





1 Department of Nuclear Medicine, University Medical Center, Johannes Gutenberg-University, 55101 Mainz, Germany
Abstract
Location and distribution of spinal sympathetic preganglionic neurons projecting to the superior cervical ganglion were investigated in a rodent model organism for photoperiodic regulation, the Djungarian hamster (Phodopus sungorus). Upon unilateral injection of Fluoro-Gold into the superior cervical ganglia, retrograde neuronal tracing demonstrated labeled neurons ipsilateral to the injection site. They were seen in spinal segments C8 to Th5 of which the segments Th1 to Th3 contained about 98% of the labeled cells. Neurons were found in the spinal cord predominantly in the intermediolateral nucleus pars principalis and pars funicularis. At the same time, the central autonomic area and the intercalated region contained only very few labeled cells. In the intermediolateral nucleus, cells often were arranged in clusters, of which several were seen in each spinal segment. Selected sections were exposed to antibodies directed against arginine-vasopressin, neuronal nitric oxide synthase, neuropeptide Y, neurotensin, oxytocin or substance P. It was found that about two-thirds of sympathetic preganglionic neurons produced the gaseous neuroactive substance nitric oxide and that few contained small amounts of neuropeptide Y. Fibers of putative supraspinal origin immunopositive for either arginine-vasopressin, neuronal nitric oxide synthase, neuropeptide Y, neurotensin, oxytocin or, in particular, substance P were found in the vicinity of labeled sympathetic preganglionic neurons. These results demonstrate the location of relay neurons for autonomic control of cranial and cardial structures and provide further knowledge on neurochemical properties of sympathetic preganglionic neurons and related structures.
Keywords
- Arginine-vasopressin
- Djungarian hamster
- Fluoro-Gold
- Neuronal nitric oxide synthase
- Neuropeptide tyrosine
- Neurotensin
- Oxytocin
- Phodopus sungorus
- Sympathetic preganglionic neurons
- Superior cervical ganglion
- Spinal Cord
- Substance P
The Djungarian or Siberian dwarf hamster, Phodopus sungorus (hereinafter referred to as Phodopus), is a widely used model organism for studying seasonal dependency of functional body parameters such as reproduction ingestive behavior and obesity as well as aggression [1, 2, 3, 4, 5, 6, 7, 8]. All these involve the components of the autonomic system, i.e., parasympathetic and intramural neurons and, in particular, the sympathetic system.
Phodopus features robust changes of neural, physiological and behavioral parameters in response to photoperiodic variations. The animals are long-day breeders, exhibit gonadal regression and body weight and fur color changes when exposed to short-day conditions [1]. The species is special since animals lose weight in winter by decreasing body mass on a metabolic basis [9].
There is consensus that the crucial seasonal cue is day-length translated into a hormonal signal, i.e., melatonin secretion from the pineal gland into the blood and cerebrospinal fluid. A clear relation was found between day-length and, among other parameters, gonadal function as a response to the melatonin signal’s daily duration. In animals with short gestation periods such as Phodopus, melatonin acts anti-gonadotropic [2, 10].
The pineal gland is a member of the circadian timing system that consists of several brain structures. The endogenous clock is located in the suprachiasmatic nuclei and exerts its influence on pineal function by multisynaptic pathways [11, 12]. The functionally most important route includes sympathetic regulation of pineal melatonin synthesis via noradrenaline released from the superior cervical ganglia (SCG) neurons, usually denominated as postganglionic neurons. Neurons of the SCG are innervated by fibers originating from perikarya located in the lateral horn of lower cervical and upper thoracic spinal cord segments, i.e., the sympathetic neurons of the intermediolateral nucleus usually denominated as sympathetic preganglionic neurons (SPN). This has been shown previously in several species by many studies using retrograde tracing techniques [13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26], without claiming completeness. See also the reviews [27, 28] and the discussion below for more details.
The SPN provide the connection between the central and the autonomic nervous system. They are the last central component, are under the descending and local influence of the CNS, and innervate postganglionic neurons. Among various other functions, the sympathetic trunk couples the pineal organ to the hypothalamus and is thus a pivotal part of the system that generates and maintains pineal-dependent circadian rhythms in mammals.
As part of a long-standing program, we studied structures and pathways mediating photoperiodic control of body functions in Phodopus, particularly the hypothalamus, retinohypothalamic tract, suprachiasmatic nucleus, superior cervical ganglia and pineal gland [29, 30, 31, 32, 33, 34, 35, 36, 37]. Since Reuss [36] focused on the distribution of calcitonin gene-related peptides in the spinal cord referred to SPN in Phodopus without detailed quantification, the present research was conducted to provide a detailed analysis location and distribution of SPN projecting to the SCG. We also sought to find the possible presence of selected neuroactive substances in or neighboring identified SPN. We, therefore, labeled SPN by retrograde neuronal tracing upon injection of the neuronal tracer, Fluoro-Gold (FG) [38], into the SCG of anesthetized animals and analyzed these neurons in sections from the perfusion-fixed spinal cord. We then used immunofluorescent staining of selected spinal sections containing SPN by antibodies directed to either arginine-vasopressin, neuronal nitric oxide-synthase, neuropeptide Y, neurotensin, oxytocin or substance P.
Fourteen (adult male) Djungarian hamsters (Phodopus sungorus) with body weights of 40–55 g were used. The animals stem from a breeding colony descending basically from hamsters provided by Dr. Klaus Hoffmann (Institute of Reproductive Medicine, University of Münster, Germany). They were reared and held under long-day conditions (light:dark 16:8) with food and water ad libitum in the Department of Anatomy and Cell Biology animal facility. The procedures concerning animals complied with German and European laws to protect animals and were approved by the county-government office (Bezirksregierung Rheinhessen-Pfalz). All efforts were made to minimize animal suffering.
For the tracer application, hamsters were anesthetized with
tribromoethanol (0.3 g/kg b.wt.,
intraperitoneal, Merck, Darmstadt, Germany). All surgical procedures were
performed aseptically. Following exposure of the right SCG under a surgical
microscope, 150 nL of a 5% Fluoro-Gold (FG, see Table 1, Ref. [16, 31, 37, 38, 39, 40, 41])
solution dissolved in distilled water were slowly pressure-injected unilaterally
into the SCG using a glass micropipette (tip diameter
| Substances | Abbreviation | Host | Dilution | Source | References | |
| Neuronal tracer | ||||||
| Fluoro-Gold | FG | 5% | Fluorochrome, Englewood, CO | [16, 38] | ||
| Primary antibodies | ||||||
| Arginine-Vasopressin | AVP | Guinea pig, pc | 1:200 | Peninsula Labs, San Carlos, CA | [37] | |
| Neuronal nitric oxide synthase | nNOS | Rabbit, pc | 1:1500 | Laboserv, Gießen, Germany | [39, 40] | |
| Neuropeptide Y | NPY | Rabbit, pc | 1:500 | Amersham | [31] | |
| Neurotensin | NT | Rabbit, pc | 1:1000 | Amersham | ||
| Oxytocin | OT | Guinea pig, pc | 1:400 | Peninsula Labs, San Carlos, CA | [37] | |
| Substance P | SP | Rat, mc | 1:200 | Boehringer, Ingelheim, Germany | [16, 41] | |
| Secondary antibodies | ||||||
| Cy3-anti-rabbit F(ab) |
donkey | 1:400 | Jackson, West Grove, PA | |||
| Cy2-anti-rat F(ab) |
donkey | 1:400 | Jackson | |||
| Cy2-anti-guinea pig F(ab) |
donkey | 1:100 | Dianova | |||
| The references refer to characterizations given by the producer and/or to
previous use of the antibodies in our laboratory. mc, monoclonal; pc, polyclonal. | ||||||
Following a seven days survival, animals were anesthetized and perfused
transcardially with room-temperature phosphate-buffered 0.9% saline (PBS)
followed by ice-cold periodate-lysine-paraformaldehyde solution [42]. The right
atrium was cut to enable venous outflow. Spinal cords were exposed by laminectomy
and cut into segments, using the midpoint between the entry of two adjacent
ventral roots as rostral and caudal limits. Segments were removed, marked on one
side, postfixed for one hour and stored overnight at 4
For immunohistochemistry, sections containing retrogradely labeled cells were
incubated free-floating in antisera directed against either arginine-vasopressin
(AVP), neuronal nitric oxide synthase (nNOS), neuropeptide Y (NPY), neurotensin
(NT), oxytocin (OT), or substance P (SP) in phosphate-buffered saline to which
0.1% Triton-X and 1% normal swine serum (PBSST) were added. The binding was
visualized using Cy3 or Cy2 coupled to IgG-F(ab)
For each antibody, a total of at least thirty sections per segment stemming from at least six animals were incubated. Further sections were used for antigen distribution and antibody specificity tests. In some cases, FG-fluorescent labeling appeared weaker following free-floating immunohistochemical incubation. Details on antibodies and tracing materials are given in Table 1.
To control for unspecific tracing due to the possible spread of the tracer, three additional animals received FG-application to the respective region without injection into the SCG. In these cases, spinal neurons were not labeled.
Control incubations were carried out by (1) using antibodies preabsorbed with
10–50
Following injection of FG into the SCG and retrograde axonal transport, labeled
neurons were found ipsilateral at the lateral edge of the spinal gray matter,
Rexed-lamina VII, also known as Substantia intermedia lateralis or Zona
intermedia. Cell counting in cross-sections showed that individual preganglionic
neurons were located in segments C8 to Th5. The number of labeled cells per
animal amounted to 486
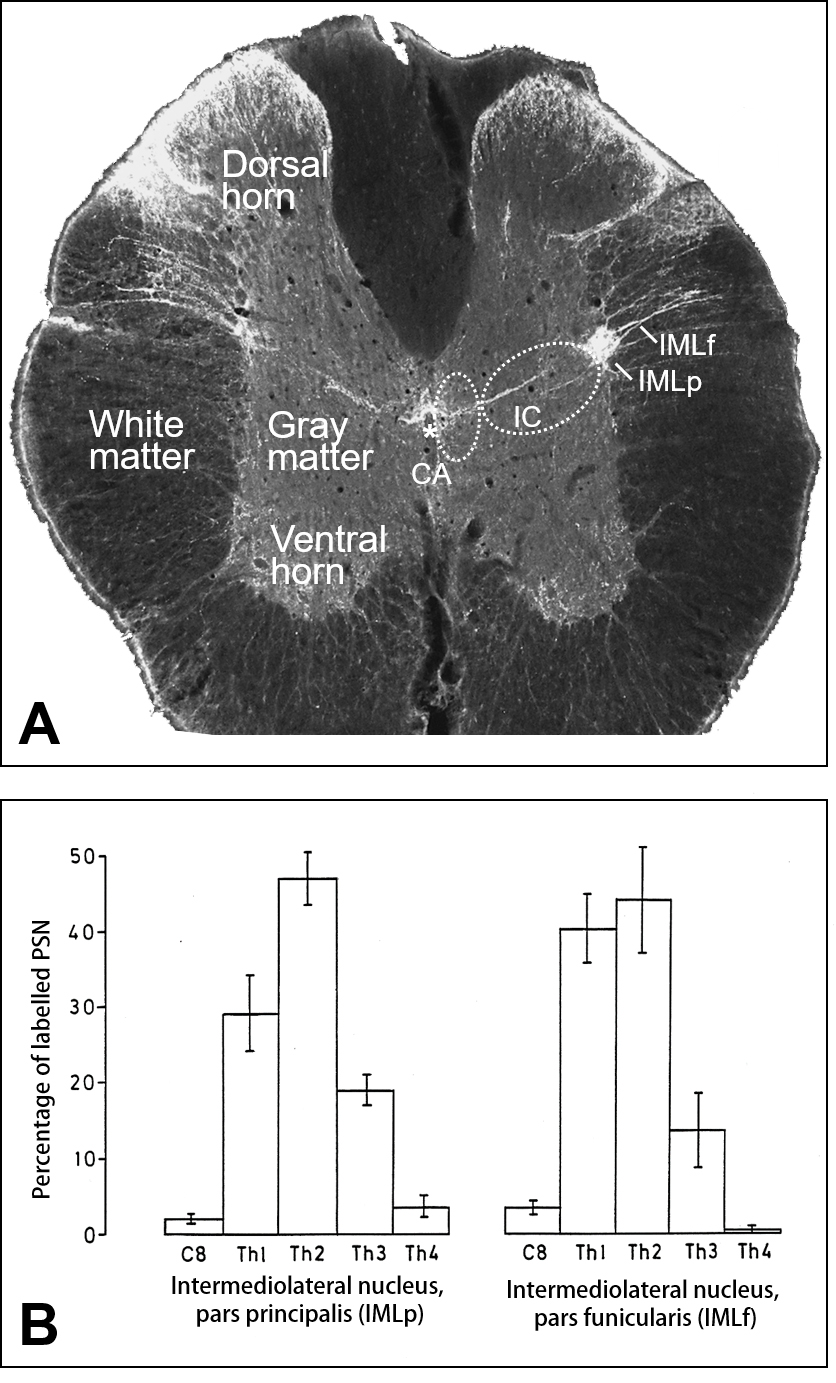 Fig. 1.
Fig. 1.Regional and segmental distribution of sympathetic
preganglionic neurons (SPN) projecting to the superior cervical ganglion (SCG).
(A) Cross-section taken from spinal segment Th2 counterstained for substance
P-immunofluorescence demonstrates the location of the intermediolateral nucleus
pars principalis (IMLp) and pars funicularis (IMLf), the central autonomic area
(CA) surrounding the central canal (*) and the intercalated region (IC). Note
that white matter appears dark in present fluorescence imaging. (B) Segmental distribution of identified sympathetic preganglionic neurons (SPN)
observed upon Fluoro-Gold injection into the superior cervical ganglion and
retrograde neuronal transport. The total numbers of SPN in each the IMLp (left)
and the IMLf (right), respectively, were taken as one-hundred percent (mean (n =
8)
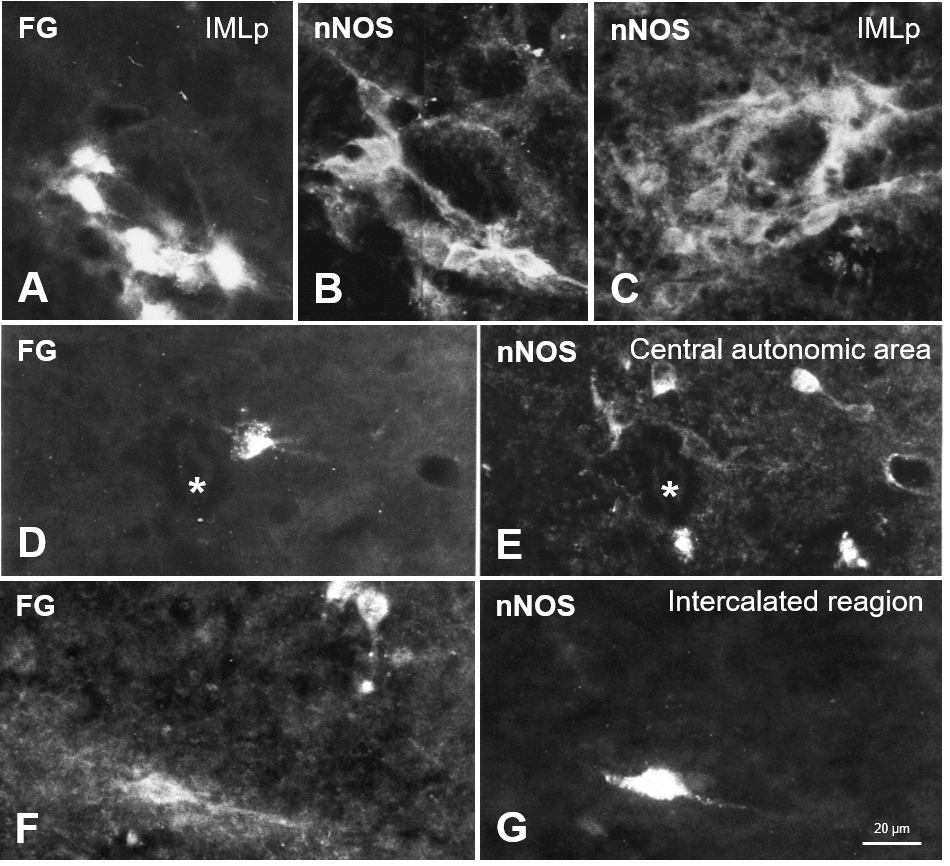 Fig. 2.
Fig. 2.Neuronal nitric oxide synthase (nNOS) in the spinal cord autonomic regions concerning retrogradely labeled neurons. (A) FG-containing sympathetic preganglionic neurons in the Th1 segment and, in the same cross-section, nNOS-immunoreactivity (B). (C) nNOS in the IML (cross-section from Th2). (D,E) In a section taken from Th3, the central autonomic area (CA) exhibited numerous nNOS-perikarya (E), which FG did not label. (D) demonstrates a retrogradely labeled neuron close to the central canal (*), which does not contain nNOS, while other cell bodies shown in the same section in (E) exhibit nNOS but not (F,G). Another example taken from the intercalated region (IC) shows partial overlapping of FG-tracing (F) and nNOS-immunoreactivity (G) in the same section.
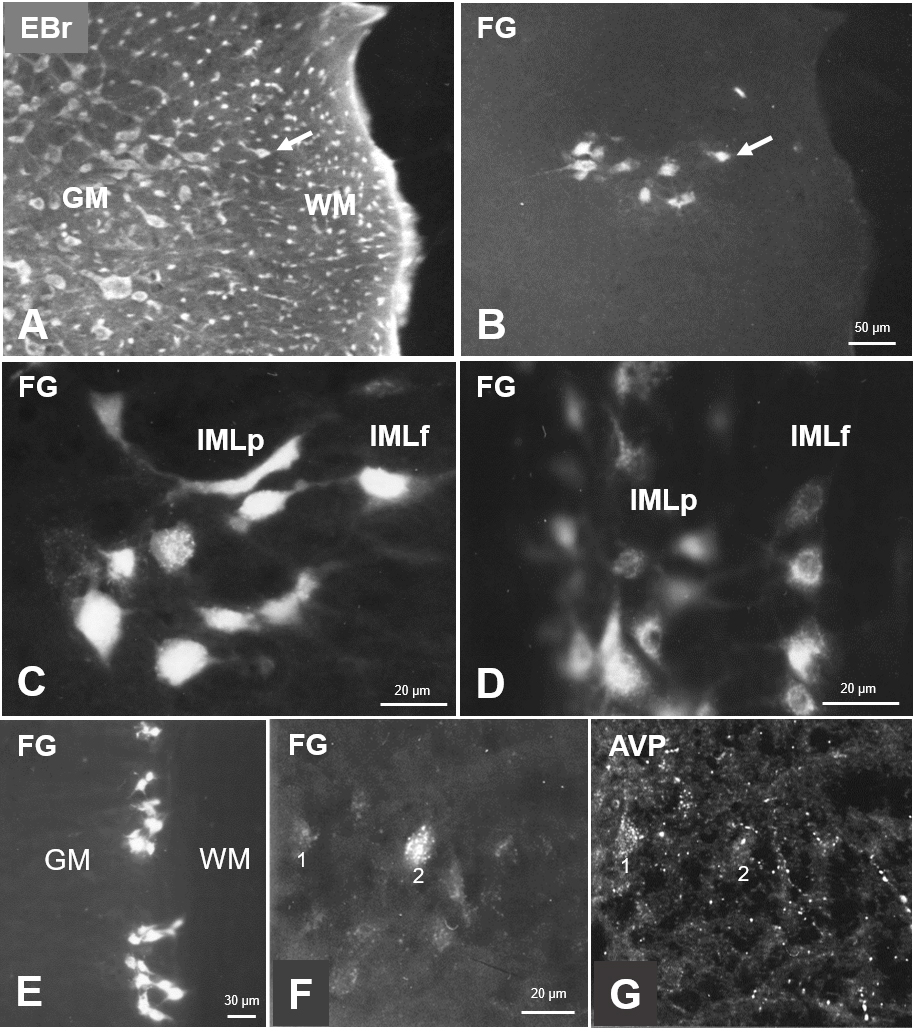 Fig. 3.
Fig. 3.Sympathetic preganglionic neurons projecting to the superior cervical ganglion, as identified by retrograde neuronal tracing with Fluoro-Gold (FG), in sections of the spinal cord. (A) Ethidium-bromide fluorescent staining of a cross-section showing the lateral horn of Th1. (B) FG-labeled cells in the same section. Arrows depict the most laterally localized SPN to ease orientation. (C) Labeled cells in both parts of the IML in a cross-section of the C8 segment. (D) Labeled cells in a horizontal section of upper thoracic segments. (E) FG-labeled cells in IMLp in a Th1 horizontal section demonstrating the location of labeled neurons in clusters. (F) FG-labeled cells in IMLp in a Th1 cross-section, and rich arginine-vasopressin (AVP)-immunoreactivity in the same section. (G) AVP is often found in the vicinity of FG-neurons. Examples are marked “1” and “2”. Note that following free-floating immunohistochemical incubation, FG-labeling may appear weaker. Abbr.: EBr, ethidium bromide; GM, gray matter; WM, white matter; IMLp, intermediolateral nucleus pars principalis; IMLf, intermediolateral nucleus pars funicularis.
The segmental distribution of labeled cells in both parts of the IML is given in
Fig. 1B, where the average total numbers of SPN in each the IMLp (326) and the
IMLf (152), respectively, were taken as one-hundred percent. In both regions, the
segments Th1 to Th3 contained about 95% of the labeled cells. In horizontal
30-
Neuronal NOS was the only substance tested that showed clear-cut immunolabeling
of identified SPN. The incubation of spinal sections in respective antibodies
demonstrated nNOS perikarya of the IML regions (Fig. 2A–C). It was found that
319
A few identified SPN exhibited very weak neuropeptide Y-immunofluorescence (Fig. 4A–D; thick arrow in D) just above the background but different from appearance in sections stained with pre-absorbed antibodies. More intensely stained fibers were seen in SPN regions (Fig. 4B,D). These fibers opposed identified SPN in some cases, but their origin was not readily apparent under the microscope.
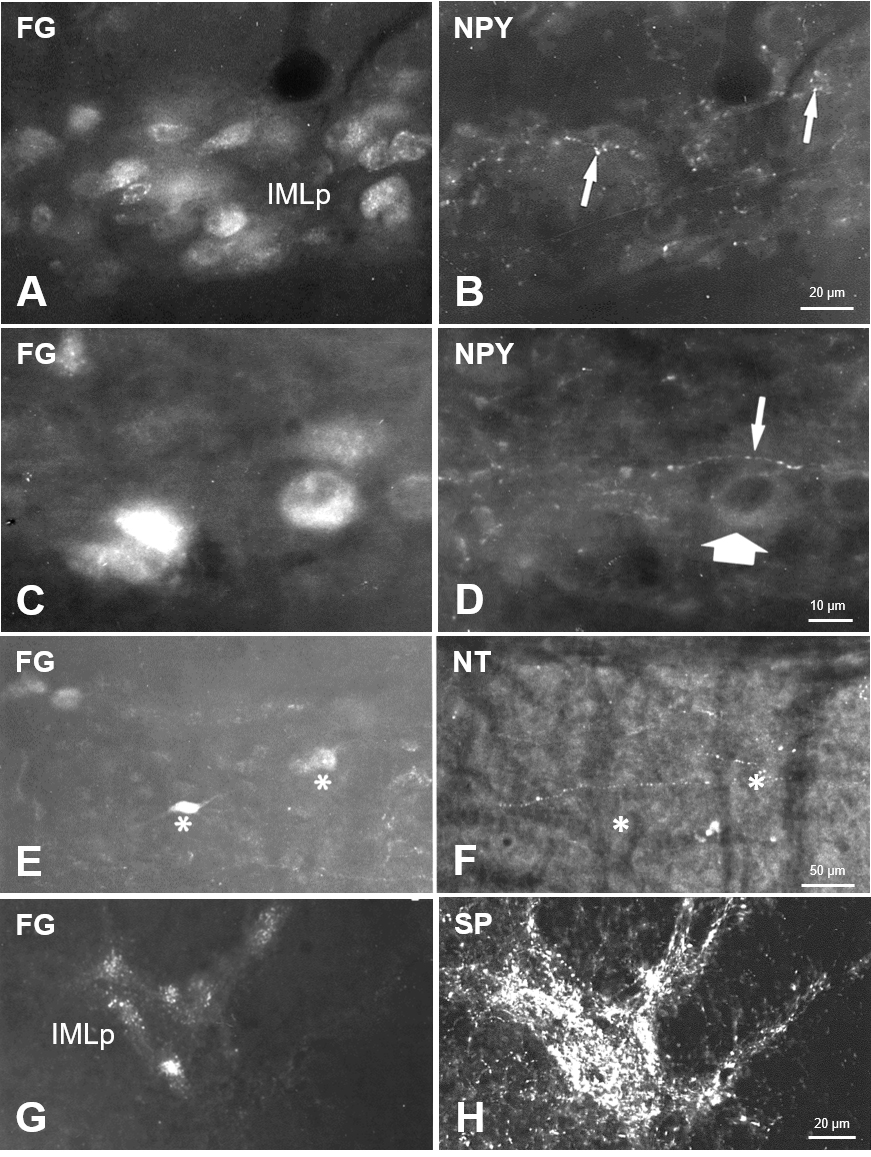 Fig. 4.
Fig. 4.Sympathetic preganglionic neurons and neuropeptide-immunoreactivity in a horizontal section of Th1. Beaded neuropeptide Y (NPY)-fibers (B,D) in the vicinity of retrogradely labeled sympathetic preganglionic neurons in the IML (A,C), which only in some cases exhibited scarce NPY-immunoreactivity (thick arrow in D). Scattered neurotensin (NT)-fibers were seen in the gray matter (F) in a horizontal section, lacking distinct association with FG-labeled SPN (E). Asterisks in E, F mark the location of two FG-neurons in both images taken from the same section. In contrast, SPN in the IML were covered with dense substance P (SP)-immunoreactive structures (G,H) in a Th1 cross-section.
Other neuropeptides tested presently were found only in fibers in the spinal segments with a close spatial relationship to identified SPN. Arginine-vasopressin was present in scattered fine fibers throughout the gray matter with an apparent accumulation at SPN (Fig. 3F,G; “1” and “2” indicated lightly labeled SPN). A very similar distribution was found for oxytocin-fibers in the lateral horn. In contrast, in the dorsal horn and intermediate zone, gray matter slightly more OT was observed than AVP. Thin varicose neurotensin-fibers were present throughout the gray matter, appeared to be concentrated in the lateral horn but did not show appositions to SPN (marked by asterisks in Fig. 4E,F).
The densest fiber staining by far was seen for substance P. The respective immunolabeling was strong in laminae I/II of the dorsal horn. The lateral horn covered the whole region where identified SPN were located (Figs. 1A,4G,H).
The fluorescent appearance of SPN was often weakened following free-floating incubation for immunohistochemistry (see Figs. 3E,4A,G).
The present research sought to characterize the main site of neuroendocrine-autonomic integration in the photoperiodic rodent, Phodopus. It describes the location of spinal SPN innervating the superior cervical ganglion and demonstrates the expression pattern of six selected neuroactive substances in the respective spinal region.
Phodopus is a seasonal breeder. Reproduction and other body parameters are regulated by photoperiod and controlled by mechanisms involving several brain sites, including the hypothalamus and pineal gland [1]. The main relay downstream to the hypothalamus is spinal SPN, where intrinsic, supraspinal and indirect peripheral (sensory) inputs converge and influence the distinct output signals that regulate sympathetic postganglionic neurons. In the context considered here, the rostralmost sympathetic ganglion, the superior cervical ganglion, is of particular interest since its neurons provide sympathetic innervation of heart and head structures such as eye, ear, choroid plexus, carotid body, salivary glands and blood vessels.
Postganglionic neurons also synapse at pinealocytes to regulate melatonin synthesis. This pathway is the route translating day-length into a melatonin secretion signal [44]. The SCG also conveys the autonomic stress response and contributes to related cardiovascular diseases [45]. It is also part of the neuroimmune connection providing the sympathetic innervation of, e.g., nasal-associated lymphoid tissue (NALT) [46]. Notably, there is also evidence for reducing SPN as part of autonomic dysfunction in Parkinson´s and Lewy-body diseases [47, 48].
The exact location of sympathetic preganglionic neurons innervating the superior cervical ganglion is thus eligible for further investigations utilizing experimental manipulations such as a specific lesion, electric stimulation or iontophoretic application of neuroactive substances in this rodent model organism.
Sympathetic preganglionic neurons projecting to the SCG were observed ipsilateral in segments C8–Th5, with the vast majority found in segments Th1–Th3. These SPN are the rostralmost specimens in mammals since their target, the superior cervical ganglion is the most cranial of the sympathetic ganglia. Most of these cells are located in one of the two parts of the IML, i.e., in the spinal nuclei IMLp and IMLf, respectively, with only a few cells occurring in the CA and the IC. Although the exact location and segmental distribution of SPN vary between species, this pattern appears to be a general feature in mammals. The injection of a retrograde neuronal tracer into the SCG, resulted in similar observations in cat, dog, pig, guinea pig, rat and golden hamster in a considerable number of studies, e.g., [13, 14, 15, 16, 17, 18, 19, 20]. While most studies agree on low cell numbers in both CA and IC, the percentage portions of neurons in IMLp/IMLf vary from 88/8 (cat [21]) and 86/9 (pig [20]) to 70/25, 64/30 and 63/33 (rat ([15, 19, 22]), 56/34 golden hamster [17], and 67/31 (Phodopus [present study]). It thus appears that species with large body sizes, such as pig and cat, exhibit most SPN in the IMLp while small species such as rat and hamster rather show a 2/1 ratio. Furthermore, from the SPN-tracing studies in which quantitative data were provided, it is evident that the mean total number of SPN per animal roughly increases with size. Described were 486 in Phodopus, 746 or 1100 in rat [19, 22], 1500 in cat [21], 5000 in pig [20].
Another distinct arrangement pattern of SPN, as apparent in horizontal sections, is their concentration in clusters. This seems to be a common SPN location in mammals since it was observed here and previously in monkeys, pigs, cats, rats and golden hamsters [13, 15, 17, 20, 21, 23, 24]. It is open whether these clusters are functionally heterogeneous and/or synapse in distinct ganglion parts. Targeted injections into restricted parts of the ganglion appeared difficult and were not described yet. In particular, in Phodopus, the structure’s small size leads to the diffusion of even nanoliter-amounts of the tracer throughout the whole ganglion. Although the projection pattern of a single SPN concerning possible convergence and divergence in the ganglion is only poorly understood, there is evidence that SPN are arranged in target-specific columns.
It is agreed that most SPN are cholinergic [20] and that they produce several additional neuroactive substances [27], one of which is the intra- and intercellular messenger nitric oxide (NO). This gaseous molecule is synthesized on demand from a terminal guanidine-nitrogen of L-arginine by neuronal nitric oxide synthase (nNOS). NO then activates its receptor soluble guanylate cyclase, an 160 kDa cytoplasmic protein, and the formation of the second messenger cyclic guanosine monophosphate (cGMP) [49, 50]. In studies of neuronal systems, the NO-system is depicted mostly by immunohistochemical detection of nNOS by specific antibodies. By using this method, three groups of neurons were presently observed with regard to nNOS-IR in Phodopus spinal autonomic regions. It was found that an average of 65% of all identified SPN exhibited distinct nNOS-immunoreactivity, with 51% of SPN in IMLp and 74% of SPN in IMLf. One-third of identified SPN did not exhibit nNOS, and some additional nNOS-positive somata not retrogradely labeled were seen in IML, CA and IC. They may represent interneurons. A more detailed analysis of nNOS-IR SPN was provided previously for rat and golden hamsters [16]. The present findings are in general accordance with data from the literature. Several studies described the presence of NOS in spinal autonomic regions. Still, SPN were in many cases, not identified by retrograde tracing from the SCG, or the ratios of SPN and nNOS-neurons were only qualitatively described. However, in golden hamsters, the SPN in the IMLp were nNOS-positive to 25% and those in the IMLf to 72% [16]. In rats, the IMLp portion accounted for 57 or 72 percent [16, 19], while in IMLf, 86 or 88 percent of SPN were found to express nNOS. In pigs, 72% of identified SPN were nNOS-immunoreactive [20]. Taken together, in the analyzed species, about two-thirds of identified SPN were nitrergic, with a higher proportion in the IMLf than in the IMLp.
Notably, we observed remarkably dense nNOS-positive fiber nets, in addition to the immunoreactive perikarya, in the IML region. They likely represent cell processes of intraspinal autonomic nNOS-immunoreactive neuronal cell bodies. Alternatively, they may originate from supraspinal sites such as the hypothalamic paraventricular nucleus (PVN; see below) that project to SPN regions. The functional role of these intra-/supraspinal structures most probably is to provide NO to induce cGMP formation in SPN target neurons. Cyclic GMP activates kinases and may exert direct effects on ion channels, thus contributing to the integration of incoming signals. Arriving action potentials of hypothalamic origin led to increased intracellular free calcium ions resulting in activation of nNOS and immediate release of NO. Notably, dorsal rhizotomy in rats affected nNOS neuron number in the ipsilateral dorsal horn but not in autonomic regions, suggesting supraspinal regulation of NO production in the IML, where close appositions of nNOS-neurons and SP-fibers were regularly observed [16]. The NO-system has potent (inhibitory) effects during increased sympathetic activity, i.e., predominantly during daytime [51]. There is, e.g., ample evidence that NO augments cholinergic transmission [52]. Thus, diminished expression of nNOS at various central sites is associated with distinct pathophysiological states of altered sympathetic output and, e.g., disturbed smooth muscle relaxation (cf. [53, 54]).
The second substance in Phodopus SPN, although in small amounts, is neuropeptide Y (NPY). This 36-amino acid peptide, also known as neuropeptide tyrosine, is a co-marker of noradrenergic rather than cholinergic neurons. While the respective immunostaining was faint and detected only in a restricted number of nerve cell bodies, it was clearly above background and antibody blocking controls. This low staining level may be interpreted as a low synthesis level. Notably, SCG neurons express NPY receptors [55], but these may also work as autoreceptors since many SCG neurons synthesize NPY, including those that project to the pineal gland [56]. There is thus some evidence for NPY-synthesis by SPN. However, the substance—and neurotensin and substance P—was described previously only below Th3 for cat SPN, which were not identified by retrograde tracing [57]. In addition, the animals received an intrathecal application of colchicine. This microtubule-disrupting substance arrests axonal transport resulting in cytoplasmic accumulation of transmitter vesicles and thus enhanced immunostaining.
However, NPY-staining of fibers close to SPN was stronger than in perikarya. These fibers may originate from brainstem neurons or arise from within the spinal cord since spinal interneurons in proximity to sympathetic preganglionic neurons express NPY-mRNA [58]. Notably, NPY-receptors in spinal regions were detected in the dorsal horn only. Interneurons that built a large portion of IML neurons (see Fig. 3) are regarded as circuits involved in sympathetic control [27].
Aside from nNOS and NPY, further peptides were detected only in fibers near SPN. These will be discussed shortly in the following.
Neurotensin, AVP and OT, and substance P were observed in fibers but not in neuronal cell bodies in the spinal autonomic regions. This is in partial contradiction to earlier reports [57, 59] describing that SPN may co-express peptides such as NT and SP in cats. The comparison to other studies and to the present findings is hampered since these substances were found only in cats. SPN were not identified by retrograde tracing. The neurotoxin colchicine used to block axonal transport is suspected of interfering with transmitter metabolism.
Hence, the main question is where the immunoreactive structures observed may originate, particularly those that may represent intra- or supraspinal inputs to SPN. Spinally projecting neurons were identified in several diencephalic regions, including hypothalamic paraventricular nucleus (PVN), Edinger-Westphal nucleus, nucleus raphe pallidus, medullary lateral tegmental field, nucleus interfascicularis hypoglossi and rostral ventrolateral medulla (RVLM) [60, 61, 62, 63]. As part of the endocrine-autonomic connectome, these may all contribute to the complex SPN control [64]. It is conceivable that they exert regulatory influence on SPN and thus on the SCG and, in turn, the structures innervated by SCG-postganglionic fibers, including the pineal gland, eye, ear, cerebral blood vessels, and heart. Notably, the IML of segments Th1–3 is the main cell group regulating sympathetic cardiac function [65].
Neurotensin (NT) was present in the IML-region in scattered, thin and varicose fibers, while not in distinct amounts in other gray matter regions. From microscopic analysis, there was no indication of clear appositions of NT-fibers and SPN. A supraspinal origin of this neuropeptide in the Phodopus spinal cord is suggested. The respective cell bodies from which the fibers may originate were observed in the hypothalamus, where the highest NT levels were found [66]. As spinal NT-receptors were not reported, and functional aspects of NT on SPN were not described, these fibers may be fibers-of-passage.
The nonapeptides arginine-vasopressin (AVP) and oxytocin (OT) were seen in sparse immunopositive fibers scattered throughout the gray substance. However, they appeared accumulated at identified SPN, predominantly in the IML, suggesting a preferential influence on autonomic neurons. Since these peptides were not found in identified SPN or other neuronal perikarya in the studied segments of the Phodopus spinal cord or other mammals, it may be assumed that these fibers originate in supraspinal regions and are part of the hypothalamo-spinal projections. Nonapeptidergic projections to identified SPN in the IML originating from parvocellular subnuclei of the PVN were shown by retrograde neuronal tracing and combined immunohistochemistry in rats [67]. In addition, iontophoresis combined with electrophysiological recordings demonstrated that AVP and OT influenced a subpopulation of SPN in rats via V1- and OT-receptors, respectively [68]. Both peptides excite SPN in rats [69].
Substance P (SP) in the Phodopus spinal cord stands out with the greatest fiber
density of all substances tested here. The upper laminae of the dorsal horn were
strongly stained, and SP-immunolabeling was prominent in the lateral horn,
covering the SPN location sites (see Fig. 1A). Immunoreactive neuronal cell
bodies were not observed. Notably, dorsal rhizotomy in rats drastically reduced
SP-fibers in the dorsal horn but did not affect SP distribution in autonomic
spinal regions where SP-fibers contact nNOS-neurons [16]. This speaks in favor of
a supraspinal origin of SP-ergic innervation of SPN, which are selectively
excited by SP via neurokinin (NK
In conclusion, the data presented here reveal that sympathetic preganglionic neurons innervating the superior cervical ganglion are located in ipsilateral clusters predominantly in the intermediolateral nucleus and the lateral funiculus of segments Th1 to Th3 in the Phodopus spinal cord, in general accordance to findings in other mammalian species. Many of these neurons produce nitric oxide as an intrinsic neuroactive gaseous substance. Some contain low amounts of neuropeptide Y. Immunohistochemical tests further revealed that other substances such as arginine-vasopressin and oxytocin are present in scarce fibers, mainly in close vicinity to identified SPN, neurotensin in fibers of passage. In contrast, substance P was observed in high density in the area of autonomic neurons. Fiber systems are most probably of supraspinal origin and relay and integrate information from hypothalamic and other sites in SPN en route to sympathetic ganglia.
AVP, arginine-vasopressin; C, cervical segment; CA, central autonomic area; DH, dorsal horn; FG, Fluoro-Gold; IC, intercalated region; IMLf, intermediolateral nucleus pars funicularis; IMLp, intermediolateral nucleus pars principalis; IR, immunoreactivity; nNOS, neuronal nitric oxide synthase; NPY, neuropeptide Y; NT, neurotensin; OT, oxytocin; SCG, superior cervical ganglion; SP, substance P; SPN, sympathetic preganglionic neurons; Th, thoracic segment; VH, ventral horn.
SR designed and performed the experiments, analyzed the data, and wrote the paper.
Animals were reared and held in the animal facility of the Department of Anatomy and Cell Biology, University of Mainz, Germany. All animal experiments were carried out in accordance with the guidelines of the European Community Council Directives 86/609/EEC and approved by the county-government office (Bezirksregierung Rheinhessen-Pfalz; 177-07/961-30).
The author thanks Ursula Disque-Kaiser and Angelika Thomas-Semm for technical assistance. This study was conducted while the author was Head of the Functional Neuroanatomy Group at the Department of Anatomy and Cell Biology, University Medical Center, Mainz. Thanks also to the anonymous reviewers for excellent and helpful criticism of the article.
This study was supported by grants from Deutsche Forschungsgemeinschaft (Re 644) and Naturwissenschaftlich-Medizinisches Forschungszentrum, Mainz.
The author declares no conflict of interest.