during imagined elbow flexion between younger and older groups. Red: Greater significant difference. HbO2, oxyhemoglobin; CH, Channel.
Younger group: The mean and standard deviation of HbO2 in the cortex areas
(right PSC, right PMC, right SMC, right PFC, right DLPFC, left PSC, left PMC,
left SMC, left PFC, left DLPFC) are as follows: 0.001
Older group: The mean and standard deviation of HbO2 in the cortex areas (right
PSC, right PMC, right SMC, right PFC, right DLPFC, left PSC, left PMC, left SMC,
left PFC, left DLPFC) are as follows: –0.001
Specifically, the HbO2 level was significantly higher in younger adults when compared to older adults in cortex areas: right PFC (t = 2.180, p = 0.033), left PFC (t = 2.878, p = 0.006), right DLPFC (t = 2.911, p = 0.005) and left DLPFC (t = 4.382, p = 0.001).
This study used fNIRS to investigate the characteristics of and differences between younger and older adults in HbO2 level during motor execution, motor passivity and motor imagery processes. During the motor execution task, younger adults presented greater activation than older younger adults in cortex area right PMC, left PMC, right PFC, left PFC, left PSC, left SMC and left DLPFC, these brain regions being strongly associated with motor execution [43, 44, 45]. Andrews-Hanna et al. [46], found that compared to healthy younger adults, older adults show weaker levels of brain activation and perform poorly in various cognitive domains such as processing speed, memory and executive functions. They concluded that the age-related coordinated disorder of large-scale brain systems leads to cognitive decline. It is also not ruled out that with aging, the reduction in gray matter volume reduces activity in brain regions directly related to executive function [47]. Healthy brain aging can lead to sensory-motor cortex integration disorders [48] and the reduced level of neuronal metabolism is associated with an associated motor decline [49].
During the motor passivity task, younger adults presented greater activation than older adults in cortex areas: right PMC, left PMC, left PSC and left SMC and no significant differences were found in cortex areas: right PFC, left SMC and left DLPFC. This may be related to the characteristics of the motor task. Loibl et al. [50] found that the differences in motor passivity between the older and younger adults were related to the complexity of the motor tasks. When performing a simple wrist flexion task, ipsilateral PMC and SMC activation increased significantly in older adults, but when performing a complex force grip task, contralateral PSC and PMC activation were more significant in younger adults. However, the main activation areas were SMC, PMC and PSC [51], which showed similar activation areas found to the results reported here.
Simultaneously, during the motor imagery task, younger adults presented greater activation than older younger adults in cortex areas: right PFC, left PFC, left PSC, right DLPFC and left DLPFC, but no significant differences were found in the remaining brain regions. Motor imagery is an advanced type of brain thought activity and there is no consensus on which brain areas cause activation. This may be related to the different experimental methods and the complexity of the designed motor imagery task [52, 53]. However, the small activation effect of motor imagery on PMC did not reflect significant differences, which is consistent with other studies [54, 55].
According to the results reported here, it is not difficult to find that three motor control conditions examined activated specific brain regions and that there were age-related differences in HbO2 level; with an older group exhibiting lower HbO2 levels when compared to a younger group. Research reports have indicated that brain blood flow in older people undergoes changes when compared to younger individuals [56, 57]. Positron emission tomography and single-photon emission computed tomography scans have shown that local brain blood flow in the association cortex and the edge cortex decreases with age [57, 58]. Furthermore, fMRI studies have revealed that, when compared to younger individuals, older individuals exhibited fewer activated voxels during motor tasks, simple reaction time tasks and visual stimulus activation [59, 60]. Additionally, Hock et al. [56] demonstrated through fNIRS that older individuals exhibit less increase in oxygenated hemoglobin and total hemoglobin in the dorsolateral prefrontal cortex during cognitive tasks when compared to younger individuals. Hogstrom et al. [61], found that cortex area decreases with age and this process is nonlinear. The results presented here show different motor control conditions are associated with different HbO2, which may be related to motor control intensity. Therefore, different motor controls have different HbO2 levels and undergo different changes with age.
HbO2 metabolism provides a direct in vivo measurement tool for assessing neuronal metabolism and biological activity, reflecting neuronal activity in normal resting, activated, or pathological states [62]. HbO2 metabolism is crucial in the normal aging process. Consistent with numerous previous research findings, this study also observed a falling trend in HbO2 level with age. In theory, the decrease in HbO2 metabolism with age may be due to structural changes. Micro anatomical changes in the cortex during normal aging include reduced synaptic density, decreased neuronal density and a decrease in average neuron size, any of which could reduce local metabolic demand, leading to the observed reduction in HbO2 levels [63, 64, 65].
It is worth noting that age-related changes in blood oxygenation have previously been reported. Ances et al. [66] noted a decrease in HbO2 level at rest but a slight increase in oxygen extraction rate with age. Furthermore, Hock et al. [56] found a significant reduction in HbO2 measured by near-infrared spectroscopy during cognitive tasks in older individuals. This finding suggests that the coupling between oxygenation and neuronal activation has changed and is consistent with the reduced blood oxygen level dependent response observed in the present study. Given the complex interplay between neuronal activation and oxygenation, the findings reported here may represent reduced neuronal activation and reduced vascular responses to different motor control conditions in older individuals.
As fNIRS is a relatively new method, it is important to consider both its advantages and limitations. Importantly, in the fNIRS method implemented here, it was not possible to obtain an absolute baseline HbO2 concentration. This would require accurate quantification of the path-length of near-infrared light in brain tissue for each wavelength obtained for every observation. Since absolute baseline values are unknown, measurements are limited to relative changes in absolute concentration. Some might argue that brain volume in the studied older individuals is smaller than in younger individuals due to brain atrophy with aging. However, brain atrophy data measured by computed tomography in adult brains indicate only mild atrophy between the ages of 20 and 60, with significant atrophy only occurring subsequently [67]. Given that the average age of the older participants in this study was 60 years, this group may only exhibit mild atrophy compared to the younger group. It is also important to note that channel placement affect.
HbO2 level and these levels vary if the angular position of optodes change [68]. To minimize the influence of different optodes, identical positions were selected in each subject. HbO2 levels in the adult head remain constant once the optode distance exceeds 2.5 cm. In this study, the optode separation was three centimeters.
Analyzing the HbO2 changes during different motor control conditions further helps understand the relationship between changes in motor control function during both the aging process and oxygen metabolism. In the future, a more in-depth exploration of HbO2 changes in different contexts should be conducted in the presence of different stimuli such as drugs, cognitive training and transcranial direct current stimulation. This will lay a foundation for disease research. Additionally, fNIRS can be combined with other methods such as magnetoencephalography and electroencephalography to assess neurological function [69]. Despite drawbacks in its current implementation, such as spatial resolution limitations and quantitative uncertainty due to the absence of precise measurement capabilities for optical path lengths, fNIRS still holds the potential to provide non-invasive assessment of cerebral HbO2, offering information in real-time at the bedside that no other non-invasive methods can currently provide.
This study provided evidence that HbO2 levels are different in the primary and association cortices during different motor control conditions in young and older adults. HbO2 levels in different brain regions under different motor control conditions can be influenced by age.
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
JL and CW contributed to conception and design. DY performed the experiments. ZY analyzed the data. DY wrote the paper. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
The present research was approved by the Biomedical Research Ethics Committee of the Second Affiliated Hospital of Nanchang University (BR/AFISG-04/1.0). After a complete description of the study, written informed consent was obtained from all patients or their families/legal guardians.
We thank two anonymous reviewers for their comments and suggestions which helped improve the manuscript.
This research was supported by Project of the Science and Technology Department of Jiangxi Province, China (20224ABC03A02).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.