






1 Department of Pain, Huadong Hospital, Shanghai Key Laboratory of Clinical Geriatric Medicine, Fudan University, 200040 Shanghai, China
2 Department of Anesthesiology, Shanghai Jiao Tong University Affiliated Sixth People's Hospital, 200233 Shanghai, China
3 Department of Anesthesiology, Shanghai Key Laboratory of Maternal Fetal Medicine, Shanghai Institute of Maternal-Fetal Medicine and Gynecologic Oncology, Shanghai First Maternity and Infant Hospital, School of Medicine, Tongji University, 20092 Shanghai, China
4 Department of Critical Care Medicine, Shanghai Jiao Tong University Affiliated Sixth People's Hospital, 200233 Shanghai, China
†These authors contributed equally.
Abstract
Background: Postoperative cognitive dysfunction (POCD) is a common postoperative complication in elderly patients. The purpose of this study was to investigate the mechanism through which metformin improves postoperative cognitive function. Methods: In the in vivo experiment, 18-month-old Sprague–Dawley (SD) rats were randomly divided into four groups (n = 12 in each group): the control, metformin, operation, and operation plus metformin groups. The animals were pretreated with metformin by gavage once daily for two weeks. The Morris water maze (MWM) was used to measure cognitive ability. In the in vitro experiment, BV2 cells were divided into five groups: the control, metformin, lipopolysaccharide (LPS), LPS plus metformin, and LPS plus metformin plus compound C groups. We stimulated microglia with LPS (500 ng/mL). Immunofluorescence and Western blotting were used to assess ROS (reactive oxygen species) levels, autophagy-associated protein levels and adenosine monophosphate-activated protein kinase (AMPK)/regulator factor 2-related enzyme 1 (SIRT1) signaling pathway activity in the rat cortex and microglial cells. Results: In the MWM test, the metformin-pretreated rats spent a higher proportion of time in the target quadrant. Immunofluorescence showed that the fluorescence intensity of LC3 in the cortex was increased in rats pretreated with metformin. Western blotting indicated that metformin upregulated the expression of autophagy-related and AMPK/SIRT1 signaling pathway-related proteins in the cortex after surgery. By activating the AMPK/SIRT1 signaling pathway in vitro, metformin reduced microglial activation and oxidative stress and promoted autophagy. Conclusions: Through the AMPK/SIRT1 pathway, metformin can boost autophagy and reduce oxidative stress in cortical microglia in older rats, in turn improving postoperative cognitive function.
Keywords
- metformin
- postoperative cognitive dysfunction
- autophagy
- oxidative stress
- AMPK/SIRT1 pathway
Postoperative cognitive dysfunction (POCD) is characterized by decreased attention and consciousness and impaired learning, memory and cognition after surgery and is especially common among elderly patients [1]. The mechanism is still unclear. Therefore, there is a lack of effective prevention and treatment methods.
Autophagy is essential for maintaining homeostasis and related activity, such as the breakdown of metabolic waste and misfolded proteins [2]. Studies have shown that a sufficient level of autophagy, especially in the brain and heart, is essential for extending life and maintaining organ function [3, 4]. Several studies have found that autophagy dysfunction occurs in the brain after surgery [5, 6, 7]. Moreover, a declining level of autophagy is strongly associated with cognitive impairment in multiple neurodegenerative diseases, while enhancement of autophagy can maintain normal cognition [8, 9]. Beclin1, p62 and LC3 are considered the core regulators of autophagy in mammals [10, 11]. Thus, the levels of these proteins were assessed in our study to evaluate the level of autophagy in the cortical layer in rats with POCD and cultured microglia stimulated with lipopolysaccharide (LPS).
Metformin is one of the traditional first-line medications for the treatment of type 2 diabetes mellitus (T2DM). As research on metformin has increased, it has been found to have even more therapeutic effects. For example, metformin has been confirmed to protect against cognitive decline in many neurodegenerative diseases [12, 13, 14]. It is still unclear exactly how metformin works at the molecular level to produce its effects. Metformin has been shown to easily cross the blood‒brain barrier (BBB) and to reach areas such as the hippocampus, cortex, and pituitary [15]. According to previous studies, it primarily has antineuroinflammatory and antioxidative effects and can ameliorate tau pathology and maintain synaptic function to protect cognition [16, 17, 18, 19].
The major target of metformin is adenosine monophosphate-activated protein kinase (AMPK), which is a ubiquitous serine/threonine protein kinase that controls cellular energy metabolism. An increasing body of evidence suggests that rats with cognitive decline exhibit lower levels of AMPK [20, 21, 22]. In Alzheimer’s disease and diabetes, activation of the AMPK-mTOR pathway can reduce neuronal death and improve cognitive function [23, 24]. Moreover, AMPK can participate in the entire process of autophagy [25]. A study confirmed that AMPK and autophagic flux are strongly related [26]. One of the important downstream targets of AMPK is silent information regulator factor 2-related enzyme 1 (SIRT1), which can also regulate cell energy metabolism and other physiological activities. Similar to AMPK, SIRT1 plays a vital role in age-related neurodegenerative diseases [27]. Activating the AMPK/SIRT1 pathway in Alzheimer’s disease can improve behavior, lower oxidative stress, and reduce neuroinflammation [28]. Similar results were found in Parkinson’s disease [29].
To confirm the effectiveness and potential mechanism of metformin in preventing POCD, we assessed the biomarkers of the AMPK/SIRT1 pathway, autophagy, neuroinflammation, and oxidative stress in the cortex of rats with POCD and in cultured BV2 cells (microglia) with LPS stimulation. Besides, we observed whether metformin pretreatment had an ameliorative effect on cognitive function in POCD-model rats by neurobehavioral test.
Metformin (CAS: 1115-70-4), compound C (CAS: 866405-64-3), and LPS (L2880) (all from Sigma–Aldrich, St. Louis, MO, USA) were utilized in this experiment. Metformin was dissolved in normal saline and used in the animal experiments. For the cell experiments, metformin, compound C (an AMPK inhibitor) and LPS were dissolved in PBS and used to treat microglia. AMPK (polyclonal, 2532), p-AMPK (recombinant, 2535), SIRT1 (recombinant, 9475), LC3 (polyclonal, 2775), Beclin1 (polyclonal, 3738), and p62 (polyclonal, 5114) antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA). A CD68 (monoclonal, ab955) antibody was obtained from Abcam (Cambridge, MA, USA). A glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibody (monoclonal, 60004-1-Ig) was purchased from Proteintech (Wuhan, China). Dulbecco’s modified eagle’s medium (DMEM) and fetal bovine serum (FBS) were obtained from Thermo Fisher Scientific, Inc (Waltham, MA, USA).
Male Sprague‒Dawley (SD) rats aged 18 months and weighing 500 g each were purchased from the Shanghai Laboratory Animal Center of the Chinese Academy of Science (Shanghai, China) and housed in accordance with industry standards at 22–24 °C on a 12-hour light–dark cycle. All procedures were approved by the Shanghai Huadong Hospital’s Animal Research Committee (approval number: 2022110029S) and carried out in accordance with the National Institutes of Health’s Guide for the Care and Use of Laboratory Animals. All rats were randomly divided into four groups (n = 12 per group): the control, metformin, operation, and operation plus metformin groups. The rats underwent splenectomy to establish a POCD animal model. Before surgery, the rats were fasted for 12 hours. The animals were pretreated with 200 mg/kg metformin by oral gavage once daily for two weeks.
After anesthetization via intraperitoneal injection of 2% pentobarbital sodium (Sigma-Aldrich, St. Louis, MO, USA) (40 mg/kg), the rats were placed on a sterile operating table. The skin on the abdomen was prepared, and after disinfection, we made a 1.5 cm incision along the midline of the abdomen. The spleen was located, and then the blood vessels of the root of the spleen were ligated. After successful splenectomy, we closed the abdominal incision with sterile sutures. Finally, the rats were placed on a heated blanket for recovery. The Morris water maze (MWM) test was conducted three days later.
To prevent infection after surgery, the training and test phases of the MWM test were performed from postoperative day 3 to postoperative day 8. The protocol was carried out as described previously [22]. The MWM consisted of a circular pool with a diameter of 180 cm and height of 50 cm. The pool was filled halfway with water, and the temperature of the water was maintained between 21 and 23 °C. The circular pool was divided into four quadrants (northeast, northwest, southeast, southwest). The platform, which had a diameter of 12 cm, was placed in the northwest quadrant slightly below the water surface. In our research, the process of training and testing in water maze was performed between 8 AM and 11 AM. There are opaque curtains that completely block the circular pool, so it can avoid visual interference. The animals’ movements were captured by a camera placed above the maze’s center, and Any-Maze software was used to analyze the data, which included the swimming speed, escape latency, and amount of time spent in the target quadrant. The test lasted for six days. The rats were placed in the water in a randomly selected quadrant near the pool wall on the first five days, i.e., during the training phase. If a rat could not locate the platform within 60 seconds, it was placed on the platform and kept there for 20 seconds to help it develop a memory of the platform location. On the final day, the platform was removed, the rats were placed in the water in the southeast quadrant, and the percentage of time spent in the target quadrant was recorded. After completing the water maze test, the rats were killed for sampling with inhaling excess carbon dioxide.
BV2 microglial cells were purchased from the Shanghai Laboratory Animal Center
of the Chinese Academy of Science (Shanghai, China). The cells were identified in terms of morphology, phenotype, and function. The mycoplasma testing has been done for the cell lines used in the experiments. The cells were cultured in high-glucose
DMEM containing 10% FBS and 1% penicillin‒streptomycin. The cells were grown in
a 37 °C incubator containing 5% CO
Following the in vitro and in vivo interventions, protein was
extracted from rat cortex samples and BV2 cells using 4 °C RIPA buffer
(Beyotime Institute of Biotechnology, Shanghai, China). The proteins were then
centrifuged at a speed of 12000
In addition to Western blotting, rat cortex samples were used for
immunofluorescence. They were removed immediately, stored in 4%
paraformaldehyde, embedded in paraffin, and sliced into approximately 5 µm
sections for staining. A primary antibody against LC3 (1:200) was used. The
cortical sections were washed with 4 °C PBS and placed in a wet box.
After that, they were incubated with the LC3 antibody. Following a second wash in
PBS, the sections were incubated at room temperature with a FITC-conjugated
anti-rabbit IgG secondary antibody. DAPI (4
To measure the ROS level, we used Reactive Oxygen Species Assay Kit (Beyotime, Shanghai, China). BV2 microglia were incubated with antibodies against oxidative stress indicators in 6-well plates for 30 min. The cells were then removed and washed 3 times with PBS for 3 minutes each. After that, we used a fluorescence microscope (TCSSP5, Leica Microsystems GmbH, Wetzlar, Germany) to determine the fluorescence intensity.
Statistical analysis was performed with GraphPad Prism 6.0 (GraphPad Software
Inc., La Jolla, CA, USA). The continuous variables in our experimental data have
been tested for normality. And all data in this study conform to normal
distribution. The means and standard error of the mean (SEMs) are displayed for all data. Multiple group
comparisons were analyzed with repeated measures analysis of variance (ANOVA), followed by a
Bonferroni’s post hoc test. An unpaired, two-tailed Student’s t-test was
applied to compare between two groups. The results were considered significant at
p
The experimental layout is depicted in Fig. 1A. The rats were pretreated with metformin for two weeks. Three days after the operation, cognitive tests, including the positioning navigation and spatial exploration experiments, were performed. According to a previous study [21, 32], surgery and inhalation anesthetics can both lead to cognitive dysfunction in rodent models, especially in aged animals, and the same result was found in our study. The operation clearly had a significant impact on the learning and memory abilities of old rats, as evidenced by the fact that the escape latency was significantly longer (Fig. 1B) and the percentage of time spent in the target quadrant was lower in the operation group than in the other groups (Fig. 1C). The rats pretreated with metformin did not display significant cognitive impairment, and they spent more time in the target quadrant (Fig. 1C) and had a shorter escape latency (Fig. 1B) than the rats in the operation group. Swimming speed was not significantly different among the four groups (Fig. 1D), indicating that the surgery had no impact on the motor ability of the aged rats. These results suggested that metformin may prevent age-related cognitive decline in rats after surgery.
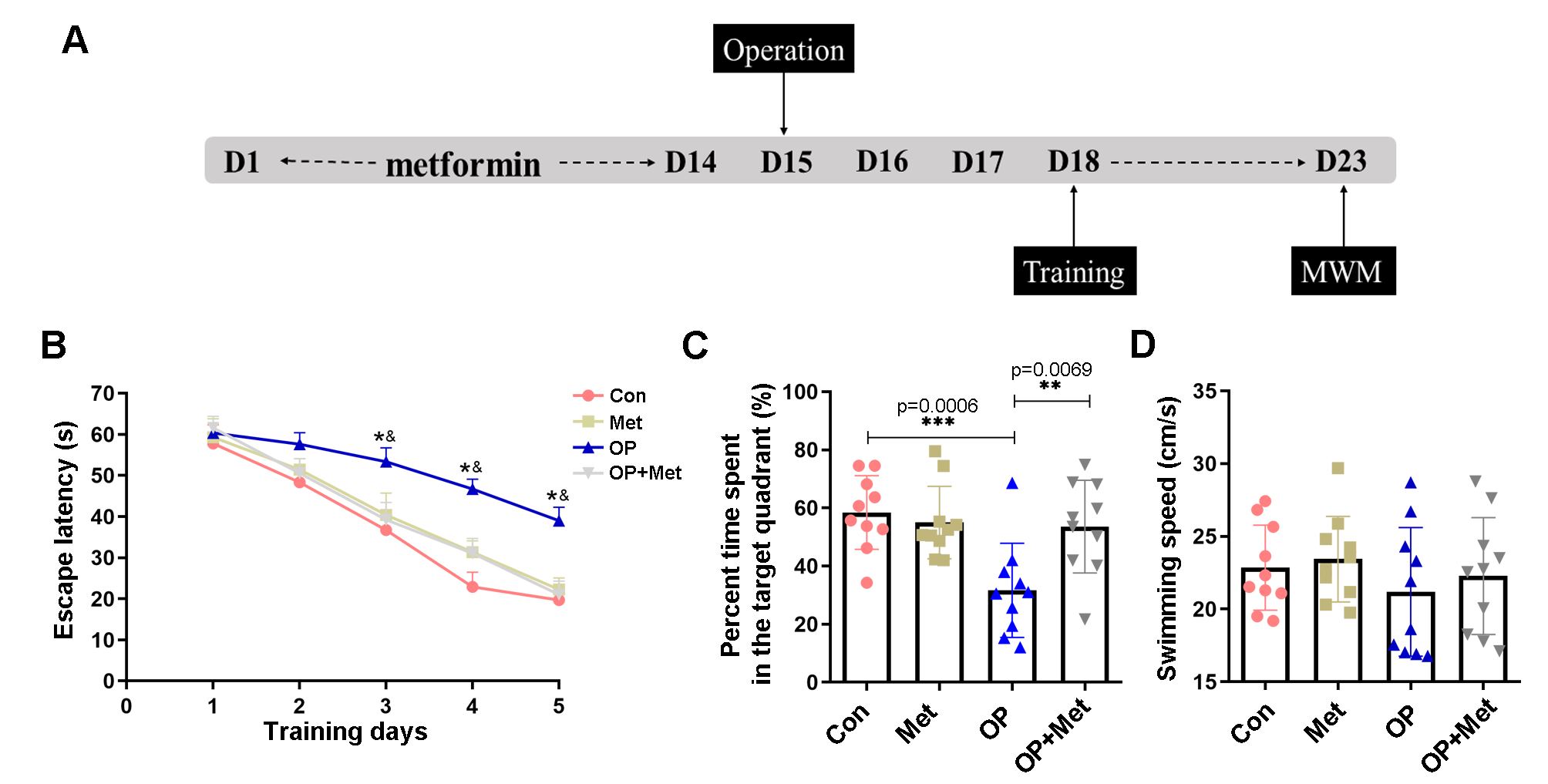 Fig. 1.
Fig. 1.Metformin alleviated surgery-induced learning and memory
impairment. (A) Timeline of metformin intervention and the MWM test (n = 12 each
group). Before splenectomy, the rats received metformin by oral gavage once daily
for two weeks and then underwent behavioral tests. (B) The escape latency refers
to the average time required to find the target platform in the training phase.
(C) During the testing phase, the metformin-treated rats spent a greater
percentage of time in the target quadrant. (D) The average swimming speed was
used to assess differences in athletic ability in the MWM test. The data are
shown as the mean
To elucidate the possible mechanism by which metformin can prevent POCD, we
investigated whether microglial autophagy occurs in the cerebral cortices of aged
rats. Autophagy was activated in the cortical homogenates of rats pretreated with
metformin, as demonstrated by a significant increase in LC3 and Beclin1 levels
(Fig. 2A,B). In contrast, p62 expression in the cortex was significantly
higher in the operation group than in the other groups (Fig. 2A,B). One of
the most commonly used autophagy markers is LC3, and the LC3II/I ratio was used
to assess the level of autophagy in the present study. We used immunofluorescence
staining to evaluate the immunoreactivity of LC3 (Fig. 2E). Consistently, the
operation group showed a lower fluorescence intensity, while metformin
pretreatment reversed the surgery-induced reduction in autophagy (Fig. 2E).
Microglia are important immune cells in the central nervous system that can be
activated by external stimulation, and one of the markers of microglial
activation is CD68. Once activated, microglia release inflammatory factors such
as IL-1
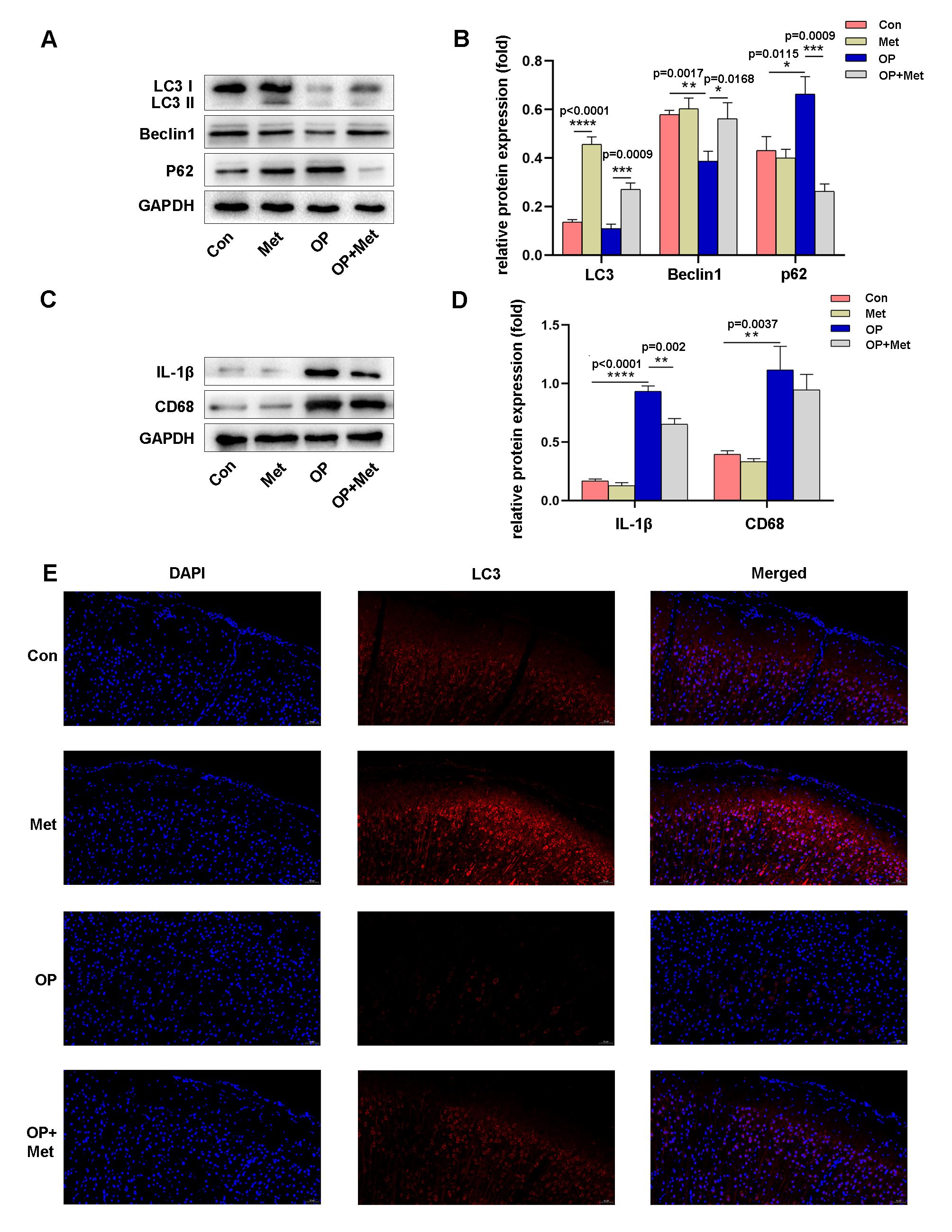 Fig. 2.
Fig. 2.Metformin increased autophagy and inhibited microglial activation in the cerebral cortex. (A,B) Western blot analysis and quantification of Autophagy associated proteins (LC3, p62, Beclin1) expression in the cortex in the control, metformin, operation and operation+metformin groups. (C,D) Western blot analysis and quantification of IL-1
Microglia typically have a round or oval shape under physiological conditions. After pretreatment with 2 mM metformin, microglia had a nearly identical morphology as those in the control group, indicating that at this dose, metformin had little effect on cell viability. After LPS stimulation, microglial cells underwent obvious morphological changes, exhibiting an irregular shape and protrusions. However, metformin prevented these morphological changes in microglia to some extent (Fig. 3A).
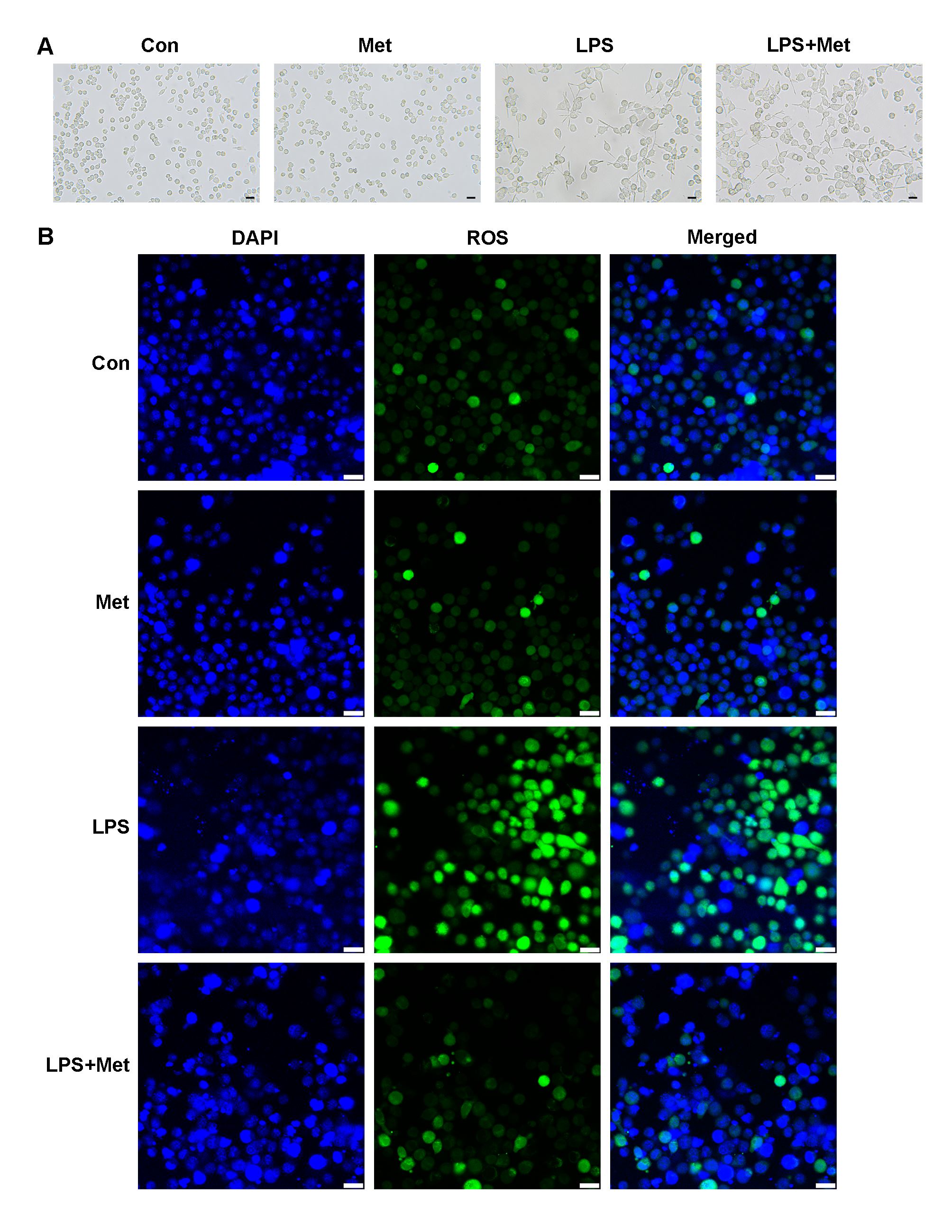 Fig. 3.
Fig. 3.Metformin changed the morphology and oxidative stress status of microglia. (A) Images of microglial morphology. (B) Immunofluorescence staining of ROS in each group. The scale bar represents 50 mm. Met, metformin; LPS, Lipopolysaccharide; ROS, reactive oxygen species.
Microglial state, in addition to cell morphology, was changed. LPS can induce oxidative stress in cells. The results are displayed in Fig. 3B. The fluorescence intensity of ROS was measured to determine ROS levels. In the LPS group, ROS levels were the highest. Although the ROS fluorescence intensity in the metformin pretreatment group was comparable to that in the control and metformin groups, the comparison between the metformin pretreatment group and the LPS group suggested that metformin had a protective effect, lowering the levels of ROS.
Microglia were stimulated with LPS in vitro to simulate neuroinflammation in rats with POCD, and changes in autophagy in the cells were assessed. The results were similar to those observed in vivo. Western blotting revealed that the levels of LC3 and Beclin1 were significantly increased in microglia pretreated with metformin, indicating that autophagy was triggered in these cells (Fig. 4A–E). However, LPS significantly inhibited autophagy in microglia (Fig. 4A–E).
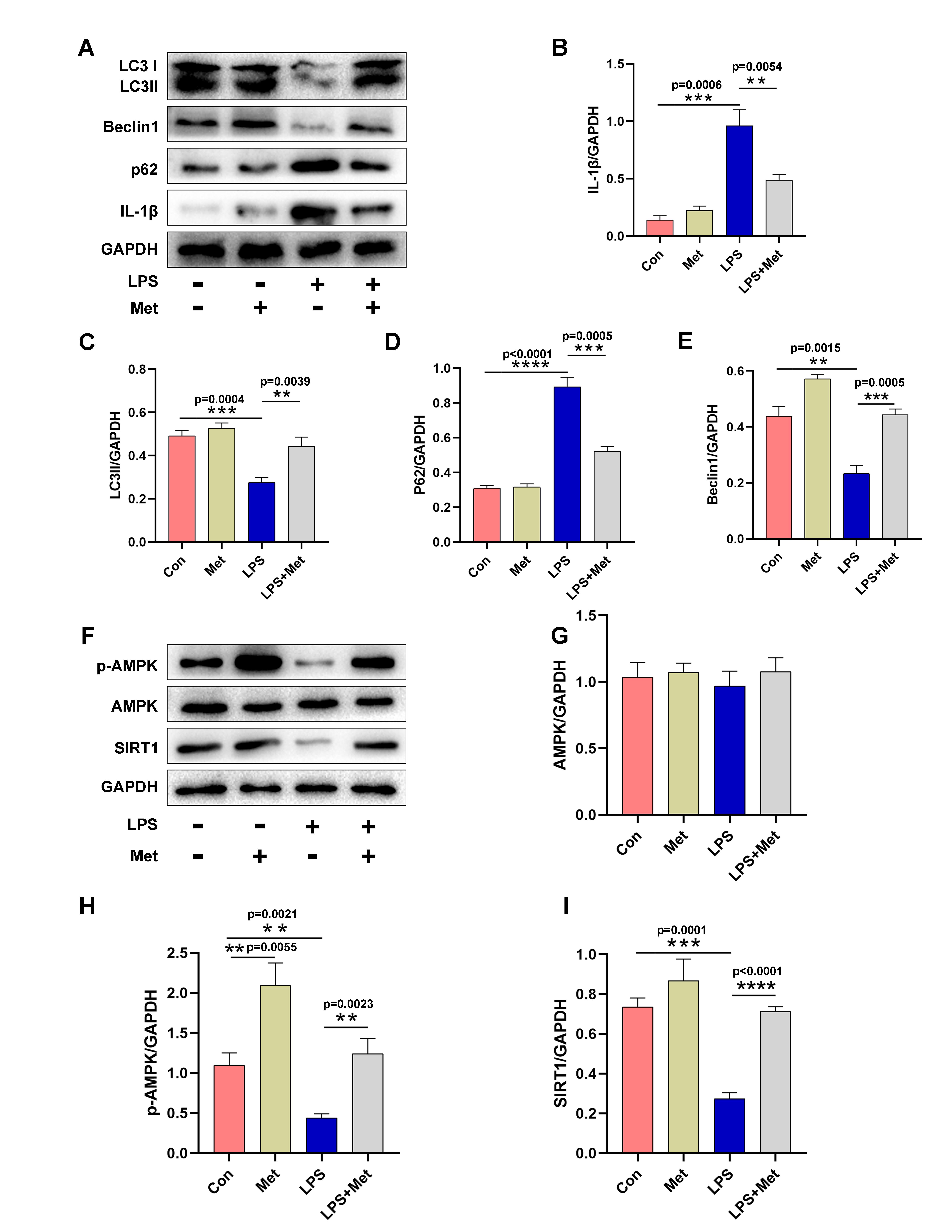 Fig. 4.
Fig. 4.Metformin increased autophagy and activated the AMPK/SIRT1 signaling pathway in vitro. (A–E) Western blot analysis and quantification of LC3, p62, Beclin1 and IL-1
According to a previous report, the regulation of autophagy is intimately tied to the AMPK/SIRT1 signaling pathway [26]. Thus, we further investigated whether this signaling pathway participates in LPS-induced neuroinflammation in microglia. In this part of the experiment, metformin pretreatment clearly enhanced the AMPK/SIRT1 interaction in microglia, as shown by an increase in AMPK and SIRT1 phosphorylation (Fig. 4F,H,I). However, compound C (a popular AMPK inhibitor) inhibited metformin-induced activation of the AMPK/SIRT1 signaling pathway, as evidenced by a reduction in AMPK and SIRT1 phosphorylation (Fig. 5C,D). Additionally, in microglia treated with LPS, compound C increased the expression of p62 while decreasing the expression of LC3 and Beclin1 (Fig. 5A,B). The ROS fluorescence intensity was not significantly different between the LPS plus metformin group and the compound C group (LPS+Met+CC). Nevertheless, the ROS fluorescence intensity in these groups was lower than that in the LPS group. According to the aforementioned findings, metformin activates the AMPK/SIRT1 pathway to reduce POCD-induced autophagy inhibition in cortical microglia.
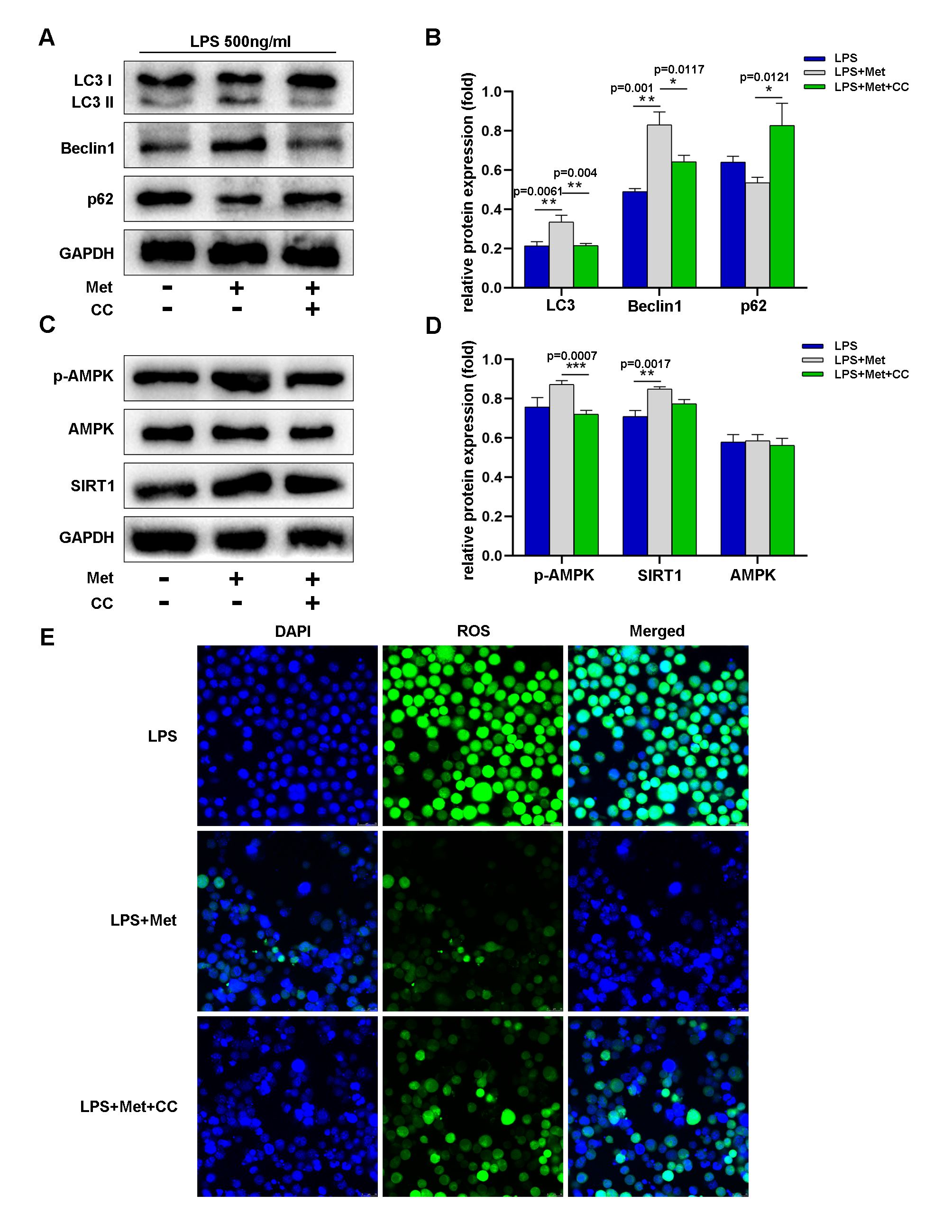 Fig. 5.
Fig. 5.During LPS-induced neuroinflammation, metformin promoted autophagy by activating the AMPK/SIRT1 signaling pathway and alleviated microglial oxidative stress in vitro. (A,B) Western blot analysis and quantification of LC3, p62 and Beclin1 levels during LPS-induced neuroinflammation in the LPS, LPS+operation, and LPS+operation+compound C groups. (C,D) Western blot analysis and quantification of phosphorylated and total AMPK and SIRT1 levels during LPS-induced neuroinflammation in the LPS, LPS+operation, and LPS+operation+compound C groups. (E) Immunofluorescence staining of ROS in each group. The scale bar represents 50 mm. Statistical differences were considered significant at *p
The Morris water maze is widely used for the measurement of visually relevant spatial and working memory in rodents. Metformin pretreated rats did not show significant cognitive impairment as they spent longer time in the target quadrant and had a shorter escape latency compared to rats in the surgical group. In in vitro experiments, metformin decreased microglial activation and oxidative stress and promoted autophagy in microglia by activating the AMPK/SIRT1 signaling pathway. Our findings suggest that metformin may reduce surgically induced cognitive impairment in old rats, presumably by enhancing autophagy in the cerebral cortex and reducing microglial activation.
After surgical anesthesia, the body experiences a stress response and neuroinflammation. Numerous studies have shown that inflammatory factors and various neurotransmitters released in the central nervous system directly affect the function of hippocampal neurons or indirectly affect the function of neurons by regulating signal transmission pathways among neurons [33, 34, 35]. Inhaled isoflurane can activate proinflammatory factors in the hippocampus and result in neuronal damage [36]. To avoid the effects of anesthetics, we did not use inhaled anesthetics in our experiment. Leptin, which is produced by adipocytes in the body, may be crucial for preserving the structure and function of the central nervous system. According to several studies, postoperative cognitive impairment is associated with leptin-related pathways [37]. Postoperative cognitive impairment is frequently caused by metabolic syndrome [38]. In a mouse model of POCD, it was discovered that surgical trauma causes metabolism to switch from oxidative phosphorylation to glycolysis in the hippocampus [39]. Therefore, postoperative cognitive impairment is caused by a variety of factors, and neuroinflammation and metabolic disorders are believed to play major roles in its occurrence.
Metformin, as a classical drug used for the treatment of T2DM, has been found to
ameliorate many neurodegenerative diseases and cerebral ischemia. Asadbegi M
et al. [40] found that metformin can protect cognitive function in rats
fed a high-fat diet by inhibiting the accumulation of
Multiple studies have suggested that metformin enters the brain through the BBB.
In the central nervous system, metformin plays many roles, including regulating
neuron and glial cell proliferation, differentiation and migration in the
hippocampus; reducing neuroinflammation and the antioxidant stress response; and
improving insulin sensitivity in the brain [43, 44, 45, 46, 47, 48]. AMPK is a key target of
metformin and is tightly associated with mitochondrial function and energy
metabolism [49]. Metformin can inhibit mitochondrial respiratory chain complex 1
through the AMPK pathway to reduce ROS levels and increase the expression of
antioxidant molecules [50, 51, 52]. These positive effects of metformin may represent
the mechanism by which it improves cognition. Moreover, metformin
dose-dependently inhibits interleukin-6 and interleukin-1
Autophagy is implicated in numerous complex neurodegenerative diseases and neurodevelopmental disorders, which suggests that autophagy is important for the development and maintenance of a healthy central nervous system [56]. There are many different signaling pathways that control autophagy. Autophagy is largely regulated by the AMPK/mTOR pathway, which is disrupted by neurodegenerative diseases and is inhibited with age [57]. Cellular growth, activation, survival, and death are all downstream processes that are influenced by AMPK/SIRT1 signaling, which is an upstream pathway that controls nutrient and redox status [58]. The AMPK/SIRT1 pathway and autophagy are closely related processes [59]. Chuan discovered that metformin inhibits the endoplasmic reticulum stress response and activates AMPK-mediated autophagy to protect against brain damage following cardiac resuscitation [60]. Zhu et al. [61] also found that in a rat model of cardiac arrest and resuscitation, metformin improves neurologic outcomes via AMPK-mediated autophagy activation. Similar results were obtained in our investigation, which demonstrated that after metformin pretreatment, AMPK-mediated autophagy was increased in LPS-stimulated microglia and the cortices of POCD model rats. Additionally, pretreatment with metformin improved the cognitive function of POCD model rats. Our in vivo experiments showed that autophagy level in cerebral cortex was significantly reduced and microglia were significantly activated in rats with POCD. When pretreated with metformin, autophagy level in the cortex of rats increased, while microglia activation decreased. Therefore, we speculate that metformin may play a role in improving POCD by increasing the autophagy level of microglia in cortex and reducing its activation. Further in vitro experiments clearly confirmed that metformin can increase the autophagy level of microglia and reduce its activation.
Metformin pretreatment can reduce postoperative cognitive impairment in elderly rats by increasing autophagy in cortical microglia and inhibiting oxidative stress via the AMPK/SIRT1 pathway.
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
JZ and QZ designed the research study. PJ, JC and YG performed the research. PJ and YG conducted the immunofluorescence staining experiments. PJ, HL and YG analyzed the data. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All protocols were performed according to the National Institute of Health Guide for the Care and Use of Laboratory Animals. Our research was approved by the Animal Ethics Committee of Huadong Hospital affiliated to Fudan University in Shanghai. The approval number is 2022110029S.
We would like to express our gratitude to all those who helped us during the writing of this manuscript. Thanks to all the peer reviewers for their opinions and suggestions.
This research was supported by the Medical Innovation Research Project of Shanghai Science and Technology Commission (21Y11904600) and Young Scientific Research Project of Anesthesia and Pain Special Committee of Shanghai Society of Integrated Traditional Chinese and Western Medicine (2021-469).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.