



1 Cognitive and Neuroscience Research Center (CNRC), Tehran Medical Sciences, Islamic Azad University, 18116-94784 Tehran, Iran
2 Institute for Cognitive Science Studies (ICSS), 16583-44575 Tehran, Iran
3 Department of Anatomical Sciences & Cognitive Neuroscience, Faculty of Medicine, Tehran Medical Sciences, Islamic Azad University, 19395-1495 Tehran, Iran
Abstract
Sleep deprivation has deteriorating effects on cognitive functions and
activation of brain inflammation mechanisms has been reported by some studies
following total sleep deprivation. Some studies have reported the health benefits
of punicalagin, a main abstract from Punica granatum L., including those for the
treatment of Alzheimer’s disease. The antioxidant characteristic of punicalagin
and the fact that sleep deprivation accelerates mediators of inflammation led us
to further explore the possible neuroprotective role of punicalagin in total
sleep deprivation memory impairment in a rat model. In this study, male Wistar
rats were implanted with a canula in the lateral ventricle to receive
intracerebroventricular injections (drug or vehicle). The animals were trained
for the passive avoidance test and then received intracerebroventricular
injections of different doses of punicalagin (0.001, 0.01, or 0.1
Keywords
- Punicalagin (PG)
- Total sleep deprivation (TSD)
- Memory deficit
- Male rats
Sleep deprivation has profound adverse effects on cognitive functions including information processing speed [1] and learning and memory [2, 3]. Sleep deprivation not only leads to a decline in cognitive functions but also generates higher levels of inflammatory cytokines [4, 5]. Previous studies have indicated that physiological and biochemical morbidity following sleep disorders is related to oxidative stress [6, 7, 8]. Besides, cognitive decline following sleep deprivation has been reported to correlate with increased oxidative stress in the brain and body [9, 10].
The cycle of sleep, as a homeostatic process, is generally divided into two main phases: rapid eye movement (REM) and non-rapid eye movement (NREM) [11]. Theta waves are dominant in REM sleep and the phase is accompanied by low muscle tone. On the other hand, EEG in NREM sleep is composed of waves with high amplitude and low frequency, and the phase is accompanied by decreased muscle activity [12, 13]. Sleep cycle in rats takes about 12-20 minutes [14] and the NREM sleep-related model of REM-sleep proposes that REM sleep is homeostatically linked to NREM sleep rather than to waking and occurs in response to NREM-sleep expression to compensate certain processes of NREM sleep [15, 16]. Similar to the sleep cycles, sleep deprivation is divided into total sleep deprivation (TSD) and rapid eye movement sleep deprivation (RSD). Reports showed that TSD impaired consolidation of declarative memory [17] and motor adaptive task memory [18]. However, selective REM sleep deprivation disrupted spatial memory consolidation in an eight-box task [19] and water maze [20]. TSD and RSD both inhibit the induction of long-term potentiation (LTP) [21, 22]. Considering that REM sleep is not the only phase that is involved in the consolidation of memory [23], in the present study we selected a 24-hour TSD model to investigate its effects on memory deficit.
Punicalagin (PG), as a potent antioxidant, is a hydrolyzable polyphenol in Punica granatum L. and is a major component of pomegranate responsible for its health benefits [24]. PG also has an anti-amyloid beta (1-42) fibril aggregation effect, a potency that can be considered in the treatment of Alzheimer’s disease [25]. The usefulness of pomegranates has also been reported by a clinical study for memory recovery after ischemic stroke [26]. Nevertheless, several issues central to a basic understanding of the modulation of cognitive performance and the possible prevention of memory decline by this natural herbal component remain unresolved.
The mentioned antioxidant characteristic of PG and the fact that sleep deprivation accelerates mediators of inflammation inspired us to investigate the possible neuroprotective effect of PG in a TSD memory impairment model in rats.
Male Wistar rats (weighing 200-220 g, age 9-11 weeks) were purchased from
Institute for Cognitive Science Studies and kept in standard conditions
(Temperature: 23
Punicalagin was purchased from Sigma-Aldrich (
Fig. 1 illustrates the general design of the study. The explanation for each step has been provided in the respective following sections:
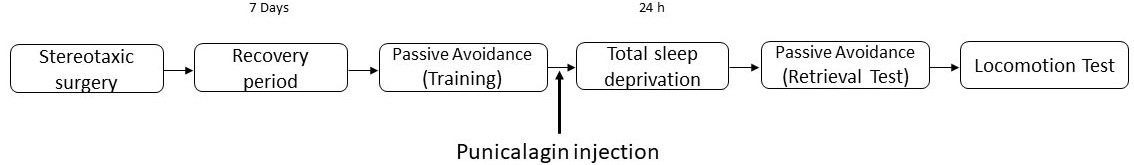 Fig. 1.
Fig. 1.The experimental design. The animals that received surgery were given one week recovery period and then passive avoidance training was conducted. Vehicle or punicalagin (PG) injection was then performed and the animals were placed in the total sleep deprivation apparatus. The apparatus was either was kept ‘off’ or turned ‘on’ for the sham and experimental groups, respectively. All animals underwent a passive avoidance test followed by a locomotion test.
Each rat was anesthetized using 2 mL/kg intraperitoneal injection of ketamine hydrochloride 10% (50 mg/kg) and xylazine 2% (4 mg/kg). The animal’s head was then fixed in a Kopf stereotaxic frame. A stainless steel guide cannula (22 gauge) was implanted above the left lateral ventricle (AP = -0.9, ML = 1.3, DV = -4.5 from skull surface) according to rat brain atlas by Paxinos and Watson [27] and fixed to the skull with instant glue and acrylic resin. After surgery, an injection of meloxicam (1 mg/kg; s.c.), as a painkiller, was made and followed by an injection of penicillin-G 200,000 IU/mL (0.2-0.3 mL/rat, single dose, intramuscular). A stainless-steel stylet was inserted into the guide cannula to prevent its occlusion during the recovery period (one week).
An automatic TSD apparatus (BorjSanatazma Co, Tehran, Iran) was used [28]. The
apparatus has four equal-sized boxes (30
To inject PG, we used a 2
Passive avoidance emotional memory test apparatus included a Plexiglas box
composed of two equal-sized partitions (20
2.3.4.1 Habituation, training, and retrieval test
To habituate the animals to the passive avoidance apparatus, the animals were allowed to habituate in the experiment room for at least 30 min prior to the experiments. Each animal in each experimental group was gently placed in the white partition of the apparatus and then, the guillotine door was raised 5 sec later. As rats are inclined to move to the dark partition, the entrance latency to the dark partition (step-through latency; STL) was recorded when the rat placed all four paws in the dark partition. The gate was closed after 10 sec. and the animal was returned from the dark partition into the home cage. Animals with a STL more than 100 sec were excluded from the experiments (2 rats).
The training trial was performed 30 min after the habituation. The animal was placed in the light compartment and 5 sec later, the guillotine door was opened. As soon as the animal crossed to the dark compartment, the door was closed and a foot shock was immediately delivered to the grid-floor of the dark compartment. After 20 sec, the animal was removed from the apparatus and temporarily placed in its home-cage. The procedure was repeated in 2 min time. The training was terminated when the rat remained in the light partition consecutively for 120 sec. The number of trials (entries into the dark partition) was also recorded. All animals learned within a maximum of three trials.
For the retrieval test of the long-term memory, each animal was placed in the light partition 24 hours after the training for 20 sec, then the door was raised and the STL to enter the dark partition was measured. The test session was ended once the animal entered the dark partition. A cut-off time of 300 sec was applied for those animals which remained in the light partition. No electric shock was applied during the retrieval session [28, 29].
Following the passive avoidance test, the rats were tested using a locomotor
activity test apparatus (BorjSanatazma Co, Tehran, Iran). The animals were
acclimatized to the test room (15 min) and then put into a clear Plexiglas
container box as the test chamber (30
To verify the cannula placement, each rat was deeply anesthetized with Ketamine
and Xylazine, the brain was removed and kept in formaldehyde (10%) for 5 days,
the brain was sliced (50
Seven groups of animals were used in this study (Table 1). The animals that received surgery were given one week recovery period and then the passive avoidance training was conducted. Vehicle or PG injection was then performed and the animals were put in the TSD apparatus. The apparatus was either kept off or turned on for the sham and experimental groups, respectively. Then, all the animals underwent the passive avoidance test followed by the locomotion test.
| Group | Surgery | Passive avoidance training | Punicalagin (i.c.v. injection) | TSD apparatus | Passive avoidance test | Locomotion test |
| Normal control | - | + | - | - | + | + |
| TSD sham control | + | + | - | off | + | + |
| TSD control | + | + | - | on | + | + |
| TSD sham vehicle | + | + | vehicle | off | + | + |
| TSD vehicle | + | + | vehicle | on | + | + |
| TSD sham + PG | + | + | + | off | + | + |
| TSD + PG | + | + | + | on | + | + |
| i.c.v., Intracerebroventricular; PG, Punicalagin; TSD, Total sleep deprivation. | ||||||
Data were analyzed by GraphPad Prism® (Version 9.0).
Kolmogorov-Smirnov statistical test did not show the normality of STL data for
all the experimental groups. Thus, a Kruskal-Wallis test followed by Dunn’s
multiple comparisons was used to identify possible differences. STL data are
expressed as median (interquartile) values. The data regarding distance traveled
followed a normal distribution in all groups and were analyzed using one-way
ANOVA test followed by Tukey-Kramer post-hoc test, as appropriated.
Distance traveled data are expressed as mean
To test the possible effects of TSD on the retrieval of long-term memory,
Kruskal-Wallis test was used on control groups data (normal control, TSD sham
control, and TSD control). The test revealed that there was a statistically
significant difference between all the three groups being tested [Chi-Square (2)
= 11.54; P
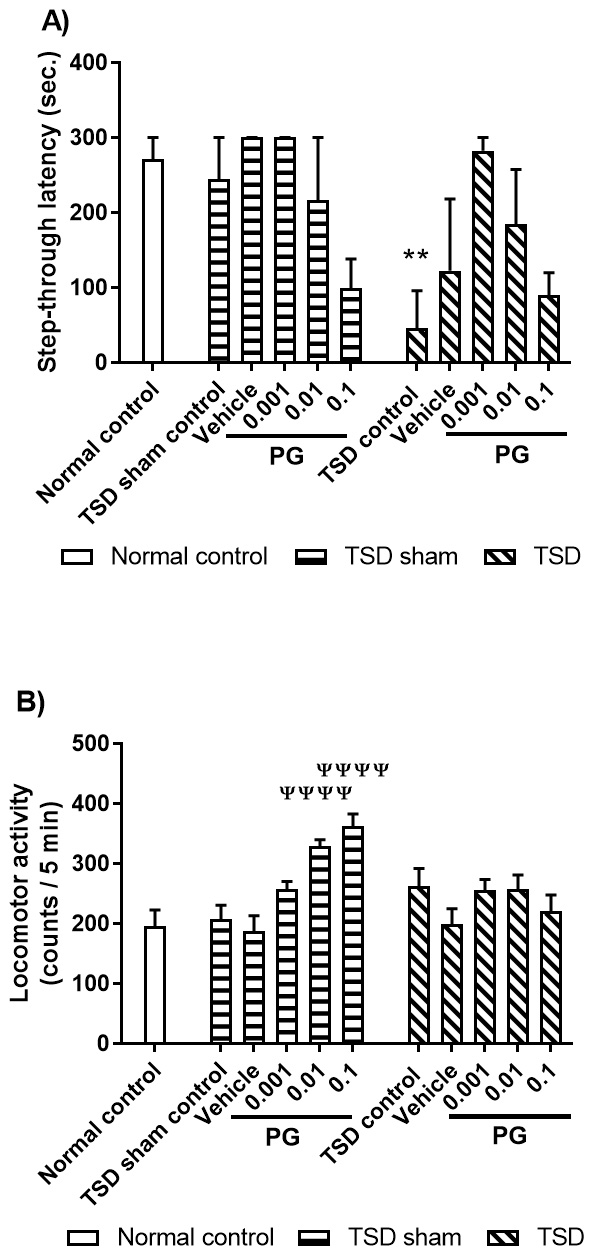 Fig. 2.
Fig. 2.The effects of post-training administration of
punicalagin (PG) on: A) memory consolidation; B) locomotor activity. In this
study, eleven groups of animals were used - from left to right: (i) The animals
in the normal control group did not receive surgery and were tested in passive
avoidance test and locomotion test; All animals in the TSD sham groups went
through the procedures while the apparatus was kept off; (ii) The animals in the
TSD sham group received surgery and spent 24 hours in the TSD apparatus (the
apparatus was kept off) before being tested in passive avoidance test and
locomotion test; (iii to vi) The animals received vehicle or different doses of
PG (0.001, 0.01, or 0.1
STL data (A) are expressed as median (interquartile) values for eight animals
per group (Kruskal-Wallis test followed by Dunn’s multiple comparisons). Distance
travelled data (B) are expressed as mean
** P
To analyze locomotor activity, univariant one-way ANOVA test was performed. The test revealed no statistically significant difference between all the three groups being tested [F (2, 21) = 1.820; P = 0.1867; Fig. 2B].
Our data revealed that 24 hours of TSD impaired retrieval of long-term memory without affecting the locomotor activity.
Comparison of the TSD sham groups using Kruskal-Wallis test revealed that there were no statistically significant differences between the groups being tested as compared to the vehicle group [Chi-Square (3) = 6.001; P = 0.1852; Fig. 2A-TSD sham groups]. Comparison of the TSD groups using Kruskal-Wallis test also revealed no statistically significant differences between the groups being tested, as compared to the vehicle group [Chi-Square (3) = 7.68; P = 0.0471; Fig. 2A-TSD groups].
Comparison of the TSD sham groups using one-way ANOVA on the results obtained
from locomotion activity tests indicated a statistically significant difference
between the experimental groups being tested [F (4, 35) = 15.20, P
Our data revealed that none of PG doses altered memory consolidation by themselves (TSD sham groups). Also, PG microinjection before TSD did not alter memory consolidation significantly and only a tendency of memory restoration was observed. While PG changed the locomotive activity in the TSD sham group, no changes was observed in the TSD group.
Lifestyle changes in modern societies have made sleep deprivation a major challenge that can increase the risk of some neurologic diseases [31]. Sleep plays a prominent role in the maintenance of mental and physical performance and inadequate sleep has deteriorating effects on cognitive functions [32] including memory functions [33].
In line with previous studies [28, 34], we here showed that acute TSD impaired memory processes in the passive avoidance task. The data also showed that PG, per se, did not have any significant effects on the memory functions in this test, but it increased the locomotor activity at the two highest applied doses. PG microinjection before TSD only showed a tendency of restoring the memory impairment and did not alter the locomotor activity. Hence, the observed tendency was not under the influence of changes in locomotor activity. Although some reports indicated no changes in locomotor activity following TSD [35, 36], other reports have indicated an increase in locomotion following sleep deprivation [37] and a recent research on the effects of pomegranate on parkinsonism proposed changes in the level of brain-derived neurotrophic factor (BDNF) as a possible mechanism for the observed increase in the locomotor activity [38]. Applied methods for sleep deprivation and the duration of sleep deprivation might be the reason for the observed differences. Sleep deprivation leads to brain inflammation [10, 39], and reports indicate the protective effect of PG in brain inflammation [40, 41].
Our results showed that 24 hours of TSD impaired retrieval of memory in the passive avoidance test. Signaling pathways underlying the deficit in LTP have been investigated by some researchers. For example, the expression of BDNF and its downstream targets (Synapsin I, cAMP response-element-binding, CREB, and calcium–calmodulin-dependent protein kinase II, CAMKII) reduced in the hippocampus following 8 and 48 h of sleep deprivation [12]. In addition, the role of a compensatory increase in cAMP signaling [42] and reduction of extracellular adenosine [43] in preventing LTP deficit following sleep deprivation have been reported. Maintaining CREB at its beneficial level might be a possible mechanism for the observed tendency, as a study showed that hydro-alcoholic leaf extract of Terminalia cattapa, which also contains PG, regulates BDNF and CREB levels [44].
Sleep deprivation promotes the production of pro-inflammatory proteins both in
humans and rodents [45, 46]. The anti-inflammatory effects of PG have been
reported in rats [41] and its neuroprotective effect on glutamate-induced
oxidative stress has been reported in the mouse hippocampal cell line, HT22 [47].
It has also been reported that PG can inhibit lipopolysaccharide-induced
inflammation in certain types of macrophages [40] and anti-inflammatory property
of PG works against memory deficit via the prevention of neuroinflammation [48].
Furthermore, it has been reported that PG inhibited the expression of NF-
It has been shown that sleep stages and its pattern vary in different animals, which is a point for consideration while the results are interpreted [49]. For example, other studies are reporting that sleep deprivation, for 96 hours, did not induce necrotic or apoptotic cell loss in rat’s brain [50]. Another study that measured antioxidant and oxidant markers in the hippocampus reported short- and long-term memory impairment after 72 hours of sleep deprivation, but found no significant differences in the markers compared to the control group [51]. Also, another study reported no significant rise in oxidative stress in 24-hour sleep-deprived rats [52].
PG only showed a tendency of restoring the memory impairment following 24 hours of TSD in the passive avoidance test. Considering the reported beneficial actions of PG by other studies, further investigation is needed into the possible effects of PG on memory alterations.
ANOVA, Analysis of Variance; BDNF, Brain-derived neurotrophic factor; CAMKII, Calcium-calmodulin-dependent protein kinase II; CREB, cAMP response-element-binding; LTP, Long-term potentiation; NREM, Non-rapid eye movement; PG, Punicalagin; REM, Rapid eye movement; RSD, Rapid eye movement sleep deprivation; TSD, Total sleep deprivation.
Shahram Zarrabian and Mohammad Nasehi were responsible for the study concept and design. Mohammad-Hossein Mohammadi-Mahdiabadi-Hasani contributed to the acquisition of behavioral data. Shahram Zarrabian, Mohammad Nasehi, and Mohammad-Reza Zarrindast assisted with data analysis and interpretation of findings. Shahram Zarrabian drafted and finalized the manuscript. Shahram Zarrabian and Mohammad Nasehi provided critical revision of the manuscript for important intellectual content. All authors critically reviewed content and approved the final version for publication.
All the experiments were conducted under the Guide for the Care and Use of Laboratory Animals (National Institutes of Health Publication No. 80-23, revised 1996). The experiments were approved by the Research and Ethics Committee of Tehran Medical Sciences, Islamic Azad University, Tehran, Iran.
We would like to thank the three anonymous reviewers for their excellent criticism of the article.
This project was supported by grant No. 1369704250005 received from Tehran Medical Sciences, Islamic Azad University, Tehran, Iran.
The authors report no conflict of interest. Given his role as the Review Board Member of JIN, Dr. Shahram Zarrabian had no involvement in the peer-review of this article and has no access to information regarding its peer-review.
Supplementary material associated with this article can be found, in the online version, at https://jin.imrpress.com/EN/10.31083/j.jin.2021.01.378.