







1 Jilin Ginseng Academy, Changchun University of Chinese Medicine, 130117 Changchun, Jilin, China
2 Department of General Surgery, Qian Wei Hospital of Jilin Province, 130117 Changchun, Jilin, China
† These authors contributed equally.
Abstract
We investigated the anti-aging effects of velvet antler polypeptide on
D-galactose (D-gal)-induced aging mice. D-gal-induced aging mice were established
and randomly divided into five groups, the control, model, vitamin E (VE), velvet
antler polypeptide low-dose and velvet antler polypeptide high-dose groups. The
Morris water maze test was used to evaluate the learning and memory abilities of
aging mice. Hippocampal neurons were observed via hematoxylin-eosin staining and
transmission electron microscopy. Biochemical methods were used to detect the
activities of superoxide dismutase, malonaldehyde and other enzymes and evaluate
the influence of velvet antler polypeptide on the antioxidant capacity of aging
mice. Using 16S rRNA gene sequencing and meristem technology, we assessed the
effect of velvet antler polypeptide on aging mice’s intestinal flora and fatty
acid metabolism. The experimental results showed that velvet antler polypeptide
could significantly improve aging mice’s learning and cognitive abilities,
increase the activities of superoxide dismutase, glutathione peroxidase, and
catalase in the serum decrease the malonaldehyde content. Intestinal
microecological analysis showed that velvet antler polypeptide could
significantly increase the beneficial bacterial genus Lactobacillus
abundance. Western blot analysis further demonstrated that velvet antler
polypeptide could promote fatty acid metabolism by activating peroxisome
proliferator-activated receptor
Keywords
- Velvet antler polypeptide
- D-galactose
- Aging
- Intestinal microecology
- Learning
- Cognitive impairment
- PPARα/APOE4 pathway
- Microbiota
Velvet antler polypeptide (VAP) accounts for 50–60% of the wet weight of velvet antler and is one of its primary active ingredients [1]. VAP is rich in proteins, amino acids, phospholipids, cholesterol, and calcium ions and exerts various health-promoting effects, with antioxidant [2], osteoporosis resistance [3], and immune regulation [4] effects having been reported. To better understand the role of VAP in the prevention and treatment of neurodegenerative diseases, previous research by our research group also confirmed that VAP could reduce the levels of the glucocorticoid receptor, mineralocorticoid receptor and corticotropin-releasing hormone. The effects of VAP on the thalamus-pituitary-adrenal axis and its ability to inhibit neuronal apoptosis have also been observed [5].
Brain aging is a natural physiological process characterized by oxidative stress (OS), the accumulation of oxidative damage molecules, and changes in the structure and function of neurons, ultimately leading to cognitive decline and memory loss [6]. Among these factors, OS is considered the primary cause of aging-associated neurodegenerative diseases [7]. When the body is subjected to harmful stimuli, there is an imbalance between the generation and removal of free radicals, resulting in an increase in free radicals to a level that exceeds the antioxidant capacity in the body. Notably, excessive free radicals are cytotoxic, causing direct damage to the central nervous system [8, 9]. The human brain accounts for only 2–3% of the human body weight. Still, the central nervous system (CNS) accounts for 20% of the total oxygen supply of the human body. Therefore, the content of neuronal free radicals in the CNS is considerable, presenting a high risk of OS [10].
The human gut microbiota, which comprises more than 100 species, comprises approximately 10-fold more bacteria than human cells [11]. Firmicutes is the most abundant species within the human gut microbiota, accounting for 60% of total bacteria, followed by Bacteroidetes (15%) and other components that account for a small proportion. The gut microbiota is in a state of dynamic balance with the human body, and intestinal microbes have an important role in human metabolism, endocrine function, and immune regulation. Therefore, when the gut microbiota structure is disturbed, neurodegenerative diseases, cardiovascular diseases, and obesity may occur [12]. Changes in the quantity and diversity of intestinal microorganisms can also affect the enteric and central nervous systems. These interactions are known as the microbiota-gut-brain axis, which is important for body function [13, 14]. Although changes in gut microbiota profiles do not necessarily affect health, the loss of diversity in the core microbiota is associated with decreased cognitive function and memory [15]. Indeed, with increasing age, the changes in bacterial flora are primarily characterized by decreases in diversity, probiotics, facultative anaerobes, and total short-chain fatty acids [16].
Although numerous studies have confirmed that VAP-associated antioxidant and anti-aging effects attenuate aging, the exact mechanism underlying this activity has remained unclear. Therefore, in the present work, the anti-aging effect of VAP was evaluated in vivo in mice via the Morris water maze (MWM) test, biochemical analysis, hematoxylin-eosin (H&E) staining, and transmission electron microscopy (TEM). Furthermore, we evaluated the changes in the intestinal microecology in aging mouse models treated with VAP using 16S ribosomal RNA (rRNA) gene sequencing technology. Our goal was to elucidate the mechanism of VAP activity in the context of aging and provide a theoretical basis for the development of new anti-aging therapeutic approaches.
VAP was provided by the Molecular Pharmacology Laboratory, Changchun University
of Chinese Medicine. D-galactose (D-gal) and VE (purity
The Morris water maze (MWM) video tracking and analysis system were purchased from Chengdu Taimeng Technology Co., Ltd. (Chengdu, China). A transmission electron microscope was purchased from Tecnai Corporation (Hillsboro, OR, USA). The high-throughput sequencing platform (MiSeq PE300) was provided by Illumina (San Diego, CA, USA).
Sixty healthy ICR mice (SPF grade, half male and half female, weighing 22–26 g)
were purchased from Changchun Yisi Laboratory Animal Technology Co., Ltd.
(certificate No. SCXK(Ji) -2018-0007). The mice were raised in separate cages
under 12 h light/dark cycles and a standard temperature (22
The MWM test was performed 1 h after D-gal administration on day 57. Briefly, a
platform was placed in the middle of the water maze. Water was added to the
platform until it exceeded 1 cm and was maintained at approximately 25
The whole brains of mice were fixed in 4% paraformaldehyde solution and embedded in paraffin. After 24 h, the brain tissues were sectioned, deparaffinized, rehydrated and stained with H&E. The sections were then observed under an optical microscope to record the pathological changes.
After perfusion, the hippocampal tissues were severed, fixed with 2% paraformaldehyde and 2.5% glutaraldehyde phosphate buffer solution, dehydrated using an alcohol gradient, cleaned with ethoxypropane, embedded, sliced, and stained with uranyl acetate and lead citrate. The ultrastructure of hippocampal neurons was observed under a transmission electron microscope.
Mouse blood samples were collected and centrifuged at 3000 rpm for 15 min at 4
Total DNA was extracted from fecal samples and used as a template to PCR amplify the V3–V4 region of the bacterial 16S rRNA gene. The raw dataset is deposited in NCBI BioProject PRJNA731623 under the BioSample accession numbers SAMN19291644 to SAMN19291668. The PCR products were quantitatively analyzed, and 16S rRNA gene sequencing was performed using an Illumina MiSeq platform (Illumina, San Diego, USA). The raw data is based on the minimum overlap length of 10 bp, with a maximum allowable mismatch ratio in the overlap area of 0.2 (default). The reads of each sample were spliced, and tags with lengths less than 75% of the tag length after quality control were filtered. The chimeras (that is, if the two parents have a sequence that is more than 80% similar to the query sequence, then the query is judged to be a chimera) were removed to obtain high-quality tag sequences. Usearch software (Sonoma, USA) [17] was used to cluster tags at a 97% similarity level, classify operational taxonomic units (OTUs), obtain species classification based on OTU sequence composition, and plot sample dilution [18] and Shannon index curves. PICRUSt (Halifax, CAN) [19] was used to determine the functional gene composition of each sample via comparison of the species composition information according to 16S rRNA gene sequencing results. In this way, functional differences between different samples or groups could be determined. The abundances, pathways, EC information, and OTU abundances in the context of each functional category were obtained using the Kyoto Encyclopedia of Genes and Genomes [20] database.
Total protein was extracted from brain tissue, and the Coomassie brilliant blue
method was used to assess the protein concentration. Then, after SDS-PAGE
electrophoresis, the proteins have transferred to the membrane, which was then
incubated with primary antibodies against target proteins apolipoprotein E
(APOE4), acyl-CoA oxidase 1 (ACOX1), peroxisome proliferator-activated receptor
Mouse brain tissues were collected, and an appropriate amount of normal saline
was added. Then, the samples were centrifuged at 4
Mouse brain tissues were collected and lysed at a brain tissue: lysis buffer
ratio of 1:5. Then, the samples were centrifuged at 12,000
SPSS 19.0 (Chicago, IL, USA) was used for data analysis. The results are
expressed as the mean
The MWM test relies on the survival instinct of animals to find the escape
platform and involves searching for a memory-related spatial position. Fig. 1
shows representative tracks of mice administered the MWM test on the last day.
The mouse behavior trajectory diagram of the model group was more complicated and
irregular, and the mice did not find the platform. Of note, the behavior
trajectory diagram of the mice in the VAP1 and VAP2 groups was simple, and a
platform was found. Compared with the Control group, the percentage of retention
time in the middle ring and the percentage of the distance moved by the middle
ring in the Model group were significantly reduced (P
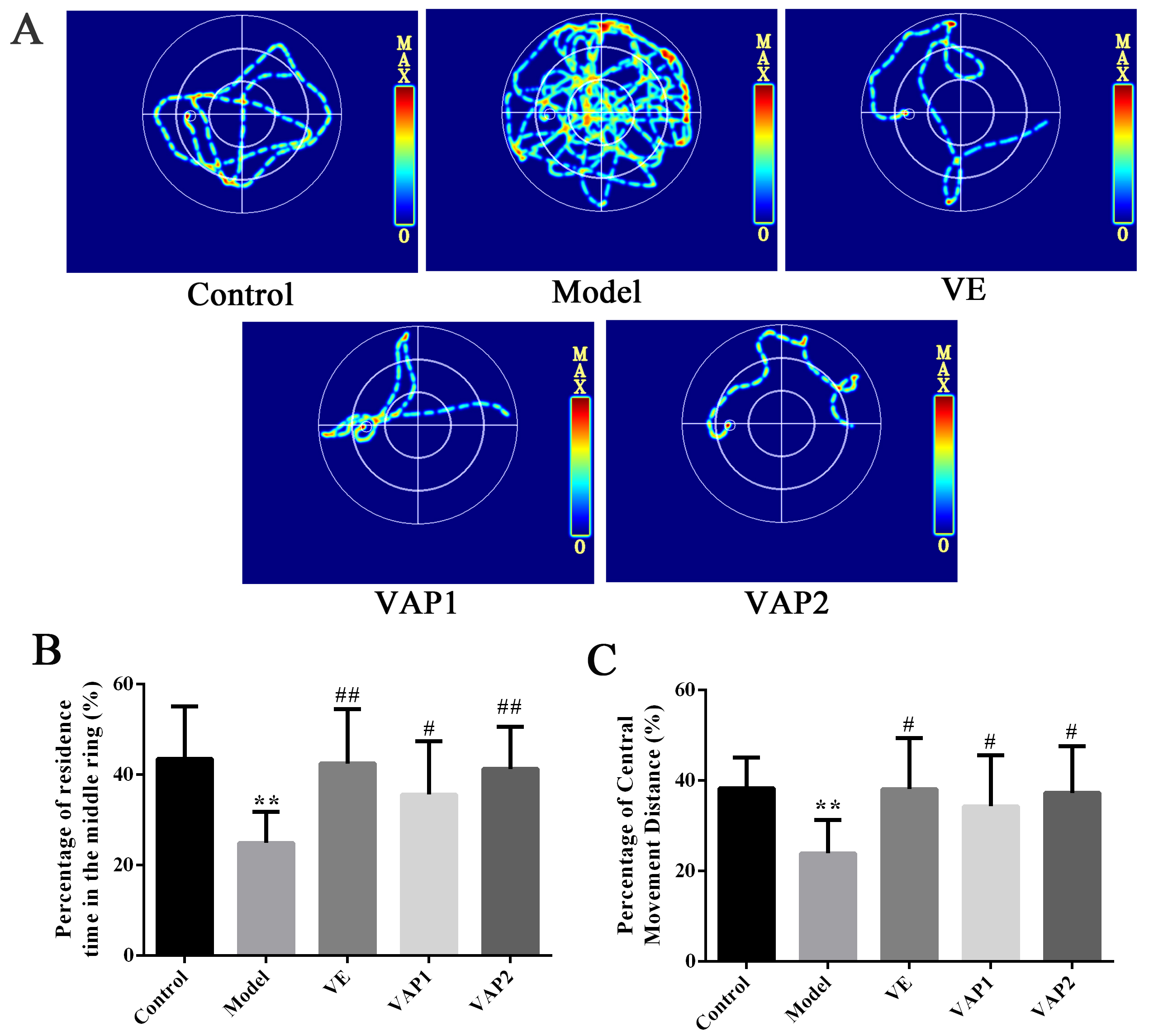 Fig. 1.
Fig. 1. MWM testing to evaluate the influence of VAP on the neurological
function and behaviors of aging mice (n = 8). (A) Behavioral route of mice in
each group. (B,C) Percentage of residence time in the Middle ring and Percentage
of Central Movement Distance. Significant differences are denoted as follows:
compared with the control group, *P
H&E staining was used to observe the pathological morphology of brain tissues in aging mice to explore whether VAP could improve D-gal-induced neuronal pathological damage (Fig. 2A). The CA1 cells in the hippocampi from the control group were abundant, with normal cell structure, complete morphology, and obvious nuclei. In contrast, the number of CA1 cells in hippocampi from the model group decreased and showed irregular morphology, with no obvious nucleoli observed under the microscope. The neurons in the CA1 area of hippocampi were relatively normal, and nuclear shrinkage was reduced in the VE group compared to the model group. Remarkably, the CA1 area in hippocampi from the VAP1 and VAP2 groups was significantly different from that observed in the model group and similar to that of the control group. The structure was relatively normal, the cells were closely arranged and regular, and the morphology was complete.
TEM was further used to observe the ultrastructure of hippocampal neurons in aging mice and to understand whether the reversal effect of VAP on the decline in learning and memory was associated with regulating neuron microstructure in the brain (Fig. 2B). The hippocampal neurons showed an intact morphology, uniform chromatin distribution, and abundant organelles in the cytoplasm in the control group. At high magnification, the mitochondria were elliptical, with a clear double outer membrane and intact cristae. In contrast, the neurons in the hippocampal region of the model group were abnormal; early apoptotic changes such as chromatin condensation were observed; lipofuscin corpuscles were more frequent; the synaptic structure was unclear; lipid droplets were scattered, and vesicle-like expansion of the Golgi complex and reduced rough endoplasmic reticulum were detected. Remarkably, compared to the model group, the VAP1 and VAP2 groups had hippocampal neurons that tended to be normal in structure and morphology, with smooth nuclear membranes, normal nucleoli, and abundant organelles. Notably, more synapses were observed, the synaptic vesicles in the anterior membrane did not have abnormal aggregation, the morphology of the intracytoplasmic Golgi body and mitochondria was normal, and lipofuscin was rare.
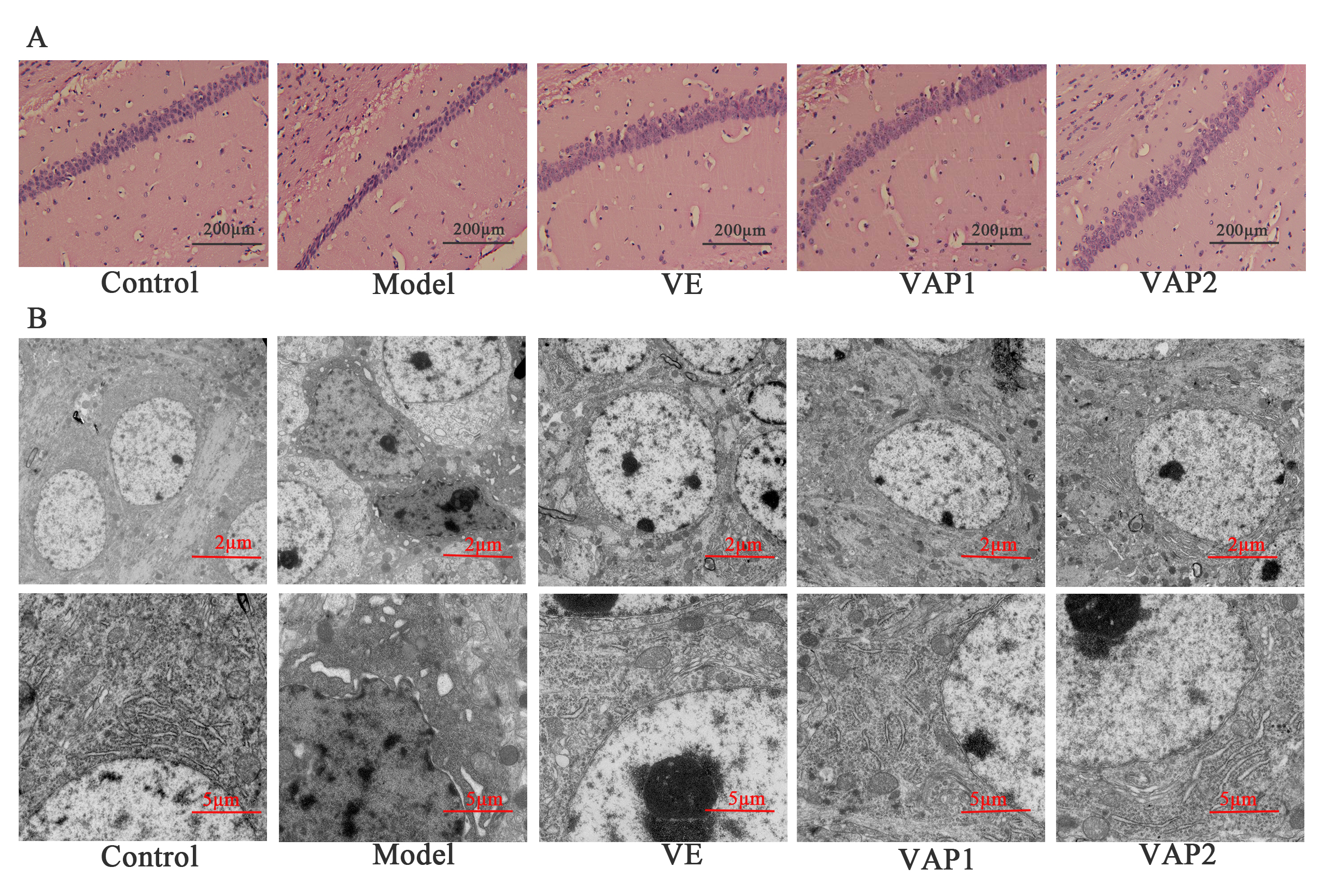 Fig. 2.
Fig. 2.Effects of VAP on the morphology of neurons in the hippocampal
CA1 region in aging mice (n = 3). (A) H&E staining of the hippocampal sections
of mice from each group. Scale bars = 200
A negative effect of OS is the production of free radicals in the body, one of
the primary causes of aging and disease (Fig. 3). Compared to the control group,
the activities of SOD, CAT, and GSH-Px in the serum of D-gal-administered model
mice were significantly decreased (P
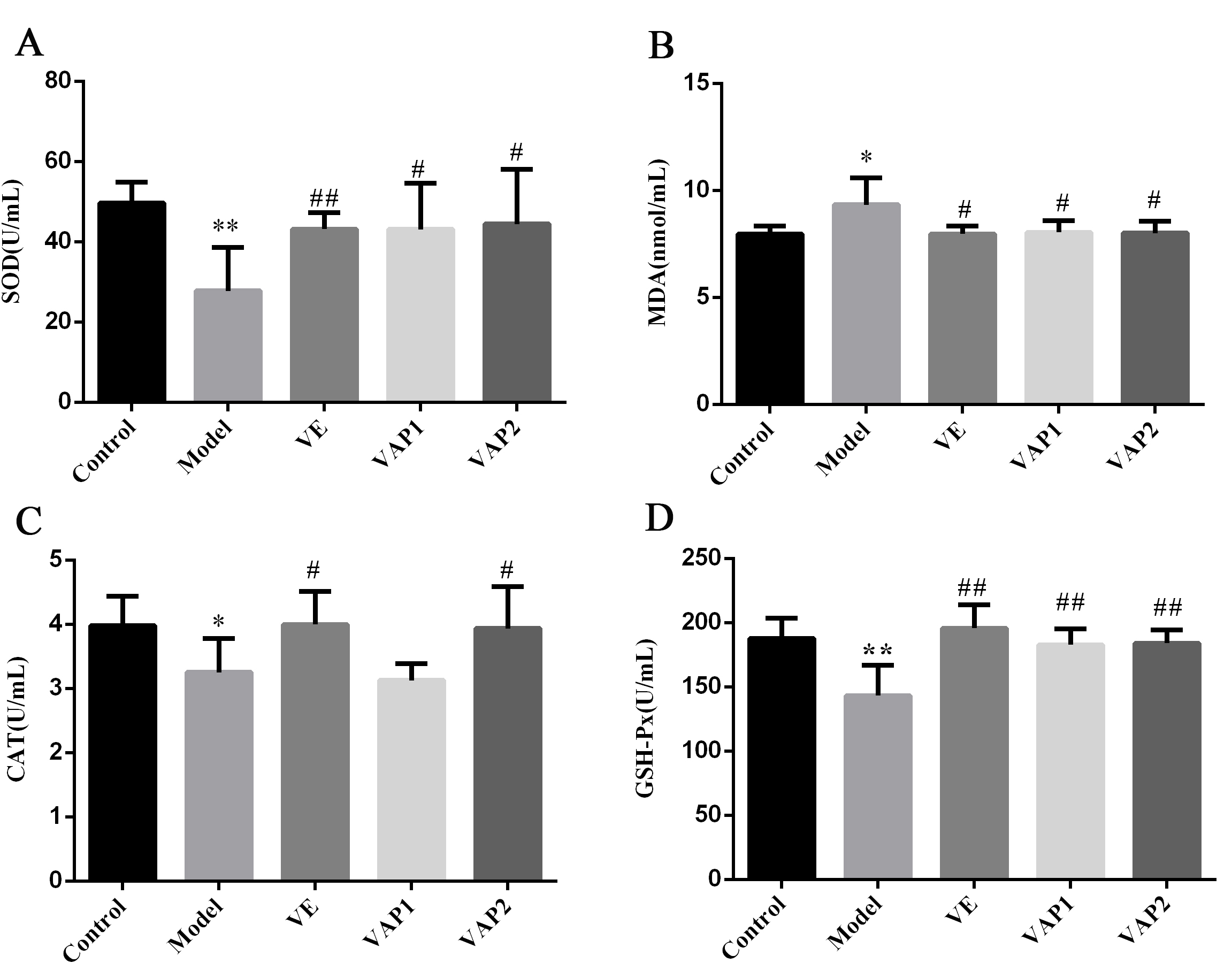 Fig. 3.
Fig. 3.Effects of VAP on the levels of SOD (A), MDA (B), CAT (C), and
GSH-Px (D) in the serum of aging mice (n = 8). Significant differences are
denoted as follows: compared to the control group, *P
The 16S rRNA gene sequencing results yielded 1,999,460 PE Reads, and after filtering and removing chimeras, 1,776,708 effective tags were obtained. The average sequence lengths of the samples in the control, model, VE, VAP1, and VAP2 groups were 417.2, 416.2, 418.2, 421.0, and 420.4 bp, respectively. The ratio of reads with a Phred quality score over 30 (Q30) ranged from 94.15 to 95.71%.
Subsequently, we determined the gut microbiota composition using the 16S rRNA gene reads. The tags obtained by high-throughput sequencing were clustered at a similarity level of 97%, and a Venn diagram was generated (Fig. 4A). Each group had 490 OTUs, and there were essentially no specific OTUs per group. To verify whether the sequencing depth met species identification requirements, we plotted Shannon and rarefaction curves for all samples (Fig. 4B,C). Both curves in all samples were close to the saturation plateau, revealing a high coverage (99%). These data suggest that the vast majority of the microbial diversity information in the samples was extracted and that the bacterial community structure and diversity could be accurately determined.
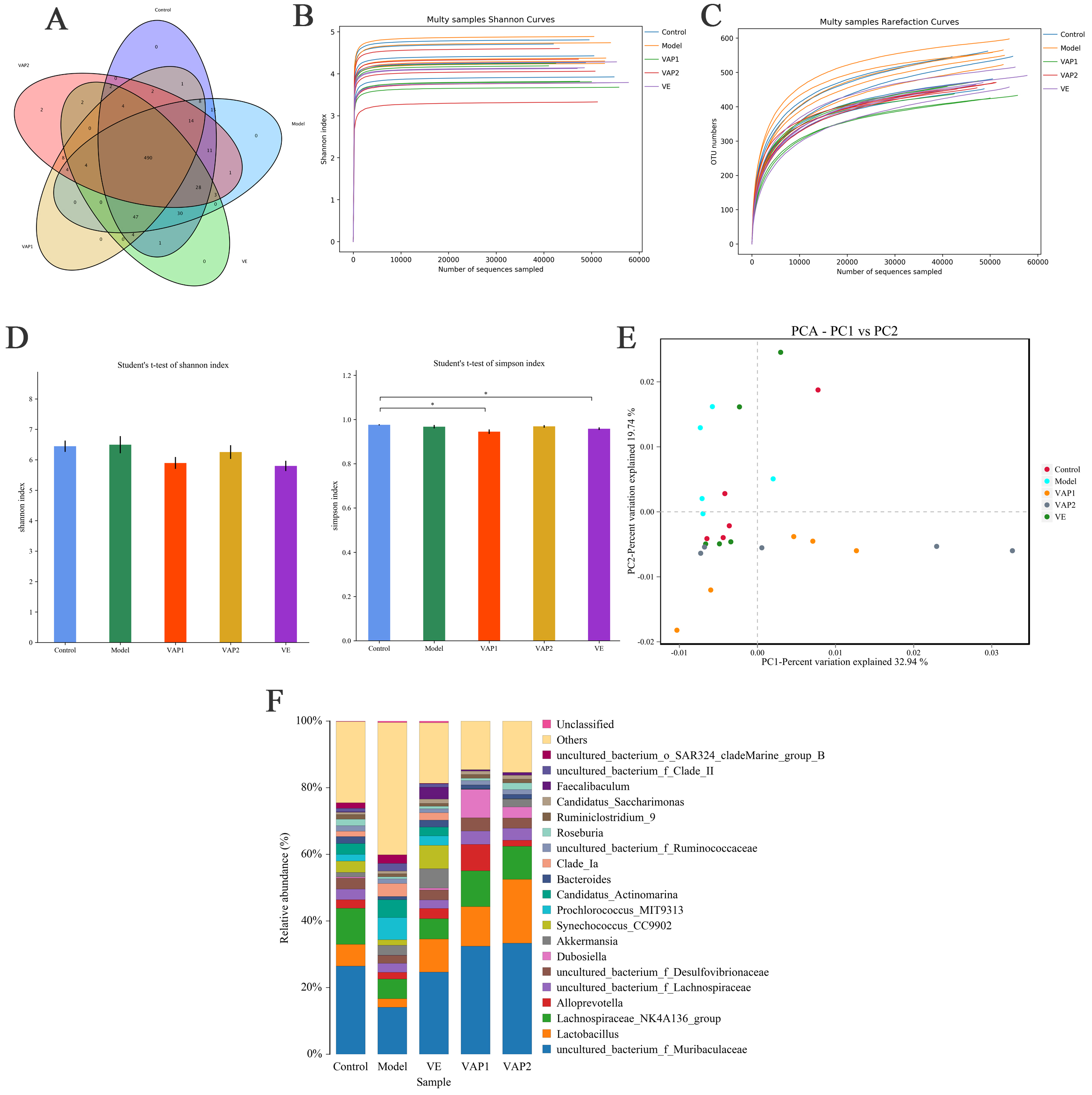 Fig. 4.
Fig. 4.Effects of VAP on the overall structure of the intestinal flora
in aging mice (n = 5). (A) Venn diagram of OTUs. Each OTU represents a microbe.
(B) Shannon curves. Each curve represents a sample, and each group is marked with
a different color. (C) Rarefaction curves. The graphs show the rate of emergence
of new OTUs under continuous sampling conditions. (D) Bacterial
Alpha diversity reflects species abundance (richness) and species diversity. Shannon and Simpson indexes are used to measure species diversity and are affected by the abundance of species in the sample community and the evenness of species (community evenness). The larger the Shannon index value, the smaller the Simpson index value, indicating that the species diversity of the sample is higher [21]. The results presented in Fig. 4D show no significant differences in the Shannon index values of the mouse flora between the groups.
Compared with the control group, the Simpson index value of the model group has no significant difference. No significant difference was observed between the model group and each treatment group. These results show no significant difference in the diversity of the intestinal flora of mice between the groups.
PCA was performed on the mouse intestinal flora (Fig. 4E), where PC1 and PC2 are the two characteristic values that cause the greatest difference between samples. The results showed that 32.94 and 19.74% of the degree of influence was accounted for by PC1 and PC2, respectively. The farther the points are, the greater the difference. The five groups of samples clustered into three categories, with the model group samples distributed in the upper left corner of the figure, far away from the other groups. The control, VE and VAP2 groups showed more overlaps, and the three groups of samples were close together. The upper middle of Fig. 4E shows that after VAP intervention, the similarity of the intestinal flora of the VAP2 group of mice to that of the control mice was improved.
The differences in the abundances of bacteria in the control model, VE, VAP1, and VAP2 groups were evaluated for the top 20 genera (Fig. 4F). The abundances of Lachnospiraceae_NK4A136_group and Lactobacillus were different between the control and model animals, where the abundance of Lactobacillus in the model group was significantly decreased. In contrast, the abundances of the remaining genera were significantly increased. Notably, Lactobacillus members are probiotics with anti-aging and anti-inflammatory effects [22, 23]. Therefore, these results were not surprising. Remarkably, the abundance of Lactobacillus was significantly increased in the VAP1 and VAP2 groups compared to the model group, suggesting that VAP may increase the content of beneficial bacteria in the gut, preventing aging.
Using KEGG pathway analysis, differences in the metabolic pathways (functional genes) in microbial communities from the different groups were investigated. Pathways with an average abundance greater than 0.1% were selected according to the 16S rRNA gene functional prediction results. Moreover, pathways with a significant impact on the differences between groups were evaluated through random forest analysis (Fig. 5). Through KEGG analysis, the following five pathways with a significant influence on the difference between the groups were identified: ko00310 (lysine degradation), ko00071 (fatty acid degradation), ko00300 (lysine biosynthesis), ko00920 (sulfur metabolism), and ko00280 (valine, leucine, and isoleucine degradation). Notably, the value of ko00071 (fatty acid degradation) ranked second but had a greater impact on each group.
According to the predicted gut microbiota functions, we speculated that the aging cognitive impairment caused by D-gal might be related to abnormal fatty acid metabolism in vivo.
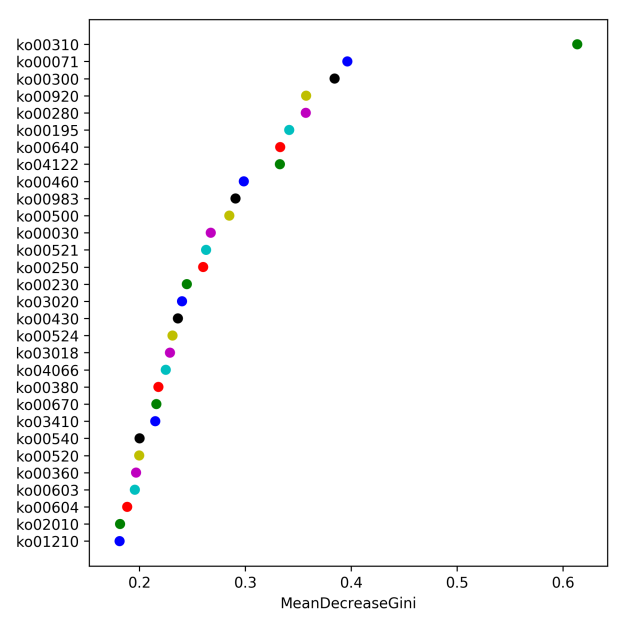 Fig. 5.
Fig. 5.16S rRNA gene functional prediction analysis (n = 5). The greater the mean decrease in the Gini value is, the greater the influence of the pathway on the different groups. Each circle represents a pathway.
According to the 16S rRNA gene functional prediction results, we speculated that
D-gal-induced aging and the consequent cognitive impairment might be associated
with abnormal fatty acid metabolism. Based on this hypothesis, we selected key
enzymes involved in beta-fatty acid oxidation, such as ACOX1 and CPT1A and the
key regulators PPAR
As shown in Fig. 6A,B, the protein expression levels of PPAR
As shown in Fig. 6C, compared to the control group, FFA levels in the model
group were significantly higher (P
The ATP content of mouse brain tissue was analyzed to determine the effect of
VAP on ATP contents. As shown in Fig. 6D, the ATP content in the model group was
significantly lower than that observed in the control group (P
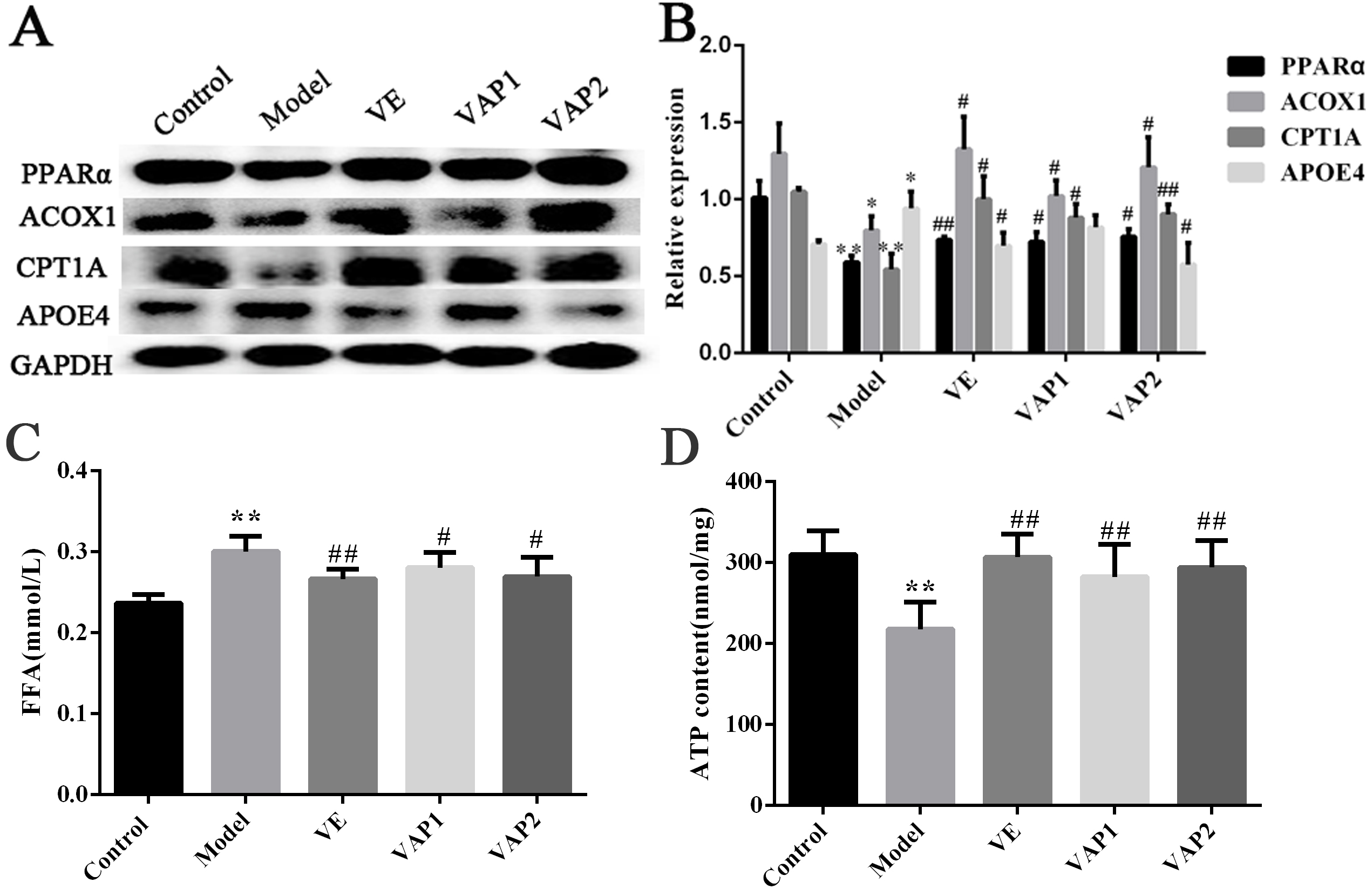 Fig. 6.
Fig. 6.Influence of VAP on fatty acid metabolism. (A,B) The
expression of PPAR
Aging is marked by changes in the physiological functions of the brain. Brain aging is accompanied by learning and memory disorders [23], and the hippocampus plays a central role in the process of memory. The MWM test is one of the classic methods used to detect changes in ethology in learning and memory ability [24]. The behavioral results showed that D-gal-induced learning and memory disorders occurred in mice, with the behavior of mice demonstrating senescence-associated changes. Remarkably, the administration of VAP significantly improved the cognitive ability of aging mice. Moreover, H&E staining and TEM results revealed that the number of neurons in the model group decreased, and their morphology was abnormal. Altogether, these results suggest that VAP can protect the microstructure of neurons and improve cognitive impairment in D-gal-induced aging mice.
Aging is an inevitable stage in the process of life., during which the body
experiences varying degrees of damage, leading to the occurrence of
neurodegenerative diseases and the consequent impairment of learning and
cognitive functions. The excessive accumulation of reactive oxygen species (ROS)
is also closely associated with aging [25, 26]. Normally, ROS are involved in
protein phosphorylation in various networks of the transport system. However,
when ROS levels are excessive, lipid peroxidation occurs, resulting in oxidative
damage [27, 28]. As important antioxidant enzymes in the body, SOD, CAT and
GSH-Px protect cells from damage by scavenging free radicals [29]. SOD can
catalyze the reduction of O
The gut microbiota, also known as the “second genome”, has been increasingly associated with human health [34]. Indeed, with the recent increase in the number of studies on this topic, the important role of the gut microbiota in the body has been widely recognized [35]. We combined behavioral experiments with high-throughput sequencing technology and observed that VAP administration significantly improved aging mice’s learning and memory abilities. Analysis of the alpha diversity of the intestinal flora of mice in each group showed no significant difference in the alpha diversity of the intestinal flora of mice between the model group and each treatment group. PCA results showed that D-gal induced a large deviation in the type of flora in aging mice. The administration of VAP caused an increase in the similarity of the type of flora observed between aging and normal mice. For instance, at the genus level, VAP significantly increased the abundance of beneficial Lactobacillus. It is worth noting that Lactobacillus probiotics were demonstrated to protect cognitive function and improve lipid metabolism.
Based on the predicted functions of 16S rRNA gene functional genes, the KEGG analysis results [36] highlighted the fatty acid degradation pathway. We speculate that the aging caused by D-gal may be related to the abnormal metabolism of fatty acids in the body. Fatty acids are important structural components and energy sources of the body, and their content is affected by decomposition and synthesis rates [37]. As the brain is rich in lipids [38], this organ is particularly sensitive to OS due to its low free radical scavenging ability and weak antioxidant environment. Clinical studies have shown that the accumulation of extremely long-chain saturated fatty acids increases in patients with defects in peroxidase oxidase. At the same time, such patients suffer from impaired brain function and cognitive impairment [39]. Intestinal probiotics such as Lactobacillus help to regulate brain function and behavior [40]. Through neuroendocrine-immune system regulation, directly or indirectly mediates the microbial gut-brain axis, which has an important impact on the physiological function of the host [41, 42]. The gut-brain axis refers to the two-way signal mechanism between the gastrointestinal tract and the central nervous system. Related metabolites of intestinal microbes can effectively improve cognitive function by participating in oxidative stress and fatty acid metabolism [43, 44]. Therefore, VAP may play a role through the brain-gut axis by regulating the composition of the intestinal flora, reducing oxidative stress and fatty acid accumulation, promoting ATP energy supply, and ultimately improving cognitive impairment in aging mice.
The metabolism of fatty acids primarily occurs in the mitochondria and
peroxisomes via
Interestingly, in neurodegenerative diseases, increased SPM content has been
shown to effectively improve the survival rate of neurons and the pathogenesis of
diseases [49]. CPT1A is located in the outer mitochondrial membrane and is also a
key enzyme in the mitochondrial
APOE is the primary component of plasma lipoproteins and the most important carrier of cerebral cholesterol [51], playing a role in regulating lipid metabolism in the central nervous system and maintaining lipid metabolism balance in the brain [52]. Studies have shown that APOE can participate in the growth and repair of neurons, affect dendritic reconstruction and promote synapse generation via the regulation of lipid metabolism and possibly via the regulation of the cytoskeleton (e.g., affecting the phosphorylation of the tau protein) [53, 54]. These biological activities of APOE suggest that it may play an important role in nerve tissue repair [55]. APOE has three common isotypes (APOE2, APOE3, and APOE4), and studies have shown that APOE4 can lead to mitochondrial dysfunction in the brain. Indeed, the cerebral synapses in APOE4 gene carriers are severely damaged [56] and are more likely to suffer from neurodegenerative diseases [57]. Importantly, we showed that VAP could reduce the expression of APOE4 in the brain.
Results conclusively show that VAP promotes the expression of CPT1A and ACOX1 by
activating PPAR
VA, velvet antler; VAP, velvet antler polypeptide; D-gal, D-galactose; H&E,
hematoxylin-eosin; TEM, transmission electron microscopy; SOD, superoxide
dismutase; MDA, malonaldehyde; GSH-Px, glutathione peroxidase; CAT, catalase;
CNS, central nervous system; OS, oxidative stress; ROS, reactive oxygen species;
OTUs, operational taxonomic units; KEGG, Kyoto Encyclopedia of Genes and Genomes;
PPAR
NL and QY conceived and designed the experiments; XRL, ZZ, STM, ZL, YXL, YHZ, QHP and SG performed the experiments; XCL, MK, JNL and JFW analyzed the data; HL contributed materials; XRL wrote the paper.
All animal experiments performed in this study aligned with the relevant guidelines and were approved by the Laboratory Animal Ethics Committee at the Changchun University of Chinese Medicine (20180056).
We thank the anonymous reviewers for their excellent criticism of the article.
This study was supported by the National Key Research and Development Program of China (2018YFC1706603-05).
The authors declare no conflict of interest.